
A Potential Role of Keratinocyte-Derived Bilirubin in Human Skin Yellowness and Its Amelioration by Sucrose Laurate/Dilaurate

 ,
,
Abstract
:1. Introduction
2. Results
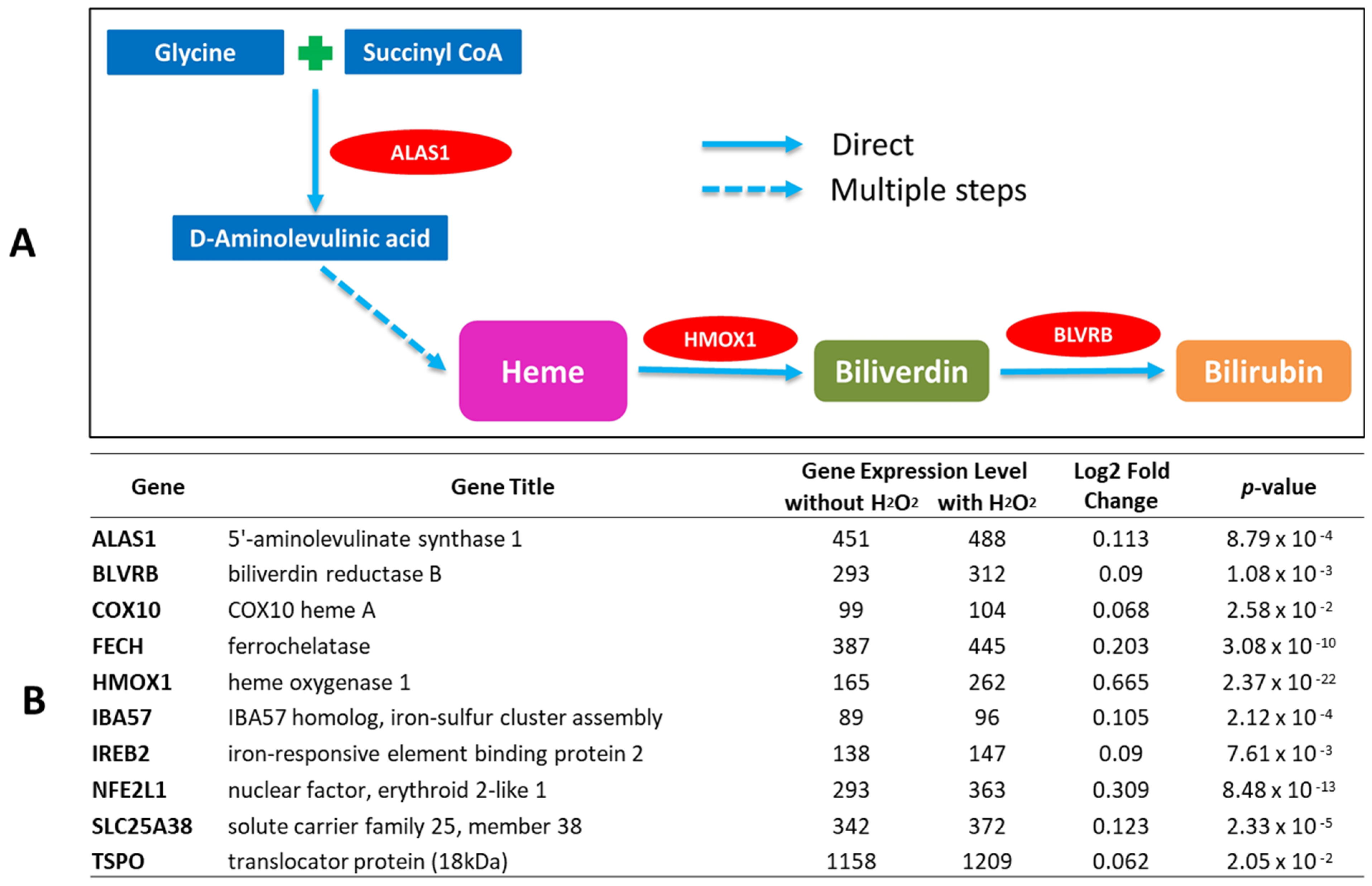
2.1. Bilirubin-Synthesis-Related Genes Are Expressed in Human Keratinocytes and Upregulated by Oxidative Stress
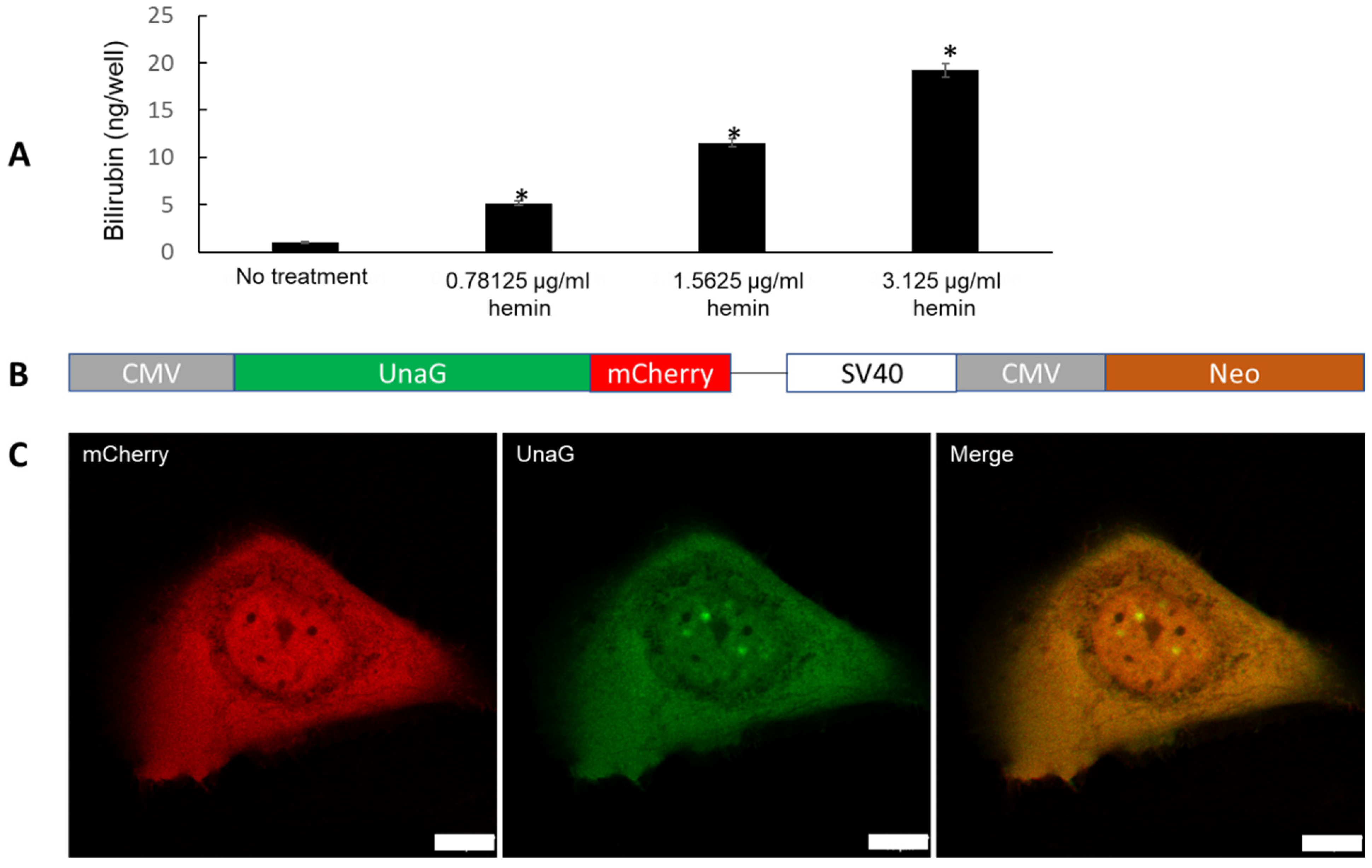
2.2. Evidence for De Novo Synthesis of Bilirubin via Heme Synthesis in Human Keratinocytes
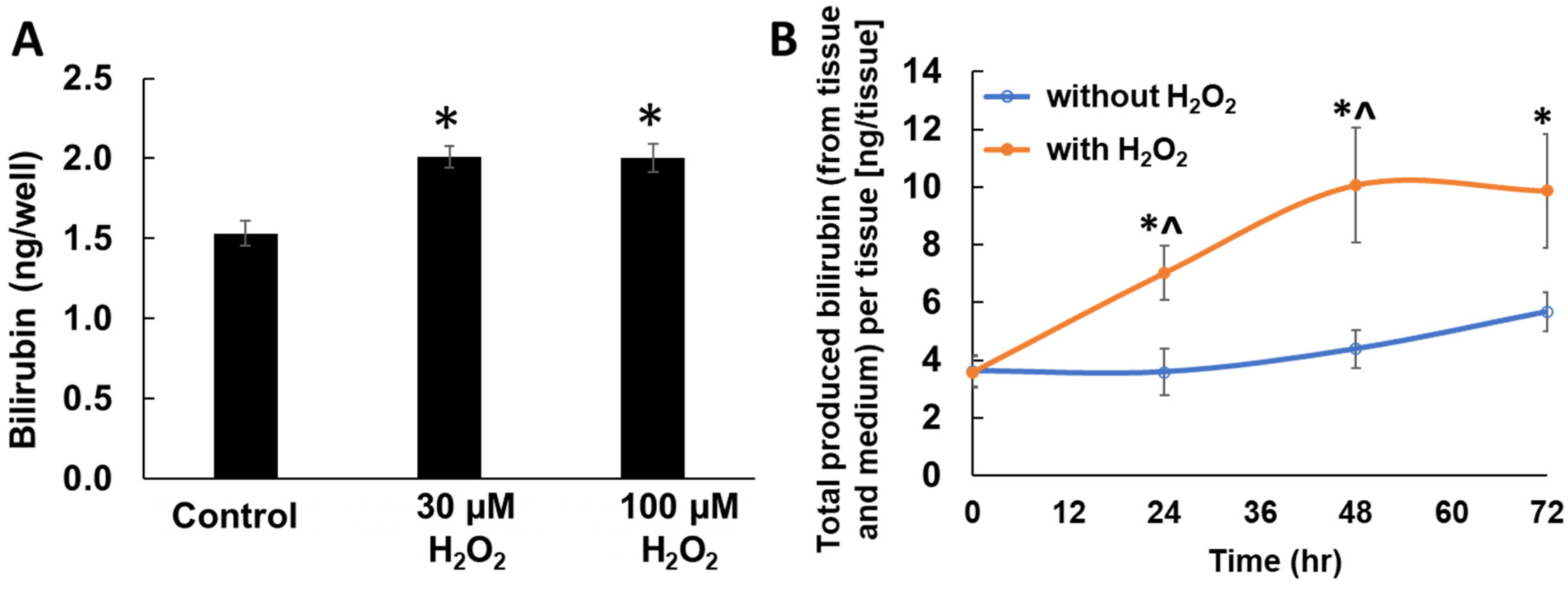
2.3. Enhanced Bilirubin Production by H2O2 Treatment in Keratinocytes
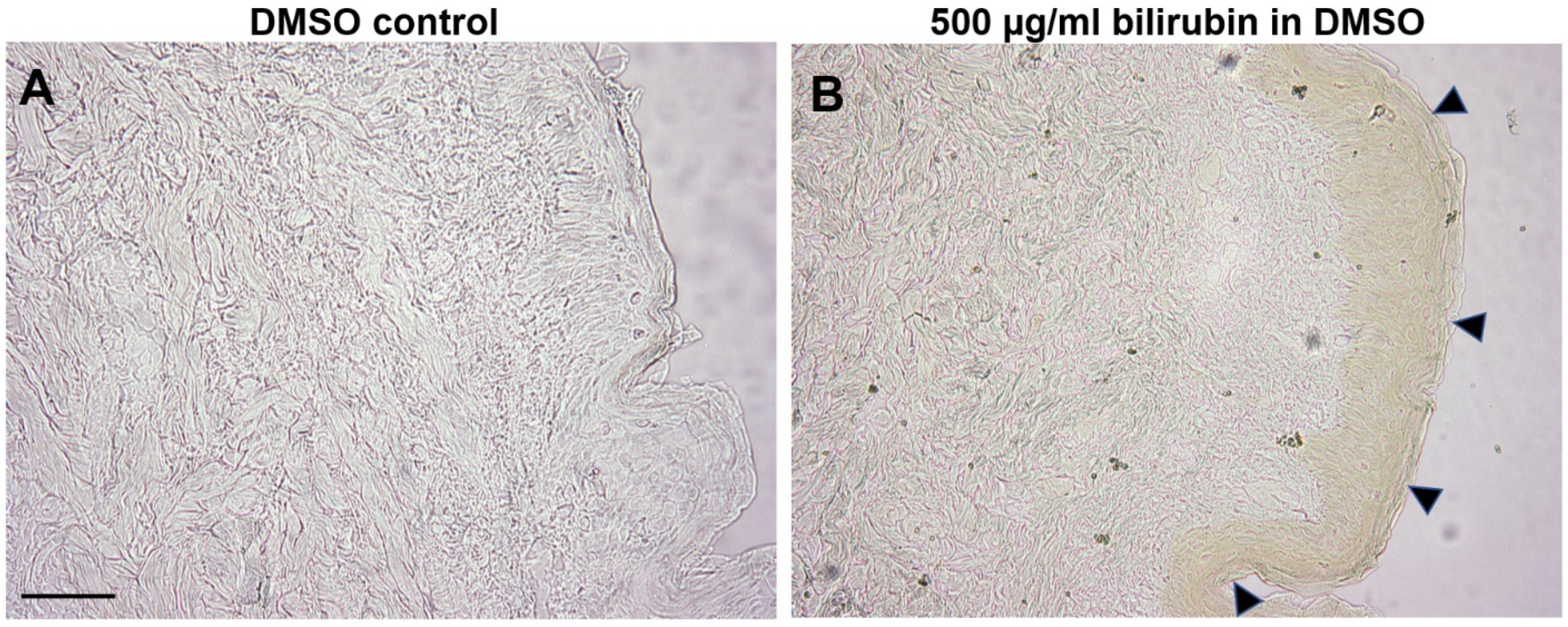
2.4. Bilirubin Has Natural Affinity toward Epidermis of Human Skin
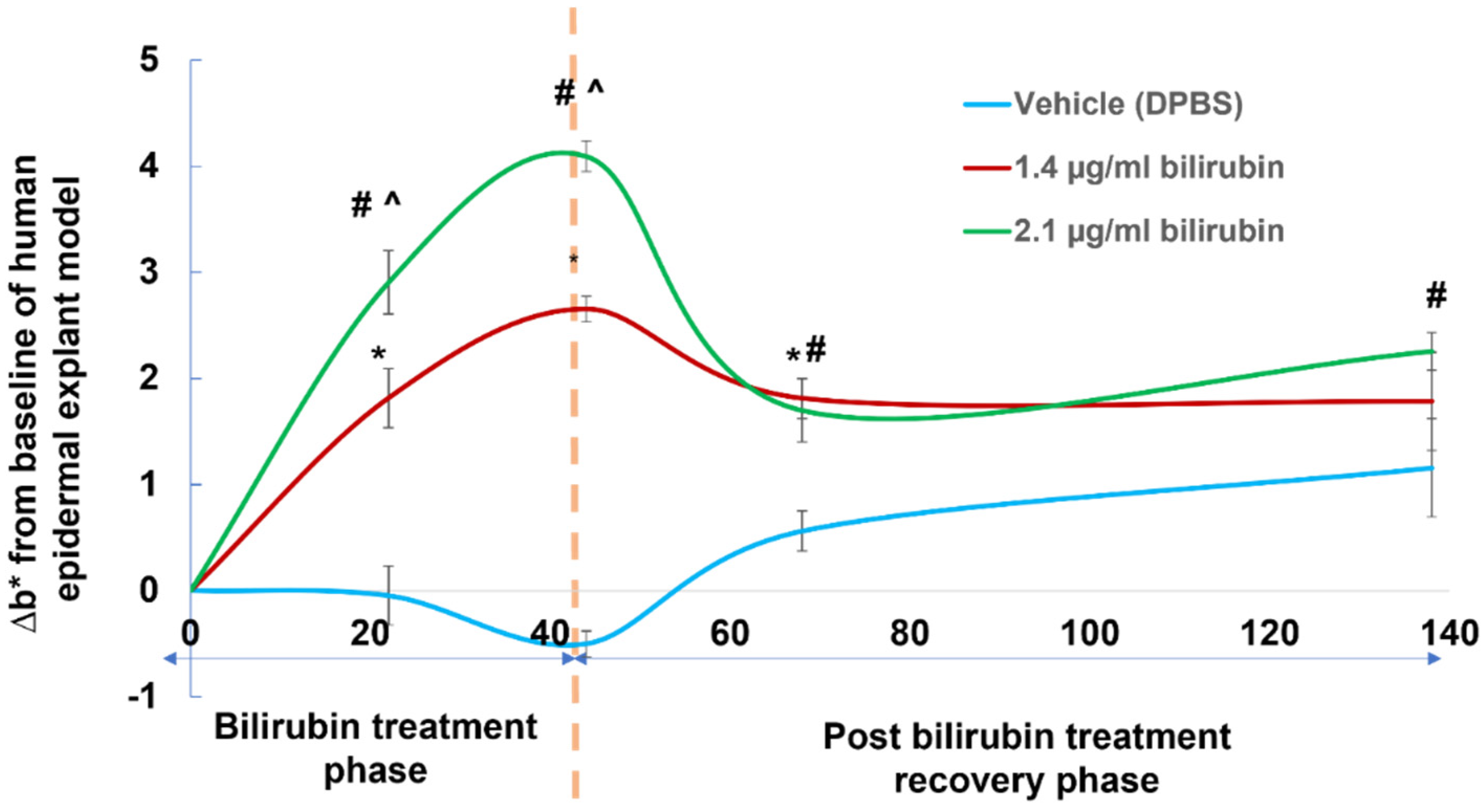
2.5. Assessment of Yellowness (b* Value) in Human Epidermal Explant Model Treated with Bioavailable Concentrations of Bilirubin
2.6. Transcriptomic Profiling Analysis of Keratinocytes by Bilirubin Treatment
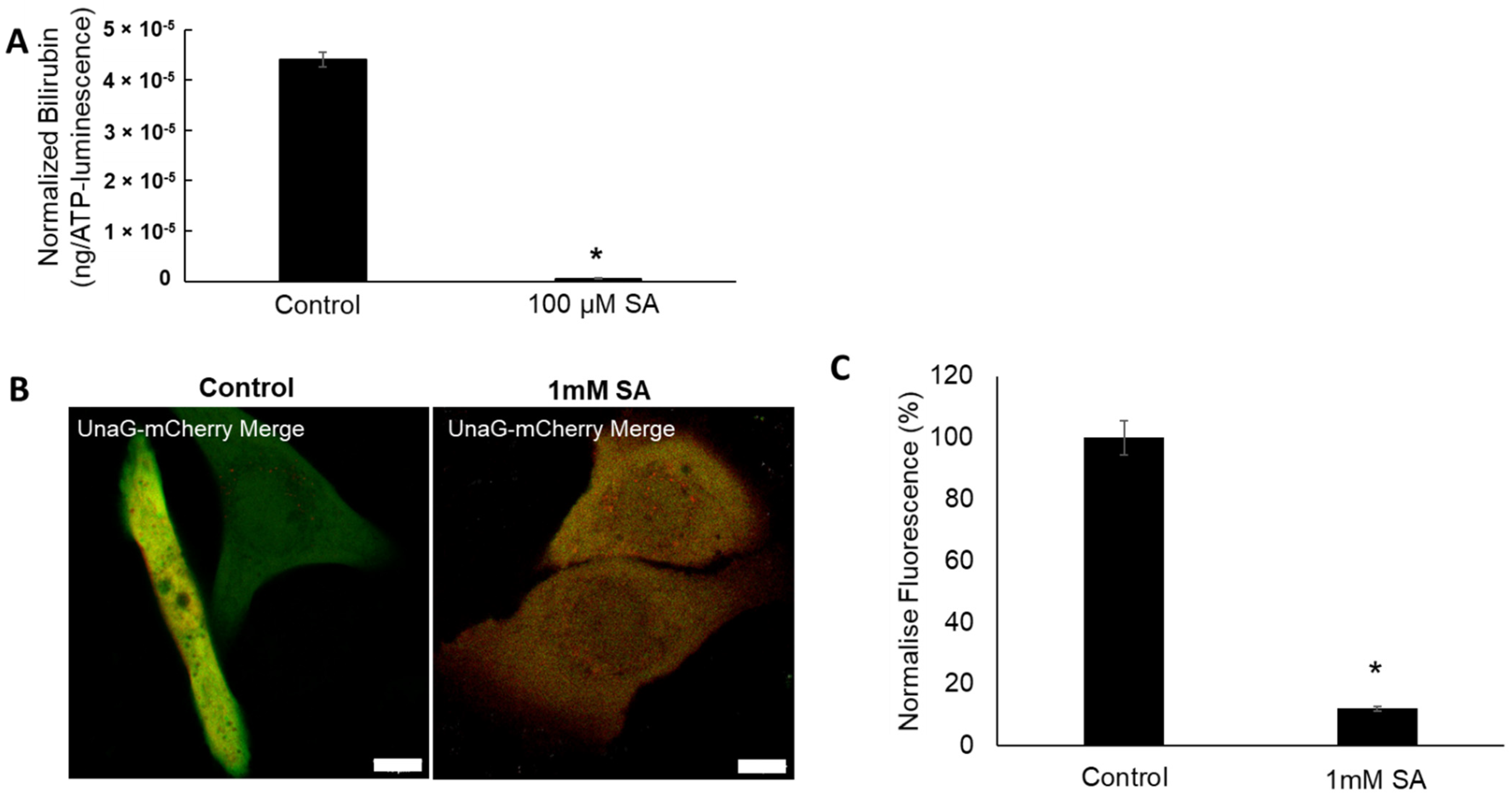
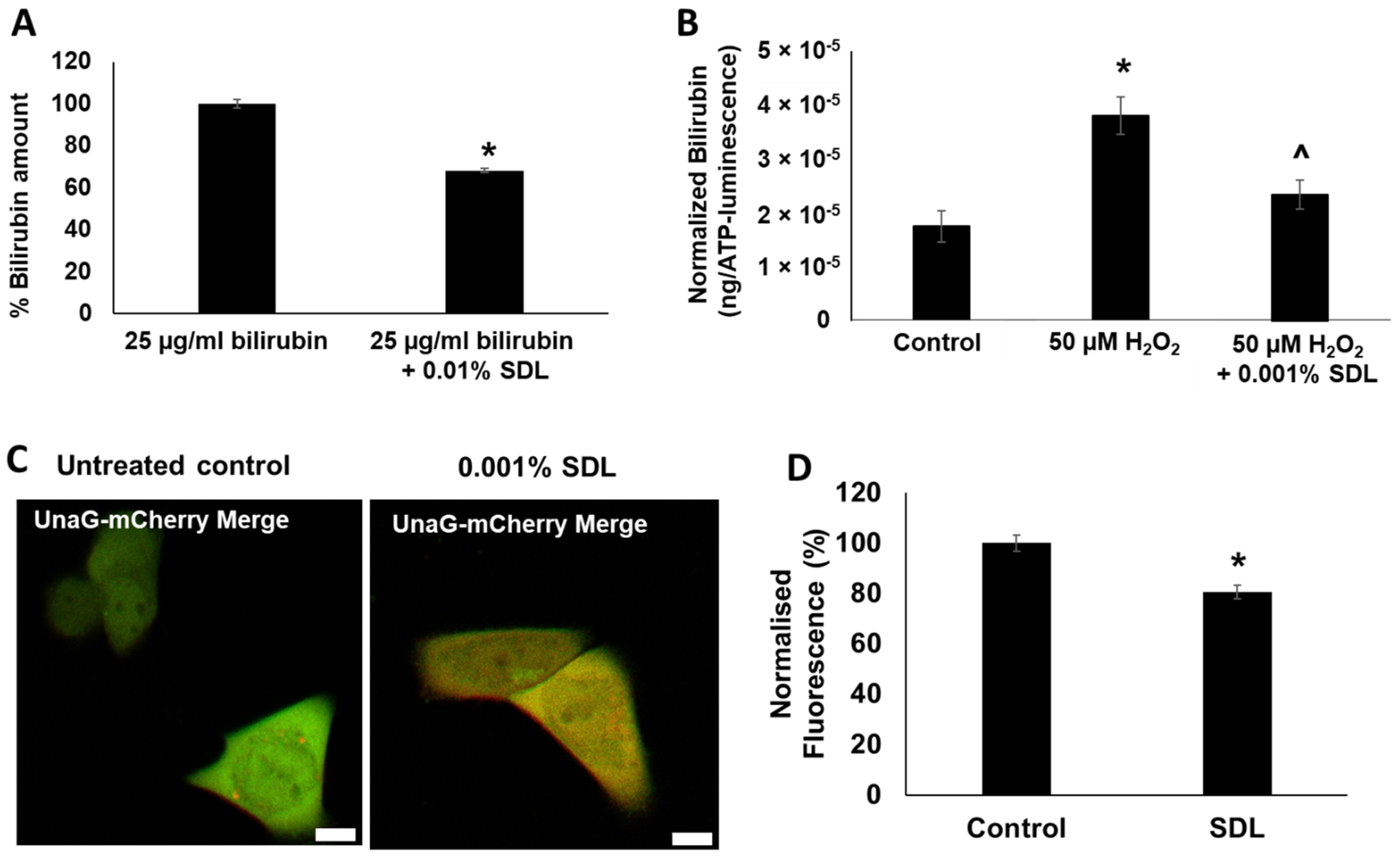
2.7. SDL Diminishes Bilirubin Levels in Keratinocytes
3. Discussion
4. Materials and Methods
4.1. Chemicals, Reagents, and Cell Lines
4.2. tKC Culture, Treatment, and Viability Assessment
4.3. Oxidative Stress Challenge to 3D Human Skin-Equivalent Model
4.4. UnaG Bilirubin Quantification Method for Cultured tKC
4.5. Bilirubin Quantification to Assess SDL Chemical Impact on Bilirubin Level
4.6. Human Epidermal Explant Model Treatment with Bilirubin
4.7. Bilirubin Quantification in Biological Samples Using LC-MS/MS
- -
- Multiple reaction monitoring (MRM) transitions for analytes (bilirubin: 585.2 → 299.2);
- -
- Their corresponding stable isotope-labeled internal standards (d4-bilirubin: 589.2 → 301.2).
4.8. Estimation of Bilirubin Concentration Produced in EpiDerm™ 3D Human Skin-Equivalent Model
4.9. Sample Preparation and Data Analysis for Microarray Studies
4.10. Visualization of Bilirubin Content within Cells Using UnaG Fluorescence
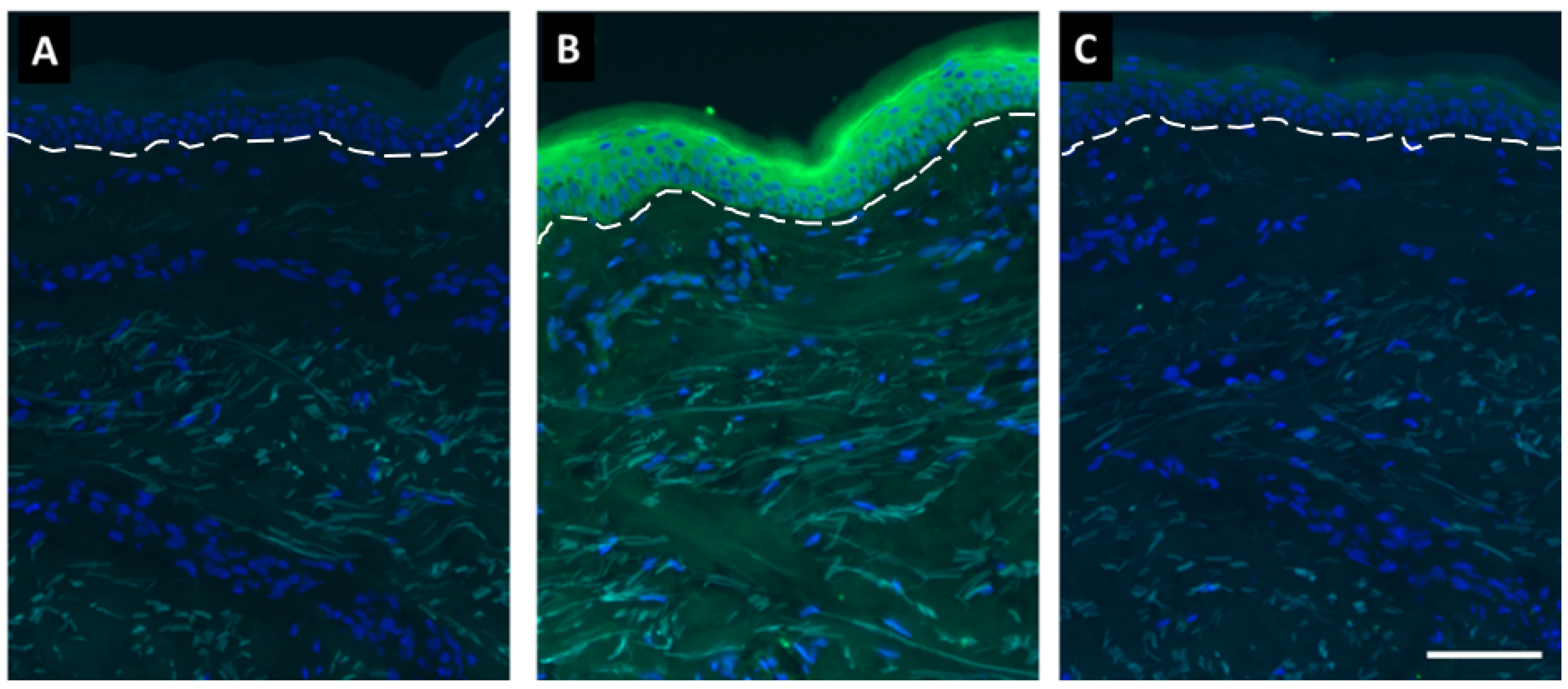
4.11. Visualization of SDL Effect on Bilirubin Reduction in Human Skin Sections
4.12. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Everson, C.A.; Thalacker, C.D.; Hogg, N. Phagocyte migration and cellular stress induced in liver, lung, and intestine during sleep loss and sleep recovery. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, R2067–R2074. [Google Scholar] [CrossRef] [PubMed]
- Buyukhatipoglu, H.; Kirhan, I.; Vural, M.; Taskin, A.; Sezen, Y.; Dag, O.F.; Turan, M.N.; Aksoy, N. Oxidative stress increased in healthcare workers working 24-hour on-call shifts. Am. J. Med. Sci. 2010, 340, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Moller-Levet, C.S.; Archer, S.N.; Bucca, G. Effects of insufficient sleep on circadian rhythmicity and expression amplitude of the human blood transcriptome. Proc. Natl. Acad. Sci. USA 2013, 110, E1132–E1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopresti, A.L.; Hood, S.D.; Drummond, P.D. A review of lifestyle factors that contribute to important pathways associated with major depression: Diet, sleep and exercise. J. Affect. Disord. 2013, 148, 12–27. [Google Scholar] [CrossRef] [Green Version]
- Weljie, A.M.; Meerlo, P.; Goel, N.; Sengupta, A.; Kayser, M.S.; Abel, T.; Birnbaum, M.J.; Dinges, D.F.; Sehgal, A. Oxalic acid and diacylglycerol 36:3 are cross-species markers of sleep debt. Proc. Natl. Acad. Sci. USA 2015, 112, 2569–2574. [Google Scholar] [CrossRef] [Green Version]
- Stocker, R.; Yamamoto, Y.; McDonagh, A.F.; Glazer, A.N.; Ames, B.N. Bilirubin is an antioxidant of possible physiological importance. Science 1987, 235, 1043–1046. [Google Scholar] [CrossRef]
- Schiff, L. Jaundice: Five-and-a-half decades in historic perspective. Selected aspects. Gastroenterology 1980, 78, 831–836. [Google Scholar] [CrossRef]
- Maines, M.D.; Trakshel, G.M.; Kutty, R.K. Characterization of two constitutive forms of rat liver microsomal heme oxygenase. Only one molecular species of the enzyme is inducible. J. Biol. Chem. 1986, 261, 411–419. [Google Scholar] [CrossRef]
- Braggins, P.E.; Trakshel, G.M.; Kutty, R.K.; Maines, M.D. Characterization of two heme oxygenase isoforms in rat spleen: Comparison with the hematin-induced and constitutive isoforms of the liver. Biochem. Biophys. Res. Commun. 1986, 141, 528–533. [Google Scholar] [CrossRef]
- Trakshel, G.M.; Kutty, R.K.; Maines, M.D. Purification and characterization of the major constitutive form of testicular heme oxygenase. The noninducible isoform. J. Biol. Chem. 1986, 261, 11131–11137. [Google Scholar] [CrossRef]
- Linnenbaum, M.; Busker, M.; Jan, R.; Kraehling, J.R.; Behrends, S. Heme oxygenase isoforms differ in their subcellular trafficking during hypoxia and are differentially modulated by cytochrome P450 reductase. PLoS ONE 2012, 7, e35483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenhunen, R.; Marver, H.S.; Schmid, R. The enzymatic conversion of heme to bilirubin by microsomal heme oxygenase. Proc. Natl. Acad. Sci. USA 1968, 61, 748–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, T.; Kikuchi, G. Purification and properties of heme oxygenase from rat liver microsomes. J. Biol. Chem. 1979, 254, 4487–4491. [Google Scholar] [CrossRef]
- Ryter, S.W.; Alam, J.; Choi, A.M. Heme oxygenase-1/carbon monoxide: From basic science to therapeutic applications. Physiol. Rev. 2006, 86, 583–650. [Google Scholar] [CrossRef] [PubMed]
- Chiabrando, D.; Vinchi, F.; Fiorito, V.; Mercurio, S.; Tolosano, E. Heme in pathophysiology: A matter of scavenging, metabolism and trafficking across cell membranes. Front. Pharmacol. 2014, 5, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- London, I.M.; West, R.; Shemin, D.; Rittenberg, D. On the origin of bile pigment in normal man. J. Biol. Chem. 1950, 184, 351–358. [Google Scholar] [CrossRef]
- Gray, C.H.; Neuberger, A.; Sneath, P.H. Studies in congenital porphyria. 2. Incorporation of 15N in the stercobilin in the normal and in the porphyric. Biochem. J. 1950, 47, 87–92. [Google Scholar] [CrossRef] [Green Version]
- Takeda, T.A.; Mu, A.; Tai, T.T.; Kitajima, S.; Taketani, S. Continuous de novo biosynthesis of haem and its rapid turnover to bilirubin are necessary for cytoprotection against cell damage. Sci. Rep. 2015, 5, 10488. [Google Scholar] [CrossRef] [Green Version]
- Numata, I.; Okuyama, R.; Memezawa, A.; Ito, Y.; Takeda, K.; Furuyama, K.; Shibahara, S.; Aiba, S. Functional expression of heme oxygenase-1 in human differentiated epidermis and its regulation by cytokines. J. Investig. Dermatol. 2009, 129, 2594–2603. [Google Scholar] [CrossRef] [Green Version]
- Shay, J.; Wright, W.E. Use of telomerase to create bioengineered tissues. Ann. N. Y. Acad. Sci. 2005, 1057, 479–491. [Google Scholar] [CrossRef]
- Rahman, M.N.; Vukomanovic, D.; Vlahakis, J.Z.; Szarek, W.A.; Nakatsu, K.; Jia, Z. Structural insights into human heme oxygenase-1 inhibition by potent and selective azole-based compounds. J. R. Soc. Interface 2013, 10, 20120697. [Google Scholar] [CrossRef] [PubMed]
- Beru, N.; Sahr, K.; Goldwasser, E. Inhibition of heme synthesis in bone marrow cells by succinylacetone: Effect on globin synthesis. J. Cell. Biochem. 1983, 21, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Sedlak, T.W.; Saleh, M.; Higginson, D.S.; Paul, B.D.; Juluri, K.R.; Snyder, S.H. Bilirubin and glutathione have complementary antioxidant and cytoprotective roles. Proc. Natl. Acad. Sci. USA 2009, 106, 5171–5176. [Google Scholar] [CrossRef] [Green Version]
- Packer, L.; Valacchi, G. Antioxidants and the response of skin to oxidative stress: Vitamin E as a key indicator. Ski. Pharmacol. Appl. Ski. Physiol. 2002, 15, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Muxin, G.; Nishida, H.; Shirakawa, C.; Sato, S.; Konishi, T. Psychological stress-induced oxidative stress as a model of sub-healthy condition and the effect of TCM. Evid. Based Complement. Alternat. Med. 2007, 4, 195–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salim, S. Oxidative stress and psychological disorders. Curr. Neuropharmacol. 2014, 12, 140–147. [Google Scholar] [CrossRef] [Green Version]
- Hellemans, L.; Corstjens, H.; Neven, A.; Declercq, L.; Maes, D. Antioxidant enzyme activity in human stratum corneum shows seasonal variation with an age-dependent recovery. J. Investig. Dermatol. 2003, 120, 434–439. [Google Scholar] [CrossRef] [Green Version]
- Zucker, S.D.; Horn, P.S.; Sherman, K.E. Serum bilirubin levels in the U.S. population: Gender effect and inverse correlation with colorectal cancer. Hepatology 2004, 40, 827–835. [Google Scholar] [CrossRef]
- Teixeira, K.R.C.; Dos Santos, C.P.; de Medeiros, L.A.; Mendes, J.A.; Cunha, T.M.; De Angelis, K.; Penha-Silva, N.; de Oliveira, E.P.; Crispim, C.A. Night workers have lower levels of antioxidant defenses and higher levels of oxidative stress damage when compared to day workers. Sci. Rep. 2019, 9, 4455. [Google Scholar] [CrossRef] [Green Version]
- Chin, K.; Ohi, M.; Shimizu, K.; Nakamura, T.; Miyaoka, F.; Mishima, M.; Nakamura, T. Increase in bilirubin levels of patients with obstructive sleep apnea in the morning-a possible explanation of induced heme oxygenase-1. Sleep 2001, 24, 218–223. [Google Scholar] [CrossRef] [Green Version]
- Shigiyama, F.; Kumashiro, N.; Tsuneoka, Y.; Igarashi, H.; Yoshikawa, F.; Kakehi, S.; Funato, H.; Hirose, T. Mechanisms of sleep deprivation-induced hepatic steatosis and insulin resistance in mice. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E848–E858. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Shioji, I.; Sugimoto, A.; Yamaoka, M. Psychological stress increases bilirubin metabolites in human urine. Biochem. Biophys. Res. Commun. 2002, 293, 517–520. [Google Scholar] [CrossRef]
- Rawat, V.; Bortolussi, G.; Gazzin, S.; Tiribelli, C.; Muro, A.F. Bilirubin-induced oxidative stress leads to DNA damage in the cerebellum of hyperbilirubinemic neonatal mice and activates DNA double-strand break repair pathways in human cells. Oxidative Med. Cell. Longev. 2018, 2018, 1801243. [Google Scholar] [CrossRef]
- Sumida, K.; Kawana, M.; Kouno, E.; Itoh, T.; Takano, S.; Narawa, T.; Tukey, R.H.; Fujiwara, R. Importance of UDP-glucuronosyltransferase 1A1 expression in skin and its induction by UVB in neonatal hyperbilirubinemia. Mol. Pharmacol. 2013, 84, 679–686. [Google Scholar] [CrossRef] [Green Version]
- Kumagai, A.; Ando, R.; Miyatake, H.; Greimel, P.; Kobayashi, T.; Hirabayashi, Y.; Shimogori, T.; Miyawaki, A. A bilirubin-inducible fluorescent protein from eel muscle. Cell 2013, 153, 1602–1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hengen, P. Purification of His-Tag fusion proteins from Escherichia coli. Trends Biochem. Sci. 1995, 20, 285–286. [Google Scholar] [CrossRef]
- Iwatani, S.; Nakamura, H.; Kurokawa, D.; Yamana, K.; Nishida, K.; Fukushima, S.; Koda, T.; Nishimura, N.; Nishio, H.; Iijima, K.; et al. Fluorescent protein-based detection of unconjugated bilirubin in newborn serum. Sci. Rep. 2016, 6, 28489. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Luo, W.; Friedman, M.S.; Shedden, K.; Hankenson, K.D.; Woolf, P.J. GAGE: Generally applicable gene set enrichment for pathway analysis. BMC Bioinform. 2009, 10, 161. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment—Time | Total BR Normalized to Estimated Tissue Volume (g/tissue-mL) | p-Value | |
|---|---|---|---|
| vs. Baseline | vs. Corresponding Control Group | ||
| Baseline | 1.44 ± 0.22 | - | - |
| No H2O2—24 h | 1.43 ± 0.32 | 0.970 | - |
| No H2O2—48 h | 1.75 ± 0.26 | 0.403 | - |
| No H2O2—72 h | 2.26 ± 0.27 | 0.056 | - |
| 30 µM H2O2—24 h | 2.80 ± 0.37 | 0.020 | 0.031 |
| 30 µM H2O2—48 h | 4.01 ± 0.79 | 0.021 | 0.035 |
| 30 µM H2O2—72 h | 3.93 ± 0.78 | 0.022 | 0.091 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, B.; Card, P.D.; Chen, J.; Li, L.; Laughlin, T.; Jarrold, B.; Zhao, W.; Benham, A.M.; Määttä, A.T.; Hawkins, T.J.; et al. A Potential Role of Keratinocyte-Derived Bilirubin in Human Skin Yellowness and Its Amelioration by Sucrose Laurate/Dilaurate. Int. J. Mol. Sci. 2022, 23, 5884. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23115884
Fang B, Card PD, Chen J, Li L, Laughlin T, Jarrold B, Zhao W, Benham AM, Määttä AT, Hawkins TJ, et al. A Potential Role of Keratinocyte-Derived Bilirubin in Human Skin Yellowness and Its Amelioration by Sucrose Laurate/Dilaurate. International Journal of Molecular Sciences. 2022; 23(11):5884. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23115884
Chicago/Turabian StyleFang, Bin, Patrick D. Card, Junjun Chen, Lijuan Li, Timothy Laughlin, Bradley Jarrold, Wenzhu Zhao, Adam M. Benham, Arto T. Määttä, Timothy J. Hawkins, and et al. 2022. "A Potential Role of Keratinocyte-Derived Bilirubin in Human Skin Yellowness and Its Amelioration by Sucrose Laurate/Dilaurate" International Journal of Molecular Sciences 23, no. 11: 5884. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23115884