
Regulatory and Enterotoxin Gene Expression and Enterotoxins Production in Staphylococcus aureus FRI913 Cultures Exposed to a Rotating Magnetic Field and trans-Anethole

 ,
,  , ,
, ,  , ,
, ,  , and
, and 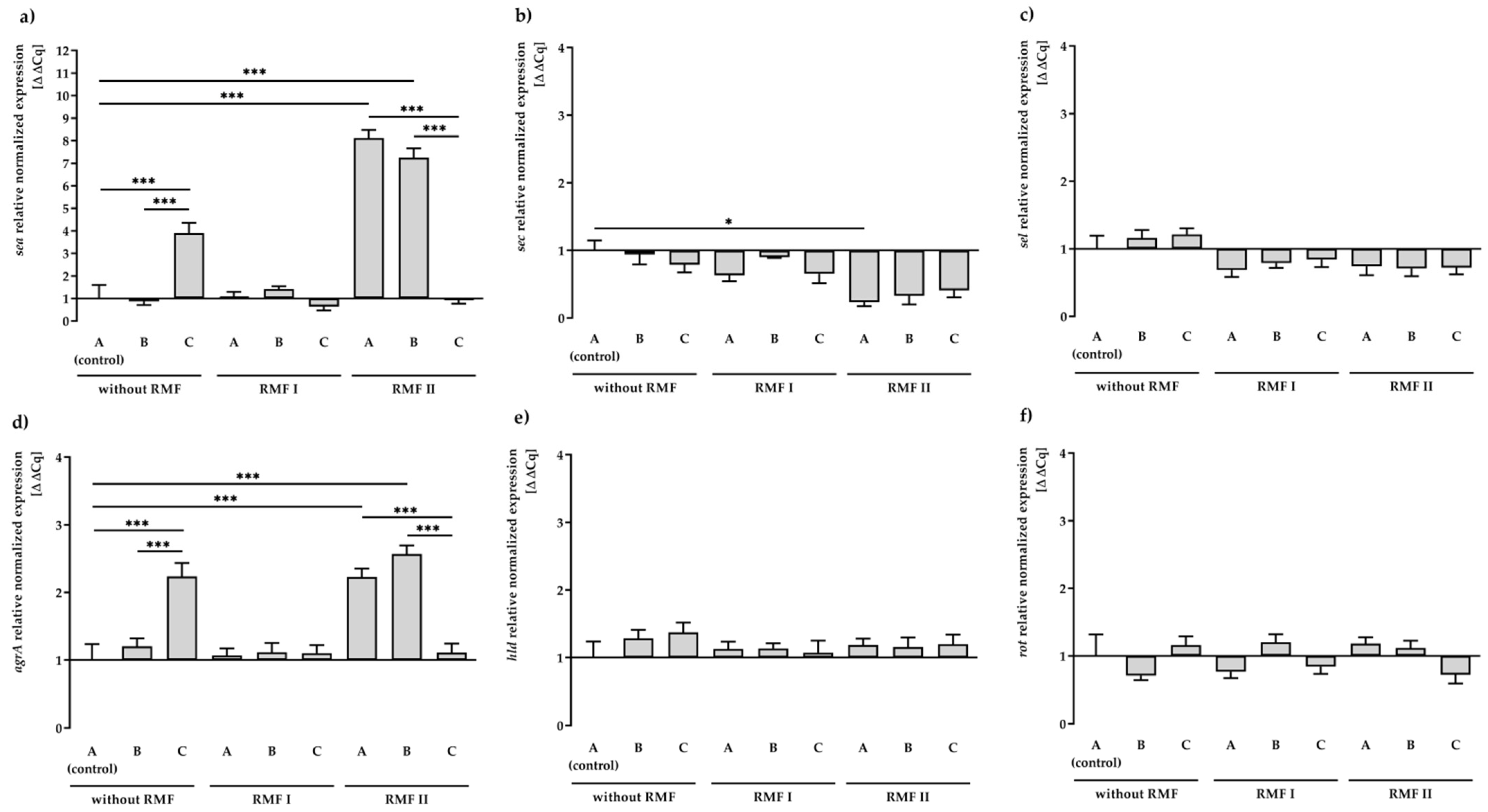{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Evaluation of Antibacterial Activity of TA
2.2. Selection of Housekeeping Gene
2.3. Effect of RMF and TA on SEs and Regulatory Gene Expression
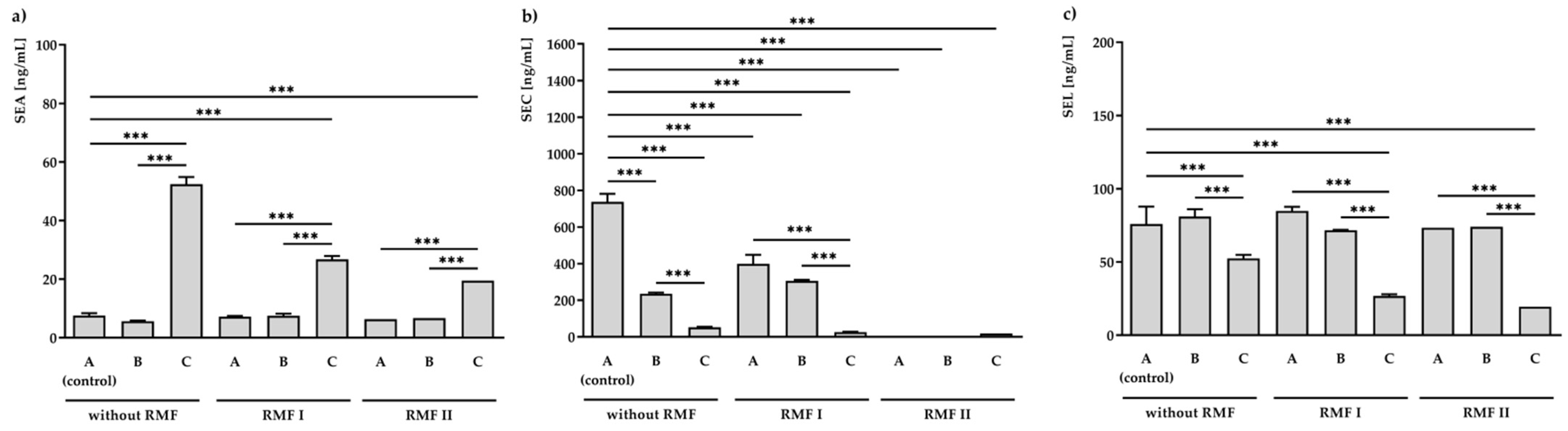
2.4. Effect of RMF and TA on SEs Production
3. Discussion
4. Materials and Methods
4.1. Reference Strain, Culture Condition, and Preparation of Staphylococcal Suspension
4.2. Minimum Inhibitory Concentration (MIC) Assay
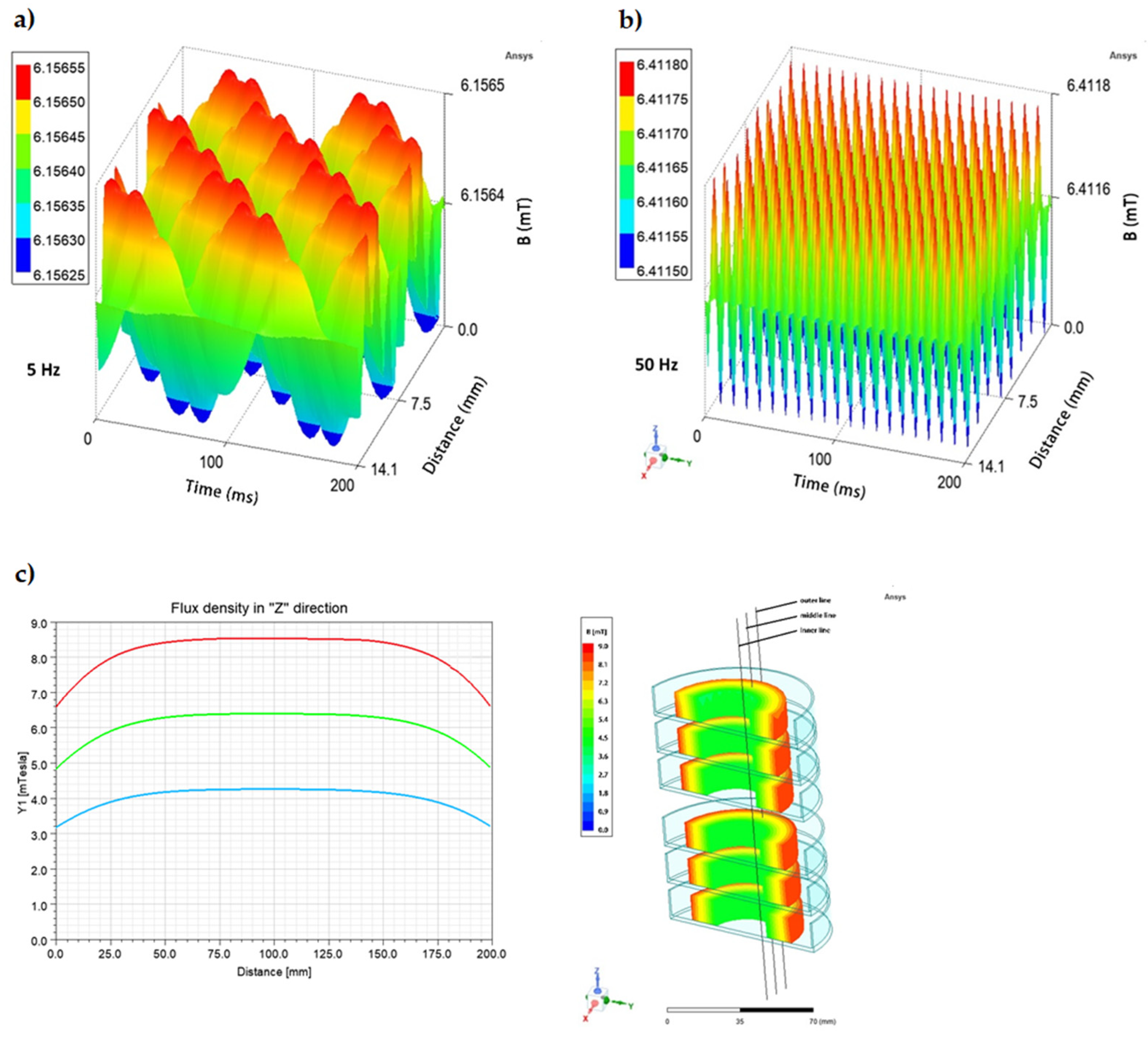
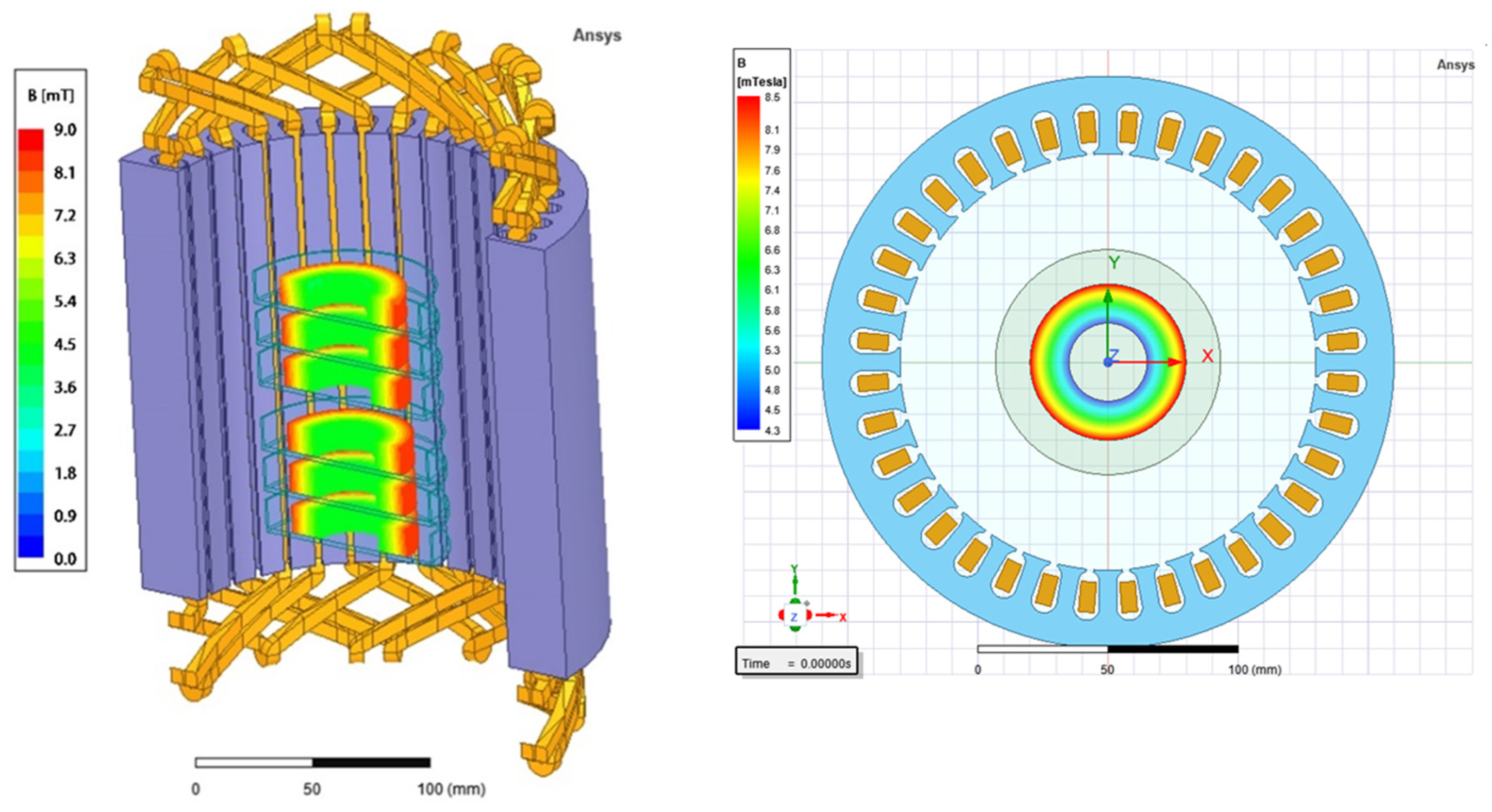
4.3. Rotating Magnetic Field Generator
4.4. Exposure of S. aureus FRI913 to RMF
4.5. RNA Isolation
4.6. Quantitative Real-Time PCR (qPCR)
4.7. Sandwich ELISA
4.7.1. Samples Preparation
4.7.2. Sandwich ELISA Preparation
4.8. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pollitt, E.J.G.; Szkuta, P.T.; Burns, N.; Foster, S.J. Staphylococcus aureus infection dynamics. PLoS Pathog. 2018, 14, e1007112. [Google Scholar] [CrossRef] [Green Version]
- Urish, K.L.; Cassat, J.E. Staphylococcus aureus osteomyelitis: Bone, bugs, and surgery. Infect. Immun. 2020, 88, e00932-19. [Google Scholar] [CrossRef]
- Cheung, G.Y.C.; Bae, J.S.; Otto, M. Pathogenicity and virulence of Staphylococcus aureus. Virulence 2021, 12, 547–569. [Google Scholar] [CrossRef]
- Adhikari, R.P.; Novick, R.P. Regulatory organization of the staphylococcal sae locus. Microbiology 2008, 154, 949–959. [Google Scholar] [CrossRef] [Green Version]
- Ono, H.K.; Sato’o, Y.; Narita, K.; Naito, I.; Hirose, S.; Hisatsune, J.; Asano, K.; Hu, D.-L.; Omoe, K.; Sugai, M.; et al. Identification and characterization of a novel staphylococcal emetic toxin. Appl. Environ. Microbiol. 2015, 81, 7034–7040. [Google Scholar] [CrossRef] [Green Version]
- Futagawa-Saito, K.; Suzuki, M.; Ohsawa, M.; Ohshima, S.; Sakurai, N.; Ba-Thein, W.; Fukuyasu, T. Identification and prevalence of an enterotoxin-related gene, se-int, in Staphylococcus intermedius isolates from dogs and pigeons. J. Appl. Microbiol. 2004, 96, 1361–1366. [Google Scholar] [CrossRef]
- Cerca, N.; Brooks, J.L.; Jefferson, K.K. Regulation of the intercellular adhesin locus regulator (icaR) by SarA, sigmaB, and IcaR in Staphylococcus aureus. J. Bacteriol. 2008, 190, 6530–6533. [Google Scholar] [CrossRef] [Green Version]
- Joo, H.-S.; Otto, M. Toxin-mediated gene regulatory mechanism in Staphylococcus aureus. Microb. Cell 2016, 4, 29–31. [Google Scholar] [CrossRef] [Green Version]
- Cheung, G.Y.C.; Wang, R.; Khan, B.A.; Sturdevant, D.E.; Otto, M. Role of the accessory gene regulator agr in community-associated methicillin-resistant Staphylococcus aureus pathogenesis. Infect. Immun. 2011, 79, 1927–1935. [Google Scholar] [CrossRef] [Green Version]
- Bronner, S.; Monteil, H.; Prévost, G. Regulation of virulence determinants in Staphylococcus aureus: Complexity and applications. FEMS Microbiol. Rev. 2004, 28, 183–200. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Zheng, L.; Landwehr, C.; Lunsford, D.; Holmes, D.; Ji, Y. Global regulation of gene expression by ArlRS, a two-component signal transduction regulatory system of Staphylococcus aureus. J. Bacteriol. 2005, 187, 5486–5492. [Google Scholar] [CrossRef] [Green Version]
- Yarwood, J.M.; McCormick, J.K.; Schlievert, P.M. Identification of a novel two-component regulatory system that acts in global regulation of virulence factors of Staphylococcus aureus. J. Bacteriol. 2001, 183, 1113–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, H.L.; Novick, R.P.; Kreiswirth, B.; Kornblum, J.; Schlievert, P. Cloning, characterization, and sequencing of an accessory gene regulator (agr) in Staphylococcus aureus. J. Bacteriol. 1988, 170, 4365–4372. [Google Scholar] [CrossRef] [Green Version]
- George, E.A.; Muir, T.W. Molecular mechanisms of agr quorum sensing in virulent staphylococci. Chembiochem 2007, 8, 847–855. [Google Scholar] [CrossRef]
- Benito, Y.; Kolb, F.A.; Romby, P.; Lina, G.; Etienne, J.; Vandenesch, F. Probing the structure of RNAIII, the Staphylococcus aureus agr regulatory RNA, and identification of the RNA domain involved in repression of protein A expression. RNA 2000, 6, 668–679. [Google Scholar] [CrossRef] [Green Version]
- Traber, K.E.; Lee, E.; Benson, S.; Corrigan, R.; Cantera, M.; Shopsin, B.; Novick, R.P. agr function in clinical Staphylococcus aureus isolates. Microbiology 2008, 154, 2265–2274. [Google Scholar] [CrossRef] [Green Version]
- Recsei, P.; Kreiswirth, B.; O’Reilly, M.; Schlievert, P.; Gruss, A.; Novick, R.P. Regulation of exoprotein gene expression in Staphylococcus aureus by agr. Mol. Gen. Genet. 1986, 202, 58–61. [Google Scholar] [CrossRef] [PubMed]
- McNamara, P.J.; Milligan-Monroe, K.C.; Khalili, S.; Proctor, R.A. Identification, cloning, and initial characterization of rot, a locus encoding a regulator of virulence factor expression in Staphylococcus aureus. J. Bacteriol. 2000, 182, 3197–3203. [Google Scholar] [CrossRef] [Green Version]
- Kwiatkowski, P.; Pruss, A.; Masiuk, H.; Mnichowska-Polanowska, M.; Kaczmarek, M.; Giedrys-Kalemba, S.; Dołęgowska, B.; Zielińska-Bliźniewska, H.; Olszewski, J.; Sienkiewicz, M. The effect of fennel essential oil and trans-anethole on antibacterial activity of mupirocin against Staphylococcus aureus isolated from asymptomatic carriers. Postep. Dermatol. I Alergol. 2019, 36, 308–314. [Google Scholar] [CrossRef]
- Asano, T.; Aida, S.; Suemasu, S.; Mizushima, T. Anethole restores delayed gastric emptying and impaired gastric accommodation in rodents. Biochem. Biophys. Res. Commun. 2016, 472, 125–130. [Google Scholar] [CrossRef] [Green Version]
- Aprotosoaie, A.C.; Costache, I.-I.; Miron, A. Anethole and its role in chronic diseases. Adv. Exp. Med. Biol. 2016, 929, 247–267. [Google Scholar] [CrossRef] [PubMed]
- Kwiatkowski, P.; Pruss, A.; Wojciuk, B.; Dołęgowska, B.; Wajs-Bonikowska, A.; Sienkiewicz, M.; Mężyńska, M.; Łopusiewicz, Ł. The influence of essential oil compounds on antibacterial activity of mupirocin-susceptible and induced low-level mupirocin-resistant MRSA strains. Molecules 2019, 24, 3105. [Google Scholar] [CrossRef] [Green Version]
- Kwiatkowski, P.; Grygorcewicz, B.; Pruss, A.; Wojciuk, B.; Dołęgowska, B.; Giedrys-Kalemba, S.; Sienkiewicz, M.; Wojciechowska-Koszko, I. The effect of subinhibitory concentrations of trans-anethole on antibacterial and antibiofilm activity of mupirocin against mupirocin-resistant Staphylococcus aureus strains. Microb. Drug Resist. 2019, 25, 1424–1429. [Google Scholar] [CrossRef]
- Moradi, J.; Abbasipour, F.; Zaringhalam, J.; Maleki, B.; Ziaee, N.; Khodadoustan, A.; Janahmadi, M. Anethole, a medicinal plant compound, decreases the production of pro-inflammatory TNF-α and IL-1β in a rat model of LPS-induced periodontitis. Iran. J. Pharm. Res. 2014, 13, 1319–1325. [Google Scholar]
- Rao, T.R.; Sonolikar, R.; Saheb, S. Influence of magnetic field on the performance of bubble columns and airlift bioreactor with submersed microorganisms. Chem. Eng. Sci. 1997, 52, 4155–4160. [Google Scholar] [CrossRef]
- Eichwald, C.; Walleczek, J. Magnetic field perturbations as a tool for controlling enzyme-regulated and oscillatory biochemical reactions. Biophys. Chem. 1998, 74, 209–224. [Google Scholar] [CrossRef]
- Al-Qodah, Z.; Lafi, W. Modeling of antibiotics production in magneto three-phase airlift fermenter. Biochem. Eng. J. 2001, 7, 7–16. [Google Scholar] [CrossRef]
- Jiang, D.-S.; Long, S.-Y.; Huang, J.; Xiao, H.-Y.; Zhou, J.-Y. Immobilization of Pycnoporus sanguineus laccase on magnetic chitosan microspheres. Biochem. Eng. J. 2005, 25, 15–23. [Google Scholar] [CrossRef]
- Krishnamurthy, S.; Yadav, A.; Phelan, P.E.; Calhoun, R.; Vuppu, A.K.; Garcia, A.A.; Hayes, M.A. Dynamics of rotating paramagnetic particle chains simulated by particle dynamics, Stokesian dynamics and lattice Boltzmann methods. Microfluid. Nanofluidics 2008, 5, 33–41. [Google Scholar] [CrossRef]
- Rakoczy, R.; Masiuk, S. Influence of transverse rotating magnetic field on enhancement of solid dissolution process. AIChE J. 2010, 56, 1416–1433. [Google Scholar] [CrossRef]
- Rakoczy, R.; Masiuk, S. Experimental study of bubble size distribution in a liquid column exposed to a rotating magnetic field. Chem. Eng. Process. 2009, 48, 1229–1240. [Google Scholar] [CrossRef]
- Masiuk, M.; Rakoczy, R.; Masiuk, S.; Kordas, M. The expression and intranuclear distribution of nucleolin in HL-60 and K-562 cells after repeated, short-term exposition to rotating magnetic fields. Int. J. Radiat. Biol. 2008, 84, 752–760. [Google Scholar] [CrossRef]
- Rakoczy, R.; Masiuk, S. Studies of a mixing process induced by a transverse rotating magnetic field. Chem. Eng. Sci. 2011, 66, 2298–2308. [Google Scholar] [CrossRef]
- Fijałkowski, K.; Nawrotek, P.; Struk, M.; Kordas, M.; Rakoczy, R. Effects of rotating magnetic field exposure on the functional parameters of different species of bacteria. Electromagn. Biol. Med. 2015, 34, 48–55. [Google Scholar] [CrossRef]
- Fijałkowski, K.; Peitler, D.; Żywicka, A.; Rakoczy, R. The effect of rotating magnetic field on enterotoxin genes expression in Staphylococcus aureus strains. J. Magn. 2016, 21, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Qiu, J.; Li, H.; Su, H.; Dong, J.; Luo, M.; Wang, J.; Leng, B.; Deng, Y.; Liu, J.; Deng, X. Chemical composition of fennel essential oil and its impact on Staphylococcus aureus exotoxin production. World J. Microbiol. Biotechnol. 2012, 28, 1399–1405. [Google Scholar] [CrossRef] [PubMed]
- Nawrotek, P.; Fijałkowski, K.; Struk, M.; Kordas, M.; Rakoczy, R. Effects of 50 Hz rotating magnetic field on the viability of Escherichia coli and Staphylococcus aureus. Electromagn. Biol. Med. 2014, 33, 29–34. [Google Scholar] [CrossRef]
- Junka, A.F.; Rakoczy, R.; Szymczyk, P.; Bartoszewicz, M.; Sedghizadeh, P.P.; Fijałkowski, K. Application of rotating magnetic fields increase the activity of antimicrobials against wound biofilm pathogens. Sci. Rep. 2018, 8, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woroszyło, M.; Ciecholewska-Juśko, D.; Junka, A.; Wardach, M.; Chodaczek, G.; Dudek, B.; Fijałkowski, K. The effect of rotating magnetic field on susceptibility profile of methicillin-resistant Staphylococcus aureus strains exposed to activity of different groups of antibiotics. Int. J. Mol. Sci. 2021, 22, 11551. [Google Scholar] [CrossRef]
- Woroszyło, M.; Ciecholewska-Juśko, D.; Junka, A.; Pruss, A.; Kwiatkowski, P.; Wardach, M.; Fijałkowski, K. The impact of intraspecies variability on growth rate and cellular metabolic activity of bacteria exposed to rotating magnetic field. Pathogens 2021, 10, 1427. [Google Scholar] [CrossRef]
- Fijałkowski, K.; Nawrotek, P.; Struk, M.; Kordas, M.; Rakoczy, R. The effects of rotating magnetic field on growth rate, cell metabolic activity and biofilm formation by Staphylococcus aureus and Escherichia coli. J. Magn. 2013, 18, 289–296. [Google Scholar] [CrossRef] [Green Version]
- Fojt, L.; Strasák, L.; Vetterl, V.; Smarda, J. Comparison of the low-frequency magnetic field effects on bacteria Escherichia coli, Leclercia adecarboxylata and Staphylococcus aureus. Bioelectrochemistry 2004, 63, 337–341. [Google Scholar] [CrossRef]
- Ahmed, I.; Istivan, T.; Cosic, I.; Pirogova, E. Evaluation of the effects of extremely low frequency (ELF) pulsed electromagnetic fields (PEMF) on survival of the bacterium Staphylococcus aureus. EPJ Nonlinear Biomed. Phys. 2013, 1, 5. [Google Scholar] [CrossRef] [Green Version]
- Dunca, S.I.; Creanga, D.E.; Ailiesei, O.; Nimitan, E. Microorganisms growth with magnetic fluids. J. Magn. Magn. Mater. 2005, 289, 445–447. [Google Scholar] [CrossRef]
- Del Re, B.; Bersani, F.; Agostini, C.; Mesirca, P.; Giorgi, G. Various effects on transposition activity and survival of Escherichia coli cells due to different ELF-MF signals. Radiat. Environ. Biophys. 2004, 43, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Del Re, B.; Garoia, F.; Mesirca, P.; Agostini, C.; Bersani, F.; Giorgi, G. Extremely low frequency magnetic fields affect transposition activity in Escherichia coli. Radiat. Environ. Biophys. 2003, 42, 113–118. [Google Scholar] [CrossRef]
- Cruz, A.R.; van Strijp, J.A.G.; Bagnoli, F.; Manetti, A.G.O. Virulence gene expression of Staphylococcus aureus in human skin. Front. Microbiol. 2021, 12, 692023. [Google Scholar] [CrossRef]
- Potenza, L.; Ubaldi, L.; De Sanctis, R.; De Bellis, R.; Cucchiarini, L.; Dachà, M. Effects of a static magnetic field on cell growth and gene expression in Escherichia coli. Mutat. Res. 2004, 561, 53–62. [Google Scholar] [CrossRef]
- Giorgi, G.; Marcantonio, P.; Bersani, F.; Gavoçi, E.; Del Re, B. Effect of extremely low frequency magnetic field exposure on DNA transposition in relation to frequency, wave shape and exposure time. Int. J. Radiat. Biol. 2011, 87, 601–608. [Google Scholar] [CrossRef]
- Azizkhani, M.; Misaghi, A.; Basti, A.A.; Gandomi, H.; Hosseini, H. Effects of Zataria multiflora Boiss. essential oil on growth and gene expression of enterotoxins A, C and E in Staphylococcus aureus ATCC 29213. Int. J. Food Microbiol. 2013, 163, 159–165. [Google Scholar] [CrossRef]
- Qiu, J.; Zhang, X.; Luo, M.; Li, H.; Dong, J.; Wang, J.; Leng, B.; Wang, X.; Feng, H.; Ren, W.; et al. Subinhibitory concentrations of perilla oil affect the expression of secreted virulence factor genes in Staphylococcus aureus. PLoS ONE 2011, 6, e16160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Dong, J.; Qiu, J.-Z.; Wang, J.-F.; Luo, M.-J.; Li, H.-E.; Leng, B.-F.; Ren, W.-Z.; Deng, X.-M. Peppermint oil decreases the production of virulence-associated exoproteins by Staphylococcus aureus. Molecules 2011, 16, 1642–1654. [Google Scholar] [CrossRef] [Green Version]
- Qiu, J.; Wang, D.; Xiang, H.; Feng, H.; Jiang, Y.; Xia, L.; Dong, J.; Lu, J.; Yu, L.; Deng, X. Subinhibitory concentrations of thymol reduce enterotoxins A and B and alpha-hemolysin production in Staphylococcus aureus isolates. PLoS ONE 2010, 5, e9736. [Google Scholar] [CrossRef] [Green Version]
- Roux, A.; Todd, D.A.; Velázquez, J.V.; Cech, N.B.; Sonenshein, A.L. CodY-mediated regulation of the Staphylococcus aureus Agr system integrates nutritional and population density signals. J. Bacteriol. 2014, 196, 1184–1196. [Google Scholar] [CrossRef] [Green Version]
- Dinges, M.M.; Orwin, P.M.; Schlievert, P.M. Exotoxins of Staphylococcus aureus. Clin. Microbiol. Rev. 2000, 13, 16–34, table of contents. [Google Scholar] [CrossRef]
- Banaszkiewicz, S.; Calland, J.K.; Mourkas, E.; Sheppard, S.K.; Pascoe, B.; Bania, J. Genetic diversity of composite enterotoxigenic Staphylococcus epidermidis pathogenicity islands. Genome Biol. Evol. 2019, 11, 3498–3509. [Google Scholar] [CrossRef]
- Cervera-Alamar, M.; Guzmán-Markevitch, K.; Žiemytė, M.; Ortí, L.; Bernabé-Quispe, P.; Pineda-Lucena, A.; Pemán, J.; Tormo-Mas, M.Á. Mobilisation mechanism of pathogenicity islands by endogenous phages in Staphylococcus aureus clinical strains. Sci. Rep. 2018, 8, 16742. [Google Scholar] [CrossRef]
- Strahl, H.; Errington, J. Bacterial membranes: Structure, domains, and function. Annu. Rev. Microbiol. 2017, 71, 519–538. [Google Scholar] [CrossRef] [PubMed]
- Dyachenko, E.N.; Dueck, J.G. Modeling of sedimentation and filtration layer formation by discrete element method. Comput. Res. Model. 2012, 4, 105–120. [Google Scholar] [CrossRef]
- Yildiz, E.; Dost, S.; Yildiz, M. A numerical simulation study for the effect of magnetic fields in liquid phase diffusion growth of SiGe single crystals. J. Cryst. Growth 2006, 291, 497–511. [Google Scholar] [CrossRef]
- de Groot, S.R.; Mazur, P. Non-Equlibrium Thermodynamics; North-holl. Publ. Co.: Amsterdam, The Netherlands, 1962. [Google Scholar]
- Stieglitz, R.; Barleon, L.; BüHler, L.; Molokov, S. Magnetohydrodynamic flow in a right-angle bend in a strong magnetic field. J. Fluid Mech. 1996, 326, 91–123. [Google Scholar] [CrossRef]
- Moreau, R. The fundamentals of MHD related to crystal growth. Prog. Cryst. Growth Charact. Mater. 1999, 38, 161–194. [Google Scholar] [CrossRef]
- Miñano, H.L.; Silva, A.C.; Souto, S.; Costa, E.J. Magnetic fields in food processing perspectives, applications and action models. Processes 2020, 8, 814. [Google Scholar] [CrossRef]
- Barz, R.U.; Gerbeth, G.; Wunderwald, U.; Buhrig, E.; Gelfgat, Y.M. Modelling of the isothermal melt flow due to rotating magnetic fields in crystal growth. J. Cryst. Growth 1997, 180, 410–421. [Google Scholar] [CrossRef]
- Hadid, H.B.; Vaux, S.; Keddeche, S. Three-dimensional flow transitions under a rotating magnetic field. J. Cryst. Growth 2001, 230, 57–62. [Google Scholar] [CrossRef]
- Krause, F. Ferrohydrodynamics. J. Appl. Math. Mech. 1987, 67, 279. [Google Scholar] [CrossRef]
- Cramer, A.; Pal, J.; Gerbeth, G. Turbulence measurements in a rotating magnetic field driven flow. Phys. Fluids 2012, 24, 45105. [Google Scholar] [CrossRef]
- Stiller, J.; Fraňa, K. Transitional and weakly turbulent flow in a rotating magnetic field. Phys. Fluids 2006, 18, 074105. [Google Scholar] [CrossRef]
- Hong, R.Y.; Feng, B.; Chen, L.L.; Liu, G.H.; Li, H.Z.; Zheng, Y.; Wei, D.G. Synthesis, characterization and MRI application of dextran-coated Fe3O4 magnetic nanoparticles. Biochem. Eng. J. 2008, 42, 290–300. [Google Scholar] [CrossRef]
- Ma, Z.; Liu, H. Synthesis and surface modification of magnetic particles for application in biotechnology and biomedicine. China Particuology 2007, 5, 1–10. [Google Scholar] [CrossRef]
- Patwa, R.; Zandraa, O.; Capáková, Z.; Saha, N.; Sáha, P. Effect of iron-oxide nanoparticles impregnated bacterial cellulose on overall properties of alginate/casein hydrogels: Potential injectable biomaterial for wound healing applications. Polymers 2020, 12, 2690. [Google Scholar] [CrossRef]
- Xu, Z.P.; Zeng, Q.H.; Lu, G.Q.; Yu, A.B. Inorganic nanoparticles as carriers for efficient cellular delivery. Chem. Eng. Sci. 2006, 61, 1027–1040. [Google Scholar] [CrossRef]
- Vékás, L.; Bica, D.; Avdeev, M. V Magnetic nanoparticles and concentrated magnetic nanofluids: Synthesis, properties and some applications. China Particuology 2007, 5, 43–49. [Google Scholar] [CrossRef]
- Vincenti, B.; Ramos, G.; Cordero, M.L.; Douarche, C.; Soto, R.; Clement, E. Magnetotactic bacteria in a droplet self-assemble into a rotary motor. Nat. Commun. 2019, 10, 5082. [Google Scholar] [CrossRef] [Green Version]
- Ērglis, K.; Wen, Q.; Ose, V.; Zeltins, A.; Sharipo, A.; Janmey, P.A.; Cēbers, A. Dynamics of magnetotactic bacteria in a rotating magnetic field. Biophys. J. 2007, 93, 1402–1412. [Google Scholar] [CrossRef] [Green Version]
- Ghaisari, S.; Winklhofer, M.; Strauch, P.; Klumpp, S.; Faivre, D. Magnetosome organization in magnetotactic bacteria unraveled by ferromagnetic resonance spectroscopy. Biophys. J. 2017, 113, 637–644. [Google Scholar] [CrossRef] [Green Version]
- Hanzlik, M.; Winklhofer, M.; Petersen, N. Pulsed-field-remanence measurements on indivudal magnetotactic bacteria. J. Magn. Magn. Mater. 2002, 248, 258–267. [Google Scholar] [CrossRef]
- Safarik, I.; Safarikova, M. Magnetically modified microbial cells: A new type of magnetic adsorbents. China Particuology 2007, 5, 19–25. [Google Scholar] [CrossRef]
- Safaríková, M.; Ptácková, L.; Kibriková, I.; Safarík, I. Biosorption of water-soluble dyes on magnetically modified Saccharomyces cerevisiae subsp. uvarum cells. Chemosphere 2005, 59, 831–835. [Google Scholar] [CrossRef]
- Lu, X.; Li, H. Fluidization of CaCO3 and Fe2O3 particle mixtures in a transverse rotating magnetic field. Powder Technol. 2000, 107, 66–78. [Google Scholar] [CrossRef]
- Auer, G.K.; Weibel, D.B. Bacterial cell mechanics. Biochemistry 2017, 56, 3710–3724. [Google Scholar] [CrossRef] [PubMed]
- Amir, A.; Babaeipour, F.; McIntosh, D.B.; Nelson, D.R.; Jun, S. Bending forces plastically deform growing bacterial cell walls. Proc. Natl. Acad. Sci. USA 2014, 111, 5778–5783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podkowik, M.; Seo, K.S.; Schubert, J.; Tolo, I.; Robinson, D.A.; Bania, J.; Bystroń, J. Genotype and enterotoxigenicity of Staphylococcus epidermidis isolate from ready to eat meat products. Int. J. Food Microbiol. 2016, 229, 52–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CLSI. Clinial and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing. Twenty-Second Informational Supplement; M100-S22; CLSI: Wayne, PA, USA, 2012; Volume 32. [Google Scholar]
- Kwiatkowski, P.; Wojciuk, B.; Wojciechowska-Koszko, I.; Łopusiewicz, Ł.; Grygorcewicz, B.; Pruss, A.; Sienkiewicz, M.; Fijałkowski, K.; Kowalczyk, E.; Dołęgowska, B. Innate immune response against Staphylococcus aureus preincubated with subinhibitory concentration of trans-anethole. Int. J. Mol. Sci. 2020, 21, 4178. [Google Scholar] [CrossRef]
- Sihto, H.-M.; Tasara, T.; Stephan, R.; Johler, S. Validation of reference genes for normalization of qPCR mRNA expression levels in Staphylococcus aureus exposed to osmotic and lactic acid stress conditions encountered during food production and preservation. FEMS Microbiol. Lett. 2014, 356, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Lis, E.; Podkowik, M.; Schubert, J.; Bystroń, J.; Stefaniak, T.; Bania, J. Production of staphylococcal enterotoxin R by Staphylococcus aureus strains. Foodborne Pathog. Dis. 2012, 9, 762–766. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Fox, L.K.; Seo, K.S.; McGuire, M.A.; Park, Y.H.; Rurangirwa, F.R.; Sischo, W.M.; Bohach, G.A. Detection of classical and newly described staphylococcal superantigen genes in coagulase-negative staphylococci isolated from bovine intramammary infections. Vet. Microbiol. 2011, 147, 149–154. [Google Scholar] [CrossRef] [Green Version]
- Schubert, J.; Podkowik, M.; Bystroń, J.; Bania, J. Production of staphylococcal enterotoxins D and R in milk and meat juice by Staphylococcus aureus strains. Foodborne Pathog. Dis. 2017, 14, 223–230. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwiatkowski, P.; Tabiś, A.; Fijałkowski, K.; Masiuk, H.; Łopusiewicz, Ł.; Pruss, A.; Sienkiewicz, M.; Wardach, M.; Kurzawski, M.; Guenther, S.; et al. Regulatory and Enterotoxin Gene Expression and Enterotoxins Production in Staphylococcus aureus FRI913 Cultures Exposed to a Rotating Magnetic Field and trans-Anethole. Int. J. Mol. Sci. 2022, 23, 6327. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23116327
Kwiatkowski P, Tabiś A, Fijałkowski K, Masiuk H, Łopusiewicz Ł, Pruss A, Sienkiewicz M, Wardach M, Kurzawski M, Guenther S, et al. Regulatory and Enterotoxin Gene Expression and Enterotoxins Production in Staphylococcus aureus FRI913 Cultures Exposed to a Rotating Magnetic Field and trans-Anethole. International Journal of Molecular Sciences. 2022; 23(11):6327. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23116327
Chicago/Turabian StyleKwiatkowski, Paweł, Aleksandra Tabiś, Karol Fijałkowski, Helena Masiuk, Łukasz Łopusiewicz, Agata Pruss, Monika Sienkiewicz, Marcin Wardach, Mateusz Kurzawski, Sebastian Guenther, and et al. 2022. "Regulatory and Enterotoxin Gene Expression and Enterotoxins Production in Staphylococcus aureus FRI913 Cultures Exposed to a Rotating Magnetic Field and trans-Anethole" International Journal of Molecular Sciences 23, no. 11: 6327. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23116327