
The Sedentary Lifestyle and Masticatory Dysfunction: Time to Review the Contribution to Age-Associated Cognitive Decline and Astrocyte Morphotypes in the Dentate Gyrus


 , ,
, ,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
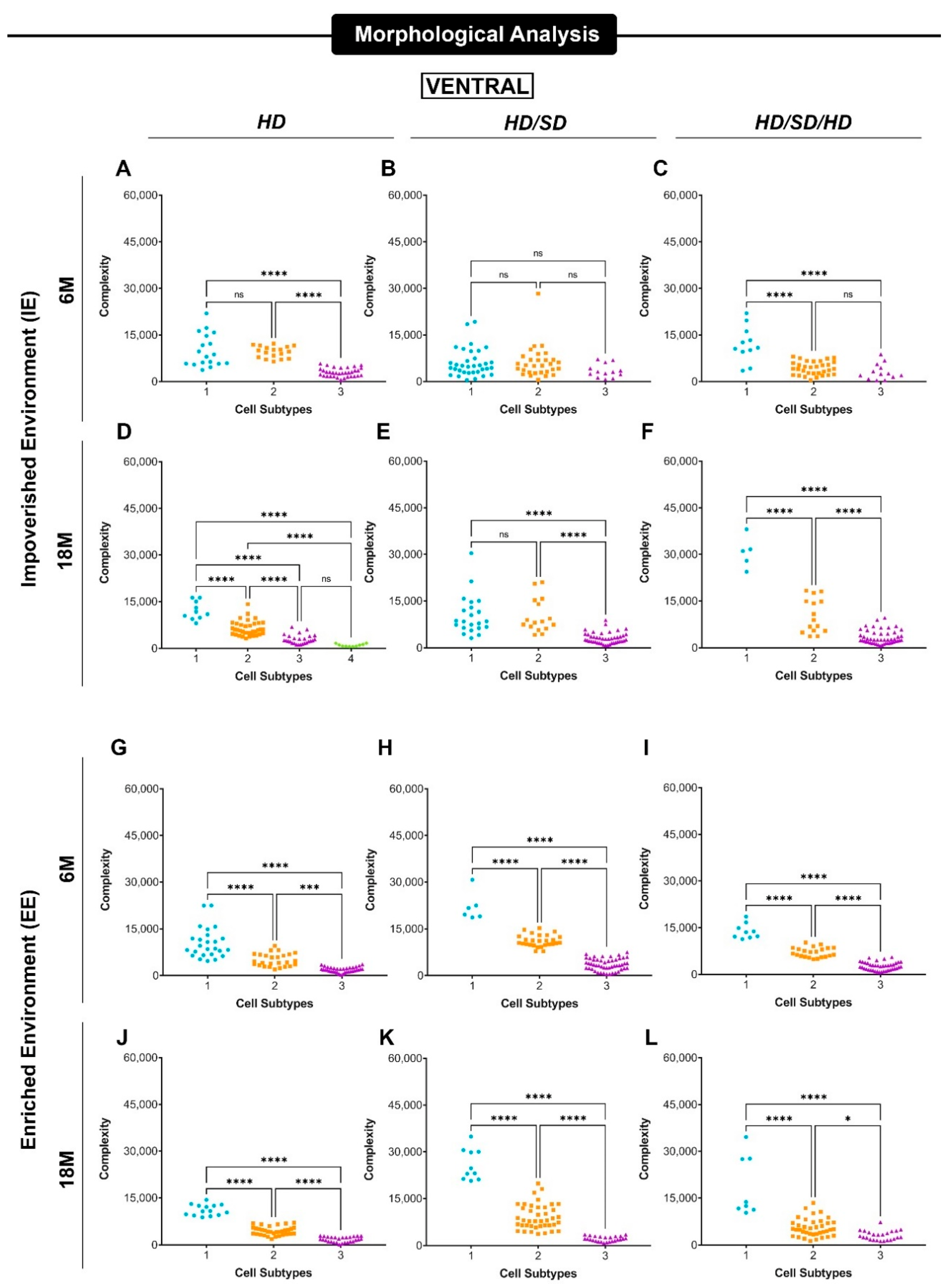{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Running, Experiencing Novelty, and Mastication to Learn Faster, Better Remember, and Enhance Individual Ethological Behavior
3. Enriched Environment and Masticatory Rehabilitation to Prevent Synaptic Dysfunction Associated with Age-Related Cognitive Decline
4. Dentate Gyrus Astrocytes, Long Life Sedentary Lifestyle and Dysfunctional Mastication
5. Differential Effects of Sedentary Lifestyle and Masticatory Dysfunction on Dorsal/Ventral Dentate Gyrus Morphological Phenotypes
6. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Latimer, C.S.; Searcy, J.L.; Bridges, M.T.; Brewer, L.D.; Popović, J.; Blalock, E.M.; Landfield, P.W.; Thibault, O.; Porter, N.M. Reversal of glial and neurovascular markers of unhealthy brain aging by exercise in middle-aged female mice. PLoS ONE 2011, 6, e26812. [Google Scholar] [CrossRef] [PubMed]
- Nishijima, T.; Llorens-Martín, M.; Tejeda, G.S.; Inoue, K.; Yamamura, Y.; Soya, H.; Trejo, J.L.; Torres-Alemán, I. Cessation of voluntary wheel running Increases anxiety-like behavior and impairs adult hippocampal neurogenesis in mice. Behav. Brain Res. 2013, 245, 34–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maasakkers, C.M.; Claassen, J.A.H.R.; Gardiner, P.A.; Olde Rikkert, M.G.M.; Lipnicki, D.M.; Scarmeas, N.; Dardiotis, E.; Yannakoulia, M.; Anstey, K.J.; Cherbuin, N.; et al. The Association of Sedentary Behaviour and Cognitive Function in People Without Dementia: A Coordinated Analysis Across Five Cohort Studies from COSMIC. Sports Med. 2020, 50, 403–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maasakkers, C.M.; Thijssen, D.H.; Knight, S.P.; Newman, L.; O’Connor, J.D.; Scarlett, S.; Carey, D.; Buckley, A.; McMorrow, J.P.; Leidhin, C.N.; et al. Hemodynamic and structural brain measures in high and low sedentary older adults. J. Cereb. Blood Flow Metab. 2021, 41, 2607–2616. [Google Scholar] [CrossRef] [PubMed]
- Puzo, C.; Labriola, C.; Sugarman, M.A.; Tripodis, Y.; Martin, B.; Palmisano, J.N.; Steinberg, E.G.; Stein, T.D.; Kowall, N.W.; McKee, A.C.; et al. Independent effects of white matter hyperintensities on cognitive, neuropsychiatric, and functional decline: A longitudinal investigation using the National Alzheimer’s Coordinating Center Uniform Data Set. Alzheimers Res. 2019, 11, 64. [Google Scholar] [CrossRef]
- Daly, B.; Thompsell, A.; Sharpling, J.; Rooney, Y.M.; Hillman, L.; Wanyonyi, K.L.; White, S.; Gallagher, J.E. Evidence summary: The relationship between oral health and dementia. Br. Dent. J. 2018, 223, 846–853. [Google Scholar] [CrossRef]
- Delwel, S.; Scherder, E.J.A.; Perez, R.S.G.M.; Hertogh, C.M.P.M.; Maier, A.B.; Lobbezoo, F. Oral function of older people with mild cognitive impairment or dementia. J. Oral. Rehabil. 2018, 45, 990–997. [Google Scholar] [CrossRef]
- Miquel, S.; Aspiras, M.; Day, J.E.L. Does reduced mastication influence cognitive and systemic health during aging? Physiol. Behav. 2018, 188, 239–250. [Google Scholar] [CrossRef]
- Saito, S.; Ohi, T.; Murakami, T.; Komiyama, T.; Miyoshi, Y.; Endo, K.; Satoh, M.; Asayama, K.; Inoue, R.; Kikuya, M.; et al. Association between tooth loss and cognitive impairment in community-dwelling older Japanese adults: A 4-year prospective cohort study from the Ohasama study. BMC Oral. Health 2018, 18, 142. [Google Scholar] [CrossRef]
- Galindo-Moreno, P.; Lopez-Chaichio, L.; Padial-Molina, M.; Avila-Ortiz, G.; O’Valle, F.; Ravida, A.; Catena, A. The impact of tooth loss on cognitive function. Clin. Oral. Investig. 2021, 26, 3493–3500. [Google Scholar] [CrossRef]
- Nakamura, T.; Zou, K.; Shibuya, Y.; Michikawa, M. Oral dysfunctions and cognitive impairment/dementia. J. Neurosci. Res. 2021, 99, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Furuya, J.; Hidaka, R.; Miyajima, S.; Matsubara, C.; Ohwada, G.; Asada, T.; Akazawa, C.; Sato, Y.; Tohara, H.; et al. Patients with mild cognitive impairment diagnosed at dementia clinic display decreased maximum occlusal force: A cross-sectional study. BMC Oral. Health 2021, 21, 665. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Huang, X.; Gong, Y.; Sun, J. Association between tooth loss rate and risk of mild cognitive impairment in older adults: A population-based longitudinal study. Aging (Albany NY) 2021, 13, 21599–21609. [Google Scholar] [CrossRef] [PubMed]
- Lauritano, D.; Moreo, G.; Della Vella, F.; Di Stasio, D.; Carinci, F.; Lucchese, A.; Petruzzi, M. Oral Health Status and Need for Oral Care in an Aging Population: A Systematic Review. Int. J. Env. Res. Public Health 2019, 16, 4558. [Google Scholar] [CrossRef] [Green Version]
- Marchini, L.; Ettinger, R.; Caprio, T.; Jucan, A. Oral health care for patients with Alzheimer’s disease: An update. Spec. Care Dent. 2019, 39, 262–273. [Google Scholar] [CrossRef]
- Scambler, S.; Curtis, S.; Manthorpe, J.; Samsi, K.; Rooney, Y.M.; Gallagher, J.E. The mouth and oral health in the field of dementia. Health 2021, 13634593211049891. [Google Scholar] [CrossRef]
- Kim, T.H. Effects of masticatory exercise on cognitive function in community-dwelling older adults. Technol. Health Care 2021, 29, 125–131. [Google Scholar] [CrossRef]
- Chen, H.; Iinuma, M.; Onozuka, M.; Kubo, K.Y. Chewing Maintains Hippocampus-Dependent Cognitive Function. Int. J. Med. Sci. 2015, 12, 502–509. [Google Scholar] [CrossRef] [Green Version]
- Tada, A.; Miura, H. Association between mastication and cognitive status: A systematic review. Arch. Gerontol. Geriatr. 2017, 70, 44–53. [Google Scholar] [CrossRef]
- Lin, C.S. Revisiting the link between cognitive decline and masticatory dysfunction. BMC Geriatr. 2018, 18, 5. [Google Scholar] [CrossRef]
- Alvarenga, M.O.P.; Ferreira, R.O.; Magno, M.B.; Fagundes, N.C.F.; Maia, L.C.; Lima, R.R. Masticatory Dysfunction by Extensive Tooth Loss as a Risk Factor for Cognitive Deficit: A Systematic Review and Meta-Analysis. Front. Physiol. 2019, 10, 832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Oh, J.K.; Wee, J.H.; Kim, Y.H.; Byun, S.H.; Choi, H.G. Association between Tooth Loss and Alzheimer’s Disease in a Nested Case-Control Study Based on a National Health Screening Cohort. J. Clin. Med. 2021, 10, 3763. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Geng, X.; Yun, H.J.; Haddad, Y.; Chen, Y.; Ding, Y. Neuroplastic Effect of Exercise Through Astrocytes Activation and Cellular Crosstalk. Aging Dis. 2021, 12, 1644–1657. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Rodríguez, G.; Ocaña-Fernández, M.A.; Vega-Rivera, N.M.; Torres-Pérez, O.M.; Gómez-Sánchez, A.; Estrada-Camarena, E.; Ortiz-López, L. Environmental enrichment induces neuroplastic changes in middle age female Balb/c mice and increases the hippocampal levels of BDNF, p-Akt and p-MAPK1/2. Neuroscience 2014, 260, 158–170. [Google Scholar] [CrossRef]
- Takeda, Y.; Oue, H.; Okada, S.; Kawano, A.; Koretake, K.; Michikawa, M.; Akagawa, Y.; Tsuga, K. Molar loss and powder diet leads to memory deficit and modifies the mRNA expression of brain-derived neurotrophic factor in the hippocampus of adult mice. BMC Neurosci. 2016, 17, 81. [Google Scholar] [CrossRef] [Green Version]
- Fukushima-Nakayama, Y.; Ono, T.; Hayashi, M.; Inoue, M.; Wake, H.; Nakashima, T. Reduced Mastication Impairs Memory Function. J. Dent. Res. 2017, 96, 1058–1066. [Google Scholar] [CrossRef]
- Piancino, M.G.; Tortarolo, A.; Polimeni, A.; Bramanti, E.; Bramanti, P. Altered mastication adversely impacts morpho-functional features of the hippocampus: A systematic review on animal studies in three different experimental conditions involving the masticatory function. PLoS ONE 2020, 15, e0237872. [Google Scholar] [CrossRef]
- Kubo, K.Y.; Ichihashi, Y.; Kurata, C.; Iinuma, M.; Mori, D.; Katayama, T.; Miyake, H.; Fujiwara, S.; Tamura, Y. Masticatory function and cognitive function. Okajimas Folia Anat. Jpn. 2010, 87, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Ono, Y.; Yamamoto, T.; Kubo, K.Y.; Onozuka, M. Occlusion and brain function: Mastication as a prevention of cognitive dysfunction. J. Oral. Rehabil. 2010, 37, 624–640. [Google Scholar] [CrossRef]
- Frota de Almeida, M.N.; de Siqueira Mendes, F.e.C.; Gurgel Felício, A.P.; Falsoni, M.; Ferreira de Andrade, M.L.; Bento-Torres, J.; da Costa Vasconcelos, P.F.; Perry, V.H.; Picanço-Diniz, C.W.; Kronka Sosthenes, M.C. Spatial memory decline after masticatory deprivation and aging is associated with altered laminar distribution of CA1 astrocytes. BMC Neurosci. 2012, 13, 23. [Google Scholar] [CrossRef] [Green Version]
- Mendes, F.e.C.; de Almeida, M.N.; Felício, A.P.; Fadel, A.C.; Silva, D.e.J.; Borralho, T.G.; da Silva, R.P.; Bento-Torres, J.; Vasconcelos, P.F.; Perry, V.H.; et al. Enriched environment and masticatory activity rehabilitation recover spatial memory decline in aged mice. BMC Neurosci. 2013, 14, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekuni, D.; Tomofuji, T.; Irie, K.; Azuma, T.; Endo, Y.; Kasuyama, K.; Morita, M. Occlusal disharmony increases amyloid-beta in the rat hippocampus. Neuromol. Med. 2011, 13, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Ekuni, D.; Endo, Y.; Tomofuji, T.; Azuma, T.; Irie, K.; Kasuyama, K.; Morita, M. Effects of apoE deficiency and occlusal disharmony on amyloid-beta production and spatial memory in rats. PLoS ONE 2013, 8, e74966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeshiba, M.; Kajiya, H.; Tsutsumi, T.; Migita, K.; Goto-T, K.; Kono, Y.; Tsuzuki, T.; Ohno, J. Occlusal disharmony transiently decrease cognition via cognitive suppressor molecules and partially restores cognitive ability via clearance molecules. Biochem. Biophys. Res. Commun. 2022, 594, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.; Torres, C. Astrocyte senescence: Evidence and significance. Aging Cell 2019, 18, e12937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales-Rosales, S.L.; Santín-Márquez, R.; Posadas-Rodriguez, P.; Rincon-Heredia, R.; Montiel, T.; Librado-Osorio, R.; Luna-López, A.; Rivero-Segura, N.A.; Torres, C.; Cano-Martínez, A.; et al. Senescence in Primary Rat Astrocytes Induces Loss of the Mitochondrial Membrane Potential and Alters Mitochondrial Dynamics in Cortical Neurons. Front. Aging Neurosci. 2021, 13, 766306. [Google Scholar] [CrossRef]
- Pannese, E. Quantitative, structural and molecular changes in neuroglia of aging mammals: A review. Eur. J. Histochem. 2021, 65, 3249. [Google Scholar] [CrossRef]
- Barter, J.D.; Foster, T.C. Aging in the Brain: New Roles of Epigenetics in Cognitive Decline. Neuroscientist 2018, 24, 516–525. [Google Scholar] [CrossRef]
- Lupo, G.; Gaetani, S.; Cacci, E.; Biagioni, S.; Negri, R. Molecular Signatures of the Aging Brain: Finding the Links Between Genes and Phenotypes. Neurotherapeutics 2019, 16, 543–553. [Google Scholar] [CrossRef]
- González-Velasco, O.; Papy-García, D.; Le Douaron, G.; Sánchez-Santos, J.M.; De Las Rivas, J. Transcriptomic landscape, gene signatures and regulatory profile of aging in the human brain. Biochim. Biophys Acta Gene Regul. Mech. 2020, 1863, 194491. [Google Scholar] [CrossRef]
- Diniz, D.G.; de Oliveira, M.A.; de Lima, C.M.; Foro, C.A.R.; Sosthenes, M.C.K.; Bento-Torres, J.; Vasconcelos, P.F.D.; Anthony, D.C.; Diniz, C.W.P. Age, environment, object recognition and morphological diversity of GFAP-immunolabeled astrocytes. Behav. Brain Funct. 2016, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maugeri, G.; D’Agata, V.; Magrì, B.; Roggio, F.; Castorina, A.; Ravalli, S.; Di Rosa, M.; Musumeci, G. Neuroprotective Effects of Physical Activity via the Adaptation of Astrocytes. Cells 2021, 10, 1542. [Google Scholar] [CrossRef] [PubMed]
- Matias, I.; Morgado, J.; Gomes, F.C.A. Astrocyte Heterogeneity: Impact to Brain Aging and Disease. Front. Aging Neurosci. 2019, 11, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matias, I.; Diniz, L.P.; Damico, I.V.; Araujo, A.P.B.; Neves, L.D.S.; Vargas, G.; Leite, R.E.P.; Suemoto, C.K.; Nitrini, R.; Jacob-Filho, W.; et al. Loss of lamin-B1 and defective nuclear morphology are hallmarks of astrocyte senescence in vitro and in the aging human hippocampus. Aging Cell 2022, 21, e13521. [Google Scholar] [CrossRef]
- Bento-Torres, J.; Sobral, L.L.; Reis, R.R.; de Oliveira, R.B.; Anthony, D.C.; Vasconcelos, P.F.C.; Diniz, C.W.P. Age and Environment Influences on Mouse Prion Disease Progression: Behavioral Changes and Morphometry and Stereology of Hippocampal Astrocytes. Oxidative Med. Cell. Longev. 2017, 2017, 4504925. [Google Scholar] [CrossRef]
- Moulson, A.J.; Squair, J.W.; Franklin, R.J.M.; Tetzlaff, W.; Assinck, P. Diversity of Reactive Astrogliosis in CNS Pathology: Heterogeneity or Plasticity? Front. Cell Neurosci. 2021, 15, 703810. [Google Scholar] [CrossRef]
- Preman, P.; Alfonso-Triguero, M.; Alberdi, E.; Verkhratsky, A.; Arranz, A.M. Astrocytes in Alzheimer’s Disease: Pathological Significance and Molecular Pathways. Cells 2021, 10, 540. [Google Scholar] [CrossRef]
- Ferrer, I. Diversity of astroglial responses across human neurodegenerative disorders and brain aging. Brain Pathol. 2017, 27, 645–674. [Google Scholar] [CrossRef]
- Escartin, C.; Galea, E.; Lakatos, A.; O’Callaghan, J.P.; Petzold, G.C.; Serrano-Pozo, A.; Steinhäuser, C.; Volterra, A.; Carmignoto, G.; Agarwal, A.; et al. Reactive astrocyte nomenclature, definitions, and future directions. Nat. Neurosci. 2021, 24, 312–325. [Google Scholar] [CrossRef]
- Viola, G.G.; Rodrigues, L.; Americo, J.C.; Hansel, G.; Vargas, R.S.; Biasibetti, R.; Swarowsky, A.; Goncalves, C.A.; Xavier, L.L.; Achaval, M.; et al. Morphological changes in hippocampal astrocytes induced by environmental enrichment in mice. Brain Res. 2009, 1274, 47–54. [Google Scholar] [CrossRef]
- Saur, L.; Baptista, P.P.; de Senna, P.N.; Paim, M.F.; do Nascimento, P.; Ilha, J.; Bagatini, P.B.; Achaval, M.; Xavier, L.L. Physical exercise increases GFAP expression and induces morphological changes in hippocampal astrocytes. Brain Struct. Funct. 2014, 219, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Viola, G.G.; Loss, C.M. Letter to Editor about: “Physical exercise increases GFAP expression and induces morphological changes in hippocampal astrocytes”. Brain Struct. Funct. 2014, 219, 1509–1510. [Google Scholar] [CrossRef] [PubMed]
- Lundquist, A.J.; Parizher, J.; Petzinger, G.M.; Jakowec, M.W. Exercise induces region-specific remodeling of astrocyte morphology and reactive astrocyte gene expression patterns in male mice. J. Neurosci. Res. 2019, 97, 1081–1094. [Google Scholar] [CrossRef]
- Cooper, C.; Moon, H.Y.; van Praag, H. On the Run for Hippocampal Plasticity. Cold Spring Harb. Perspect. Med. 2018, 8, a029736. [Google Scholar] [CrossRef] [PubMed]
- Kempermann, G. Environmental enrichment, new neurons and the neurobiology of individuality. Nat. Rev. Neurosci. 2019, 20, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Onozuka, M.; Watanabe, K.; Nagasaki, S.; Jiang, Y.; Ozono, S.; Nishiyama, K.; Kawase, T.; Karasawa, N.; Nagatsu, I. Impairment of spatial memory and changes in astroglial responsiveness following loss of molar teeth in aged SAMP8 mice. Behav. Brain Res. 2000, 108, 145–155. [Google Scholar] [CrossRef]
- Mendes, F.C.C.S.; Felício, A.P.G.; Diniz, C.W.P.; Sosthenes, M.C.K. Alteração mastigatória, ambiente enriquecido e envelhecimento: Estudos estereológicos de CA1 do hipocampo de camundongos suíços albinos. Rev. Pan-Amaz. Saude. 2016, 7, 31–40. [Google Scholar] [CrossRef]
- Kida, K.; Tsuji, T.; Tanaka, S.; Kogo, M. Zinc deficiency with reduced mastication impairs spatial memory in young adult mice. Physiol. Behav. 2015, 152, 231–237. [Google Scholar] [CrossRef] [Green Version]
- De Siqueira Mendes, F.C.C.; Paixão, L.T.V.B.; Diniz, D.G.; Anthony, D.C.; Brites, D.; Diniz, C.W.P.; Sosthenes, M.C.K. Sedentary Life and Reduced Mastication Impair Spatial Learning and Memory and Differentially Affect Dentate Gyrus Astrocyte Subtypes in the Aged Mice. Front Neurosci. 2021, 15, 632216. [Google Scholar] [CrossRef]
- Batiuk, M.Y.; Martirosyan, A.; Wahis, J.; de Vin, F.; Marneffe, C.; Kusserow, C.; Koeppen, J.; Viana, J.F.; Oliveira, J.F.; Voet, T.; et al. Identification of region-specific astrocyte subtypes at single cell resolution. Nat. Commun. 2020, 11, 1220. [Google Scholar] [CrossRef] [Green Version]
- Chai, H.; Diaz-Castro, B.; Shigetomi, E.; Monte, E.; Octeau, J.C.; Yu, X.; Cohn, W.; Rajendran, P.S.; Vondriska, T.M.; Whitelegge, J.P.; et al. Neural Circuit-Specialized Astrocytes: Transcriptomic, Proteomic, Morphological, and Functional Evidence. Neuron 2017, 95, 531–549.e539. [Google Scholar] [CrossRef] [PubMed]
- Clarke, L.E.; Liddelow, S.A.; Chakraborty, C.; Münch, A.E.; Heiman, M.; Barres, B.A. Normal aging induces A1-like astrocyte reactivity. Proc. Natl. Acad. Sci. USA 2018, 115, E1896–E1905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boisvert, M.M.; Erikson, G.A.; Shokhirev, M.N.; Allen, N.J. The Aging Astrocyte Transcriptome from Multiple Regions of the Mouse Brain. Cell Rep. 2018, 22, 269–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munger, E.L.; Edler, M.K.; Hopkins, W.D.; Ely, J.J.; Erwin, J.M.; Perl, D.P.; Mufson, E.J.; Hof, P.R.; Sherwood, C.C.; Raghanti, M.A. Astrocytic changes with aging and Alzheimer’s disease-type pathology in chimpanzees. J. Comp. Neurol. 2019, 527, 1179–1195. [Google Scholar] [CrossRef]
- Bettcher, B.M.; Olson, K.E.; Carlson, N.E.; McConnell, B.V.; Boyd, T.; Adame, V.; Solano, D.A.; Anton, P.; Markham, N.; Thaker, A.A.; et al. Astrogliosis and episodic memory in late life: Higher GFAP is related to worse memory and white matter microstructure in healthy aging and Alzheimer’s disease. Neurobiol. Aging 2021, 103, 68–77. [Google Scholar] [CrossRef]
- Adamsky, A.; Goshen, I. Astrocytes in Memory Function: Pioneering Findings and Future Directions. Neuroscience 2018, 370, 14–26. [Google Scholar] [CrossRef]
- Navarrete, M.; Cuartero, M.I.; Palenzuela, R.; Draffin, J.E.; Konomi, A.; Serra, I.; Colié, S.; Castaño-Castaño, S.; Hasan, M.T.; Nebreda, Á.; et al. Astrocytic p38α MAPK drives NMDA receptor-dependent long-term depression and modulates long-term memory. Nat. Commun. 2019, 10, 2968. [Google Scholar] [CrossRef] [Green Version]
- Iwai, Y.; Ozawa, K.; Yahagi, K.; Mishima, T.; Akther, S.; Vo, C.T.; Lee, A.B.; Tanaka, M.; Itohara, S.; Hirase, H. Transient Astrocytic Gq Signaling Underlies Remote Memory Enhancement. Front. Neural. Circuits 2021, 15, 658343. [Google Scholar] [CrossRef]
- Van Den Herrewegen, Y.; Sanderson, T.M.; Sahu, S.; De Bundel, D.; Bortolotto, Z.A.; Smolders, I. Side-by-side comparison of the effects of Gq- and Gi-DREADD-mediated astrocyte modulation on intracellular calcium dynamics and synaptic plasticity in the hippocampal CA1. Mol. Brain. 2021, 14, 144. [Google Scholar] [CrossRef]
- Corkrum, M.; Covelo, A.; Lines, J.; Bellocchio, L.; Pisansky, M.; Loke, K.; Quintana, R.; Rothwell, P.E.; Lujan, R.; Marsicano, G.; et al. Dopamine-Evoked Synaptic Regulation in the Nucleus Accumbens Requires Astrocyte Activity. Neuron 2020, 105, 1036–1047.e1035. [Google Scholar] [CrossRef] [Green Version]
- Lines, J.; Martin, E.D.; Kofuji, P.; Aguilar, J.; Araque, A. Astrocytes modulate sensory-evoked neuronal network activity. Nat. Commun. 2020, 11, 3689. [Google Scholar] [CrossRef] [PubMed]
- Vaidyanathan, T.V.; Collard, M.; Yokoyama, S.; Reitman, M.E.; Poskanzer, K.E. Cortical astrocytes independently regulate sleep depth and duration via separate GPCR pathways. Elife 2021, 10, e63329. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, P.; Qi, Y.; Xu, Z.; Yang, Y. Activation of hypothalamic astrocytes suppresses feeding without altering emotional states. Glia 2016, 64, 2263–2273. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, L.; Wu, J.; Zhu, Z.; Feng, X.; Qin, L.; Zhu, Y.; Sun, L.; Liu, Y.; Qiu, Z.; et al. Activation of astrocytes in hippocampus decreases fear memory through adenosine A. Elife 2020, 9, e57155. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Lu, C.L.; Huang, J.; Fan, J.; Guo, F.; Mo, J.W.; Huang, W.Y.; Kong, P.L.; Li, X.W.; Sun, L.R.; et al. A Distinct Metabolically Defined Central Nucleus Circuit Bidirectionally Controls Anxiety-Related Behaviors. J. Neurosci. 2022, 42, 2356–2370. [Google Scholar] [CrossRef]
- Lyon, K.A.; Allen, N.J. From Synapses to Circuits, Astrocytes Regulate Behavior. Front. Neural Circuits 2021, 15, 786293. [Google Scholar] [CrossRef]
- Nagai, J.; Yu, X.; Papouin, T.; Cheong, E.; Freeman, M.R.; Monk, K.R.; Hastings, M.H.; Haydon, P.G.; Rowitch, D.; Shaham, S.; et al. Behaviorally consequential astrocytic regulation of neural circuits. Neuron 2021, 109, 576–596. [Google Scholar] [CrossRef]
- Patten, A.R.; Sickmann, H.; Hryciw, B.N.; Kucharsky, T.; Parton, R.; Kernick, A.; Christie, B.R. Long-term exercise is needed to enhance synaptic plasticity in the hippocampus. Learn Mem. 2013, 20, 642–647. [Google Scholar] [CrossRef] [Green Version]
- Vivar, C.; van Praag, H. Running Changes the Brain: The Long and the Short of It. Physiology (Bethesda) 2017, 32, 410–424. [Google Scholar] [CrossRef]
- Van Praag, H. Neurogenesis and exercise: Past and future directions. Neuromolecular Med. 2008, 10, 128–140. [Google Scholar] [CrossRef]
- Huang, Y.Q.; Wu, C.; He, X.F.; Wu, D.; He, X.; Liang, F.Y.; Dai, G.Y.; Pei, Z.; Xu, G.Q.; Lan, Y. Effects of Voluntary Wheel-Running Types on Hippocampal Neurogenesis and Spatial Cognition in Middle-Aged Mice. Front. Cell Neurosci. 2018, 12, 177. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.Y.; Keown, C.L.; Wen, X.; Li, J.; Vousden, D.A.; Anacker, C.; Bhattacharyya, U.; Ryan, R.; Diorio, J.; O’Toole, N.; et al. Environmental enrichment increases transcriptional and epigenetic differentiation between mouse dorsal and ventral dentate gyrus. Nat. Commun. 2018, 9, 298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, D.; Li, C.; Swanson, A.M.; Villalba, R.M.; Guo, J.; Zhang, Z.; Matheny, S.; Murakami, T.; Stephenson, J.R.; Daniel, S.; et al. BAI1 regulates spatial learning and synaptic plasticity in the hippocampus. J. Clin. Investig. 2015, 125, 1497–1508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birch, A.M.; McGarry, N.B.; Kelly, A.M. Short-term environmental enrichment, in the absence of exercise, improves memory, and increases NGF concentration, early neuronal survival, and synaptogenesis in the dentate gyrus in a time-dependent manner. Hippocampus 2013, 23, 437–450. [Google Scholar] [CrossRef]
- Birch, A.M.; Kelly, Á. Lifelong environmental enrichment in the absence of exercise protects the brain from age-related cognitive decline. Neuropharmacology 2019, 145, 59–74. [Google Scholar] [CrossRef]
- Diniz, D.G.; Foro, C.A.R.; Rego, C.M.D.; Gloria, D.A.; de Oliveira, F.R.R.; Paes, J.M.P.; de Sousa, A.A.; Tokuhashi, T.P.; Trindade, L.S.; Turiel, M.C.P.; et al. Environmental impoverishment and aging alter object recognition, spatial learning, and dentate gyrus astrocytes. Eur. J. Neurosci. 2010, 32, 509–519. [Google Scholar] [CrossRef] [Green Version]
- Harland, B.C.; Dalrymple-Alford, J.C. Enriched Environment Procedures for Rodents: Creating a Standardized Protocol for Diverse Enrichment to Improve Consistency across Research Studies. Bio Protoc. 2020, 10, e3637. [Google Scholar] [CrossRef]
- Kobilo, T.; Liu, Q.R.; Gandhi, K.; Mughal, M.; Shaham, Y.; van Praag, H. Running is the neurogenic and neurotrophic stimulus in environmental enrichment. Learn Mem. 2011, 18, 605–609. [Google Scholar] [CrossRef] [Green Version]
- Grégoire, C.A.; Bonenfant, D.; Le Nguyen, A.; Aumont, A.; Fernandes, K.J. Untangling the influences of voluntary running, environmental complexity, social housing and stress on adult hippocampal neurogenesis. PLoS ONE 2014, 9, e86237. [Google Scholar] [CrossRef] [Green Version]
- Robison, L.S.; Francis, N.; Popescu, D.L.; Anderson, M.E.; Hatfield, J.; Xu, F.; Anderson, B.J.; Van Nostrand, W.E.; Robinson, J.K. Environmental Enrichment: Disentangling the Influence of Novelty, Social, and Physical Activity on Cerebral Amyloid Angiopathy in a Transgenic Mouse Model. Int. J. Mol. Sci. 2020, 21, 843. [Google Scholar] [CrossRef] [Green Version]
- Wolf, M.; Weissing, F.J. Animal personalities: Consequences for ecology and evolution. Trends Ecol. Evol. 2012, 27, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Füzesi, T.; Daviu, N.; Wamsteeker Cusulin, J.I.; Bonin, R.P.; Bains, J.S. Hypothalamic CRH neurons orchestrate complex behaviours after stress. Nat. Commun. 2016, 7, 11937. [Google Scholar] [CrossRef] [PubMed]
- Kentner, A.C.; Speno, A.V.; Doucette, J.; Roderick, R.C. The Contribution of Environmental Enrichment to Phenotypic Variation in Mice and Rats. eNeuro 2021, 8. [Google Scholar] [CrossRef]
- Komada, M.; Takao, K.; Miyakawa, T. Elevated plus maze for mice. J. Vis. Exp. 2008, 22, 1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ennaceur, A. Tests of unconditioned anxiety—Pitfalls and disappointments. Physiol. Behav. 2014, 135, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Tapper, A.R.; Molas, S. Midbrain circuits of novelty processing. Neurobiol. Learn Mem. 2020, 176, 107323. [Google Scholar] [CrossRef]
- Schomaker, J.; Meeter, M. Short- and long-lasting consequences of novelty, deviance and surprise on brain and cognition. Neurosci. Biobehav. Rev. 2015, 55, 268–279. [Google Scholar] [CrossRef]
- Girbovan, C.; Plamondon, H. Environmental enrichment in female rodents: Considerations in the effects on behavior and biochemical markers. Behav. Brain Res. 2013, 253, 178–190. [Google Scholar] [CrossRef]
- Wang, H.; Xu, X.; Gao, J.; Zhang, T. Enriched Environment and Social Isolation Affect Cognition Ability via Altering Excitatory and Inhibitory Synaptic Density in Mice Hippocampus. Neurochem. Res. 2020, 45, 2417–2432. [Google Scholar] [CrossRef]
- Da Silva, J.D.; Ni, S.C.; Lee, C.; Elani, H.; Ho, K.; Thomas, C.; Kuwajima, Y.; Ishida, Y.; Kobayashi, T.; Ishikawa-Nagai, S. Association between cognitive health and masticatory conditions: A descriptive study of the national database of the universal healthcare system in Japan. Aging (Albany N.Y.) 2021, 13, 7943–7952. [Google Scholar] [CrossRef]
- Krishnamoorthy, G.; Narayana, A.I.; Balkrishanan, D. Mastication as a tool to prevent cognitive dysfunctions. Jpn. Dent. Sci. Rev. 2018, 54, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Chuhuaicura, P.; Dias, F.J.; Arias, A.; Lezcano, M.F.; Fuentes, R. Mastication as a protective factor of the cognitive decline in adults: A qualitative systematic review. Int. Dent. J. 2019, 69, 334–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Chaichio, L.; Padial-Molina, M.; O’Valle, F.; Gil-Montoya, J.A.; Catena, A.; Galindo-Moreno, P. Oral health and healthy chewing for healthy cognitive ageing: A comprehensive narrative review. Gerodontology 2021, 38, 126–135. [Google Scholar] [CrossRef]
- Ahmed, S.E.; Mohan, J.; Kalaignan, P.; Kandasamy, S.; Raju, R.; Champakesan, B. Influence of Dental Prostheses on Cognitive Functioning in Elderly Population: A Systematic Review. J. Pharm. Bioallied. Sci. 2021, 13, S788–S794. [Google Scholar] [CrossRef]
- Tsutsui, K.; Kaku, M.; Motokawa, M.; Tohma, Y.; Kawata, T.; Fujita, T.; Kohno, S.; Ohtani, J.; Tenjoh, K.; Nakano, M.; et al. Influences of reduced masticatory sensory input from soft-diet feeding upon spatial memory/learning ability in mice. Biomed. Res. 2007, 28, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, H.; Kurahashi, M.; Mori, D.; Iinuma, M.; Tamura, Y.; Mizutani, K.; Shimpo, K.; Sonoda, S.; Azuma, K.; Kubo, K.Y. Hippocampus-dependent spatial memory impairment due to molar tooth loss is ameliorated by an enriched environment. Arch. Oral. Biol. 2016, 61, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Weijenberg, R.A.F.; Delwel, S.; Ho, B.V.; van der Maarel-Wierink, C.D.; Lobbezoo, F. Mind your teeth-The relationship between mastication and cognition. Gerodontology 2019, 36, 2–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arakawa, Y.; Ichihashi, Y.; Iinuma, M.; Tamura, Y.; Iwaku, F.; Kubo, K.Y. Duration-dependent effects of the bite-raised condition on hippocampal function in SAMP8 mice. Okajimas Folia Anat. Jpn. 2007, 84, 115–119. [Google Scholar] [CrossRef] [Green Version]
- Kubo, K.Y.; Yamada, Y.; Iinuma, M.; Iwaku, F.; Tamura, Y.; Watanabe, K.; Nakamura, H.; Onozuka, M. Occlusal disharmony induces spatial memory impairment and hippocampal neuron degeneration via stress in SAMP8 mice. Neurosci. Lett. 2007, 414, 188–191. [Google Scholar] [CrossRef]
- Mori, D.; Katayama, T.; Miyake, H.; Fujiwara, S.; Kubo, K.Y. Occlusal disharmony leads to learning deficits associated with decreased cellular proliferation in the hippocampal dentate gyrus of SAMP8 mice. Neurosci. Lett. 2013, 534, 228–232. [Google Scholar] [CrossRef]
- Yoshihara, T.; Matsumoto, Y.; Ogura, T. Occlusal disharmony affects plasma corticosterone and hypothalamic noradrenaline release in rats. J. Dent. Res. 2001, 80, 2089–2092. [Google Scholar] [CrossRef] [PubMed]
- Iinuma, M.; Ichihashi, Y.; Hioki, Y.; Kurata, C.; Tamura, Y.; Kubo, K.Y. Malocclusion induces chronic stress. Okajimas Folia Anat. Jpn. 2008, 85, 35–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.; Li, J.; Jiang, T.; Han, S.H.; Yao, D.Y. Experimental occlusal disharmony—A promoting factor for anxiety in rats under chronic psychological stress. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 75, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Kubo, K.Y.; Iinuma, M.; Chen, H. Mastication as a Stress-Coping Behavior. Biomed. Res. Int. 2015, 2015, 876409. [Google Scholar] [CrossRef] [PubMed]
- Azuma, K.; Zhou, Q.; Niwa, M.; Kubo, K.Y. Association between Mastication, the Hippocampus, and the HPA Axis: A Comprehensive Review. Int. J. Mol. Sci. 2017, 18, 1687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cait, J.; Cait, A.; Scott, R.W.; Winder, C.B.; Mason, G.J. Conventional laboratory housing increases morbidity and mortality in research rodents: Results of a meta-analysis. BMC Biol. 2022, 20, 15. [Google Scholar] [CrossRef] [PubMed]
- Ono, Y.; Koizumi, S.; Onozuka, M. Chewing prevents stress-induced hippocampal LTD formation and anxiety-related behaviors: A possible role of the dopaminergic system. Biomed. Res. Int. 2015, 2015, 294068. [Google Scholar] [CrossRef]
- Yaoita, F.; Tsuchiya, M.; Arai, Y.; Tadano, T.; Tan-No, K. Involvement of catecholaminergic and GABAAergic mediations in the anxiety-related behavior in long-term powdered diet-fed mice. Neurochem. Int. 2019, 124, 1–9. [Google Scholar] [CrossRef]
- Ono, Y.; Kataoka, T.; Miyake, S.; Cheng, S.J.; Tachibana, A.; Sasaguri, K.I.; Onozuka, M. Chewing ameliorates stress-induced suppression of hippocampal long-term potentiation. Neuroscience 2008, 154, 1352–1359. [Google Scholar] [CrossRef]
- Ono, Y.; Kataoka, T.; Miyake, S.; Sasaguri, K.; Sato, S.; Onozuka, M. Chewing rescues stress-suppressed hippocampal long-term potentiation via activation of histamine H1 receptor. Neurosci. Res. 2009, 64, 385–390. [Google Scholar] [CrossRef]
- Ogawa, M.; Nagai, T.; Saito, Y.; Miyaguchi, H.; Kumakura, K.; Abe, K.; Asakura, T. Short-term mastication after weaning upregulates GABAergic signalling and reduces dendritic spine in thalamus. Biochem. Biophys. Res. Commun. 2018, 498, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Sasaguri, K.; Yamada, K.; Yamamoto, T. Uncovering the neural circuitry involved in the stress-attenuation effects of chewing. Jpn. Dent. Sci. Rev. 2018, 54, 118–126. [Google Scholar] [CrossRef] [PubMed]
- De Siqueira Mendes, F.C.C.; da Paixão, L.T.V.B.; Diniz, C.W.P.; Sosthenes, M.C.K. Environmental Impoverishment, Aging, and Reduction in Mastication Affect Mouse Innate Repertoire to Explore Novel Environments and to Assess Risk. Front. Neurosci. 2019, 13, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, C.L.; Ma, X.T.; Wang, J.J.; Liu, H.; Chen, Y.F.; Yang, Y. Physical exercise induces hippocampal neurogenesis and prevents cognitive decline. Behav. Brain. Res. 2017, 317, 332–339. [Google Scholar] [CrossRef]
- Wang, X.; Hu, J.; Jiang, Q. Tooth Loss-Associated Mechanisms That Negatively Affect Cognitive Function: A Systematic Review of Animal Experiments Based on Occlusal Support Loss and Cognitive Impairment. Front. Neurosci. 2022, 16, 811335. [Google Scholar] [CrossRef] [PubMed]
- Eichenbaum, H.; Yonelinas, A.P.; Ranganath, C. The medial temporal lobe and recognition memory. Annu. Rev. Neurosci. 2007, 30, 123–152. [Google Scholar] [CrossRef] [Green Version]
- Hiscox, L.V.; Johnson, C.L.; McGarry, M.D.J.; Schwarb, H.; van Beek, E.J.R.; Roberts, N.; Starr, J.M. Hippocampal viscoelasticity and episodic memory performance in healthy older adults examined with magnetic resonance elastography. Brain. Imaging Behav. 2020, 14, 175–185. [Google Scholar] [CrossRef] [Green Version]
- Sander, M.C.; Fandakova, Y.; Grandy, T.H.; Shing, Y.L.; Werkle-Bergner, M. Oscillatory Mechanisms of Successful Memory Formation in Younger and Older Adults Are Related to Structural Integrity. Cereb. Cortex. 2020, 30, 3744–3758. [Google Scholar] [CrossRef] [Green Version]
- Harris, K.M. Structural LTP: From synaptogenesis to regulated synapse enlargement and clustering. Curr. Opin. Neurobiol. 2020, 63, 189–197. [Google Scholar] [CrossRef]
- Caroni, P.; Chowdhury, A.; Lahr, M. Synapse rearrangements upon learning: From divergent-sparse connectivity to dedicated sub-circuits. Trends Neurosci. 2014, 37, 604–614. [Google Scholar] [CrossRef]
- Hrybouski, S.; MacGillivray, M.; Huang, Y.; Madan, C.R.; Carter, R.; Seres, P.; Malykhin, N.V. Involvement of hippocampal subfields and anterior-posterior subregions in encoding and retrieval of item, spatial, and associative memories: Longitudinal versus transverse axis. Neuroimage 2019, 191, 568–586. [Google Scholar] [CrossRef] [PubMed]
- Scully, D.; Fedriani, R.; Desouza, I.E.; Murphy, K.J.; Regan, C.M. Regional dissociation of paradigm-specific synapse remodeling during memory consolidation in the adult rat dentate gyrus. Neuroscience 2012, 209, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Van Groen, T.; Miettinen, P.; Kadish, I. The entorhinal cortex of the mouse: Organization of the projection to the hippocampal formation. Hippocampus 2003, 13, 133–149. [Google Scholar] [CrossRef] [PubMed]
- Nilssen, E.S.; Doan, T.P.; Nigro, M.J.; Ohara, S.; Witter, M.P. Neurons and networks in the entorhinal cortex: A reappraisal of the lateral and medial entorhinal subdivisions mediating parallel cortical pathways. Hippocampus 2019, 29, 1238–1254. [Google Scholar] [CrossRef] [Green Version]
- Jourdain, P.; Bergersen, L.H.; Bhaukaurally, K.; Bezzi, P.; Santello, M.; Domercq, M.; Matute, C.; Tonello, F.; Gundersen, V.; Volterra, A. Glutamate exocytosis from astrocytes controls synaptic strength. Nat. Neurosci. 2007, 10, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Savtchouk, I.; Di Castro, M.A.; Ali, R.; Stubbe, H.; Luján, R.; Volterra, A. Circuit-specific control of the medial entorhinal inputs to the dentate gyrus by atypical presynaptic NMDARs activated by astrocytes. Proc. Natl. Acad. Sci. USA 2019, 116, 13602–13610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Castro, M.A.; Volterra, A. Astrocyte control of the entorhinal cortex-dentate gyrus circuit: Relevance to cognitive processing and impairment in pathology. Glia 2021. [Google Scholar] [CrossRef]
- Akther, S.; Hirase, H. Assessment of astrocytes as a mediator of memory and learning in rodents. Glia 2021. [Google Scholar] [CrossRef]
- Nguyen, A.Q.; Koeppen, J.; Woodruff, S.; Mina, K.; Figueroa, Z.; Ethell, I.M. Astrocytic Ephrin-B1 Controls Synapse Formation in the Hippocampus During Learning and Memory. Front. Synaptic Neurosci. 2020, 12, 10. [Google Scholar] [CrossRef]
- Kofuji, P.; Araque, A. G-Protein-Coupled Receptors in Astrocyte-Neuron Communication. Neuroscience 2021, 456, 71–84. [Google Scholar] [CrossRef]
- Adamsky, A.; Kol, A.; Kreisel, T.; Doron, A.; Ozeri-Engelhard, N.; Melcer, T.; Refaeli, R.; Horn, H.; Regev, L.; Groysman, M.; et al. Astrocytic Activation Generates De Novo Neuronal Potentiation and Memory Enhancement. Cell 2018, 174, 59–71.e14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Covelo, A.; Araque, A. Stimulating Astrocytes to Remember. Cell 2018, 174, 12–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loprinzi, P.D. The role of astrocytes on the effects of exercise on episodic memory function. Physiol. Int. 2019, 106, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Pillai, A.G.; de Jong, D.; Kanatsou, S.; Krugers, H.; Knapman, A.; Heinzmann, J.M.; Holsboer, F.; Landgraf, R.; Joëls, M.; Touma, C. Dendritic morphology of hippocampal and amygdalar neurons in adolescent mice is resilient to genetic differences in stress reactivity. PLoS ONE 2012, 7, e38971. [Google Scholar] [CrossRef] [Green Version]
- Verkhratsky, A.; Augusto-Oliveira, M.; Pivoriūnas, A.; Popov, A.; Brazhe, A.; Semyanov, A. Astroglial asthenia and loss of function, rather than reactivity, contribute to the ageing of the brain. Pflug. Arch 2021, 473, 753–774. [Google Scholar] [CrossRef]
- Porchet, R.; Probst, A.; Bouras, C.; Dráberová, E.; Dráber, P.; Riederer, B.M. Analysis of glial acidic fibrillary protein in the human entorhinal cortex during aging and in Alzheimer’s disease. Proteomics 2003, 3, 1476–1485. [Google Scholar] [CrossRef]
- Han, X.; Zhang, T.; Liu, H.; Mi, Y.; Gou, X. Astrocyte Senescence and Alzheimer’s Disease: A Review. Front. Aging Neurosci. 2020, 12, 148. [Google Scholar] [CrossRef]
- Morita, M.; Ikeshima-Kataoka, H.; Kreft, M.; Vardjan, N.; Zorec, R.; Noda, M. Metabolic Plasticity of Astrocytes and Aging of the Brain. Int. J. Mol. Sci. 2019, 20, 941. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Tonosaki, K.; Kawase, T.; Karasawa, N.; Nagatsu, I.; Fujita, M.; Onozuka, M. Evidence for involvement of dysfunctional teeth in the senile process in the hippocampus of SAMP8 mice. Exp. Gerontol. 2001, 36, 283–295. [Google Scholar] [CrossRef]
- Yamamoto, T.; Hirayama, A. Effects of soft-diet feeding on synaptic density in the hippocampus and parietal cortex of senescence-accelerated mice. Brain Res. 2001, 902, 255–263. [Google Scholar] [CrossRef]
- Kubo, K.Y.; Iwaku, F.; Watanabe, K.; Fujita, M.; Onozuka, M. Molarless-induced changes of spines in hippocampal region of SAMP8 mice. Brain Res. 2005, 1057, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Sofroniew, M.V. Astrocyte Reactivity: Subtypes, States, and Functions in CNS Innate Immunity. Trends Immunol. 2020, 41, 758–770. [Google Scholar] [CrossRef] [PubMed]
- Fanselow, M.S.; Dong, H.W. Are the dorsal and ventral hippocampus functionally distinct structures? Neuron 2010, 65, 7–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, W.B.; Cho, J.H. Synaptic Targeting of Double-Projecting Ventral CA1 Hippocampal Neurons to the Medial Prefrontal Cortex and Basal Amygdala. J. Neurosci. 2017, 37, 4868–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahlstrom, K.L.; Huff, M.L.; Emmons, E.B.; Freeman, J.H.; Narayanan, N.S.; McIntyre, C.K.; LaLumiere, R.T. Basolateral Amygdala Inputs to the Medial Entorhinal Cortex Selectively Modulate the Consolidation of Spatial and Contextual Learning. J. Neurosci. 2018, 38, 2698–2712. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.B.; Cho, J.H. Encoding of contextual fear memory in hippocampal-amygdala circuit. Nat. Commun. 2020, 11, 1382. [Google Scholar] [CrossRef] [PubMed]
- Jarzebowski, P.; Hay, Y.A.; Grewe, B.F.; Paulsen, O. Different encoding of reward location in dorsal and intermediate hippocampus. Curr. Biol. 2022, 32, 834–841.e835. [Google Scholar] [CrossRef]
- Papatheodoropoulos, C. Striking differences in synaptic facilitation along the dorsoventral axis of the hippocampus. Neuroscience 2015, 301, 454–470. [Google Scholar] [CrossRef]
- Dubovyk, V.; Manahan-Vaughan, D. Gradient of Expression of Dopamine D2 Receptors Along the Dorso-Ventral Axis of the Hippocampus. Front. Synaptic Neurosci. 2019, 11, 28. [Google Scholar] [CrossRef] [Green Version]
- Trompoukis, G.; Papatheodoropoulos, C. Dorsal-Ventral Differences in Modulation of Synaptic Transmission in the Hippocampus. Front. Synaptic Neurosci. 2020, 12, 24. [Google Scholar] [CrossRef]
- Chawla, M.K.; Sutherland, V.L.; Olson, K.; McNaughton, B.L.; Barnes, C.A. Behavior-driven arc expression is reduced in all ventral hippocampal subfields compared to CA1, CA3, and dentate gyrus in rat dorsal hippocampus. Hippocampus 2018, 28, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Frey, S.; Schieweck, R.; Forné, I.; Imhof, A.; Straub, T.; Popper, B.; Kiebler, M.A. Physical Activity Dynamically Regulates the Hippocampal Proteome along the Dorso-Ventral Axis. Int. J. Mol. Sci. 2020, 21, 3501. [Google Scholar] [CrossRef] [PubMed]
- Bondi, H.; Bortolotto, V.; Canonico, P.L.; Grilli, M. Complex and regional-specific changes in the morphological complexity of GFAP. Neurobiol. Aging 2021, 100, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Padilla-Coreano, N.; Bolkan, S.S.; Pierce, G.M.; Blackman, D.R.; Hardin, W.D.; Garcia-Garcia, A.L.; Spellman, T.J.; Gordon, J.A. Direct Ventral Hippocampal-Prefrontal Input Is Required for Anxiety-Related Neural Activity and Behavior. Neuron 2016, 89, 857–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parfitt, G.M.; Nguyen, R.; Bang, J.Y.; Aqrabawi, A.J.; Tran, M.M.; Seo, D.K.; Richards, B.A.; Kim, J.C. Bidirectional Control of Anxiety-Related Behaviors in Mice: Role of Inputs Arising from the Ventral Hippocampus to the Lateral Septum and Medial Prefrontal Cortex. Neuropsychopharmacology 2017, 42, 1715–1728. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.; D’Ambra, A.F.; Jung, S.J.; Teratani-Ota, Y.; Vishwakarma, N.; Venkatesh, R.; Parigi, A.; Antzoulatos, E.G.; Fioravante, D.; Wiltgen, B.J. High-Frequency Stimulation of Ventral CA1 Neurons Reduces Amygdala Activity and Inhibits Fear. Front. Behav. Neurosci. 2021, 15, 595049. [Google Scholar] [CrossRef]
- Hauser, J.; Llano López, L.H.; Feldon, J.; Gargiulo, P.A.; Yee, B.K. Small lesions of the dorsal or ventral hippocampus subregions are associated with distinct impairments in working memory and reference memory retrieval, and combining them attenuates the acquisition rate of spatial reference memory. Hippocampus 2020, 30, 938–957. [Google Scholar] [CrossRef]
- Gage, F.H. Adult neurogenesis in neurological diseases. Science 2021, 374, 1049–1050. [Google Scholar] [CrossRef]
- Terreros-Roncal, J.; Moreno-Jiménez, E.P.; Flor-García, M.; Rodríguez-Moreno, C.B.; Trinchero, M.F.; Cafini, F.; Rábano, A.; Llorens-Martín, M. Impact of neurodegenerative diseases on human adult hippocampal neurogenesis. Science 2021, 374, 1106–1113. [Google Scholar] [CrossRef]
- Gomes-Leal, W. Adult Hippocampal Neurogenesis and Affective Disorders: New Neurons for Psychic Well-Being. Front. Neurosci. 2021, 15, 594448. [Google Scholar] [CrossRef]
- Takahashi, T.; Amano, N.; Asamura, H.; Nomiyama, T.; Hanihara, T.; Nakayama, J.; Fukushima, H. Correlation between glial fibrillary acidic protein-positive astrocytes and age in the human hippocampus. Leg. Med. 2006, 8, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Jinno, S. Regional and laminar differences in antigen profiles and spatial distributions of astrocytes in the mouse hippocampus, with reference to aging. Neuroscience 2011, 180, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.J.; Yeh, C.Y.; Terzieva, S.; Olabarria, M.; Kulijewicz-Nawrot, M.; Verkhratsky, A. Complex and region-specific changes in astroglial markers in the aging brain. Neurobiol. Aging 2014, 35, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Arellano, J.J.; Parpura, V.; Zorec, R.; Verkhratsky, A. Astrocytes in physiological aging and Alzheimer’s disease. Neuroscience 2016, 323, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Verkhratsky, A.; Ho, M.S.; Vardjan, N.; Zorec, R.; Parpura, V. General Pathophysiology of Astroglia. Adv. Exp. Med. Biol. 2019, 1175, 149–179. [Google Scholar] [CrossRef]
- Sampedro-Piquero, P.; De Bartolo, P.; Petrosini, L.; Zancada-Menendez, C.; Arias, J.L.; Begega, A. Astrocytic plasticity as a possible mediator of the cognitive improvements after environmental enrichment in aged rats. Neurobiol. Learn Mem. 2014, 114, 16–25. [Google Scholar] [CrossRef]
- Ward, J. Hierarchical Grouping to Optimize an Objective Function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siqueira Mendes, F.d.C.C.d.; Almeida, M.N.F.d.; Falsoni, M.; Andrade, M.L.F.; Felício, A.P.G.; Paixão, L.T.V.B.d.; Júnior, F.L.d.A.; Anthony, D.C.; Brites, D.; Diniz, C.W.P.; et al. The Sedentary Lifestyle and Masticatory Dysfunction: Time to Review the Contribution to Age-Associated Cognitive Decline and Astrocyte Morphotypes in the Dentate Gyrus. Int. J. Mol. Sci. 2022, 23, 6342. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23116342
Siqueira Mendes FdCCd, Almeida MNFd, Falsoni M, Andrade MLF, Felício APG, Paixão LTVBd, Júnior FLdA, Anthony DC, Brites D, Diniz CWP, et al. The Sedentary Lifestyle and Masticatory Dysfunction: Time to Review the Contribution to Age-Associated Cognitive Decline and Astrocyte Morphotypes in the Dentate Gyrus. International Journal of Molecular Sciences. 2022; 23(11):6342. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23116342
Chicago/Turabian StyleSiqueira Mendes, Fabíola de Carvalho Chaves de, Marina Negrão Frota de Almeida, Manoela Falsoni, Marcia Lorena Ferreira Andrade, André Pinheiro Gurgel Felício, Luisa Taynah Vasconcelos Barbosa da Paixão, Fábio Leite do Amaral Júnior, Daniel Clive Anthony, Dora Brites, Cristovam Wanderley Picanço Diniz, and et al. 2022. "The Sedentary Lifestyle and Masticatory Dysfunction: Time to Review the Contribution to Age-Associated Cognitive Decline and Astrocyte Morphotypes in the Dentate Gyrus" International Journal of Molecular Sciences 23, no. 11: 6342. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23116342