
Regionally Altered Immunosignals of Surfactant Protein-G, Vascular and Non-Vascular Elements of the Neurovascular Unit after Experimental Focal Cerebral Ischemia in Mice, Rats, and Sheep

Abstract
:1. Introduction
2. Results
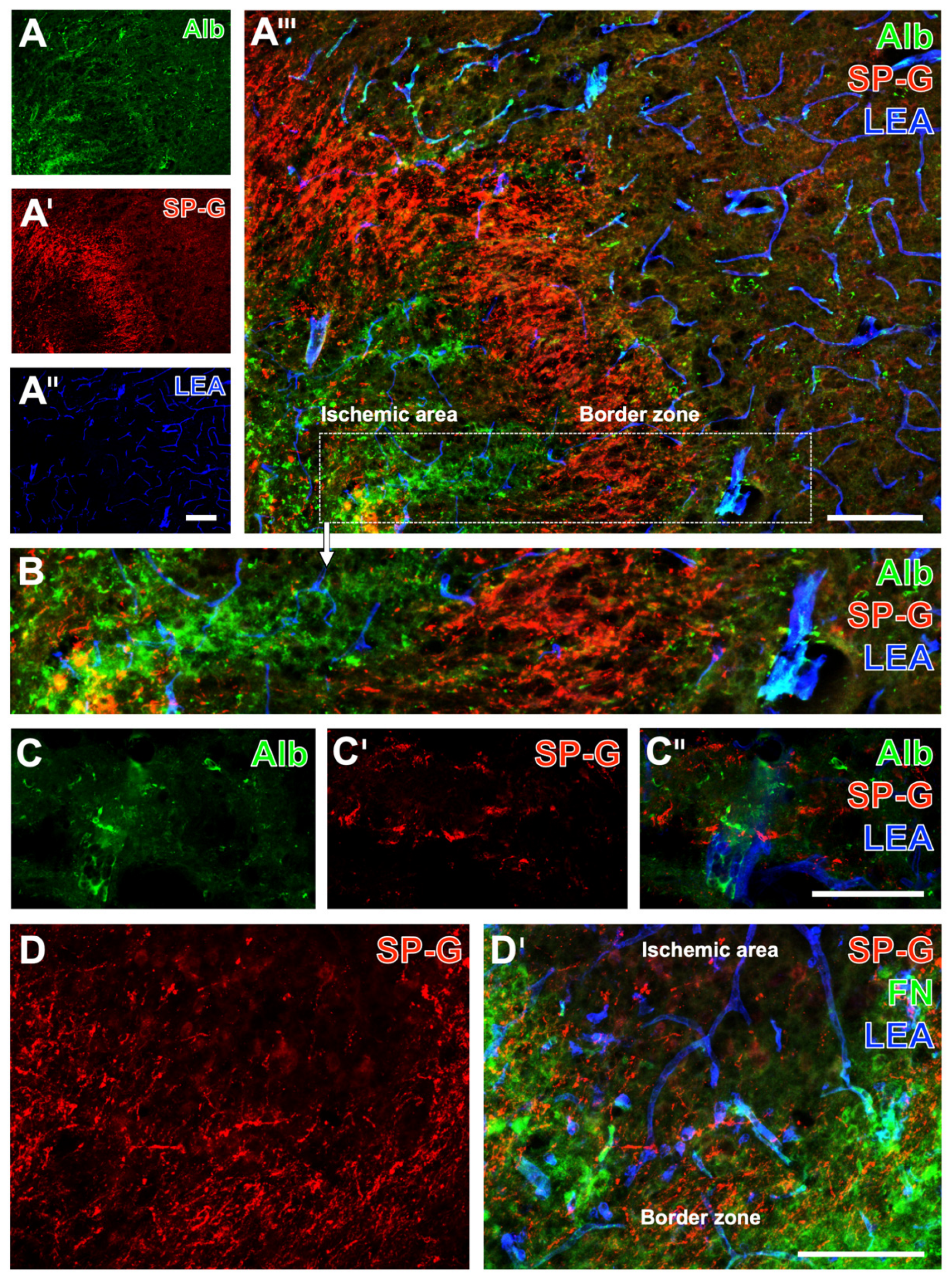
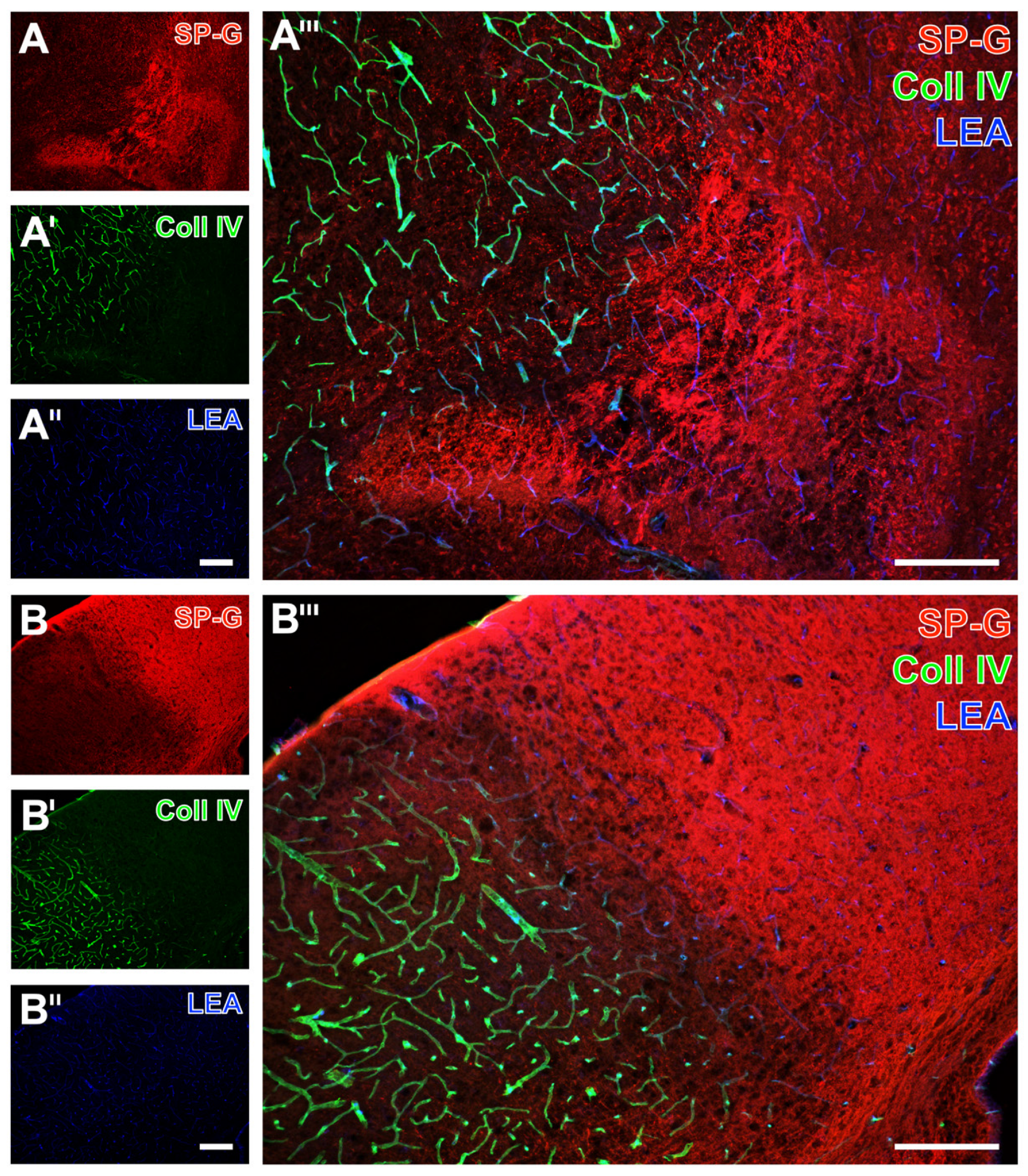
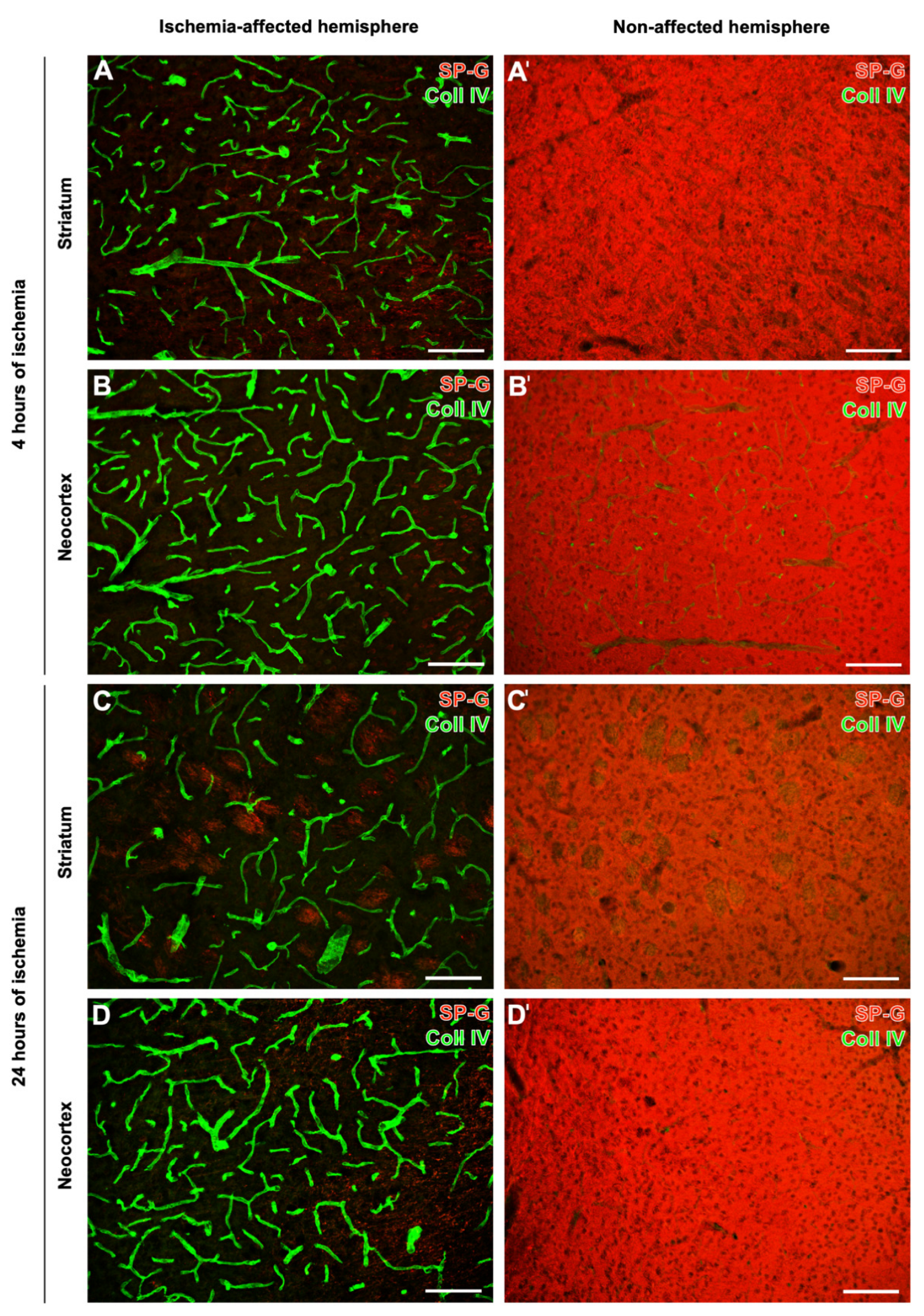
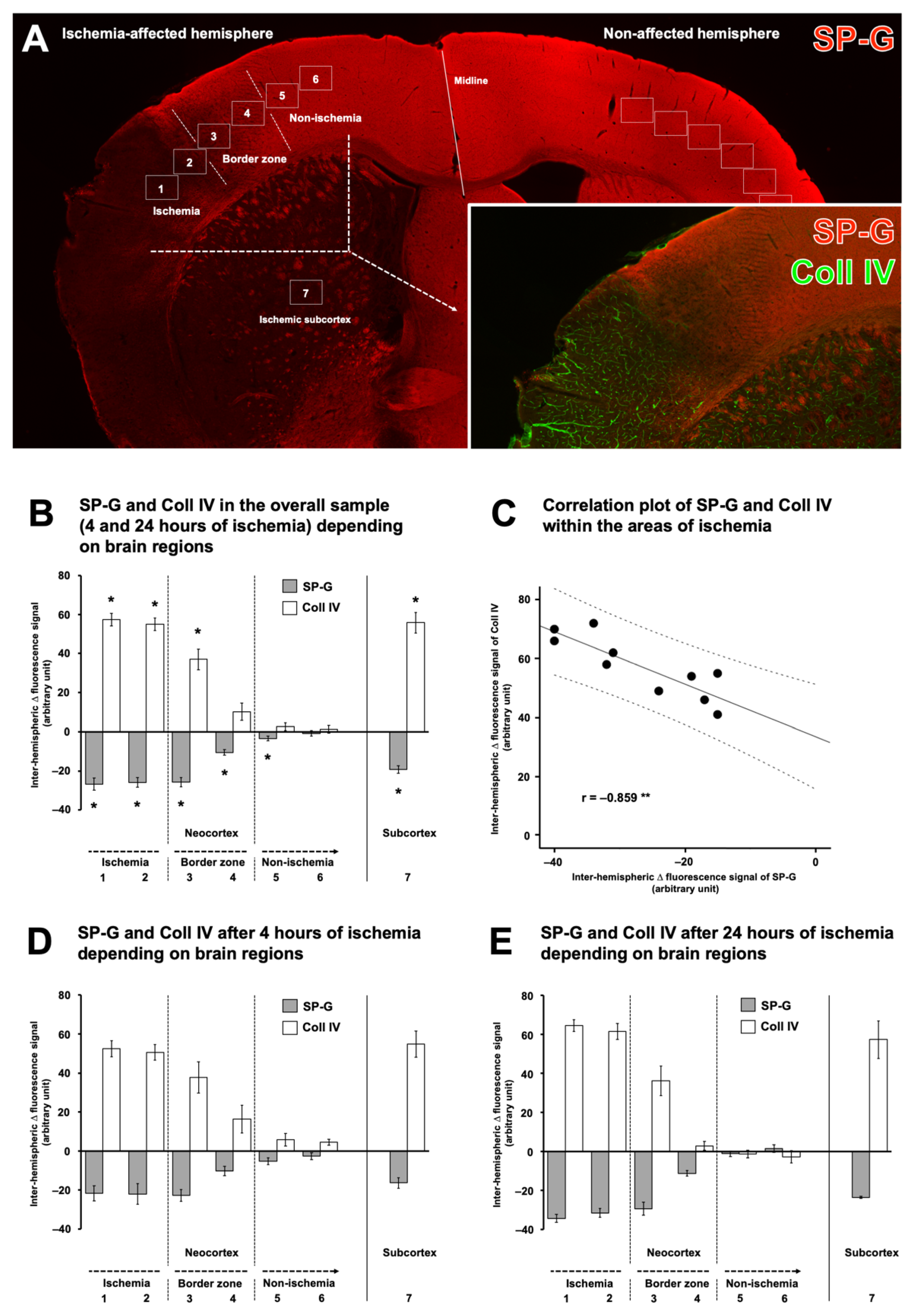
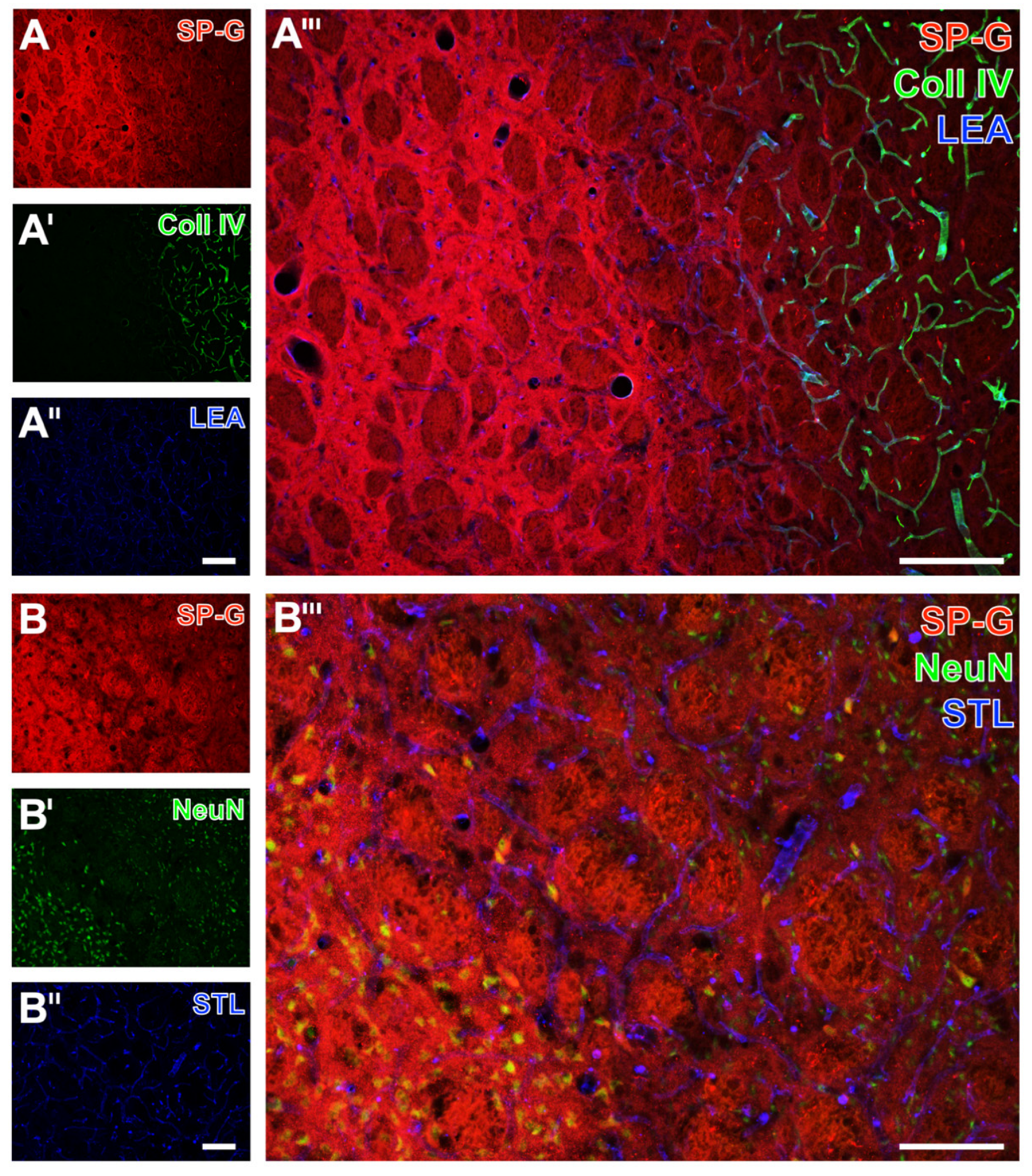
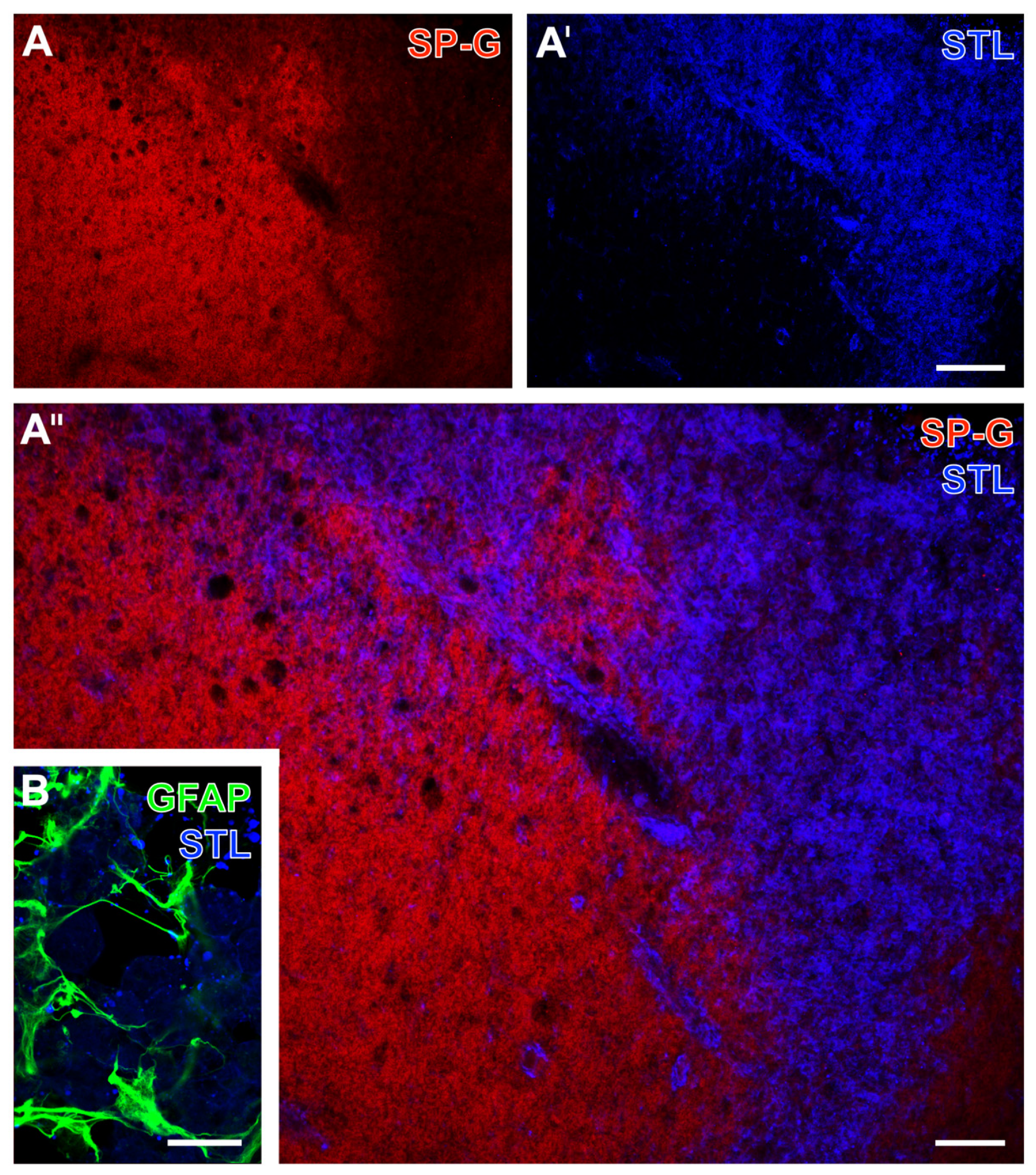
2.1. SP-G and Vascular Elements in Ischemia-Affected Brain Areas in Mice
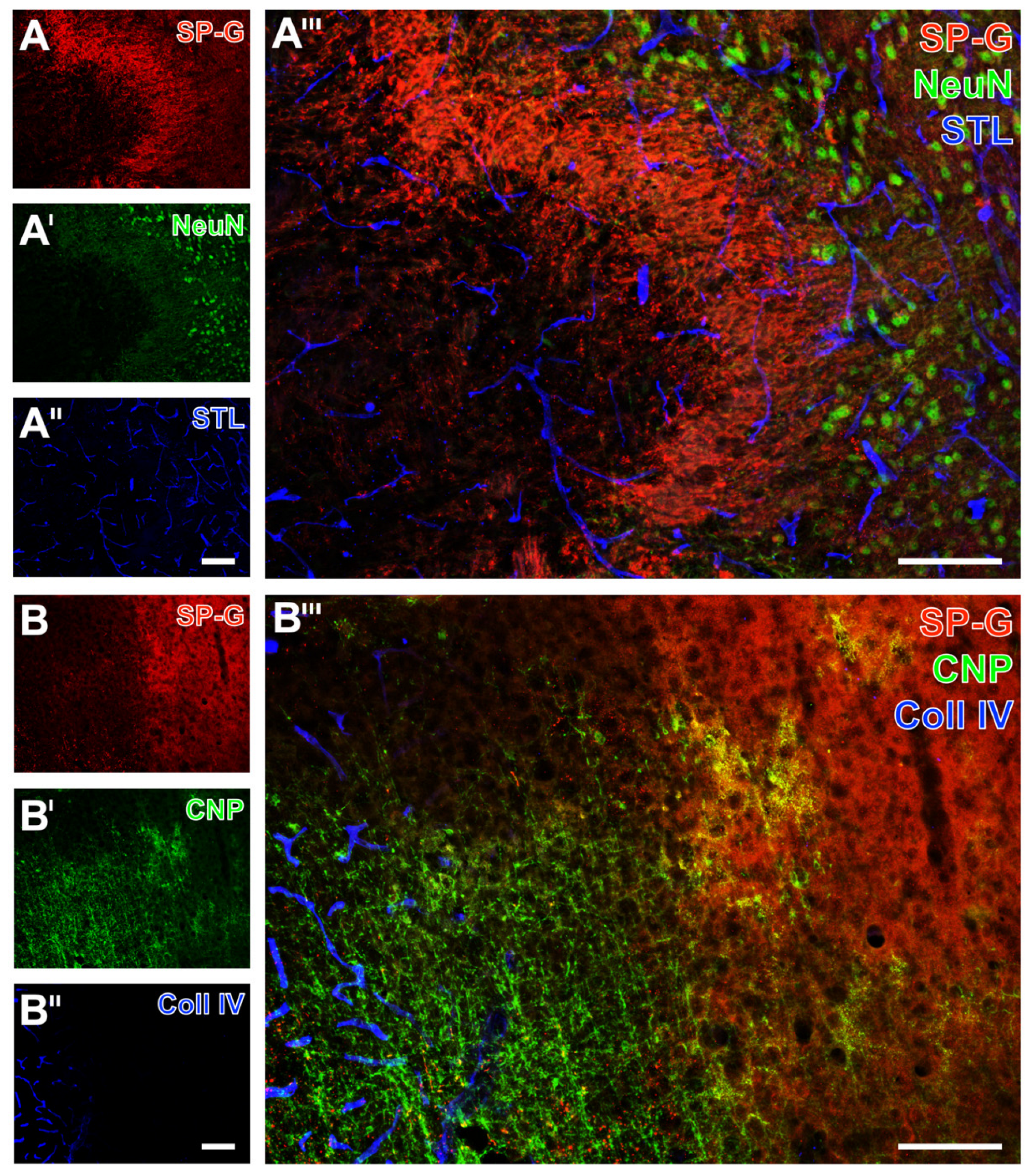
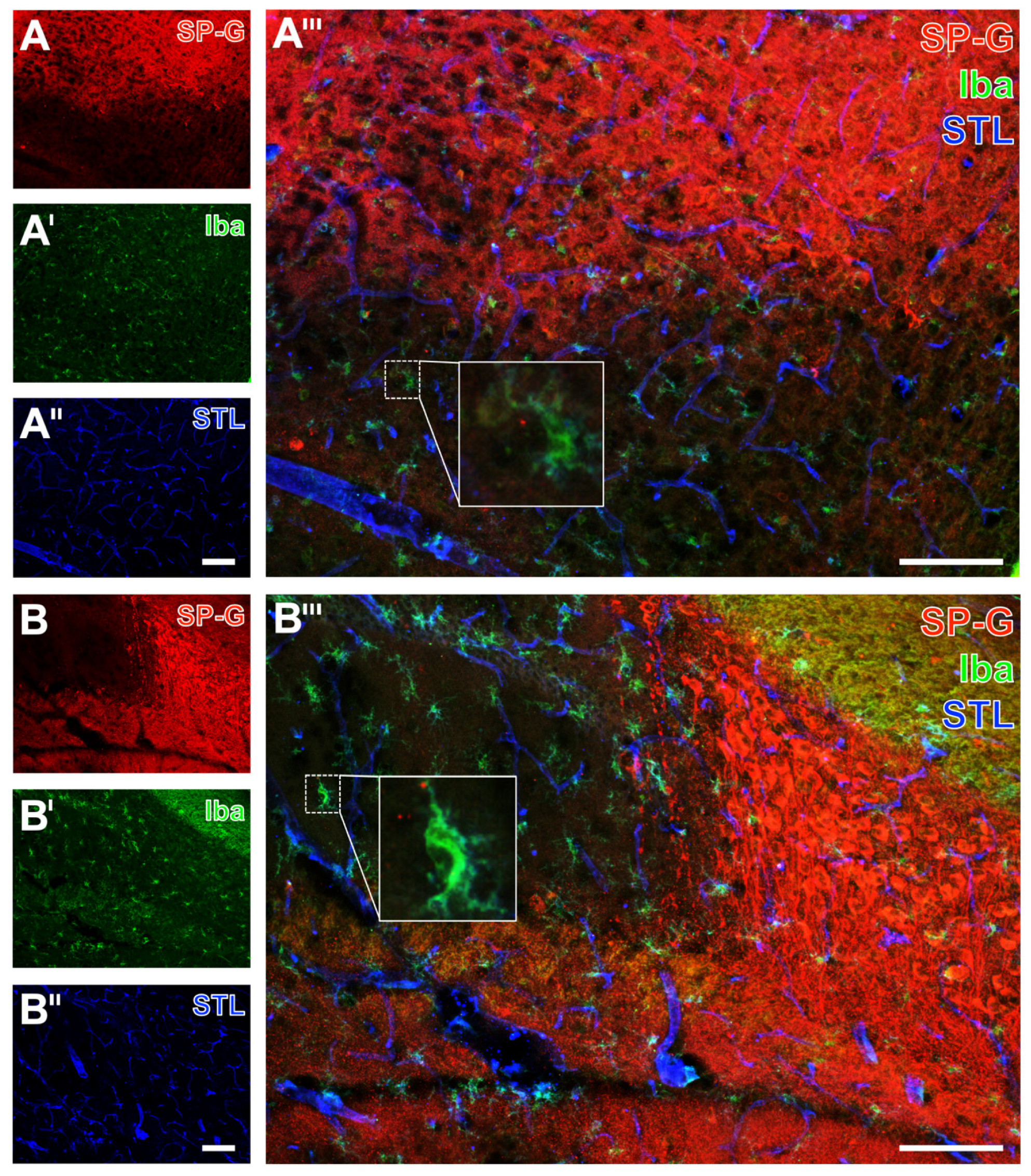
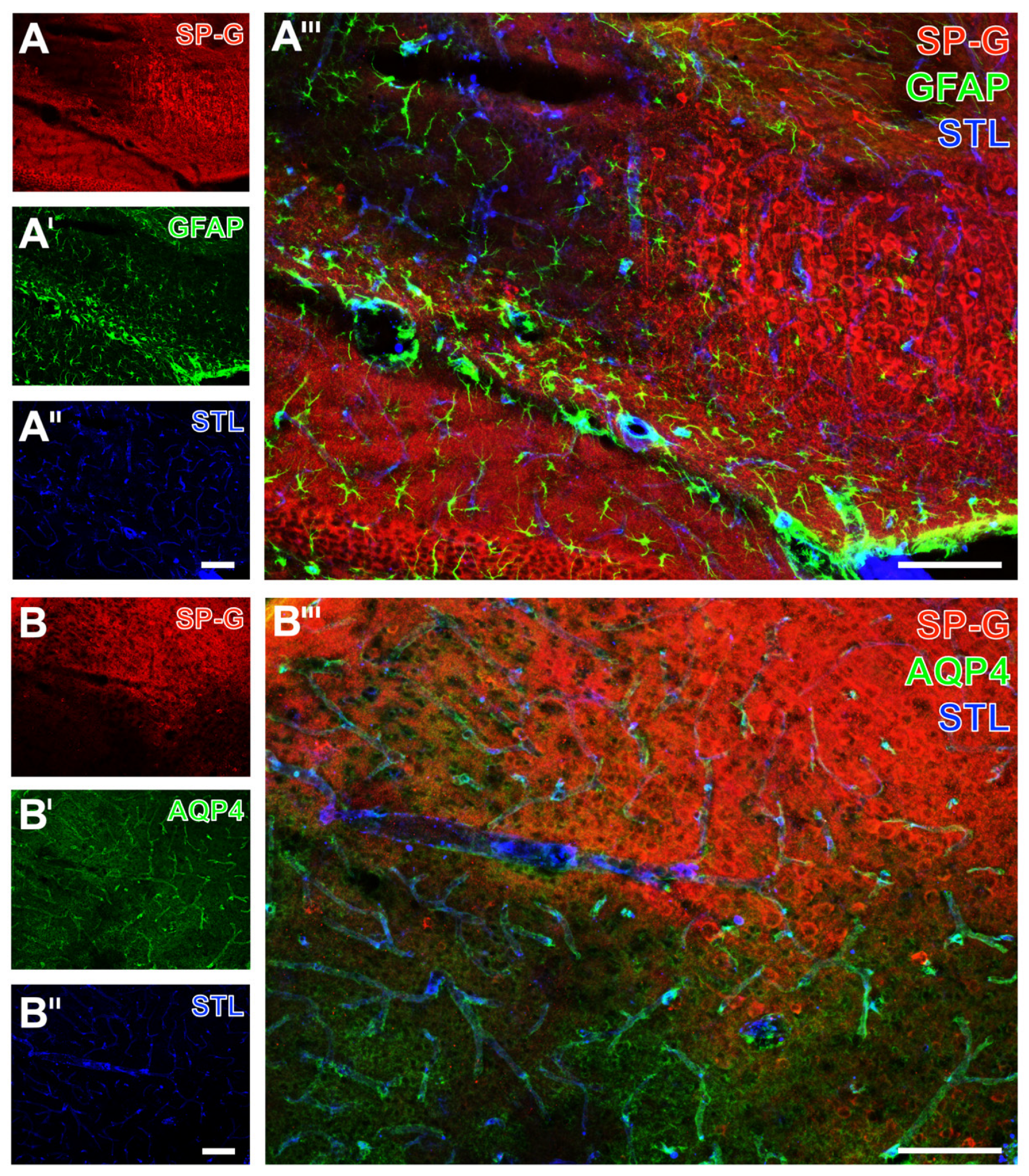
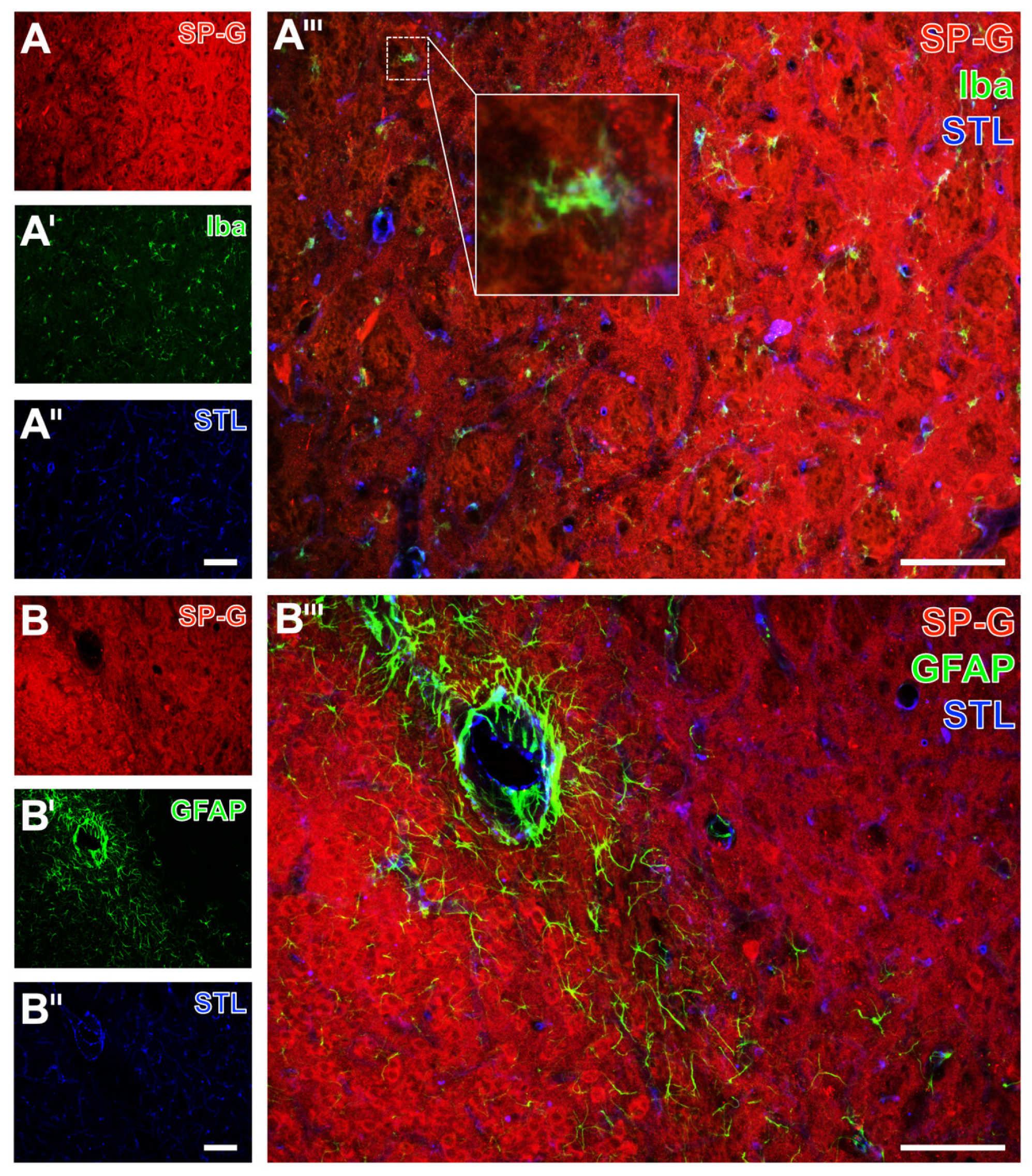
2.2. Regional Characteristics of SP-G and Neurovascular Unit Elements after Ischemia
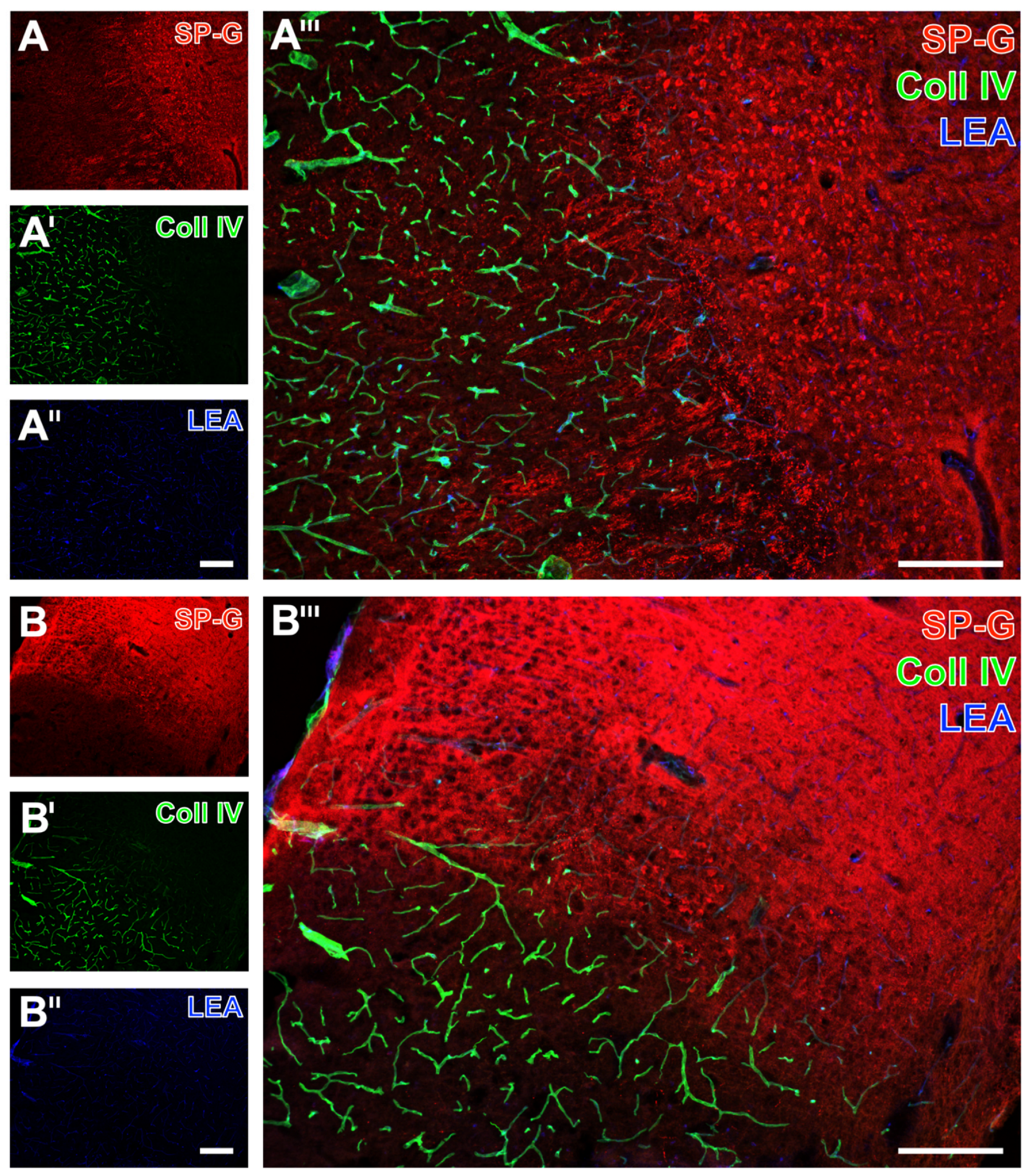
2.3. SP-G and the Associated Vasculature in Ischemia-Affected Brains from Rat and Sheep
3. Discussion
4. Materials and Methods
4.1. Study Design and Content
4.2. Experimental Focal Cerebral Ischemia in Mice, Rats, and Sheep
4.3. Tissue Preparation and Fluorescence Labeling
4.4. Microscopy and Processing of Images
4.5. Quantification of Fluorescence Signals
4.6. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hankey, G.J. Stroke. Lancet 2017, 389, 641–654. [Google Scholar] [CrossRef]
- Feigin, V.L.; Brainin, M.; Norrving, B.; Martins, S.; Sacco, R.L.; Hacke, W.; Fisher, M.; Pandian, J.; Lindsay, P. World Stroke Organization (WSO): Global stroke fact sheet 2022. Int. J. Stroke 2022, 17, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Dirnagl, U.; Iadecola, C.; Moskowitz, M.A. Pathobiology of ischaemic stroke: An integrated view. Trends Neurosci. 1999, 22, 391–397. [Google Scholar] [CrossRef]
- del Zoppo, G.J. The neurovascular unit in the setting of stroke. J. Intern. Med. 2010, 267, 156–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaeffer, S.; Iadecola, C. Revisiting the neurovascular unit. Nat. Neurosci. 2021, 24, 1198–1209. [Google Scholar] [CrossRef] [PubMed]
- Michalski, D.; Grosche, J.; Pelz, J.; Schneider, D.; Weise, C.; Bauer, U.; Kacza, J.; Gärtner, U.; Hobohm, C.; Härtig, W. A novel quantification of blood-brain barrier damage and histochemical typing after embolic stroke in rats. Brain Res. 2010, 1359, 186–200. [Google Scholar] [CrossRef]
- Michalski, D.; Pitsch, R.; Pillai, D.R.; Mages, B.; Aleithe, S.; Grosche, J.; Martens, H.; Schlachetzki, F.; Härtig, W. Delayed histochemical alterations within the neurovascular unit due to transient focal cerebral ischemia and experimental treatment with neurotrophic factors. PLoS ONE 2017, 12, e0174996. [Google Scholar] [CrossRef] [Green Version]
- Michalski, D.; Keck, A.L.; Grosche, J.; Martens, H.; Härtig, W. Immunosignals of oligodendrocyte markers and myelin-associated proteins are critically affected after experimental stroke in wild-type and alzheimer modeling mice of different ages. Front. Cell Neurosci. 2018, 12, 23. [Google Scholar] [CrossRef] [Green Version]
- Kestner, R.I.; Mayser, F.; Vutukuri, R.; Hansen, L.; Günther, S.; Brunkhorst, R.; Devraj, K.; Pfeilschifter, W. Gene Expression Dynamics at the neurovascular unit during early regeneration after cerebral ischemia/reperfusion injury in mice. Front. Neurosci. 2020, 14, 280. [Google Scholar] [CrossRef] [Green Version]
- Bernardo-Castro, S.; Sousa, J.A.; Brás, A.; Cecília, C.; Rodrigues, B.; Almendra, L.; Machado, C.; Santo, G.; Silva, F.; Ferreira, L.; et al. Pathophysiology of blood-brain barrier permeability throughout the different stages of ischemic stroke and its implication on hemorrhagic transformation and recovery. Front. Neurol. 2020, 11, 594672. [Google Scholar] [CrossRef]
- Krueger, M.; Bechmann, I.; Immig, K.; Reichenbach, A.; Härtig, W.; Michalski, D. Blood-brain barrier breakdown involves four distinct stages of vascular damage in various models of experimental focal cerebral ischemia. J. Cereb. Blood Flow. Metab. 2015, 35, 292–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, F.C.; Churilov, L.; Yassi, N.; Kleinig, T.J.; Thijs, V.; Wu, T.Y.; Shah, D.G.; Dewey, H.M.; Sharma, G.; Desmond, P.M.; et al. Microvascular dysfunction in blood-brain barrier disruption and hypoperfusion within the infarct posttreatment are associated with cerebral edema. Stroke 2022, 53, 1597–1605. [Google Scholar] [CrossRef] [PubMed]
- Mestre, H.; Du, T.; Sweeney, A.M.; Liu, G.; Samson, A.J.; Peng, W.; Mortensen, K.N.; Stæger, F.F.; Bork, P.A.R.; Bashford, L.; et al. Cerebrospinal fluid influx drives acute ischemic tissue swelling. Science 2020, 367, eaax7171. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Shao, L.; Ma, L. Cerebral edema formation after stroke: Emphasis on blood-brain barrier and the lymphatic drainage system of the brain. Front. Cell Neurosci. 2021, 15, 716825. [Google Scholar] [CrossRef] [PubMed]
- Lv, T.; Zhao, B.; Hu, Q.; Zhang, X. The glymphatic system: A novel therapeutic target for stroke treatment. Front. Aging Neurosci. 2021, 13, 689098. [Google Scholar] [CrossRef]
- Zhou, X.; Li, Y.; Lenahan, C.; Ou, Y.; Wang, M.; He, Y. Glymphatic system in the central nervous system, a novel therapeutic direction against brain edema after stroke. Front. Aging Neurosci. 2021, 13, 698036. [Google Scholar] [CrossRef]
- Ji, C.; Yu, X.; Xu, W.; Lenahan, C.; Tu, S.; Shao, A. The role of glymphatic system in the cerebral edema formation after ischemic stroke. Exp. Neurol. 2021, 340, 113685. [Google Scholar] [CrossRef]
- Mestre, H.; Mori, Y.; Nedergaard, M. The brain’s glymphatic system: Current controversies. Trends Neurosci. 2020, 43, 458–466. [Google Scholar] [CrossRef]
- Mader, S.; Brimberg, L. Aquaporin-4 water channel in the brain and its implication for health and disease. Cells 2019, 8, 90. [Google Scholar] [CrossRef] [Green Version]
- MacAulay, N. Molecular mechanisms of brain water transport. Nat. Rev. Neurosci. 2021, 22, 326–344. [Google Scholar] [CrossRef]
- Wolburg, H.; Noell, S.; Wolburg-Buchholz, K.; Mack, A.; Fallier-Becker, P. Agrin, aquaporin-4, and astrocyte polarity as an important feature of the blood-brain barrier. Neuroscientist 2009, 15, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Mulder, I.A.; van Bavel, E.T.; de Vries, H.E.; Coutinho, J.M. Adjunctive cytoprotective therapies in acute ischemic stroke: A systematic review. Fluids Barriers CNS 2021, 18, 46. [Google Scholar] [CrossRef] [PubMed]
- O’Collins, V.E.; Macleod, M.R.; Donnan, G.A.; Horky, L.L.; van der Worp, B.H.; Howells, D.W. 1026 Experimental treatments in acute stroke. Ann. Neurol. 2006, 59, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Griese, M. Pulmonary surfactant in health and human lung diseases: State of the art. Eur. Respir. J. 1999, 13, 1455–1476. [Google Scholar] [CrossRef] [PubMed]
- Schob, S.; Weiß, A.; Surov, A.; Dieckow, J.; Richter, C.; Pirlich, M.; Horvath-Rizea, D.; Härtig, W.; Hoffmann, K.T.; Krause, M.; et al. Elevated surfactant protein levels and increased flow of cerebrospinal fluid in cranial magnetic resonance imaging. Mol. Neurobiol. 2018, 55, 6227–6236. [Google Scholar] [CrossRef] [PubMed]
- Schob, S.; Schicht, M.; Sel, S.; Stiller, D.; Kekulé, A.S.; Paulsen, F.; Maronde, E.; Bräuer, L. The detection of surfactant proteins A, B, C and D in the human brain and their regulation in cerebral infarction, autoimmune conditions and infections of the CNS. PLoS ONE. 2013, 8, e74412. [Google Scholar] [CrossRef]
- Rausch, F.; Schicht, M.; Paulsen, F.; Ngueya, I.; Bräuer, L.; Brandt, W. “SP-G”, a putative new surfactant protein--tissue localization and 3D structure. PLoS ONE 2012, 7, e47789. [Google Scholar] [CrossRef]
- Meinicke, A.; Härtig, W.; Winter, K.; Puchta, J.; Mages, B.; Michalski, D.; Emmer, A.; Otto, M.; Hoffmann, K.T.; Reimann, W.; et al. Surfactant protein-G in wildtype and 3xTg-AD mice: Localization in the forebrain, age-dependent hippocampal dot-like deposits and brain content. Biomolecules 2022, 12, 96. [Google Scholar] [CrossRef]
- Krause, M.; Peukert, N.; Härtig, W.; Emmer, A.; Mahr, C.V.; Richter, C.; Dieckow, J.; Puchta, J.; Pirlich, M.; Hoffmann, K.T.; et al. Localization, occurrence, and CSF changes of SP-G, a new surface active protein with assumable immunoregulatory functions in the CNS. Mol. Neurobiol. 2019, 56, 2433–2439. [Google Scholar] [CrossRef]
- Fisher, M.; Feuerstein, G.; Howells, D.W.; Hurn, P.D.; Kent, T.A.; Savitz, S.I.; Lo, E.H.; STAIR Group. Update of the stroke therapy academic industry roundtable preclinical recommendations. Stroke 2009, 40, 2244–2250. [Google Scholar] [CrossRef]
- Durukan, A.; Tatlisumak, T. Acute ischemic stroke: Overview of major experimental rodent models, pathophysiology, and therapy of focal cerebral ischemia. Pharmacol. Biochem. Behav. 2007, 87, 179–197. [Google Scholar] [CrossRef] [PubMed]
- Sommer, C.J. Ischemic stroke: Experimental models and reality. Acta Neuropathol. 2017, 133, 245–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michalski, D.; Spielvogel, E.; Puchta, J.; Reimann, W.; Barthel, H.; Nitzsche, B.; Mages, B.; Jäger, C.; Martens, H.; Horn, A.K.E.; et al. Increased immunosignals of collagen IV and fibronectin indicate ischemic consequences for the neurovascular matrix adhesion zone in various animal models and human stroke tissue. Front. Physiol. 2020, 11, 575598. [Google Scholar] [CrossRef] [PubMed]
- Mages, B.; Aleithe, S.; Blietz, A.; Krueger, M.; Härtig, W.; Michalski, D. Simultaneous alterations of oligodendrocyte-specific CNP, astrocyte-specific AQP4 and neuronal NF-L demarcate ischemic tissue after experimental stroke in mice. Neurosci. Lett. 2019, 711, 134405. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Carraher, C.; Schwarzbauer, J.E. Assembly of fibronectin extracellular matrix. Ann. Rev. Cell Dev. Biol. 2010, 26, 397–419. [Google Scholar] [CrossRef] [Green Version]
- Krueger, M.; Mages, B.; Hobusch, C.; Michalski, D. Endothelial edema precedes blood-brain barrier breakdown in early time points after experimental focal cerebral ischemia. Acta Neuropathol. Commun. 2019, 7, 17. [Google Scholar] [CrossRef]
- Pillai, D.R.; Dittmar, M.S.; Baldaranov, D.; Heidemann, R.M.; Henning, E.C.; Schuierer, G.; Bogdahn, U.; Schlachetzki, F. Cerebral ischemia-reperfusion injury in rats—A 3 T MRI study on biphasic blood-brain barrier opening and the dynamics of edema formation. J. Cereb. Blood Flow Metab. 2009, 29, 1846–1855. [Google Scholar] [CrossRef]
- Strbian, D.; Durukan, A.; Pitkonen, M.; Marinkovic, I.; Tatlisumak, E.; Pedrono, E.; Abo-Ramadan, U.; Tatlisumak, T. The blood-brain barrier is continuously open for several weeks following transient focal cerebral ischemia. Neuroscience 2008, 153, 175–181. [Google Scholar] [CrossRef]
- Nielsen, S.; Nagelhus, E.A.; Amiry-Moghaddam, M.; Bourque, C.; Agre, P.; Ottersen, O.P. Specialized membrane domains for water transport in glial cells: High-resolution immunogold cytochemistry of aquaporin-4 in rat brain. J. Neurosci. 1997, 17, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, M.K.; Mestre, H.; Nedergaard, M. The glymphatic pathway in neurological disorders. Lancet Neurol. 2018, 17, 1016–1024. [Google Scholar] [CrossRef] [Green Version]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving bioscience research reporting: The ARRIVE guidelines for reporting animal research. PLoS Biol. 2010, 8, e1000412. [Google Scholar] [CrossRef] [PubMed]
- Longa, E.Z.; Weinstein, P.R.; Carlson, S.; Cummins, R. Reversible middle cerebral artery occlusion without craniectomy in rats. Stroke 1989, 20, 84–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawkes, C.A.; Michalski, D.; Anders, R.; Nissel, S.; Grosche, J.; Bechmann, I.; Carare, R.O.; Härtig, W. Stroke-induced opposite and age-dependent changes of vessel-associated markers in co-morbid transgenic mice with Alzheimer-like alterations. Exp. Neurol. 2013, 250, 270–281. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.L.; Chopp, M.; Zhang, Z.G.; Jiang, Q.; Ewing, J.R. A rat model of focal embolic cerebral ischemia. Brain Res. 1997, 766, 83–92. [Google Scholar] [CrossRef]
- Menzies, S.A.; Hoff, J.T.; Betz, A.L. Middle cerebral artery occlusion in rats: A neurological and pathological evaluation of a reproducible model. Neurosurgery 1992, 31, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Nitzsche, B.; Barthel, H.; Lobsien, D.; Boltze, J.; Zeisig, V.; Dreyer, A.Y. Focal cerebral ischemia by permanent middle cerebral artery occlusion in sheep: Surgical technique, clinical imaging and histopathological results. In Experimental Neurosurgery in Animal Models; Janowski, M., Ed.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 195–225. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Host Species | Supplier | Product Number | Dilution | Fluoro-phor |
|---|---|---|---|---|---|
| Albumin | Sheep | AbD Serotec, Oxford, UK | 0220-2424 | 1:500 | Cy2 |
| Aquaporin 4 | Guinea pig | Synaptic Systems, Göttingen, Germany | 429004 | 1:200 | Cy2 |
| CNP | Guinea pig | Synaptic Systems | 355004 | 1:200 | Cy2 |
| Collagen IV | Goat | Merck Millipore; Billerica, MA, USA | AB749 | 1:100 | Alexa Fluor647 |
| Fibronectin | Sheep | Bio-Techne; Wies-baden, Germany * | AF1918 | 1:100 | Cy2 |
| GFAP | Guinea pig | Synaptic Systems | 173004 | 1:200 | Cy2 |
| Iba | Guinea pig | Synaptic Systems | 234004 | 1:100 | Cy2 |
| NeuN | Guinea pig | Synaptic Systems | 266004 | 1:200 | Cy2 |
| SP-G | Rabbit | Hölzel; Cologne, Germany ** | PAD755 Hu01 | 1:70 *** | Cy3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michalski, D.; Reimann, W.; Spielvogel, E.; Mages, B.; Biedermann, B.; Barthel, H.; Nitzsche, B.; Schob, S.; Härtig, W. Regionally Altered Immunosignals of Surfactant Protein-G, Vascular and Non-Vascular Elements of the Neurovascular Unit after Experimental Focal Cerebral Ischemia in Mice, Rats, and Sheep. Int. J. Mol. Sci. 2022, 23, 5875. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23115875
Michalski D, Reimann W, Spielvogel E, Mages B, Biedermann B, Barthel H, Nitzsche B, Schob S, Härtig W. Regionally Altered Immunosignals of Surfactant Protein-G, Vascular and Non-Vascular Elements of the Neurovascular Unit after Experimental Focal Cerebral Ischemia in Mice, Rats, and Sheep. International Journal of Molecular Sciences. 2022; 23(11):5875. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23115875
Chicago/Turabian StyleMichalski, Dominik, Willi Reimann, Emma Spielvogel, Bianca Mages, Bernd Biedermann, Henryk Barthel, Björn Nitzsche, Stefan Schob, and Wolfgang Härtig. 2022. "Regionally Altered Immunosignals of Surfactant Protein-G, Vascular and Non-Vascular Elements of the Neurovascular Unit after Experimental Focal Cerebral Ischemia in Mice, Rats, and Sheep" International Journal of Molecular Sciences 23, no. 11: 5875. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23115875