
Impact of N-Terminal Tags on De Novo Vimentin Intermediate Filament Assembly


 and
and
Abstract
:1. Introduction
2. Results
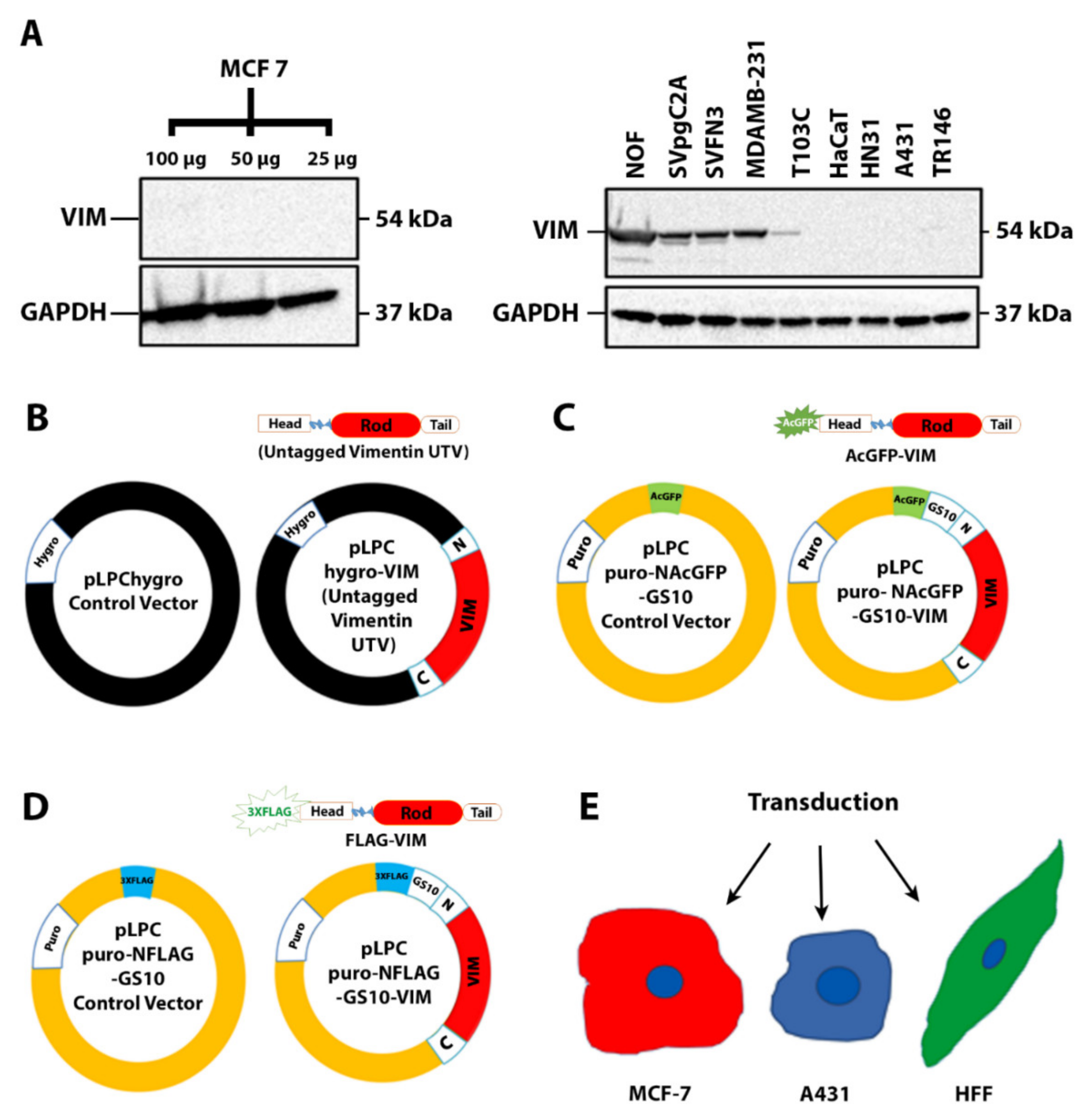
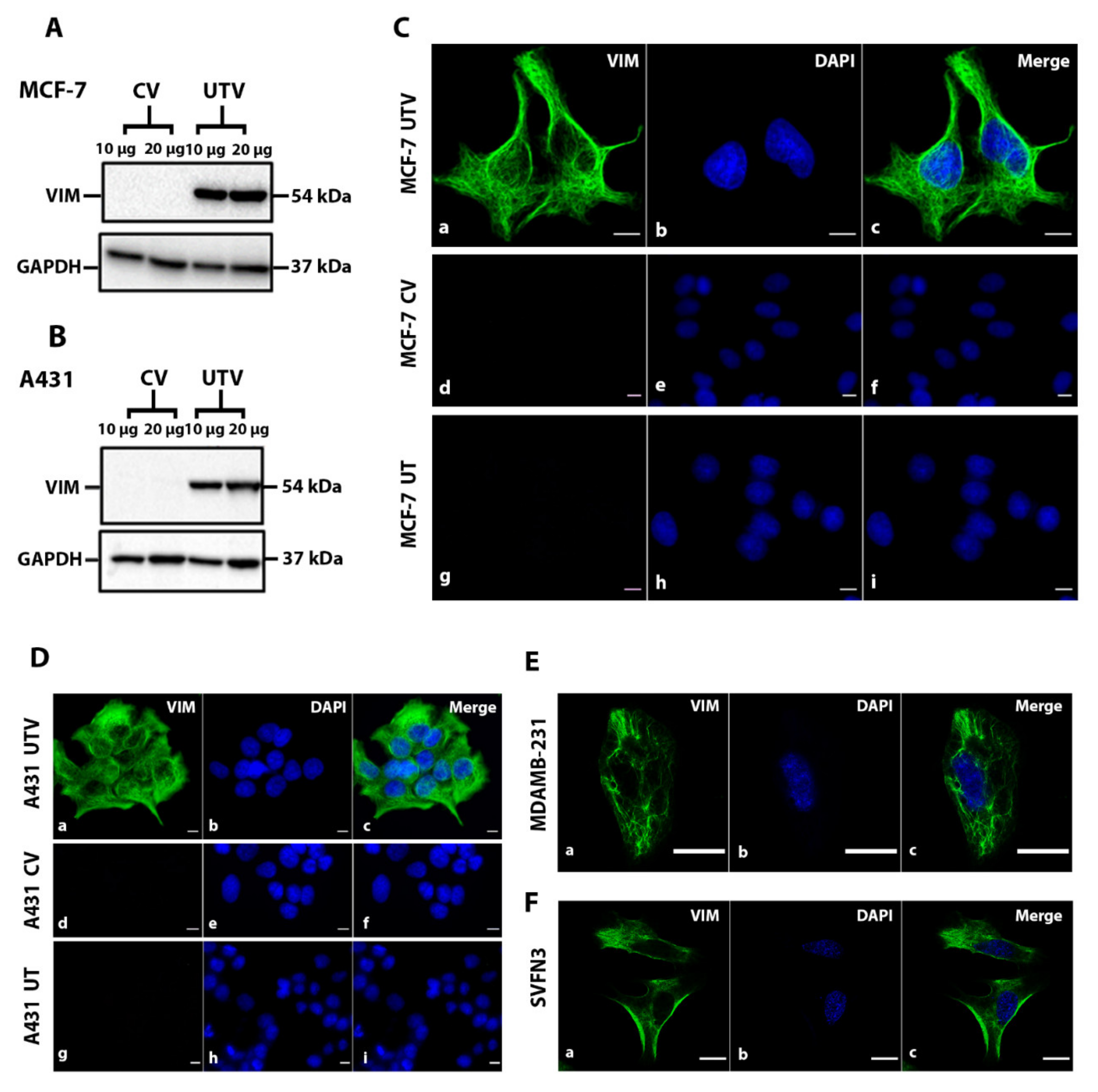
2.1. Filament Formation by Ectopically Expressed Untagged Vimentin (UTV) in the Absence of Endogenous Vimentin
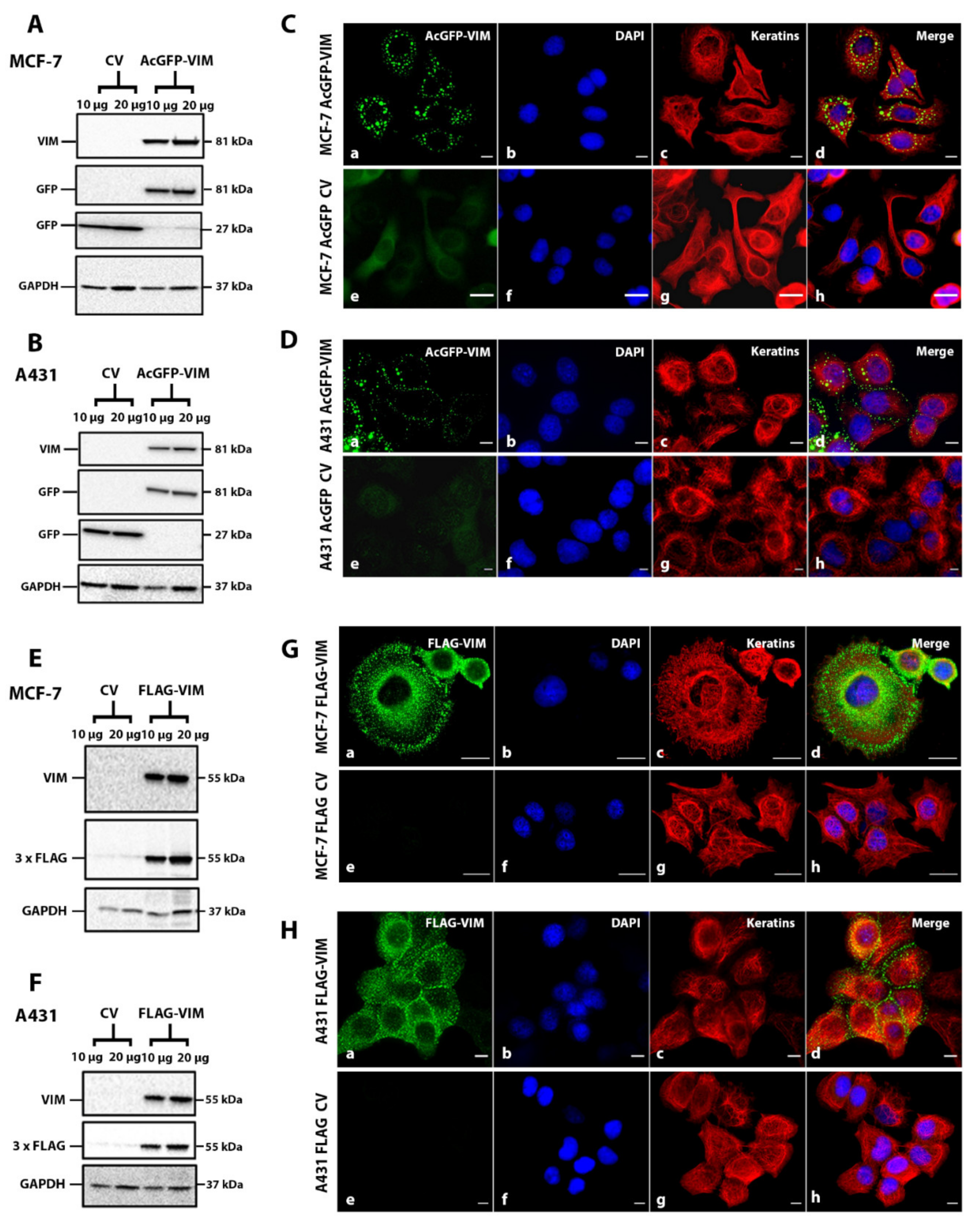
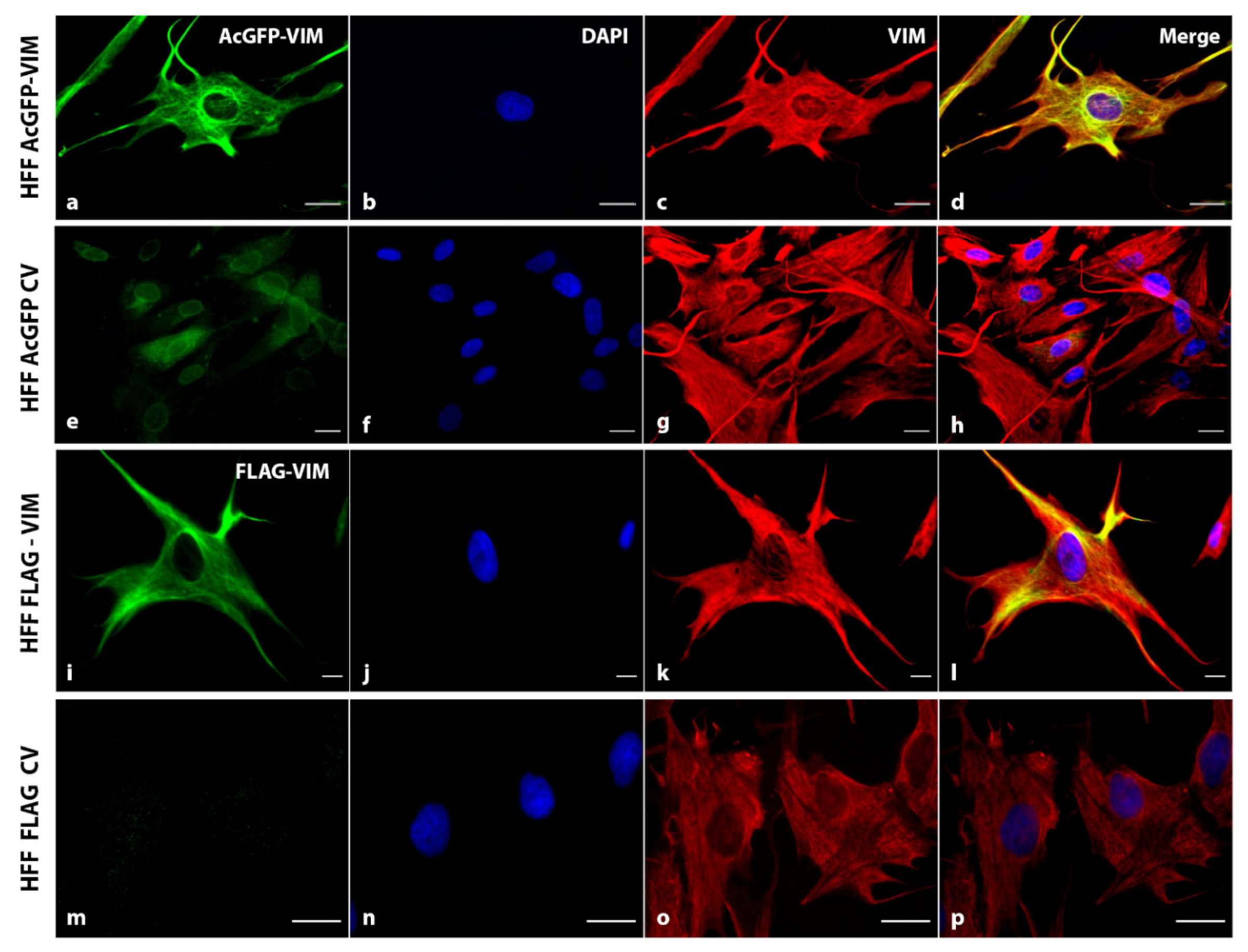
2.2. Ectopic Expression of N-Terminally Tagged Vimentin in Cells Devoid of Endogenous Vimentin
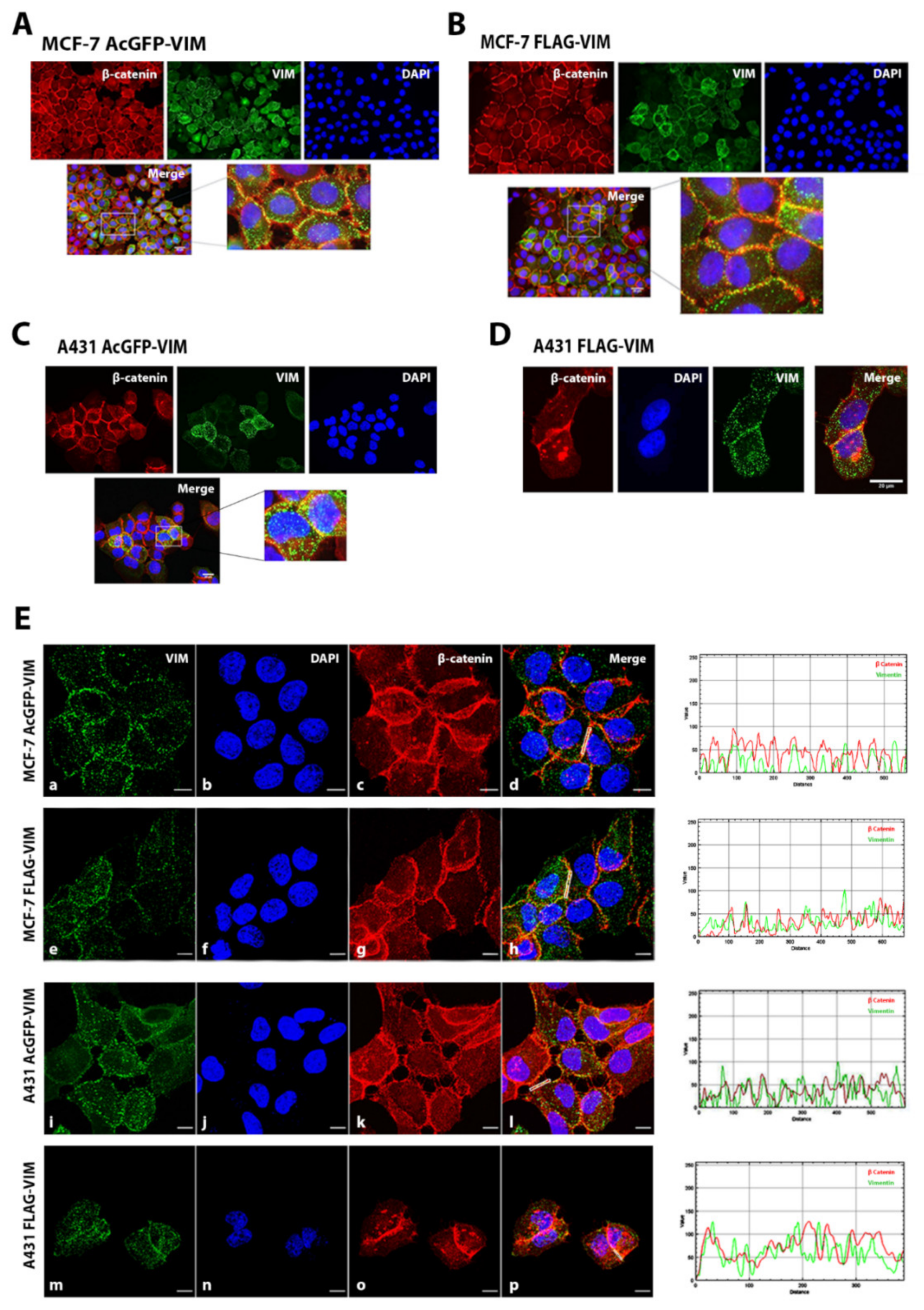
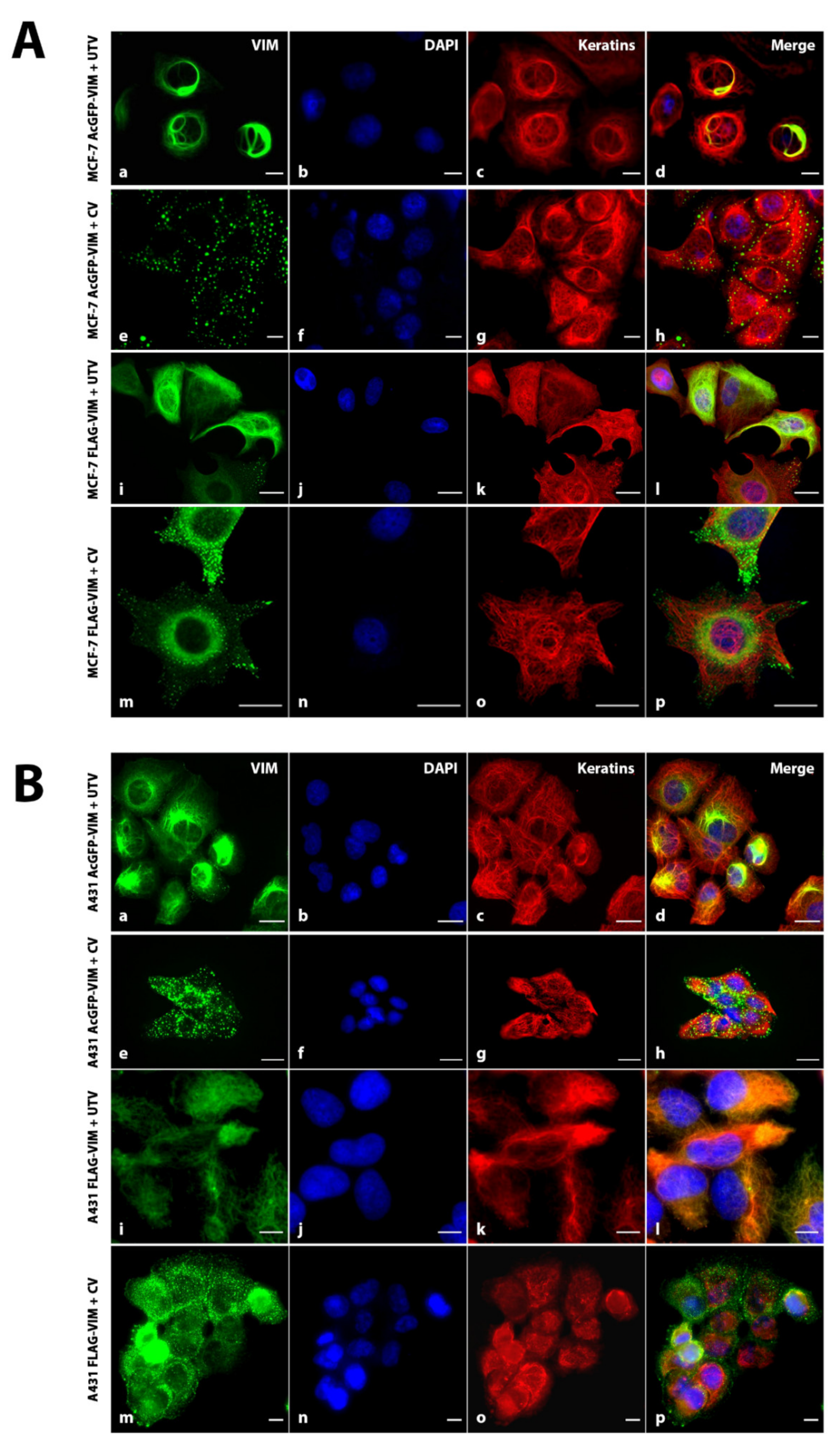
2.3. Co-Alignment of N-Terminally Tagged Vimentin Aggregates with β-Catenin
2.4. N-Terminally Tagged Vimentin Did Not Act as Dominant Negative
2.5. Aggregation of N-Terminally Tagged Vimentin Was Reversible
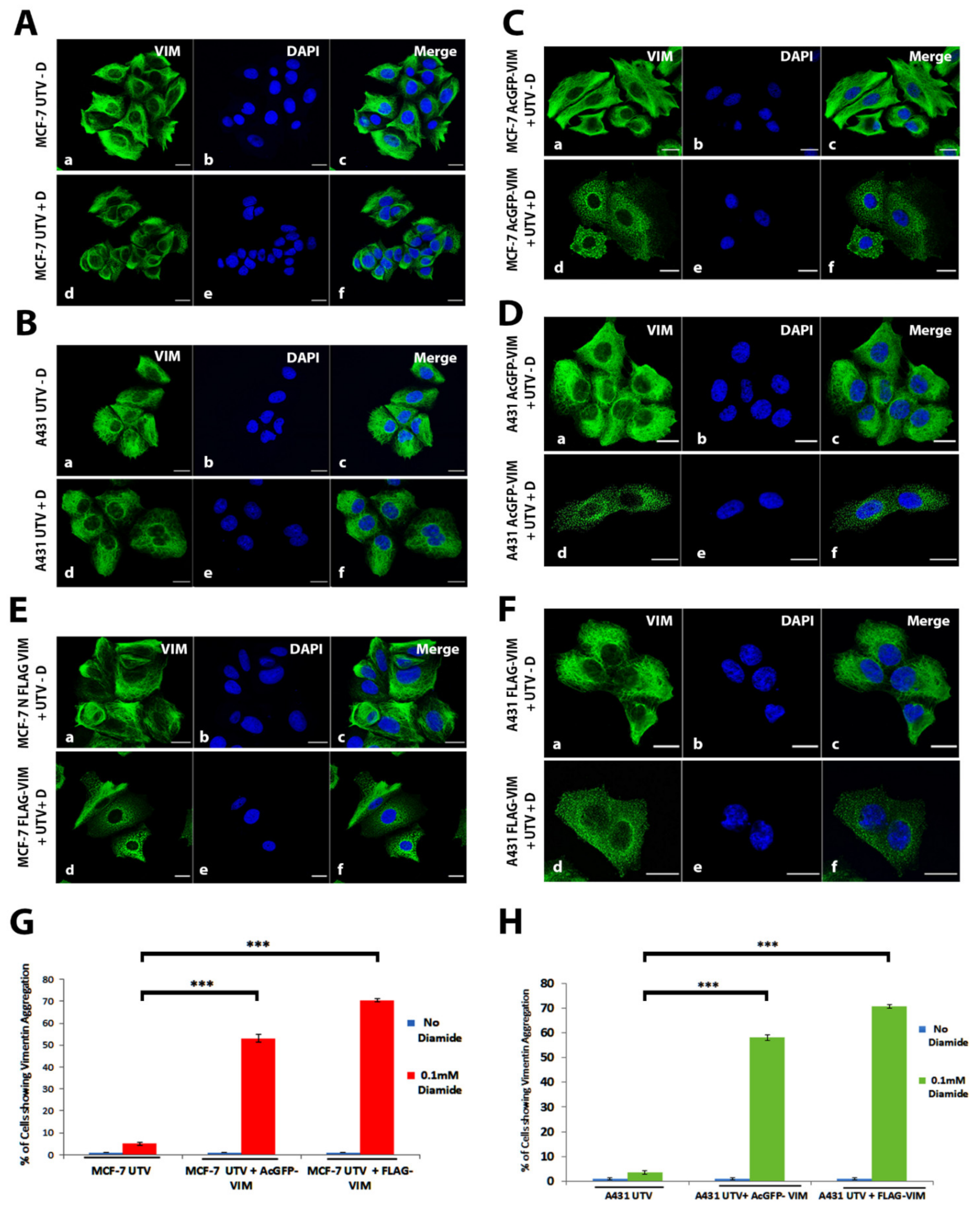
2.6. Response of Ectopically Expressed Vimentin Filament Network to Oxidative Stress
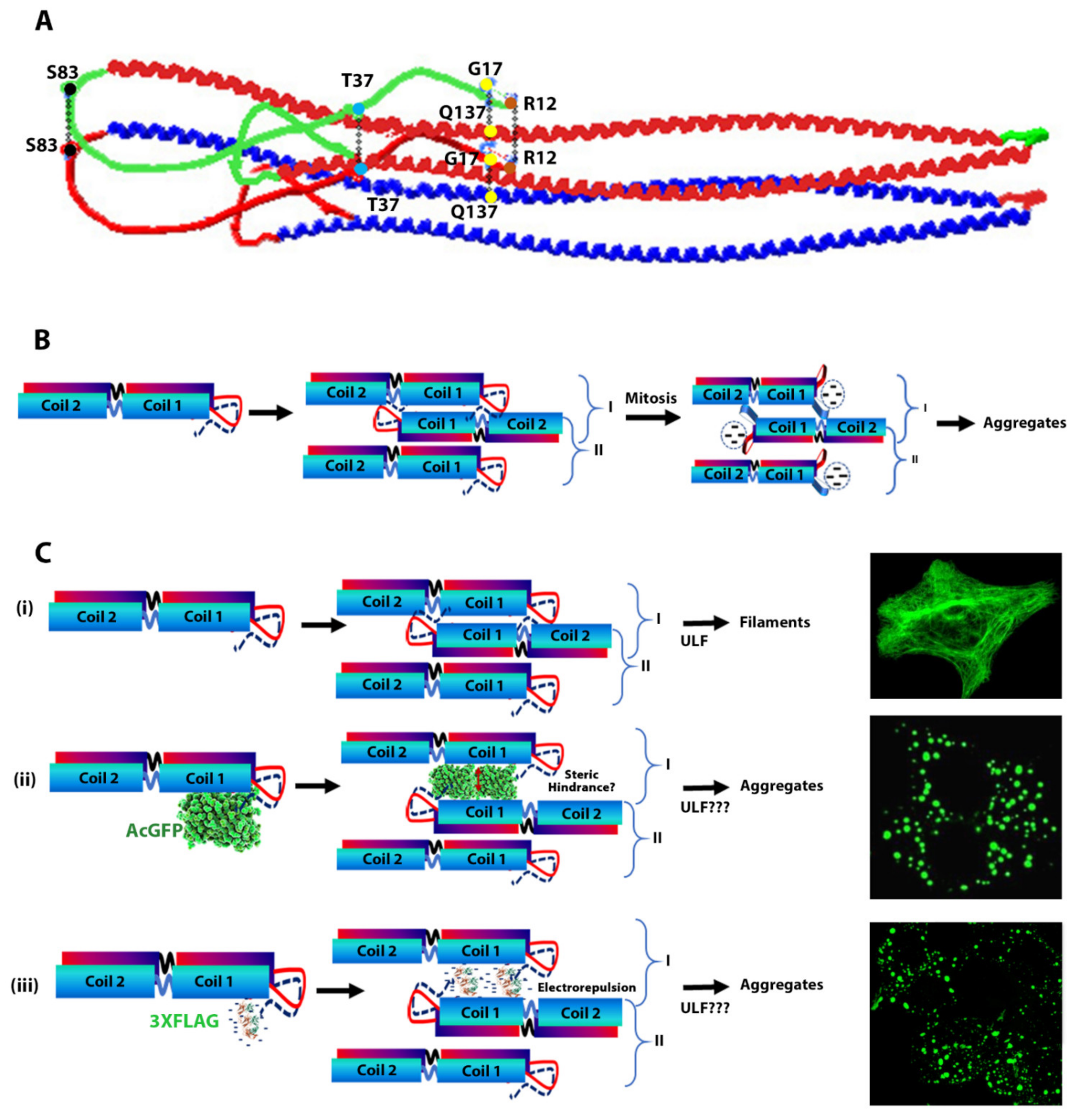
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatments
4.2. Protein Extraction and Western Blotting
4.3. Plasmid Constructs and cDNA Cloning
- pLPChygro-VIM (untagged vimentin, UTV) and pLPChygro (control vector, CV), (Figure 1B).
- pLPCpuro-NAcGFP-GS10-VIM (AcGFP tagged at the N-terminus of full-length vimentin, AcGFP-VIM) and pLPCpuro-NAcGFP-GS10 (AcGFP) (Figure 1C)
- pLPCpuro-NFLAG-GS10-VIM (3 × FLAG tagged at the N-terminus of full-length vimentin, FLAG-VIM) and pLPCpuro-NFLAG-GS10 (FLAG) (Figure 1D).
4.4. Recombinant Retrovirus Production
4.5. Transduction by Spinfection in Cultured Cells
4.6. Immunofluorescence Staining
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Satelli, A.; Li, S. Vimentin in cancer and its potential as a molecular target for cancer therapy. Cell. Mol. Life Sci. 2011, 68, 3033–3046. [Google Scholar] [CrossRef] [Green Version]
- Pollard, T.D.; Goldman, R.D. Overview of the Cytoskeleton from an Evolutionary Perspective. Cold Spring Harb. Perspect. Biol. 2018, 10, a030288. [Google Scholar] [CrossRef] [PubMed]
- Danielsson, F.; Peterson, M.K.; Caldeira Araújo, H.; Lautenschläger, F.; Gad, A.K. Vimentin Diversity in Health and Disease. Cells 2018, 7, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nekrasova, O.E.; Mendez, M.G.; Chernoivanenko, I.S.; Tyurin-Kuzmin, P.A.; Kuczmarski, E.R.; Gelfand, V.I.; Goldman, R.D.; Minin, A.A. Vimentin intermediate filaments modulate the motility of mitochondria. Mol. Biol. Cell 2011, 22, 2282–2289. [Google Scholar] [CrossRef]
- Schwarz, N.; Leube, R.E. Intermediate Filaments as Organizers of Cellular Space: How They Affect Mitochondrial Structure and Function. Cells 2016, 5, 30. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Kang, H.; Zou, F. Low Concentration of GA Activates a Preconditioning Response in HepG2 Cells during Oxidative Stress: Roles of Hsp90 and Vimentin. Cell Stress Chaperones 2009, 14, 381–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, T.J.; Lai, Y.K. Transient increase in vimentin phosphorylation and vimentin-HSC70 association in 9L rat brain tumor cells experiencing heat-shock. J. Cell Biochem. 1994, 54, 100–109. [Google Scholar] [CrossRef]
- Lee, J.-S.; Zhang, M.-H.; Yun, E.K.; Geum, D.; Kim, K.; Kim, T.-H.; Lim, Y.S.; Seo, J.-S. Heat shock protein 27 interacts with vimentin and prevents insolubilization of vimentin subunits induced by cadmium. Exp. Mol. Med. 2005, 37, 427–435. [Google Scholar] [CrossRef] [Green Version]
- Mónico, A.; Duarte, S.; Pajares, M.A.; Pérez-Sala, D. Vimentin disruption by lipoxidation and electrophiles: Role of the cysteine residue and filament dynamics. Redox Biol. 2019, 23, 101098. [Google Scholar] [CrossRef]
- Mendez, M.G.; Kojima, S.-I.; Goldman, R.D. Vimentin induces changes in cell shape, motility, and adhesion during the epithelial to mesenchymal transition. FASEB J. 2010, 24, 1838–1851. [Google Scholar] [CrossRef] [Green Version]
- Usman, S.; Waseem, N.H.; Nguyen, T.K.N.; Mohsin, S.; Jamal, A.; Teh, M.T.; Waseem, A. Vimentin Is at the Heart of Epithelial Mesenchymal Transition (EMT) Mediated Metastasis. Cancers 2021, 13, 4985. [Google Scholar] [CrossRef] [PubMed]
- Challa, A.A.; Stefanovic, B. A novel role of vimentin filaments: Binding and stabilization of collagen mRNAs. Mol. Cell. Biol. 2011, 31, 3773–3789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrmann, H.; Häner, M.; Brettel, M.; Müller, S.A.; Goldie, K.N.; Fedtke, B.; Lustig, A.; Franke, W.W.; Aebi, U. Structure and Assembly Properties of the Intermediate Filament Protein Vimentin: The Role of its Head, Rod and Tail Domains. J. Mol. Biol. 1996, 264, 933–953. [Google Scholar] [CrossRef]
- Tang, D.D. Intermediate filaments in smooth muscle. Am. J. Physiol.-Cell Physiol. 2008, 294, C869–C878. [Google Scholar] [CrossRef] [Green Version]
- Aziz, A.; Hess, J.F.; Budamagunta, M.S.; Voss, J.C.; Kuzin, A.P.; Huang, Y.J.; Xiao, R.; Montelione, G.T.; FitzGerald, P.G.; Hunt, J.F. The Structure of Vimentin Linker 1 and Rod 1B Domains Characterized by Site-directed Spin-labeling Electron Paramagnetic Resonance (SDSL-EPR) and X-ray Crystallography. J. Biol. Chem. 2012, 287, 28349–28361. [Google Scholar] [CrossRef] [Green Version]
- Vermeire, P.J.; Stalmans, G.; Lilina, A.V.; Fiala, J.; Novak, P.; Herrmann, H.; Strelkov, S.V. Molecular Interactions Driving Intermediate Filament Assembly. Cells 2021, 10, 2457. [Google Scholar] [CrossRef] [PubMed]
- Strelkov, S.V.; Herrmann, H.; Geisler, N.; Wedig, T.; Zimbelmann, R.; Aebi, U.; Burkhard, P. Conserved segments 1A and 2B of the intermediate filament dimer: Their atomic structures and role in filament assembly. EMBO J. 2002, 21, 1255–1266. [Google Scholar] [CrossRef] [Green Version]
- Hess, J.F.; Budamagunta, M.S.; Voss, J.C.; FitzGerald, P.G. Structural Characterization of Human Vimentin Rod 1 and the Sequencing of Assembly Steps in Intermediate Filament Formation in Vitro Using Site-directed Spin Labeling and Electron Paramagnetic Resonance. J. Biol. Chem. 2004, 279, 44841–44846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chernyatina, A.A.; Guzenko, D.; Strelkov, S.V. Intermediate filament structure: The bottom-up approach. Curr. Opin. Cell Biol. 2015, 32, 65–72. [Google Scholar] [CrossRef]
- Steinert, P.M.; Marekov, L.N.; Parry, D.A. Diversity of intermediate filament structure. Evidence that the alignment of coiled-coil molecules in vimentin is different from that in keratin intermediate filaments. J. Biol. Chem. 1993, 268, 24916–24925. [Google Scholar] [CrossRef]
- Rogers, K.R.; Herrmann, H.; Franke, W.W. Characterization of disulfide crosslink formation of human vimentin at the dimer, tetramer, and intermediate filament levels. J. Struct. Biol. 1996, 117, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Mücke, N.; Kämmerer, L.; Winheim, S.; Kirmse, R.; Krieger, J.; Mildenberger, M.; Baßler, J.; Hurt, E.; Goldmann, W.H.; Aebi, U.; et al. Assembly Kinetics of Vimentin Tetramers to Unit-Length Filaments: A Stopped-Flow Study. Biophys. J. 2018, 114, 2408–2418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mucke, N.; Wedig, T.; Burer, A.; Marekov, L.N.; Steinert, P.M.; Langowski, J.; Aebi, U.; Herrmann, H. Molecular and biophysical characterization of assembly-starter units of human vimentin. J. Mol. Biol. 2004, 340, 97–114. [Google Scholar] [CrossRef]
- Georgakopoulou, S.; Moller, D.; Sachs, N.; Herrmann, H.; Aebi, U. Near-UV circular dichroism reveals structural transitions of vimentin subunits during intermediate filament assembly. J. Mol. Biol. 2009, 386, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, H.; Aebi, U. Intermediate Filaments: Structure and Assembly. Cold Spring Harb. Perspect. Biol. 2016, 8, a018242. [Google Scholar] [CrossRef]
- Cary, R.B.; Klymkowsky, M.W.; Evans, R.M.; Domingo, A.; Dent, J.A.; Backhus, L.E. Vimentin's tail interacts with actin-containing structures in vivo. J. Cell Sci. 1994, 107 Pt 6, 1609–1622. [Google Scholar] [CrossRef]
- Esue, O.; Carson, A.A.; Tseng, Y.; Wirtz, D. A direct interaction between actin and vimentin filaments mediated by the tail domain of vimentin. J. Biol. Chem. 2006, 281, 30393–30399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, S.; Viedma-Poyatos, A.; Navarro-Carrasco, E.; Martinez, A.E.; Pajares, M.A.; Perez-Sala, D. Vimentin filaments interact with the actin cortex in mitosis allowing normal cell division. Nat. Commun. 2019, 10, 4200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helfand, B.T.; Mikami, A.; Vallee, R.B.; Goldman, R.D. A requirement for cytoplasmic dynein and dynactin in intermediate filament network assembly and organization. J. Cell Biol. 2002, 157, 795–806. [Google Scholar] [CrossRef] [PubMed]
- Prahlad, V.; Yoon, M.; Moir, R.D.; Vale, R.D.; Goldman, R.D. Rapid movements of vimentin on microtubule tracks: Kinesin-dependent assembly of intermediate filament networks. J. Cell Biol. 1998, 143, 159–170. [Google Scholar] [CrossRef] [Green Version]
- Traub, P.; Scherbarth, A.; Wiegers, W.; Shoeman, R.L. Salt-stable interaction of the amino-terminal head region of vimentin with the alpha-helical rod domain of cytoplasmic intermediate filament proteins and its relevance to protofilament structure and filament formation and stability. J. Cell Sci. 1992, 101 Pt 2, 363–381. [Google Scholar] [CrossRef] [PubMed]
- Shoeman, R.L.; Hartig, R.; Berthel, M.; Traub, P. Deletion Mutagenesis of the Amino-Terminal Head Domain of Vimentin Reveals Dispensability of Large Internal Regions for Intermediate Filament Assembly and Stability. Exp. Cell Res. 2002, 279, 344–353. [Google Scholar] [CrossRef]
- Aziz, A.; Hess, J.F.; Budamagunta, M.S.; Voss, J.C.; FitzGerald, P.G. Site-directed spin labeling and electron paramagnetic resonance determination of vimentin head domain structure. J. Biol. Chem. 2010, 285, 15278–15285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowrie, D.J.; Stickney, J.T.; Ip, W. Properties of the Nonhelical End Domains of Vimentin Suggest a Role in Maintaining Intermediate Filament Network Structure. J. Struct. Biol. 2000, 132, 83–94. [Google Scholar] [CrossRef]
- Chernyatina, A.A.; Nicolet, S.; Aebi, U.; Herrmann, H.; Strelkov, S.V. Atomic structure of the vimentin central α-helical domain and its implications for intermediate filament assembly. Proc. Natl. Acad. Sci. USA 2012, 109, 13620–13625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martys, J.L.; Ho, C.L.; Liem, R.K.; Gundersen, G.G. Intermediate filaments in motion: Observations of intermediate filaments in cells using green fluorescent protein-vimentin. Mol. Biol. Cell 1999, 10, 1289–1295. [Google Scholar] [CrossRef]
- Thorn, K. Genetically encoded fluorescent tags. Mol. Biol. Cell 2017, 28, 848–857. [Google Scholar] [CrossRef] [PubMed]
- Ridge, K.M.; Shumaker, D.; Robert, A.; Hookway, C.; Gelfand, V.I.; Janmey, P.A.; Lowery, J.; Guo, M.; Weitz, D.A.; Kuczmarski, E.; et al. Chapter Fourteen—Methods for Determining the Cellular Functions of Vimentin Intermediate Filaments. In Methods in Enzymology; Omary, M.B., Liem, R.K.H., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 568, pp. 389–426. [Google Scholar]
- Ho, C.L.; Martys, J.L.; Mikhailov, A.; Gundersen, G.G.; Liem, R.K. Novel features of intermediate filament dynamics revealed by green fluorescent protein chimeras. J. Cell Sci. 1998, 111 Pt 13, 1767–1778. [Google Scholar] [CrossRef] [PubMed]
- Yoon, M.; Moir, R.D.; Prahlad, V.; Goldman, R.D. Motile Properties of Vimentin Intermediate Filament Networks in Living Cells. J. Cell Biol. 1998, 143, 147–157. [Google Scholar] [CrossRef] [Green Version]
- McCormick, M.B.; Kouklis, P.; Syder, A.; Fuchs, E. The roles of the rod end and the tail in vimentin IF assembly and IF network formation. J. Cell Biol. 1993, 122, 395–407. [Google Scholar] [CrossRef] [Green Version]
- Aziz, A.; Hess, J.F.; Budamagunta, M.S.; FitzGerald, P.G.; Voss, J.C. Head and Rod 1 Interactions in Vimentin. J. Biol. Chem. 2009, 284, 7330–7338. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Sala, D.; Oeste, C.L.; Martínez, A.E.; Carrasco, M.J.; Garzón, B.; Cañada, F.J. Vimentin filament organization and stress sensing depend on its single cysteine residue and zinc binding. Nat. Commun. 2015, 6, 7287. [Google Scholar] [CrossRef] [Green Version]
- Kural, C.; Serpinskaya, A.S.; Chou, Y.H.; Goldman, R.D.; Gelfand, V.I.; Selvin, P.R. Tracking melanosomes inside a cell to study molecular motors and their interaction. Proc. Natl. Acad. Sci. USA 2007, 104, 5378–5382. [Google Scholar] [CrossRef] [Green Version]
- Serres, M.P.; Samwer, M.; Truong Quang, B.A.; Lavoie, G.; Perera, U.; Görlich, D.; Charras, G.; Petronczki, M.; Roux, P.P.; Paluch, E.K. F-Actin Interactome Reveals Vimentin as a Key Regulator of Actin Organization and Cell Mechanics in Mitosis. Dev. Cell 2020, 52, 210–222.e217. [Google Scholar] [CrossRef] [Green Version]
- Amraei, R.; Xia, C.; Olejnik, J.; White, M.R.; Napoleon, M.A.; Lotfollahzadeh, S.; Hauser, B.M.; Schmidt, A.G.; Chitalia, V.; Mühlberger, E.; et al. Extracellular vimentin is an attachment factor that facilitates SARS-CoV-2 entry into human endothelial cells. Proc. Natl. Acad. Sci. USA 2022, 119, e2113874119. [Google Scholar] [CrossRef]
- Pattabiraman, S.; Azad, G.K.; Amen, T.; Brielle, S.; Park, J.E.; Sze, S.K.; Meshorer, E.; Kaganovich, D. Vimentin protects differentiating stem cells from stress. Sci. Rep. 2020, 10, 19525. [Google Scholar] [CrossRef]
- Song, S.; Hanson, M.J.; Liu, B.-F.; Chylack, L.T.; Liang, J.J.N. Protein-protein interactions between lens vimentin and alphaB-crystallin using FRET acceptor photobleaching. Mol. Vis. 2008, 14, 1282–1287. [Google Scholar]
- Cogli, L.; Progida, C.; Bramato, R.; Bucci, C. Vimentin phosphorylation and assembly are regulated by the small GTPase Rab7a. Biochim. Biophys. Acta 2013, 1833, 1283–1293. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Nakamoto, T.; Ogawa, S.; Seo, S.; Matsumura, T.; Tachibana, K.; Morimoto, C.; Hirai, H. MICAL, a Novel CasL Interacting Molecule, Associates with Vimentin. J. Biol. Chem. 2002, 277, 14933–14941. [Google Scholar] [CrossRef] [Green Version]
- Makise, M.; Nakamura, H.; Kuniyasu, A. The role of vimentin in the tumor marker Nup88-dependent multinucleated phenotype. BMC Cancer 2018, 18, 519. [Google Scholar] [CrossRef]
- Shabbir, S.H.; Cleland, M.M.; Goldman, R.D.; Mrksich, M. Geometric control of vimentin intermediate filaments. Biomaterials 2014, 35, 1359–1366. [Google Scholar] [CrossRef] [Green Version]
- You, H.; Yuan, D.; Bi, Y.; Zhang, N.; Li, Q.; Tu, T.; Wei, X.; Lian, Q.; Yu, T.; Kong, D.; et al. Hepatitis B virus X protein promotes vimentin expression via LIM and SH3 domain protein 1 to facilitate epithelial-mesenchymal transition and hepatocarcinogenesis. Cell Commun. Signal. 2021, 19, 33. [Google Scholar] [CrossRef] [PubMed]
- Virant, D.; Traenkle, B.; Maier, J.; Kaiser, P.D.; Bodenhöfer, M.; Schmees, C.; Vojnovic, I.; Pisak-Lukáts, B.; Endesfelder, U.; Rothbauer, U. A peptide tag-specific nanobody enables high-quality labeling for dSTORM imaging. Nat. Commun. 2018, 9, 930. [Google Scholar] [CrossRef]
- Vicente, F.N.; Lelek, M.; Tinevez, J.-Y.; Tran, Q.D.; Pehau-Arnaudet, G.; Zimmer, C.; Etienne-Manneville, S.; Giannone, G.; Leduc, C. Molecular organization and mechanics of single vimentin filaments revealed by super-resolution imaging. HAL Open Sci. 2021, hal-03366936, 1–35. [Google Scholar]
- Zeng, F.; Beck, V.; Schuierer, S.; Garnier, I.; Manneville, C.; Agarinis, C.; Morelli, L.; Quinn, L.; Knehr, J.; Roma, G.; et al. A Simple and Efficient CRISPR Technique for Protein Tagging. Cells 2020, 9, 2618. [Google Scholar] [CrossRef]
- Kimple, M.E.; Brill, A.L.; Pasker, R.L. Overview of affinity tags for protein purification. Curr. Protoc. Protein. Sci. 2013, 73, 9.9.1–9.9.23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.Y.; Chang, P.W.; Hsu, W.H.; Chang, H.C.; Chen, C.L.; Lai, C.C.; Chiu, W.T.; Chen, H.C. Src and SHP2 coordinately regulate the dynamics and organization of vimentin filaments during cell migration. Oncogene 2019, 38, 4075–4094. [Google Scholar] [CrossRef] [Green Version]
- Hao, F.; Liu, J.; Zhong, M.; Wang, J.; Liu, J. Expression of E-cadherin, vimentin and β-catenin in ameloblastoma and association with clinicopathological characteristics of ameloblastoma. Int. J. Clin. Exp. Pathol. 2018, 11, 199–207. [Google Scholar]
- Gilles, C.; Polette, M.; Mestdagt, M.l.; Nawrocki-Raby, B.a.; Ruggeri, P.; Birembaut, P.; Foidart, J.-M. Transactivation of Vimentin by β-Catenin in Human Breast Cancer Cells1. Cancer Res. 2003, 63, 2658–2664. [Google Scholar]
- Ivaska, J.; Pallari, H.M.; Nevo, J.; Eriksson, J.E. Novel functions of vimentin in cell adhesion, migration, and signaling. Exp. Cell Res. 2007, 313, 2050–2062. [Google Scholar] [CrossRef] [PubMed]
- Hendrix, M.J.; Seftor, E.A.; Seftor, R.E.; Trevor, K.T. Experimental co-expression of vimentin and keratin intermediate filaments in human breast cancer cells results in phenotypic interconversion and increased invasive behavior. Am. J. Pathol. 1997, 150, 483–495. [Google Scholar] [PubMed]
- Liu, C.-Y.; Lin, H.-H.; Tang, M.-J.; Wang, Y.-K. Vimentin contributes to epithelial-mesenchymal transition cancer cell mechanics by mediating cytoskeletal organization and focal adhesion maturation. Oncotarget 2015, 6, 15966–15983. [Google Scholar] [CrossRef] [Green Version]
- Bornheim, R.; Muller, M.; Reuter, U.; Herrmann, H.; Bussow, H.; Magin, T.M. A dominant vimentin mutant upregulates Hsp70 and the activity of the ubiquitin-proteasome system, and causes posterior cataracts in transgenic mice. J. Cell Sci. 2008, 121, 3737–3746. [Google Scholar] [CrossRef] [Green Version]
- Sarria, A.J.; Nordeen, S.K.; Evans, R.M. Regulated expression of vimentin cDNA in cells in the presence and absence of a preexisting vimentin filament network. J. Cell Biol. 1990, 111, 553–565. [Google Scholar] [CrossRef]
- Sarria, A.J.; Lieber, J.G.; Nordeen, S.K.; Evans, R.M. The presence or absence of a vimentin-type intermediate filament network affects the shape of the nucleus in human SW-13 cells. J. Cell Sci. 1994, 107 Pt 6, 1593–1607. [Google Scholar] [CrossRef] [PubMed]
- Raats, J.M.; Pieper, F.R.; Vree Egberts, W.T.; Verrijp, K.N.; Ramaekers, F.C.; Bloemendal, H. Assembly of amino-terminally deleted desmin in vimentin-free cells. J. Cell Biol. 1990, 111, 1971–1985. [Google Scholar] [CrossRef] [Green Version]
- Vassar, R.; Coulombe, P.A.; Degenstein, L.; Albers, K.; Fuchs, E. Mutant keratin expression in transgenic mice causes marked abnormalities resembling a human genetic skin disease. Cell 1991, 64, 365–380. [Google Scholar] [CrossRef]
- Lane, E.B.; Rugg, E.L.; Navsaria, H.; Leigh, I.M.; Heagerty, A.H.; Ishida-Yamamoto, A.; Eady, R.A. A mutation in the conserved helix termination peptide of keratin 5 in hereditary skin blistering. Nature 1992, 356, 244–246. [Google Scholar] [CrossRef] [PubMed]
- Coulombe, P.A.; Hutton, M.E.; Letai, A.; Hebert, A.; Paller, A.S.; Fuchs, E. Point mutations in human keratin 14 genes of epidermolysis bullosa simplex patients: Genetic and functional analyses. Cell 1991, 66, 1301–1311. [Google Scholar] [CrossRef]
- Morley, S.M.; Dundas, S.R.; James, J.L.; Gupta, T.; Brown, R.A.; Sexton, C.J.; Navsaria, H.A.; Leigh, I.M.; Lane, E.B. Temperature sensitivity of the keratin cytoskeleton and delayed spreading of keratinocyte lines derived from EBS patients. J. Cell Sci. 1995, 108 Pt 11, 3463–3471. [Google Scholar] [CrossRef] [PubMed]
- Werner, N.S.; Windoffer, R.; Strnad, P.; Grund, C.; Leube, R.E.; Magin, T.M. Epidermolysis bullosa simplex-type mutations alter the dynamics of the keratin cytoskeleton and reveal a contribution of actin to the transport of keratin subunits. Mol. Biol. Cell 2004, 15, 990–1002. [Google Scholar] [CrossRef] [Green Version]
- Gouveia, M.; Zemljic-Jokhadar, S.; Vidak, M.; Stojkovic, B.; Derganc, J.; Travasso, R.; Liovic, M. Keratin Dynamics and Spatial Distribution in Wild-Type and K14 R125P Mutant Cells-A Computational Model. Int. J. Mol. Sci. 2020, 21, 2596. [Google Scholar] [CrossRef] [Green Version]
- Woll, S.; Windoffer, R.; Leube, R.E. p38 MAPK-dependent shaping of the keratin cytoskeleton in cultured cells. J. Cell Biol. 2007, 177, 795–807. [Google Scholar] [CrossRef] [PubMed]
- Traub, P.; Vorgias, C.E. Involvement of the N-terminal polypeptide of vimentin in the formation of intermediate filaments. J. Cell Sci. 1983, 63, 43–67. [Google Scholar] [CrossRef]
- Shoeman, R.L.; Mothes, E.; Kesselmeier, C.; Traub, P. Intermediate filament assembly and stability in vitro: Effect and implications of the removal of head and tail domains of vimentin by the human immunodeficiency virus type 1 protease. Cell Biol. Int. Rep. 1990, 14, 583–594. [Google Scholar] [CrossRef]
- Beuttenmüller, M.; Chen, M.; Janetzko, A.; Kühn, S.; Traub, P. Structural Elements the Amino-Terminal Head Domain of Vimentin Essential for Intermediate Filament Formation in Vivo and in Vitro. Exp. Cell Res. 1994, 213, 128–142. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, M.; Nishi, Y.; Nishizawa, K.; Matsuyama, M.; Sato, C. Site-specific phosphorylation induces disassembly of vimentin filaments in vitro. Nature 1987, 328, 649–652. [Google Scholar] [CrossRef]
- Inagaki, M.; Takahara, H.; Nishi, Y.; Sugawara, K.; Sato, C. Ca2+-dependent deimination-induced disassembly of intermediate filaments involves specific modification of the amino-terminal head domain. J. Biol. Chem. 1989, 264, 18119–18127. [Google Scholar] [CrossRef]
- Hofmann, I.; Herrmann, H. Interference in vimentin assembly in vitro by synthetic peptides derived from the vimentin head domain. J. Cell Sci. 1992, 101 Pt 3, 687–700. [Google Scholar] [CrossRef]
- Yasui, Y.; Goto, H.; Matsui, S.; Manser, E.; Lim, L.; Nagata, K.; Inagaki, M. Protein kinases required for segregation of vimentin filaments in mitotic process. Oncogene 2001, 20, 2868–2876. [Google Scholar] [CrossRef] [Green Version]
- Goto, H.; Yasui, Y.; Kawajiri, A.; Nigg, E.A.; Terada, Y.; Tatsuka, M.; Nagata, K.; Inagaki, M. Aurora-B regulates the cleavage furrow-specific vimentin phosphorylation in the cytokinetic process. J. Biol. Chem. 2003, 278, 8526–8530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almazan, G.; Afar, D.E.; Bell, J.C. Phosphorylation and disruption of intermediate filament proteins in oligodendrocyte precursor cultures treated with calyculin A. J. Neurosci. Res. 1993, 36, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Tzivion, G.; Luo, Z.J.; Avruch, J. Calyculin A-induced vimentin phosphorylation sequesters 14-3-3 and displaces other 14-3-3 partners in vivo. J. Biol. Chem. 2000, 275, 29772–29778. [Google Scholar] [CrossRef] [Green Version]
- Chou, Y.H.; Bischoff, J.R.; Beach, D.; Goldman, R.D. Intermediate filament reorganization during mitosis is mediated by p34cdc2 phosphorylation of vimentin. Cell 1990, 62, 1063–1071. [Google Scholar] [CrossRef]
- Eckert, C. Flag-Tag and 3x Flag-Tag: An Epitope Tag for Capture and Detection Experiments. Available online: https://resources.chromotek.com/blog/flag-tag-and-3x-flag-tag-an-epitope-tag-for-capture-and-detection-experiments (accessed on 15 February 2022).
- Soule, H.D.; Vazquez, J.; Long, A.; Albert, S.; Brennan, M. A Human Cell Line from a Pleural Effusion Derived From a Breast Carcinoma2. JNCI J. Natl. Cancer Inst. 1973, 51, 1409–1416. [Google Scholar] [CrossRef] [PubMed]
- Giard, D.J.; Aaronson, S.A.; Todaro, G.J.; Arnstein, P.; Kersey, J.H.; Dosik, H.; Parks, W.P. In vitro cultivation of human tumors: Establishment of cell lines derived from a series of solid tumors. J. Natl. Cancer Inst. 1973, 51, 1417–1423. [Google Scholar] [CrossRef]
- Eiselleova, L.; Peterková, I.; Neradil, J.; Slaninová, I.; Hampl, A.; DvpYák, P. Comparative study of mouse and human feeder cells for human embryonic stem cells. Int. J. Dev. Biol. 2008, 52, 353–363. [Google Scholar] [CrossRef]
- Cailleau, R.; Young, R.; Olivé, M.; Reeves, W.J., Jr. Breast Tumor Cell Lines From Pleural Effusions2. JNCI J. Natl. Cancer Inst. 1974, 53, 661–674. [Google Scholar] [CrossRef] [PubMed]
- Gemenetzidis, E.; Bose, A.; Riaz, A.M.; Chaplin, T.; Young, B.D.; Ali, M.; Sugden, D.; Thurlow, J.K.; Cheong, S.-C.; Teo, S.-H.; et al. FOXM1 Upregulation Is an Early Event in Human Squamous Cell Carcinoma and it Is Enhanced by Nicotine during Malignant Transformation. PLoS ONE 2009, 4, e4849. [Google Scholar] [CrossRef]
- Rheinwald, J.G.; Beckett, M.A. Defective terminal differentiation in culture as a consistent and selectable character of malignant human keratinocytes. Cell 1980, 22, 629–632. [Google Scholar] [CrossRef]
- Kulkarnl, P.S.; Sundqvist, K.; Betsholtz, C.; Höglund, P.; Wiman, K.G.; Zhivotovsky, B.; Bertolero, F.; Liu, Y.; Grafström, R.C. Characterization of human buccal epithelial cells transfected with the simian virus 40 T-antigen gene. Carcinogenesis 1995, 16, 2515–2521. [Google Scholar] [CrossRef]
- Bryan, D.; Sexton, C.J.; Williams, D.; Leigh, I.M.; McKay, I.A. Oral keratinocytes immortalized with the early region of human papillomavirus type 16 show elevated expression of interleukin 6, which acts as an autocrine growth factor for the derived T103C cell line. Cell Growth Differ 1995, 6, 1245–1250. [Google Scholar] [PubMed]
- Boukamp, P.; Petrussevska, R.T.; Breitkreutz, D.; Hornung, J.; Markham, A.; Fusenig, N.E. Normal keratinization in a spontaneously immortalized aneuploid human keratinocyte cell line. J. Cell Biol. 1988, 106, 761–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singchat, W.; Hitakomate, E.; Rerkarmnuaychoke, B.; Suntronpong, A.; Fu, B.; Bodhisuwan, W.; Peyachoknagul, S.; Yang, F.; Koontongkaew, S.; Srikulnath, K. Genomic Alteration in Head and Neck Squamous Cell Carcinoma (HNSCC) Cell Lines Inferred from Karyotyping, Molecular Cytogenetics, and Array Comparative Genomic Hybridization. PLoS ONE 2016, 11, e0160901. [Google Scholar] [CrossRef]
- Nielsen, H.; Rassing, M. Nicotine permeability across the buccal TR146 cell culture model and porcine buccal mucosa in vitro: Effect of pH and concentration. Eur. J. Pharm. Sci. Off. J. Eur. Fed. Pharm. Sci. 2002, 16, 151–157. [Google Scholar] [CrossRef]
- Pear, W.S.; Nolan, G.P.; Scott, M.L.; Baltimore, D. Production of high-titer helper-free retroviruses by transient transfection. Proc. Natl. Acad. Sci. USA 1993, 90, 8392–8396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, A.D.; Chen, F. Retrovirus packaging cells based on 10A1 murine leukemia virus for production of vectors that use multiple receptors for cell entry. J. Virol. 1996, 70, 5564–5571. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Origin | Reference |
|---|---|---|
| TR146 | Human squamous cell carcinoma | [97] |
| A431 | Epidermoid carcinoma of the vulva | [88] |
| MDA-MB-231 | Invasive and poorly differentiated cell line derived from breast adenocarcinoma. Derived from metastatic site: Pleural effusion | [12] |
| MCF-7 | Epithelial cancer cell line derived from breast adenocarcinoma. Derived from metastatic site: Pleural effusion | [87] |
| SVpgC2a | Simian virus 40 transformed human premalignant buccal keratinocyte | [93] |
| SVFN3 | Malignant buccal keratinocyte derived after treating SVpgC2a with nicotine and over-expression of FOXM1B | [91] |
| HaCaT | Spontaneously immortalized human keratinocyte line | [95] |
| HFF-1 | HPV type 16 E6/E7 immortalised human foreskin fibroblasts | [89] |
| NHOF | Normal human oral keratinocytes | [92] |
| T103C | HPV type 16 E6/E7 immortalized oral keratinocytes | [94] |
| HN31 | Head and neck squamous cell carcinoma line | [96] |
| Antibody | Dilution | Host | Catalogue # | Supplier |
|---|---|---|---|---|
| Anti-vimentin, V9 | IF = 1:700 WB = 1:2000 | Mouse | ab8069 | Abcam |
| Anti-cytokeratin K18 | IF = 1:100 WB = 1:1000 | Rabbit | ab24561 | Abcam |
| Anti-GFP | IF = 1:500 WB = 1:10,000 | Rabbit | ab183734 | Abcam |
| Anti-GAPDH | WB = 1:2000 | Rabbit | Ab9485 | Abcam |
| Anti-FLAG | IF = 1:500 WB = 1:1000 | Rabbit | F7425 | Millipore UK |
| Mouse IgG peroxidase conjugated | WB = 1:1000 | Goat | NA931V | GE Healthcare UK |
| Rabbit IgG peroxidase conjugated | WB = 1:1000 | Donkey | NA934 | GE Healthcare UK |
| A45-B/B3 (anti-pancytokeratin) | IF = Neat hybridoma supernatant | Mouse | In-house, neat hybridoma supernatant | In-house, neat hybridoma supernatant40 |
| Anti-Integrin α6 | IF = 1:100 | Mouse | ab181551 | Abcam |
| Anti-β-catenin | IF = 1:100 | Mouse | 610154 | BD Biosciences |
| Anti-E cadherin | IF = 1:100 | Mouse | ab1416 | Abcam |
| Anti-vinculin | IF = 1:150 | Mouse | V4505 | Sigma-Aldrich |
| Anti-mouse Alexa Fluor® 488 IgG H+L | IF = 1:1000 | Goat | A-11001 | Life Technologies, UK |
| Anti-mouse Alexa Fluor® 594 IgG H+L | IF = 1:1000 | Goat | A-11005 | Molecular Probes, UK |
| Anti-rabbit Alexa Fluor® 488 IgG H+L | IF = 1:1000 | Goat | A-11008 | Life Technologies, UK |
| Anti-rabbit Alexa Fluor® 594 IgG H+L | IF = 1:1000 | Goat | A-11012 | Life Technologies, UK |
| Name of the Transduced Cell Lines Used in This Study | Abbreviated as |
|---|---|
| MCF-7 pLPChygro-VIM (untagged Vimentin) | MCF-7 UTV |
| MCF-7 pLPChygro (control vector) | MCF-7 CV |
| A431 pLPChygro-VIM (untagged Vimentin) | A431 UTV |
| A431 pLPChygro (control vector) | A431 CV |
| MCF-7 pLPCpuro-NAcGFP-GS10-VIM | MCF-7 AcGFP-VIM |
| MCF-7 pLPCpuro-NAcGFP-GS10 | MCF-7 AcGFP CV |
| A431 pLPCpuro-NAcGFP-GS10-VIM | A431 AcGFP-VIM |
| A431 pLPCpuro-NAcGFP-GS10 | A431 AcGFP CV |
| MCF-7 pLPCpuro-NFLAG-GS10-VIM | MCF-7 FLAG-VIM |
| MCF-7 pLPCpuro-NFLAG-GS10 | MCF-7 FLAG CV |
| A431 pLPCpuro-NFLAG-GS10-VIM | A431 FLAG-VIM |
| A431 pLPCpuro-NFLAG-GS10 | A431 FLAG CV |
| HFF-1 pLPCpuro-NAcGFP-GS10-VIM | HFF-1 AcGFP-VIM |
| HFF-1 pLPCpuro-NAcGFP-GS10 | HFF-1 AcGFP CV |
| HFF-1 pLPCpuro-NFLAG-GS10-VIM | HFF-1 FLAG-VIM |
| HFF-1 pLPCpuro-NFLAG-GS10 | HFF-1 FLAG CV |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Usman, S.; Aldehlawi, H.; Nguyen, T.K.N.; Teh, M.-T.; Waseem, A. Impact of N-Terminal Tags on De Novo Vimentin Intermediate Filament Assembly. Int. J. Mol. Sci. 2022, 23, 6349. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23116349
Usman S, Aldehlawi H, Nguyen TKN, Teh M-T, Waseem A. Impact of N-Terminal Tags on De Novo Vimentin Intermediate Filament Assembly. International Journal of Molecular Sciences. 2022; 23(11):6349. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23116349
Chicago/Turabian StyleUsman, Saima, Hebah Aldehlawi, Thuan Khanh Ngoc Nguyen, Muy-Teck Teh, and Ahmad Waseem. 2022. "Impact of N-Terminal Tags on De Novo Vimentin Intermediate Filament Assembly" International Journal of Molecular Sciences 23, no. 11: 6349. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23116349