
Analysis of Huntington’s Disease Modifiers Using the Hyperbolic Mapping of the Protein Interaction Network




Abstract
:1. Introduction
2. Results
2.1. Human Protein Interaction Network Embedding to Hyperbolic Disc (hPIN)
2.2. Identification of Protein Clusters in the Angular Dimension
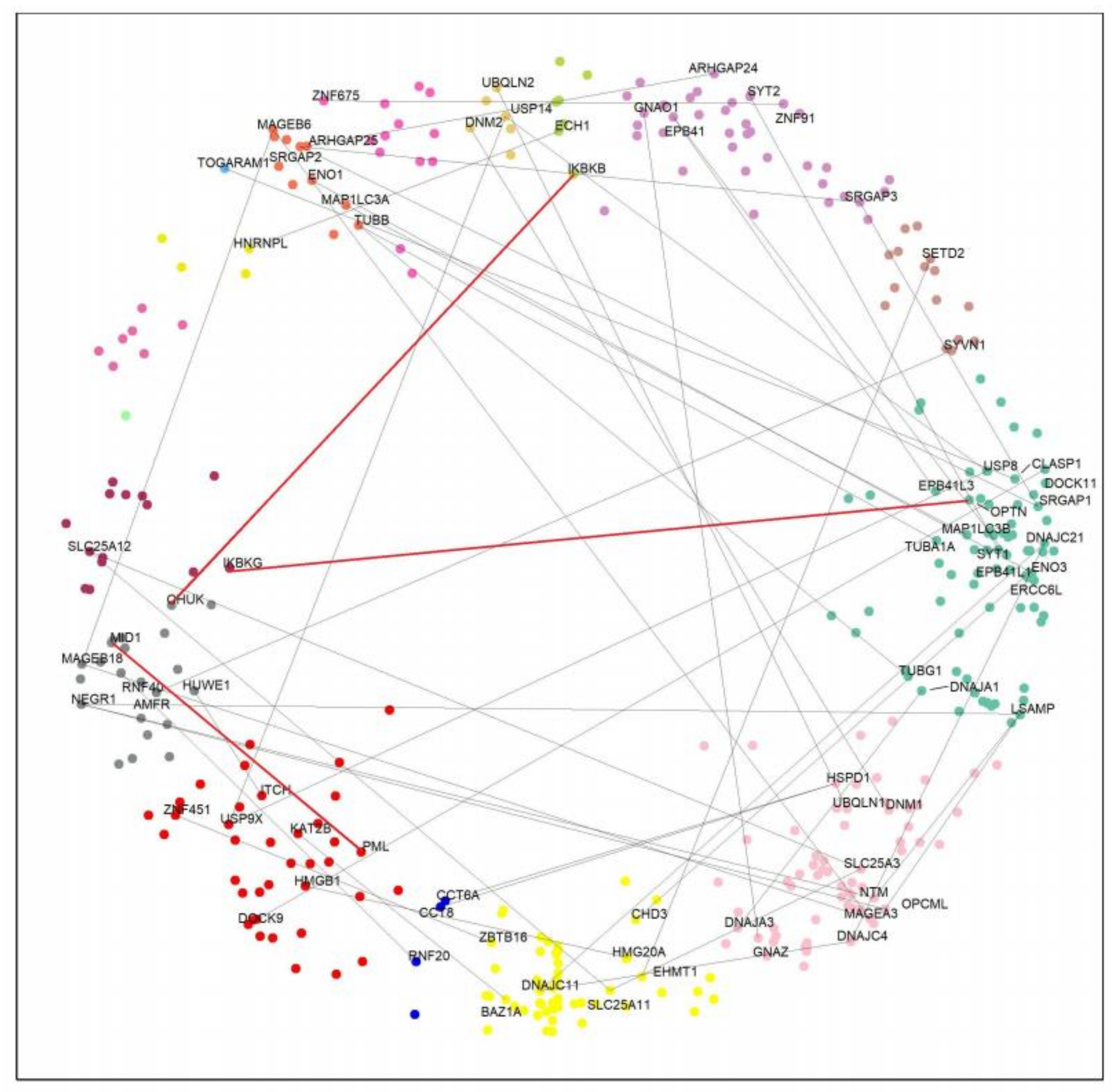
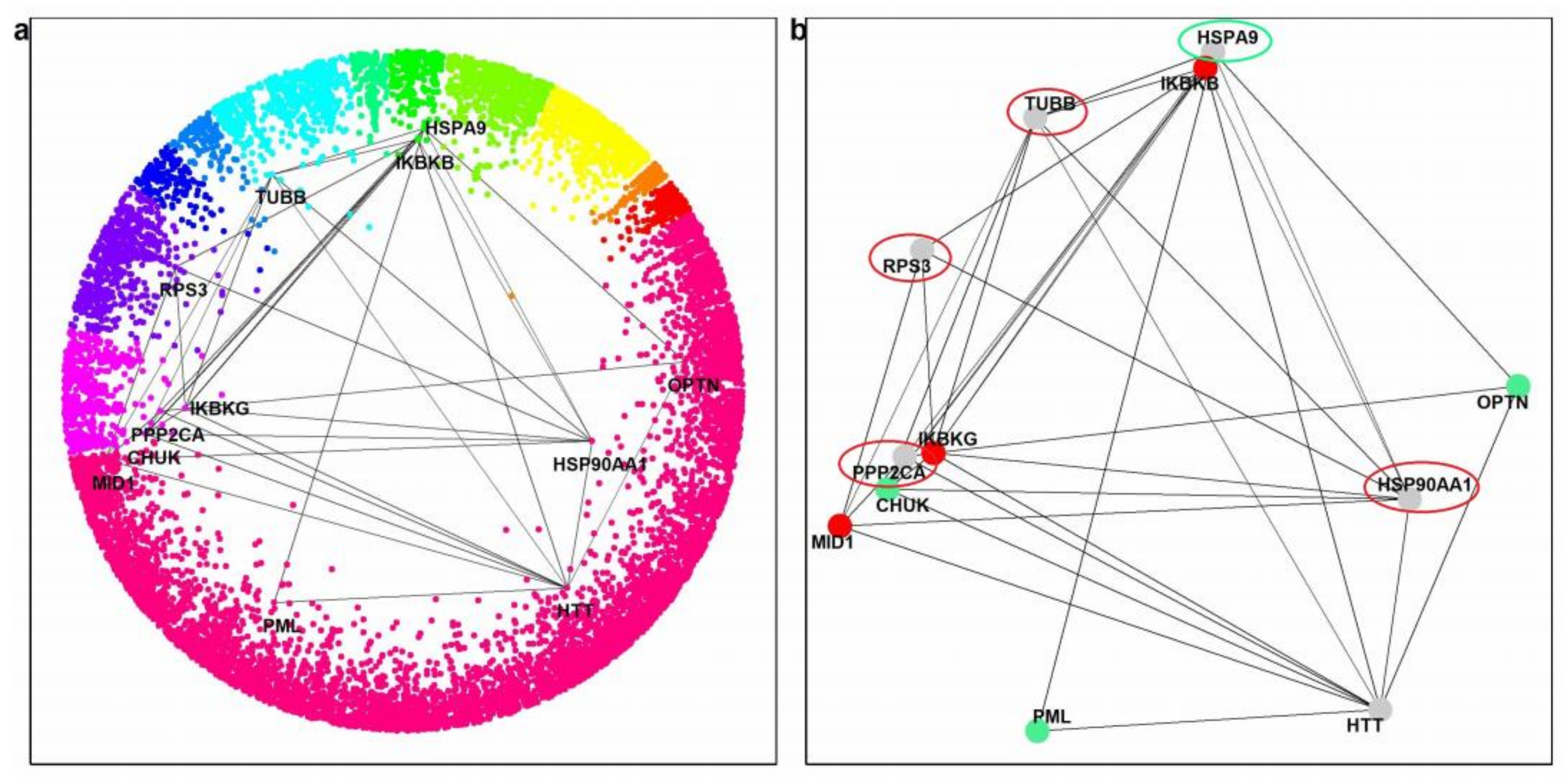
2.3. HTT-Interactors in the Hyperbolic Disc
2.4. Paralog Pairs of HTT Interactors
2.5. Effects of Paralog Pairs on HD
2.6. Common Interactors between Positive and Negative Paralogs
3. Discussion
4. Materials and Methods
4.1. Construction of the hPIN
4.2. Mapping the hPIN to Hyperbolic Space
4.3. Clustering in the Similarity Dimension
4.4. HTT Interactors in the Hyperbolic Space
4.5. Paralog Pairs and Common Interacting Partners
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Macdonald, M. A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington’s disease chromosomes. Cell 1993, 72, 971–983. [Google Scholar] [CrossRef]
- Ordway, J.M.; Tallaksen-Greene, S.; Gutekunst, C.-A.; Bernstein, E.M.; Cearley, J.A.; Wiener, H.W.; Dure, L.S.; Lindsey, R.; Hersch, S.M.; Jope, R.S.; et al. Ectopically Expressed CAG Repeats Cause Intranuclear Inclusions and a Progressive Late Onset Neurological Phenotype in the Mouse. Cell 1997, 91, 753–763. [Google Scholar] [CrossRef] [Green Version]
- Kaminosono, S.; Saito, T.; Oyama, F.; Ohshima, T.; Asada, A.; Nagai, Y.; Nukina, N.; Hisanaga, S.I. Suppression of Mutant Huntingtin Aggregate Formation by Cdk5/p35 through the Effect on Microtubule Stability. J. Neurosci. 2008, 28, 8747–8755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harjes, P.; Wanker, E.E. The hunt for huntingtin function: Interaction partners tell many different stories. Trends Biochem. Sci. 2003, 28, 425–433. [Google Scholar] [CrossRef]
- Kaltenbach, L.S.; Romero, E.; Becklin, R.R.; Chettier, R.; Bell, R.; Phansalkar, A.; Strand, A.; Torcassi, C.; Savage, J.; Hurlburt, A.; et al. Huntingtin Interacting Proteins Are Genetic Modifiers of Neurodegeneration. PLoS Genet. 2007, 3, e82. [Google Scholar] [CrossRef] [Green Version]
- Li, S.-H.; Li, X.-J. Huntingtin–protein interactions and the pathogenesis of Huntington’s disease. Trends Genet. 2004, 20, 146–154. [Google Scholar] [CrossRef]
- DiFiglia, M.; Sapp, E.; Chase, K.O.; Davies, S.W.; Bates, G.P.; Vonsattel, J.P.; Aronin, N. Aggregation of Huntingtin in Neuronal Intranuclear Inclusions and Dystrophic Neurites in Brain. Science 1997, 277, 1990–1993. [Google Scholar] [CrossRef]
- Hoffner, G.; Island, M.L.; Djian, P. Purification of neuronal inclusions of patients with Huntington’s disease reveals a broad range of N-terminal fragments of expanded huntingtin and insoluble polymers: Huntingtin fragments and polymers in inclusions. J. Neurochem. 2005, 95, 125–136. [Google Scholar] [CrossRef]
- Boutell, J.M.; Thomas, P.; Neal, J.W.; Weston, V.J.; Duce, J.; Harper, P.S.; Jones, A.L. Aberrant Interactions of Transcriptional Repressor Proteins with the Huntington’s Disease Gene Product, Huntingtin. Hum. Mol. Genet. 1999, 8, 1647–1655. [Google Scholar] [CrossRef] [Green Version]
- Nucifora, F.C.; Sasaki, M.; Peters, M.F.; Huang, H.; Cooper, J.K.; Yamada, M.; Takahashi, H.; Tsuji, S.; Troncoso, J.; Dawson, V.L.; et al. Interference by Huntingtin and Atrophin-1 with CBP-Mediated Transcription Leading to Cellular Toxicity. Science 2001, 291, 2423–2428. [Google Scholar] [CrossRef] [Green Version]
- Suhr, S.T.; Senut, M.-C.; Whitelegge, J.P.; Faull, K.F.; Cuizon, D.B.; Gage, F.H. Identities of Sequestered Proteins in Aggregates from Cells with Induced Polyglutamine Expression. J. Cell Biol. 2001, 153, 283–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerson, J.E.; Safren, N.; Fischer, S.; Patel, R.; Crowley, E.V.; Welday, J.P.; Windle, A.K.; Barmada, S.; Paulson, H.L.; Sharkey, L.M. Ubiquilin-2 differentially regulates polyglutamine disease proteins. Hum. Mol. Genetics 2020, 29, 2596–2610. [Google Scholar] [CrossRef] [PubMed]
- Heinz, A.; Schilling, J.; van Roon-Mom, W.; Krauß, S. The MID1 Protein: A Promising Therapeutic Target in Huntington’s Disease. Front. Genet. 2021, 12, 761714. [Google Scholar] [CrossRef] [PubMed]
- Hyrskyluoto, A.; Bruelle, C.; Lundh, S.H.; Do, H.T.; Kivinen, J.; Rappou, E.; Reijonen, S.; Waltimo, T.; Petersén, Å.; Lindholm, D.; et al. Ubiquitin-specific protease-14 reduces cellular aggregates and protects against mutant huntingtin-induced cell degeneration: Involvement of the proteasome and ER stress-activated kinase IRE1. Hum. Mol. Genet. 2014, 23, 5928–5939. [Google Scholar] [CrossRef] [Green Version]
- Min, H.J.; Ko, E.A.; Wu, J.; Kim, E.S.; Kwon, M.K.; Kwak, M.S.; Choi, J.E.; Lee, J.E.; Shin, J.-S. Chaperone-like Activity of High-Mobility Group Box 1 Protein and Its Role in Reducing the Formation of Polyglutamine Aggregates. J. Immunol. 2013, 190, 1797–1806. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.; Bacos, K.; Fedele, V.; Soulet, D.; Walz, H.A.; Obermüller, S.; Lindqvist, A.; Björkqvist, M.; Klein, P.; Önnerfjord, P.; et al. Mutant huntingtin interacts with -tubulin and disrupts vesicular transport and insulin secretion. Hum. Mol. Genet. 2009, 18, 3942–3954. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.Q.; Xu, Y.X.; Zhao, X.Y.; Zhao, H.; Yan, J.; Sun, X.B.; Guo, J.-C.; Zhu, C.-Q. Overexpression of F0F1-ATP synthase α suppresses mutant huntingtin aggregation and toxicity in vitro. Biochem. Biophys. Res. Commun. 2009, 390, 1294–1298. [Google Scholar] [CrossRef]
- Yang, H.; Liu, C.; Zhong, Y.; Luo, S.; Monteiro, M.J.; Fang, S. Huntingtin Interacts with the Cue Domain of gp78 and Inhibits gp78 Binding to Ubiquitin and p97/VCP. PLoS ONE 2010, 5, e8905. [Google Scholar] [CrossRef]
- Cariulo, C.; Azzollini, L.; Verani, M.; Martufi, P.; Boggio, R.; Chiki, A.; Deguire, S.M.; Cherubini, M.; Gines, S.; Marsh, J.L.; et al. Phosphorylation of huntingtin at residue T3 is decreased in Huntington’s disease and modulates mutant huntingtin protein conformation. Proc. Natl. Acad. Sci. USA 2017, 114, E10809–E10818. [Google Scholar] [CrossRef] [Green Version]
- Mishra, R.; Hoop, C.L.; Kodali, R.; Sahoo, B.; van der Wel, P.C.A.; Wetzel, R. Serine Phosphorylation Suppresses Huntingtin Amyloid Accumulation by Altering Protein Aggregation Properties. J. Mol. Biol. 2012, 424, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Petrakis, S.; Rasko, T.; Russ, J.; Friedrich, R.P.; Stroedicke, M.; Riechers, S.-P.; Muehlenberg, K.; Möller, A.; Reinhardt, A.; Vinayagam, A.; et al. Identification of Human Proteins That Modify Misfolding and Proteotoxicity of Pathogenic Ataxin-1. PLoS Genet. 2012, 8, e1002897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrakis, S.; Schaefer, M.H.; Wanker, E.E.; Andrade-Navarro, M.A. Aggregation of polyQ-extended proteins is promoted by interaction with their natural coiled-coil partners. BioEssays 2013, 35, 503–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kastano, K.; Mier, P.; Andrade-Navarro, M.A. The Role of Low Complexity Regions in Protein Interaction Modes: An Illustration in Huntingtin. Int. J. Mol. Sci. 2021, 22, 1727. [Google Scholar] [CrossRef] [PubMed]
- Jensen, L.J.; Ussery, D.W.; Brunak, S. Functionality of System Components: Conservation of Protein Function in Protein Feature Space. Genome Res. 2003, 13, 2444–2449. [Google Scholar] [CrossRef]
- Zallot, R.; Harrison, K.J.; Kolaczkowski, B.; De Crécy-Lagard, V. Functional Annotations of Paralogs: A Blessing and a Curse. Life 2016, 6, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serlidaki, D.; van Waarde, M.A.W.H.; Rohland, L.; Wentink, A.S.; Dekker, S.L.; Kamphuis, M.J.; Boertien, J.M.; Brunsting, J.F.; Nillegoda, N.B.; Bukau, B.; et al. Functional diversity between HSP70 paralogs caused by variable interactions with specific co-chaperones. J. Biol. Chem. 2020, 295, 7301–7316. [Google Scholar] [CrossRef] [Green Version]
- Favaro, P.D.; Huang, X.; Hosang, L.; Stodieck, S.; Cui, L.; Liu, Y.-z.; Engelhardt, K.-A.; Schmitz, F.; Dong, Y.; Löwel, S.; et al. An opposing function of paralogs in balancing developmental synapse maturation. PLoS Biol. 2018, 16, e2006838. [Google Scholar] [CrossRef]
- Gosak, M.; Markovič, R.; Dolenšek, J.; Slak Rupnik, M.; Marhl, M.; Stožer, A.; Perc, M. Network science of biological systems at different scales: A review. Phys. Life Rev. 2018, 24, 118–135. [Google Scholar] [CrossRef]
- Alanis-Lobato, G.; Mier, P.; Andrade-Navarro, M. The latent geometry of the human protein interaction network. Bioinformatics 2018, 34, 2826–2834. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulos, F.; Kitsak, M.; Serrano, M.Á.; Boguñá, M.; Krioukov, D. Popularity versus similarity in growing networks. Nature 2012, 489, 537–540. [Google Scholar] [CrossRef] [Green Version]
- Härtner, F.; Andrade-Navarro, M.A.; Alanis-Lobato, G. Geometric characterisation of disease modules. Appl. Netw. Sci. 2018, 3, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alanis-Lobato, G.; Andrade-Navarro, M.A.; Schaefer, M.H. HIPPIE v2.0: Enhancing meaningfulness and reliability of protein–protein interaction networks. Nucleic Acids Res. 2017, 45, D408–D414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaefer, M.H.; Fontaine, J.F.; Vinayagam, A.; Porras, P.; Wanker, E.E.; Andrade-Navarro, M.A. HIPPIE: Integrating Protein Interaction Networks with Experiment Based Quality Scores. PLoS ONE 2012, 7, e31826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alanis-Lobato, G.; Mier, P.; Andrade-Navarro, M.A. Efficient embedding of complex networks to hyperbolic space via their Laplacian. Sci. Rep. 2016, 6, 30108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alanis-Lobato, G.; Mier, P.; Andrade-Navarro, M.A. Manifold learning and maximum likelihood estimation for hyperbolic network embedding. Appl. Netw. Sci. 2016, 1, 10. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulos, F.; Aldecoa, R.; Krioukov, D. Network geometry inference using common neighbors. Phys. Rev. E 2015, 92, 022807. [Google Scholar] [CrossRef] [Green Version]
- Musso, G.; Zhang, Z.; Emili, A. Retention of protein complex membership by ancient duplicated gene products in budding yeast. Trends Genet. 2007, 23, 266–269. [Google Scholar] [CrossRef]
- Wagner, A. The Yeast Protein Interaction Network Evolves Rapidly and Contains Few Redundant Duplicate Genes. Mol. Biol. Evol. 2001, 18, 1283–1292. [Google Scholar] [CrossRef] [Green Version]
- Wacker, J.L.; Zareie, M.H.; Fong, H.; Sarikaya, M.; Muchowski, P.J. Hsp70 and Hsp40 attenuate formation of spherical and annular polyglutamine oligomers by partitioning monomer. Nat. Struct. Mol. Biol. 2004, 11, 1215–1222. [Google Scholar] [CrossRef]
- Chai, Y.; Koppenhafer, S.L.; Bonini, N.M.; Paulson, H.L. Analysis of the Role of Heat Shock Protein (Hsp) Molecular Chaperones in Polyglutamine Disease. J. Neurosci. 1999, 19, 10338–10347. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-González, C.; Lin, S.; Arkan, S.; Hansen, C. Co-chaperones DNAJA1 and DNAJB6 are critical for regulation of polyglutamine aggregation. Sci. Rep. 2020, 10, 8130. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, A.; Kubota, H.; Pack, C.G.; Matsumoto, G.; Hirayama, S.; Takahashi, Y.; Kimura, H.; Kinjo, M.; Morimoto, R.; Nagata, K. Cytosolic chaperonin prevents polyglutamine toxicity with altering the aggregation state. Nat. Cell Biol. 2006, 8, 1163–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langfelder, P.; Cantle, J.P.; Chatzopoulou, D.; Wang, N.; Gao, F.; Al-Ramahi, I.; Lu, X.-H.; Ramos, E.M.; El-Zein, K.; Zhao, Y.; et al. Integrated genomics and proteomics define huntingtin CAG length–dependent networks in mice. Nat. Neurosci. 2016, 19, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Gärtner, A.; Muller, S. PML, SUMO, and RNF4: Guardians of Nuclear Protein Quality. Mol. Cell 2014, 55, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Giasson, B.I.; Glavis-Bloom, A.; Brewer, M.D.; Shorter, J.; Gitler, A.D.; Yang, X. A Cellular System that Degrades Misfolded Proteins and Protects against Neurodegeneration. Mol. Cell 2014, 55, 15–30. [Google Scholar] [CrossRef] [Green Version]
- Krauß, S.; Griesche, N.; Jastrzebska, E.; Chen, C.; Rutschow, D.; Achmüller, C.; Dorn, S.; Boesch, S.M.; Lalowski, M.; Wanker, E.; et al. Translation of HTT mRNA with expanded CAG repeats is regulated by the MID1–PP2A protein complex. Nat. Commun. 2013, 4, 1511. [Google Scholar] [CrossRef] [Green Version]
- Khoshnan, A.; Ko, J.; Tescu, S.; Brundin, P.; Patterson, P.H. IKKα and IKKβ Regulation of DNA Damage-Induced Cleavage of Huntingtin. PLoS ONE 2009, 4, e5768. [Google Scholar] [CrossRef] [Green Version]
- Anborgh, P.H.; Godin, C.; Pampillo, M.; Dhami, G.K.; Dale, L.B.; Cregan, S.P.; Truant, R.; Ferguson, S.S.G. Inhibition of Metabotropic Glutamate Receptor Signaling by the Huntingtin-binding Protein Optineurin. J. Biol. Chem. 2005, 280, 34840–34848. [Google Scholar] [CrossRef] [Green Version]
- Khoshnan, A.; Ko, J.; Watkin, E.E.; Paige, L.A.; Reinhart, P.H.; Patterson, P.H. Activation of the I B Kinase Complex and Nuclear Factor- B Contributes to Mutant Huntingtin Neurotoxicity. J. Neurosci. 2004, 24, 7999–8008. [Google Scholar] [CrossRef] [Green Version]
- Shen, W.-C.; Li, H.-Y.; Chen, G.-C.; Chern, Y.; Tu, P.-h. Mutations in the ubiquitin-binding domain of OPTN/optineurin interfere with autophagy-mediated degradation of misfolded proteins by a dominant-negative mechanism. Autophagy 2015, 11, 685–700. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Sharma, B. Pharmacological benefit of I1-imidazoline receptors activation and nuclear factor kappa-B (NF-κB) modulation in experimental Huntington’s disease. Brain Res. Bull. 2014, 102, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Kalathur, R.K.R.; Giner-Lamia, J.; Machado, S.; Barata, T.; Ayasolla, K.R.S.; Futschik, M.E. The unfolded protein response and its potential role in Huntington’s disease elucidated by a systems biology approach. F1000Research 2015, 4, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griesche, N.; Schilling, J.; Weber, S.; Rohm, M.; Pesch, V.; Matthes, F.; Auburger, G.; Krauss, S. Regulation of mRNA Translation by MID1: A Common Mechanism of Expanded CAG Repeat RNAs. Front. Cell. Neurosci. 2016, 10, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Häcker, H.; Karin, M. Regulation and Function of IKK and IKK-Related Kinases. Sci. STKE 2006, 2006, re13. [Google Scholar] [CrossRef] [PubMed]
- De Luca, G.; Russo, M.T.; Degan, P.; Tiveron, C.; Zijno, A.; Meccia, E.; Ventura, I.; Mattei, E.; Nakabeppu, Y.; Crescenzi, M.; et al. A Role for Oxidized DNA Precursors in Huntington’s Disease–Like Striatal Neurodegeneration. PLoS Genet. 2008, 4, e1000266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, A.; Roy, A.; Liu, X.; Kordower, J.H.; Mufson, E.J.; Hartley, D.M.; Ghosh, S.; Mosley, R.L.; Gendelman, H.E.; Pahan, K. Selective inhibition of NF-κB activation prevents dopaminergic neuronal loss in a mouse model of Parkinson’s disease. Proc. Natl. Acad. Sci. USA 2007, 104, 18754–18759. [Google Scholar] [CrossRef] [Green Version]
- Mattson, M.P.; Culmsee, C.; Yu, Z.; Camandola, S. Roles of Nuclear Factor κB in Neuronal Survival and Plasticity. J. Neurochem. 2001, 74, 443–456. [Google Scholar] [CrossRef]
- Soylu-Kucharz, R.; Khoshnan, A.; Petersén, Å. IKKβ signaling mediates metabolic changes in the hypothalamus of a Huntington disease mouse model. iScience 2022, 25, 103771. [Google Scholar] [CrossRef]
- Napolitano, M.; Zei, D.; Centonze, D.; Palermo, R.; Bernardi, G.; Vacca, A.; Calabresi, P.; Gulino, A. NF-kB/NOS cross-talk induced by mitochondrial complex II inhibition: Implications for Huntington’s disease. Neurosci. Lett. 2008, 434, 241–246. [Google Scholar] [CrossRef]
- Yakovleva, T.; Bazov, I.; Watanabe, H.; Hauser, K.F.; Bakalkin, G. Transcriptional control of maladaptive and protective responses in alcoholics: A role of the NF-κB system. Brain Behav. Immun. 2011, 25, S29–S38. [Google Scholar] [CrossRef] [Green Version]
- De Marco, N.; Buono, M.; Troise, F.; Diez-Roux, G. Optineurin Increases Cell Survival and Translocates to the Nucleus in a Rab8-dependent Manner upon an Apoptotic Stimulus. J. Biol. Chem. 2006, 281, 16147–16156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vagiona, A.-C.; Andrade-Navarro, M.A.; Psomopoulos, F.; Petrakis, S. Dynamics of a Protein Interaction Network Associated to the Aggregation of polyQ-Expanded Ataxin-1. Genes 2020, 11, 1129. [Google Scholar] [CrossRef] [PubMed]
- Brheme, M.; Voisine, C.; Rolland, T.; Wachi, S.; Soper, J.H.; Zhu, Y.; Orton, K.; Villella, A.; Garza, D.; Vidal, M.; et al. A chaperone subnetwork safeguards proteostasis in aging and neurodegenerative disease. Cell Rep. 2014, 9, 1135–1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metzler, M.; Gan, L.; Mazarei, G.; Graham, R.K.; Liu, L.; Bissada, N.; Lu, G.; Leavitt, B.R.; Hayden, M.R. Phosphorylation of Huntingtin at Ser421 in YAC128 Neurons Is Associated with Protection of YAC128 Neurons from NMDA-Mediated Excitotoxicity and Is Modulated by PP1 and PP2A. J. Neurosci. 2010, 30, 14318–14329. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Luo, W.; Zeng, C.; Zhang, Y.; Wang, L.; Yao, W.; Nie, C. PP2A mediates apoptosis or autophagic cell death in multiple myeloma cell lines. Oncotarget 2017, 8, 80770–80789. [Google Scholar] [CrossRef] [Green Version]
- Haenig, C.; Atias, N.; Taylor, A.K.; Mazza, A.; Schaefer, M.H.; Russ, J.; Riechers, S.-P.; Jain, S.; Coughlin, M.; Fontaine, J.-F.; et al. Interactome Mapping Provides a Network of Neurodegenerative Disease Proteins and Uncovers Widespread Protein Aggregation in Affected Brains. Cell Rep. 2020, 32, 108050. [Google Scholar] [CrossRef]
- Laidou, S.; Alanis-Lobato, G.; Pribyl, J.; Raskó, T.; Tichy, B.; Mikulasek, K.; Tsagiopoulou, M.; Oppelt, J.; Kastrinaki, G.; Lefaki, M.; et al. Nuclear inclusions of pathogenic ataxin-1 induce oxidative stress and perturb the protein synthesis machinery. Redox Biol. 2020, 32, 101458. [Google Scholar] [CrossRef]
- Yadavilli, S.; Hegde, V.; Deutsch, W.A. Translocation of human ribosomal protein S3 to sites of DNA damage is dependant on ERK-mediated phosphorylation following genotoxic stress. DNA Repair 2007, 6, 1453–1462. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Kim, H.D.; Kim, J. Cytoplasmic ribosomal protein S3 (rpS3) plays a pivotal role in mitochondrial DNA damage surveillance. Biochim. Biophys. Acta BBA Mol. Cell Res. 2013, 1833, 2943–2952. [Google Scholar] [CrossRef] [Green Version]
- Bell, R.; Hubbard, A.; Chettier, R.; Chen, D.; Miller, J.P.; Kapahi, P.; Tarnopolsky, M.; Sahasrabuhde, S.; Melov, S.; Hughes, R.E. A Human Protein Interaction Network Shows Conservation of Aging Processes between Human and Invertebrate Species. PLoS Genet. 2009, 5, e1000414. [Google Scholar] [CrossRef] [Green Version]
- Bouwmeester, T.; Bauch, A.; Ruffner, H.; Angrand, P.O.; Bergamini, G.; Croughton, K.; Cruciat, C.; Eberhard, D.; Gagneur, J.; Ghidelli, S.; et al. A physical and functional map of the human TNF-α/NF-κB signal transduction pathway. Nat. Cell Biol. 2004, 6, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto-Sato, E.; Fujimori, S.; Ishizaka, M.; Hirai, N.; Masuoka, K.; Saito, R.; Ozawa, Y.; Hino, K.; Washio, T.; Tomita, M.; et al. A Comprehensive Resource of Interacting Protein Regions for Refining Human Transcription Factor Networks. PLoS ONE 2010, 5, e9289. [Google Scholar] [CrossRef] [PubMed]
- Jo, D.S.; Park, S.J.; Kim, A.K.; Park, N.Y.; Kim, J.B.; Bae, J.E.; Park, J.H.; Shin, J.H.; Chang, J.W.; Kim, P.K.; et al. Loss of HSPA9 induces peroxisomal degradation by increasing pexophagy. Autophagy 2020, 16, 1989–2003. [Google Scholar] [CrossRef] [PubMed]
- Burbulla, L.F.; Schelling, C.; Kato, H.; Rapaport, D.; Woitalla, D.; Schiesling, C.; Schulte, C.; Sharma, M.; Illig, T.; Bauer, P.; et al. Dissecting the role of the mitochondrial chaperone mortalin in Parkinson’s disease: Functional impact of disease-related variants on mitochondrial homeostasis. Hum. Mol. Genet. 2010, 19, 4437–4452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.J.; Shin, J.H.; Jeong, J.I.; Song, J.H.; Jo, Y.K.; Kim, E.S.; Lee, E.H.; Hwang, J.J.; Lee, E.K.; Chung, J.S.; et al. Down-regulation of Mortalin Exacerbates Aβ-mediated Mitochondrial Fragmentation and Dysfunction. J. Biol. Chem. 2014, 289, 2195–2204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krioukov, D.; Papadopoulos, F.; Kitsak, M.; Vahdat, A.; Boguna, M. Hyperbolic geometry of complex networks. Phys. Rev. E 2010, 82, 036106. [Google Scholar] [CrossRef] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, F.; Allen, J.E.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Austine-Orimoloye, O.; Azov, A.G.; Barnes, I.; Bennett, R.; et al. Ensembl 2022. Nucleic Acids Res. 2022, 50, D988–D995. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Proteins | Effects on HD | References |
|---|---|---|---|
| MID1/PML | E3 ubiquitin-protein ligase Midline-1/Protein PML | Mutant HTT recruits MID1 protein complex resulting in overproduction of polyQ HTT; PML plays a protective role against neuronal toxicity associated with polyQ proteins. | [13,44,45,46] |
| IKBKB/IKKA(CHUK) | Inhibitor of nuclear factor kappa B kinase subunit beta/Inhibitor of nuclear factor kappa-B kinase subunit alpha | Inhibition of IKBKB may promote neuronal survival in HD; IKKA has a protective role in preventing HTT proteolysis. | [47] |
| IKBKG/OPTN | NF-kappa-B essential modulator/Optineurin | Inhibition of IKBKG activity reduces HTT-polyQ toxicity; OPTN has a protective effect on polyQ neurotoxicity associated with mutant HTT. | [48,49,50] |
| Paralogs | Effect on HD | Common Interactors between Paralogs | Confidence Score | Interaction of Common Interactors with HTT | Ranking by # of Interactions with HTT-Interactors * |
|---|---|---|---|---|---|
| MID1, IKBKB, IKBKG | Negative | PPP2CA | ≥0.72 | Yes | 402 (20) |
| TUBB | ≥0.63 | Yes | 81 (41) | ||
| HSP90AA1 | ≥0.63 | Yes | 51 (47) | ||
| RPS3 | ≥0.63 | No | 295 (23) | ||
| PML, OPTN, CHUK | Positive | HSPA9 | ≥0.49 | No | 47 (48) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vagiona, A.-C.; Mier, P.; Petrakis, S.; Andrade-Navarro, M.A. Analysis of Huntington’s Disease Modifiers Using the Hyperbolic Mapping of the Protein Interaction Network. Int. J. Mol. Sci. 2022, 23, 5853. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105853
Vagiona A-C, Mier P, Petrakis S, Andrade-Navarro MA. Analysis of Huntington’s Disease Modifiers Using the Hyperbolic Mapping of the Protein Interaction Network. International Journal of Molecular Sciences. 2022; 23(10):5853. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105853
Chicago/Turabian StyleVagiona, Aimilia-Christina, Pablo Mier, Spyros Petrakis, and Miguel A. Andrade-Navarro. 2022. "Analysis of Huntington’s Disease Modifiers Using the Hyperbolic Mapping of the Protein Interaction Network" International Journal of Molecular Sciences 23, no. 10: 5853. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105853