
A Comprehensive Identification and Expression Analysis of VQ Motif-Containing Proteins in Sugarcane (Saccharum spontaneum L.) under Phytohormone Treatment and Cold Stress

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Identification and Sequence Analysis of SsVQ Genes
2.2. Phylogenetic Analysis and Multiple Sequence Alignment
2.3. Conserved Motifs and Gene Structures of the VQ Gene Family
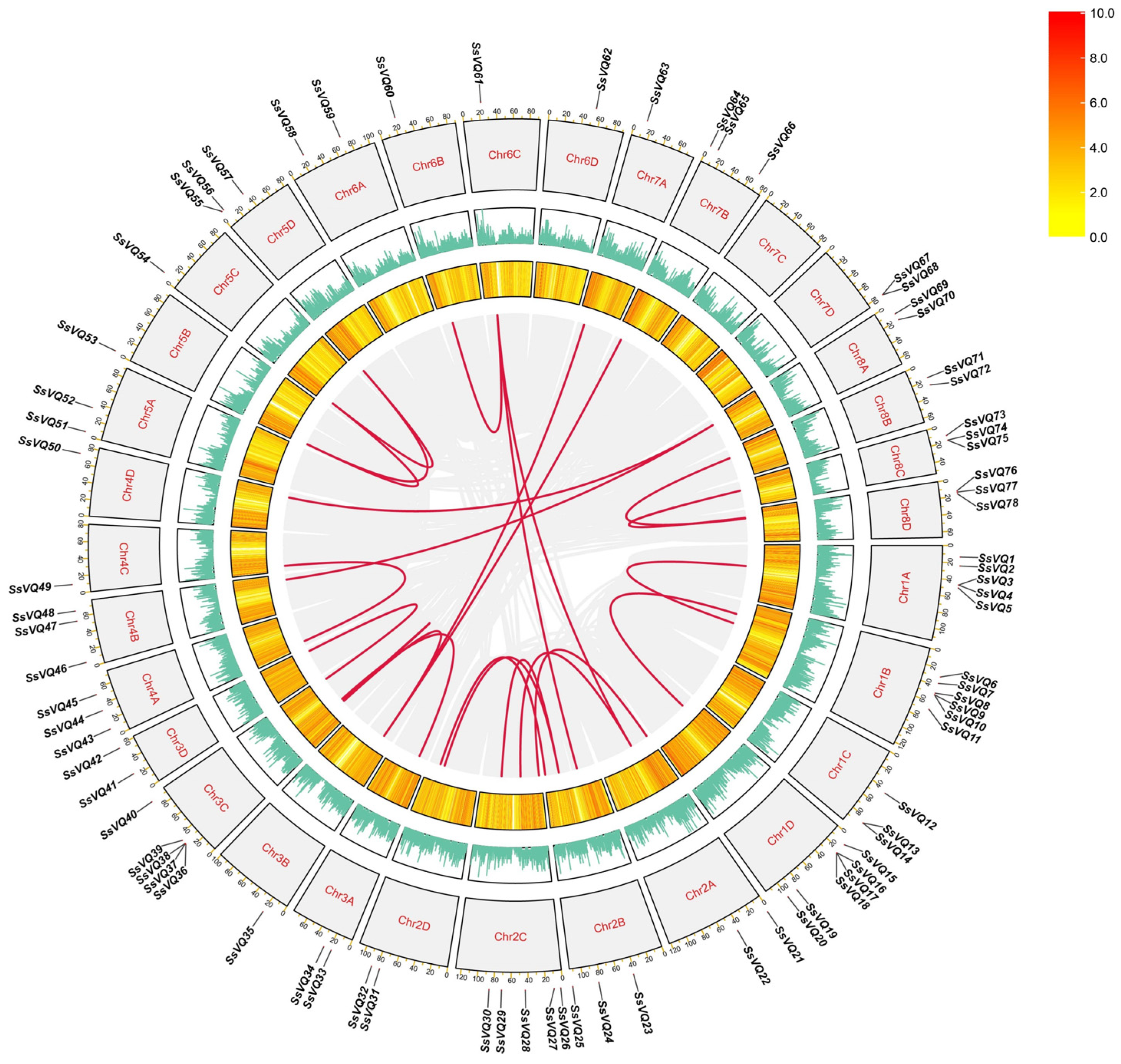
2.4. Chromosome Distribution and Gene Duplication Analysis
2.5. Identification of CREs in the Promoter Regions of SsVQ Genes
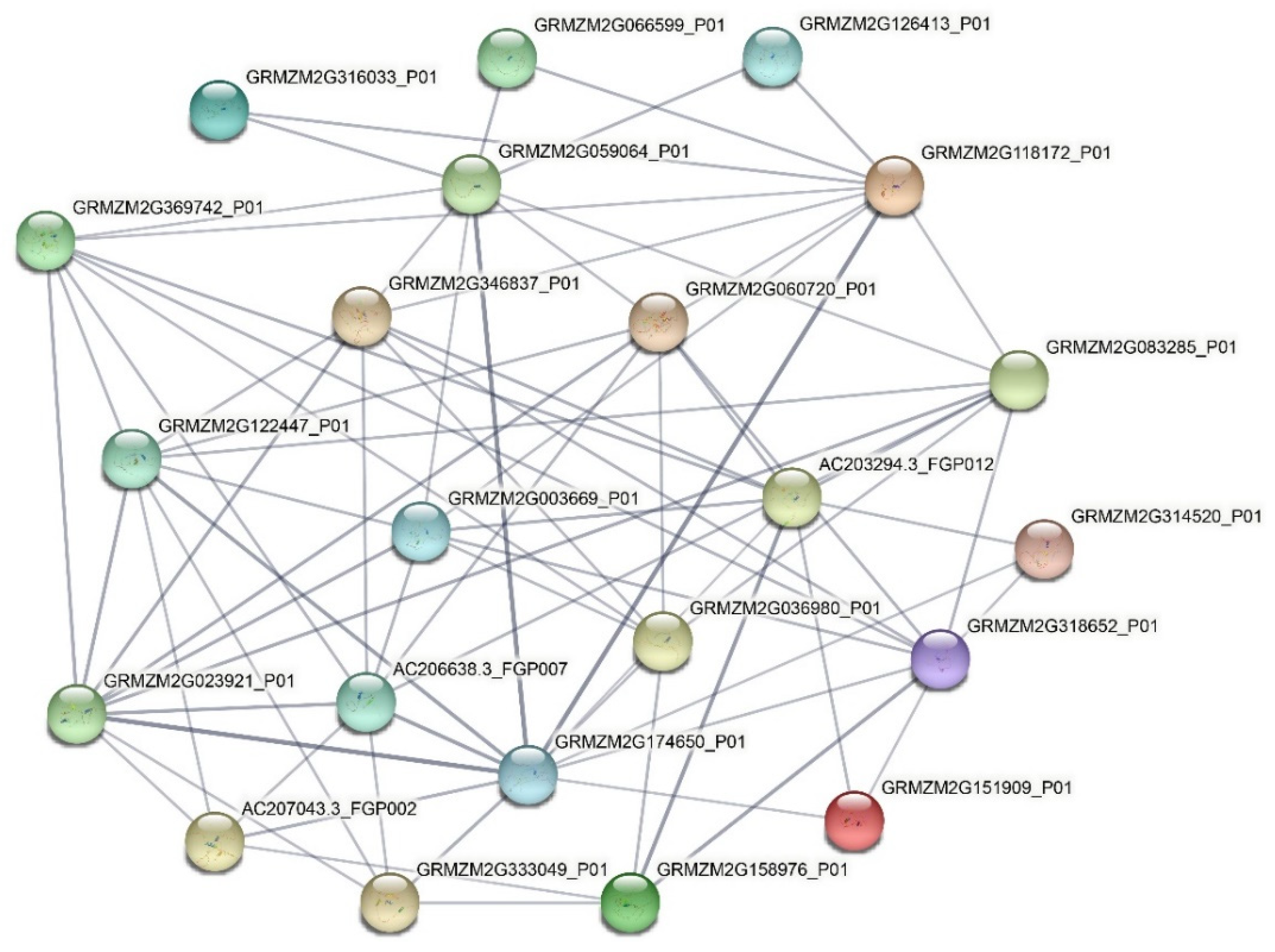
2.6. Protein-Protein Network of the VQ Proteins
2.7. Expression Pattern for SsVQ Genes during Sugarcane Development Based on RNA-Seq Data
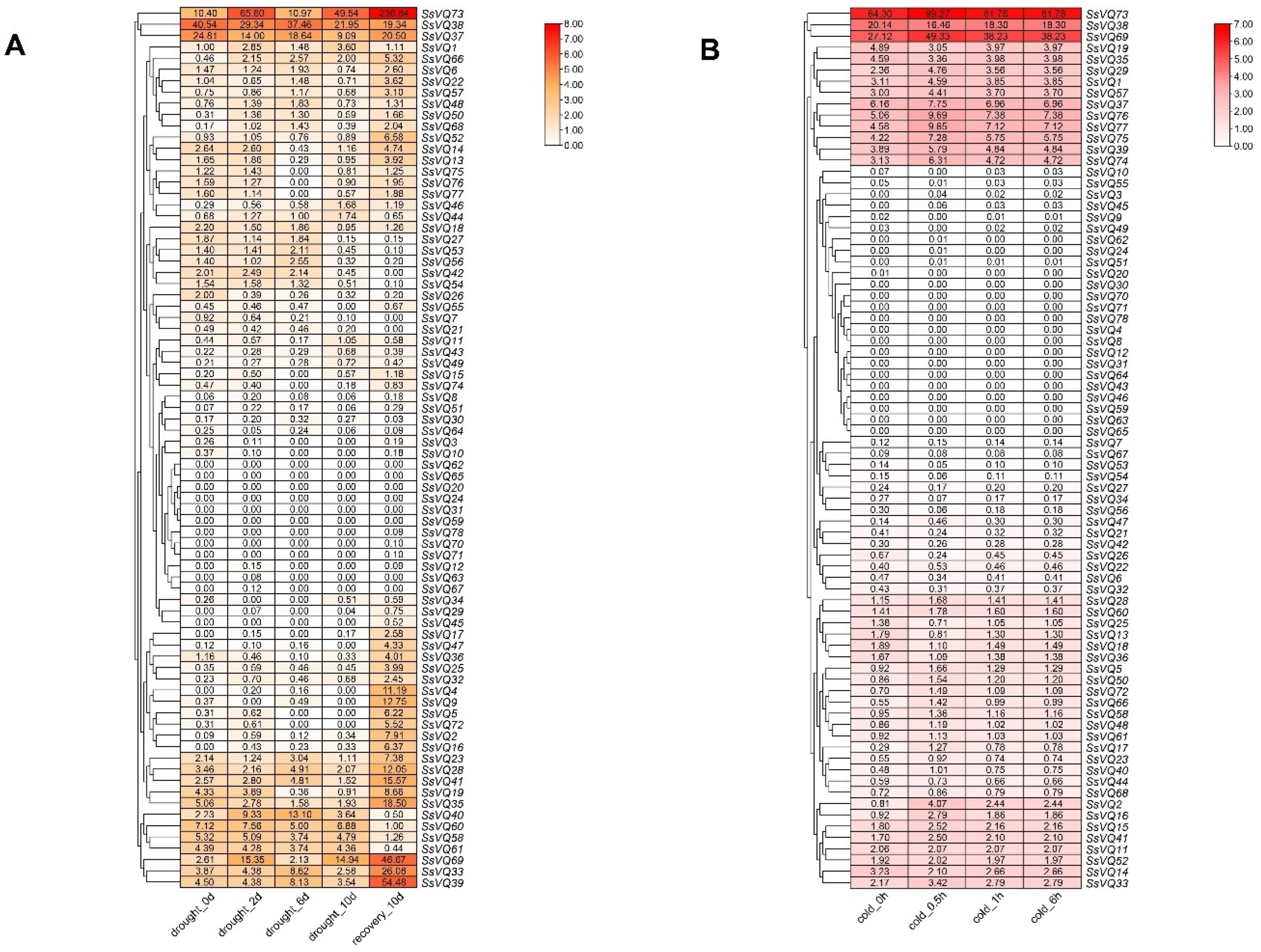
2.8. Expression Pattern for SsVQ Genes under Cold and Drought Treatments Based on SRA Data
2.9. Expression Analyses of VQ genes in Response to Hormone Treatments Using qRT-PCR
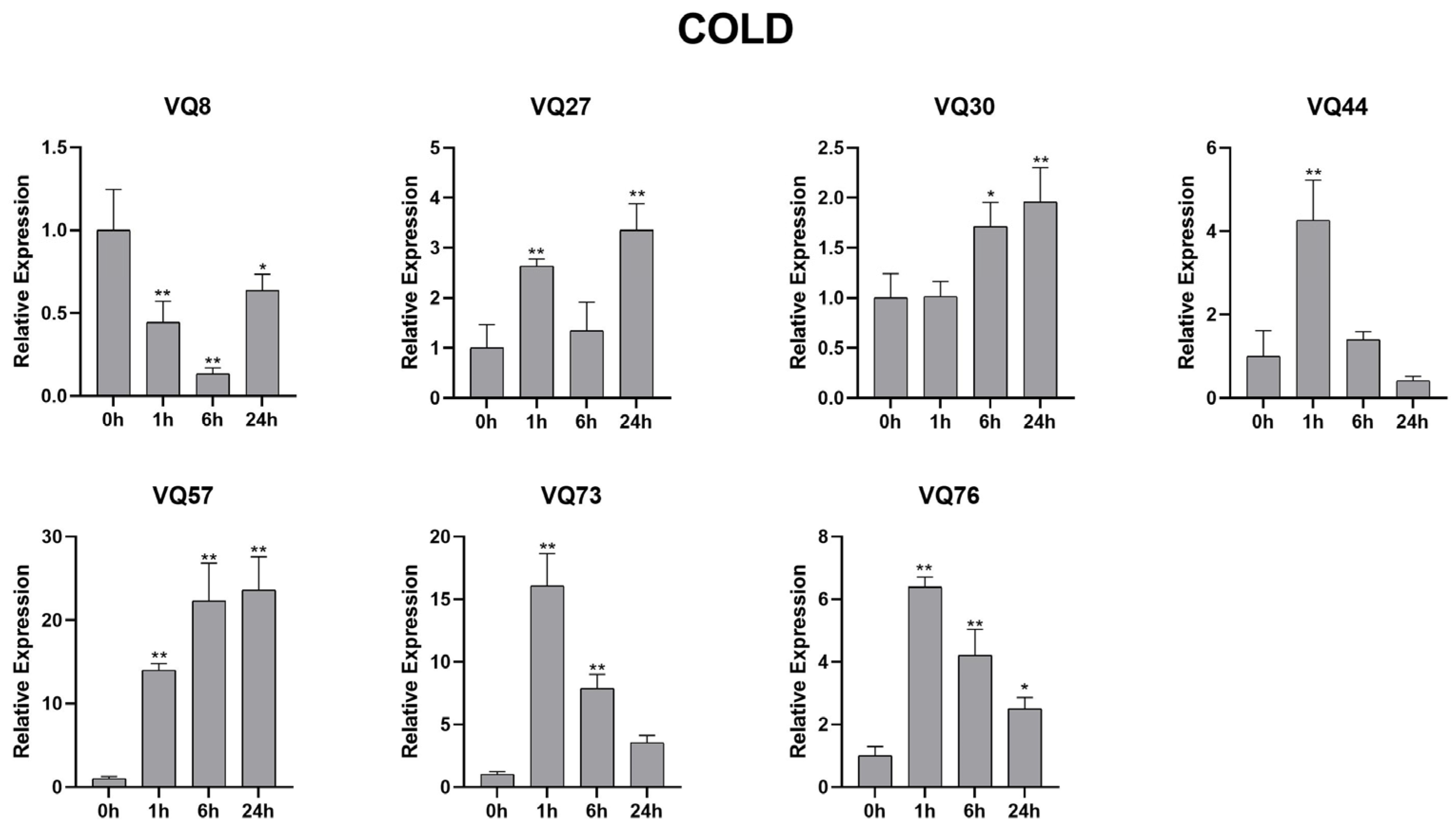
2.10. SsVQ Gene Expression following Abiotic Treatments Using qRT-PCR
3. Discussion
4. Materials and Methods
4.1. Identification and Classification of VQ Gene Family in Sugarcane
4.2. Phylogenetic Analysis and Multiple Sequence Alignment of SsVQ Proteins
4.3. Gene Structure, Conserved Domains and Motif Composition of SsVQ Genes
4.4. Promoter Analysis of SsVQ Genes
4.5. Chromosomal Distribution and Duplication Analysis of SsVQ Genes
4.6. Prediction of the Protein-Protein Interaction Network
4.7. Expression Analysis of SsVQs Based on RNA-Seq
4.8. Plant Materials, Phytohormone and Cold Treatments
4.9. Quantitative Real-Time PCR (qRT-PCR) Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhou, D.; Liu, X.; Gao, S.; Guo, J.; Su, Y.; Ling, H.; Wang, C.; Li, Z.; Xu, L.; Que, Y. Foreign cry1Ac gene integration and endogenous borer stress-related genes synergistically improve insect resistance in sugarcane. BMC Plant Biol. 2018, 18, 342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.; He, L.; Gao, S.; Su, Y.; Li, F.; Xu, L. Comparative analysis of two sugarcane ancestors Saccharum officinarum and S. spontaneum based on complete chloroplast genome sequences and photosynthetic ability in cold stress. Int. J. Mol. Sci. 2019, 20, 3828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Xu, L.; Que, Y.; Ling, H.; Yin, Z.; Liu, Y.; Zhou, D. The complete mitochondrial genome and phylogenetic analysis of sugarcane (Saccharum spp. hybrids) line 15a-53. Mitochondrial DNA B Resour. 2020, 5, 3389–3391. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Hua, X.T.; Zhong, W.M.; Yuan, Y.; Wang, Y.J.; Wang, Z.C.; Ming, R.; Zhang, J.S. Genome-wide identification and expression profile analysis of WRKY family genes in the autopolyploid Saccharum spontaneum. Plant Cell Physiol. 2020, 61, 616–630. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, X.; Tang, H.; Zhang, Q.; Hua, X.; Ma, X.; Zhu, F.; Jones, T.; Zhu, X.; Bowers, J.; et al. Allele-defined genome of the autopolyploid sugarcane Saccharum spontaneum L. Nat. Genet. 2018, 50, 1565–1573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lembke, C.G.; Nishiyama, M.Y., Jr.; Sato, P.M.; de Andrade, R.F.; Souza, G.M. Identification of sense and antisense transcripts regulated by drought in sugarcane. Plant Mol. Biol. 2012, 79, 461–477. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Song, X.P.; Chen, R.F.; Zhang, B.Q.; Li, C.N.; Liang, Y.S.; Qiu, L.H.; Fan, Y.G.; Zhou, Z.F.; Zhou, H.W.; et al. Genome-wide analysis of the DREB subfamily in Saccharum spontaneum reveals their functional divergence during cold and drought stresses. Front. Genet. 2020, 10, 1326. [Google Scholar] [CrossRef] [Green Version]
- Rice, J.L.; Hoy, J.W.; Grisham, M.P. Sugarcane mosaic distribution, incidence, increase, and spatial pattern in Louisiana. Plant Dis. 2019, 103, 2051–2056. [Google Scholar] [CrossRef]
- Dessoky, E.S.; Ismail, R.M.; Elarabi, N.I.; Abdelhadi, A.A.; Abdallah, N.A. Improvement of sugarcane for borer resistance using Agrobacterium mediated transformation of cry1Ac gene. GM Crops Food 2021, 12, 47–56. [Google Scholar] [CrossRef]
- Basnayake, J.; Jackson, P.A.; Inman-Bamber, N.G.; Lakshmanan, P. Sugarcane for water-limited environments. Genetic variation in cane yield and sugar content in response to water stress. J. Exp. Bot. 2012, 63, 6023–6033. [Google Scholar] [CrossRef]
- Viswanathan, R.; Balamuralikrishnan, M. Impact of mosaic infection on growth and yield of sugarcane. Sugar Tech 2005, 7, 61–65. [Google Scholar] [CrossRef]
- Lu, G.; Wang, Z.; Xu, F.; Pan, Y.-B.; Grisham, M.P.; Xu, L. Sugarcane mosaic disease: Characteristics, identification and control. Microorganisms 2021, 9, 1984. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.Z.; Yang, B.P.; Feng, X.Y.; Cao, Z.Y.; Feng, C.L.; Wang, J.G.; Xiong, G.R.; Shen, L.B.; Zeng, J.; Zhao, T.T.; et al. Development and characterization of transgenic sugarcane with insect resistance and herbicide tolerance. Front. Plant Sci. 2017, 8, 1535. [Google Scholar] [CrossRef] [PubMed]
- Qamar, Z.; Nasir, I.A.; Abouhaidar, M.G.; Hefferon, K.L.; Rao, A.Q.; Latif, A.; Ali, Q.; Anwar, S.; Rashid, B.; Shahid, A.A. Novel approaches to circumvent the devastating effects of pests on sugarcane. Sci. Rep. 2021, 11, 12428. [Google Scholar] [CrossRef]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, H.; Zhang, M.; Liu, Y.; He, Q.; Chai, M.; Liu, L.; Chen, F.; Huang, Y.; Yan, M.; Zhao, H.; et al. Genome-wide classification and evolutionary and functional analyses of the VQ family. Trop. Plant Biol. 2019, 12, 117–131. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, Z.; Li, Z.; Zhao, Y.; Tan, W.; Liu, Z.; Cui, S.; Yu, X.; Ma, J.; Wang, G.; et al. Genome-wide identification and expression analysis of the VQ gene family in soybean (Glycine max). PeerJ 2019, 7, e7509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, Z.; Li, Y.; Wang, F.; Cheng, Y.; Fan, B.; Yu, J.Q.; Chen, Z. Arabidopsis sigma factor binding proteins are activators of the WRKY33 transcription factor in plant defense. Plant Cell 2011, 23, 3824–3841. [Google Scholar] [CrossRef] [Green Version]
- Song, W.; Zhao, H.; Zhang, X.; Lei, L.; Lai, J. Genome-wide identification of VQ motif-containing proteins and their expression profiles under abiotic stresses in maize. Front. Plant Sci. 2015, 6, 1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Y.J.; Xiao, Y.Y.; Han, Y.C.; Shan, W.; Fan, Z.Q.; Xu, Q.G.; Kuang, J.F.; Lu, W.J.; Lakshmanan, P.; Chen, J.Y. Banana fruit VQ motif-containing protein5 represses cold-responsive transcription factor MaWRKY26 involved in the regulation of JA biosynthetic genes. Sci. Rep. 2016, 6, 23632. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Wang, F.; Li, J.; Ding, Q.; Zhang, Y.; Li, H.; Zhang, J.; Gao, J. Genome-wide identification and analysis of the VQ motif-containing protein family in Chinese cabbage (Brassica rapa L. ssp. Pekinensis). Int. J. Mol. Sci. 2015, 16, 28683–28704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Liu, H.; Zhou, C.; Timko, M.P. Genome-wide identification of the VQ protein gene family of tobacco (Nicotiana tabacum L.) and analysis of its expression in response to phytohormones and abiotic and biotic stresses. Genes 2020, 11, 284. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Wang, Y.; Hu, B.; Qiu, Z.; Zeng, B.; Fan, C. Genome-wide characterization, evolution, and expression profiling of VQ gene family in response to phytohormone treatments and abiotic stress in Eucalyptus grandis. Int. J. Mol. Sci. 2019, 20, 1765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.; Wei, F.; Cheng, S.; Ma, L.; Wang, H.; Zhang, M.; Mao, G.; Lu, J.; Hao, P.; Ahmad, A.; et al. A comprehensive analysis of cotton VQ gene superfamily reveals their potential and extensive roles in regulating cotton abiotic stress. BMC Genom. 2020, 21, 795. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Zhou, Y.; Yang, Y.; Chi, Y.J.; Zhou, J.; Chen, J.Y.; Wang, F.; Fan, B.; Shi, K.; Zhou, Y.H.; et al. Structural and functional analysis of VQ motif-containing proteins in Arabidopsis as interacting proteins of WRKY transcription factors. Plant Physiol. 2012, 159, 810–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, Y.; Lin, R. The VQ motif-containing protein family of plant-specific transcriptional regulators. Plant Physiol. 2015, 169, 371–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Q.; Zhao, S.; Duan, D.; Tian, Y.; Wang, Y.; Mao, K.; Zhou, Z.; Ma, F. Structural and functional analyses of genes encoding VQ proteins in apple. Plant Sci. 2018, 272, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Kwon, S.I.; Choi, C.; Lee, H.; Ahn, I.; Park, S.R.; Bae, S.C.; Lee, S.C.; Hwang, D.J. Expression analysis of rice VQ genes in response to biotic and abiotic stresses. Gene 2013, 529, 208–214. [Google Scholar] [CrossRef]
- Jiang, S.Y.; Sevugan, M.; Ramachandran, S. Valine-glutamine (VQ) motif coding genes are ancient and non-plant-specific with comprehensive expression regulation by various biotic and abiotic stresses. BMC Genom. 2018, 19, 342. [Google Scholar] [CrossRef]
- Wang, A.; Garcia, D.; Zhang, H.; Feng, K.; Chaudhury, A.; Berger, F.; Peacock, W.J.; Dennis, E.S.; Luo, M. The VQ motif protein IKU1 regulates endosperm growth and seed size in Arabidopsis. Plant J. 2010, 63, 670–679. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Zhou, W.; Cheng, Z.; Fan, M.; Wang, L.; Xie, D. JAV1 controls jasmonate-regulated plant defense. Mol. Cell 2013, 50, 504–515. [Google Scholar] [CrossRef] [Green Version]
- Perruc, E.; Charpenteau, M.; Ramirez, B.C.; Jauneau, A.; Galaud, J.P.; Ranjeva, R.; Ranty, B. A novel calmodulin-binding protein functions as a negative regulator of osmotic stress tolerance in Arabidopsis thaliana seedlings. Plant J. 2004, 38, 410–420. [Google Scholar] [CrossRef] [PubMed]
- Xixu, P.; Ting, X.; Jiao, M.; Zong, T.; Dinggang, Z.; Xinke, T.; Haihua, W. Differential expression of rice valine-qlutamine gene family in response to nitric oxide and regulatory circuit of OsVQ7 and OsWRKY24. Rice Sci. 2020, 27, 10–20. [Google Scholar] [CrossRef]
- Kaltenegger, E.; Leng, S.; Heyl, A. The effects of repeated whole genome duplication events on the evolution of cytokinin signaling pathway. BMC Evol. Biol. 2018, 18, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Yin, Z.; Liu, Y.; Li, Z.; Yan, M.; Que, Y.; Xu, L.; Zhou, D. The complete mitochondrial genome of sugarcane (Saccharum spp.) variety FN15. Mitochondrial DNA Part B 2020, 5, 2163–2165. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Ni, P.; Wong, G.K. Comparing the whole-genome-shotgun and map-based sequences of the rice genome. Trends Plant Sci. 2006, 11, 387–391. [Google Scholar] [CrossRef]
- Arabidopsis Genome, I. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 2000, 408, 796–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.; Liu, H.; Wu, L.; Warburton, M.; Yan, J. Genome-wide association studies in maize: Praise and Stargaze. Mol. Plant 2017, 10, 359–374. [Google Scholar] [CrossRef] [Green Version]
- Garsmeur, O.; Droc, G.; Antonise, R.; Grimwood, J.; Potier, B.; Aitken, K.; Jenkins, J.; Martin, G.; Charron, C.; Hervouet, C.; et al. A mosaic monoploid reference sequence for the highly complex genome of sugarcane. Nat. Commun. 2018, 9, 2638. [Google Scholar] [CrossRef] [PubMed]
- Ling, L.; Qu, Y.; Zhu, J.; Wang, D.; Guo, C. Genome-wide identification and expression analysis of the VQ gene family in Cicer arietinum and Medicago truncatula. PeerJ 2020, 8, e8471. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Vannozzi, A.; Wang, G.; Zhong, Y.; Corso, M.; Cavallini, E.; Cheng, Z.M. A comprehensive survey of the grapevine VQ gene family and its transcriptional correlation with WRKY proteins. Front. Plant Sci. 2015, 6, 417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, H.; Yuan, G.; Mo, S.; Qian, Y.; Wu, Y.; Chen, Q.; Xu, X.; Wu, X.; Ge, C. Genome-wide analysis of the plant-specific VQ motif-containing proteins in tomato (Solanum lycopersicum) and characterization of SlVQ6 in thermotolerance. Plant Physiol. Biochem. 2019, 143, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J. Evolution by gene duplication: An update. Trends Ecol. Evol. 2003, 18, 292–298. [Google Scholar] [CrossRef] [Green Version]
- Krishnamurthy, P.; Hong, J.K.; Kim, J.A.; Jeong, M.J.; Lee, Y.H.; Lee, S.I. Genome-wide analysis of the expansin gene superfamily reveals Brassica rapa-specific evolutionary dynamics upon whole genome triplication. Mol. Genet. Genom. 2015, 290, 521–530. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, H.; Sun, G.; Jin, Y.; Qiu, L. Identification of active VQ motif-containing genes and the expression patterns under low nitrogen treatment in soybean. Gene 2014, 543, 237–243. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Clamp, M.; Cuff, J.; Searle, S.M.; Barton, G.J. The Jalview Java alignment editor. Bioinformatics 2004, 20, 426–427. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Li, P.T.; Chai, Z.; Lin, P.P.; Huang, C.H.; Huang, G.Q.; Xu, L.N.; Deng, Z.H.; Zhang, M.Q.; Zhang, Y.; Zhao, X.W. Genome-wide identification and expression analysis of AP2/ERF transcription factors in sugarcane (Saccharum spontaneum L.). BMC Genom. 2020, 21, 685. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhang, Y.; Zhang, Z.; Zhu, J.; Yu, J. KaKs_Calculator 2.0: A toolkit incorporating gamma-series methods and sliding window strategies. Genom. Proteom. Bioinform. 2010, 8, 77–80. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Zhang, C.; Wang, D.; Ren, Y.; Zhang, J.; Zang, S.; Zou, W.; Su, Y.; You, C.; Xu, L.; et al. A comprehensive survey of the aldehyde dehydrogenase gene superfamily in Saccharum and the role of ScALDH2B-1 in the stress response. Environ. Exp. Bot. 2022, 194, 104725. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. The R package Rsubread is easier, faster, cheaper and better for alignment and quantification of RNA sequencing reads. Nucleic Acids Res. 2019, 47, e47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.D.; Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 2010, 11, R25. [Google Scholar] [CrossRef] [PubMed] [Green Version]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Liu, X.; Yang, D.; Yin, Z.; Jiang, Y.; Ling, H.; Huang, N.; Zhang, D.; Wu, J.; Liu, L.; et al. A Comprehensive Identification and Expression Analysis of VQ Motif-Containing Proteins in Sugarcane (Saccharum spontaneum L.) under Phytohormone Treatment and Cold Stress. Int. J. Mol. Sci. 2022, 23, 6334. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23116334
Liu Y, Liu X, Yang D, Yin Z, Jiang Y, Ling H, Huang N, Zhang D, Wu J, Liu L, et al. A Comprehensive Identification and Expression Analysis of VQ Motif-Containing Proteins in Sugarcane (Saccharum spontaneum L.) under Phytohormone Treatment and Cold Stress. International Journal of Molecular Sciences. 2022; 23(11):6334. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23116334
Chicago/Turabian StyleLiu, Ying, Xiaolan Liu, Dandan Yang, Ze Yin, Yaolan Jiang, Hui Ling, Ning Huang, Dawei Zhang, Jinfeng Wu, Lili Liu, and et al. 2022. "A Comprehensive Identification and Expression Analysis of VQ Motif-Containing Proteins in Sugarcane (Saccharum spontaneum L.) under Phytohormone Treatment and Cold Stress" International Journal of Molecular Sciences 23, no. 11: 6334. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23116334