
Erythrocyte Membrane Nanomechanical Rigidity Is Decreased in Obese Patients †

 , ,
, ,  ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Patient Recruitment and Anthropometry
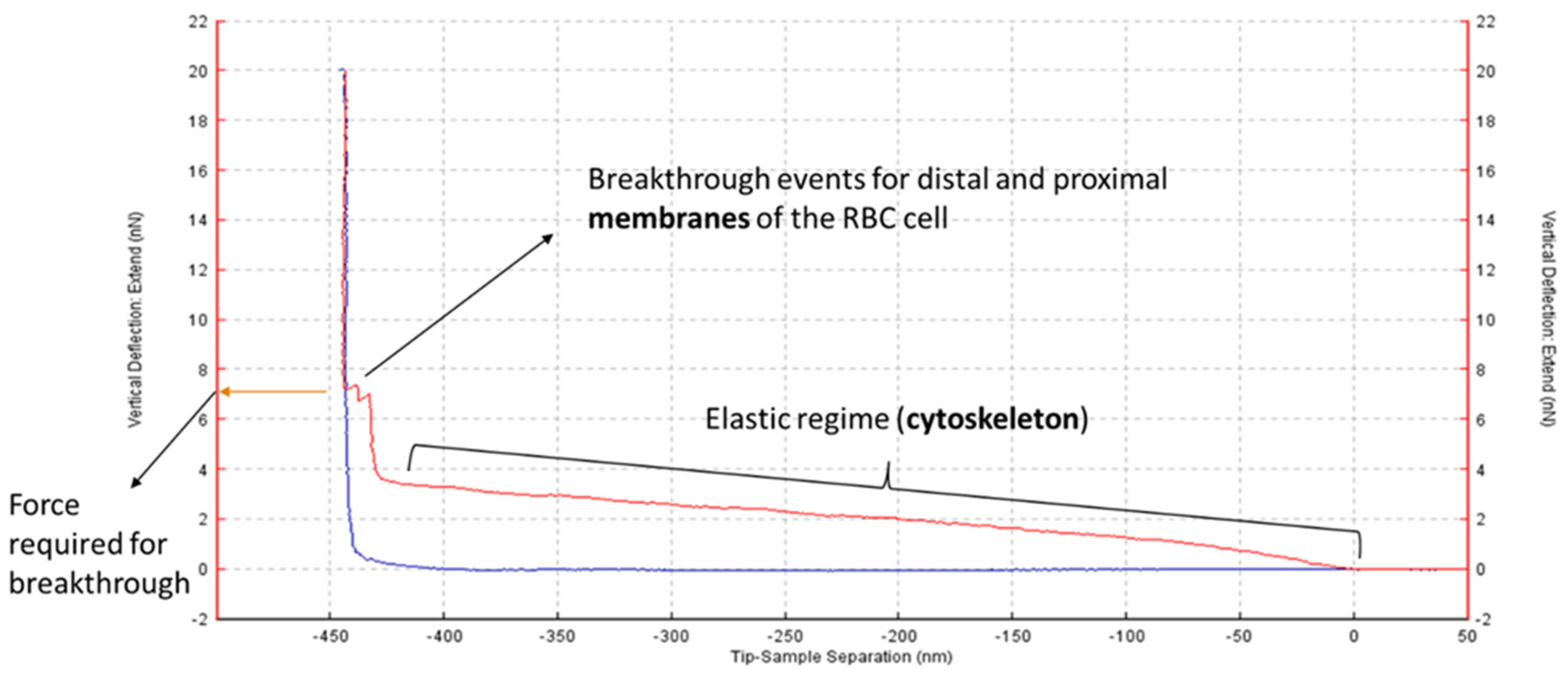
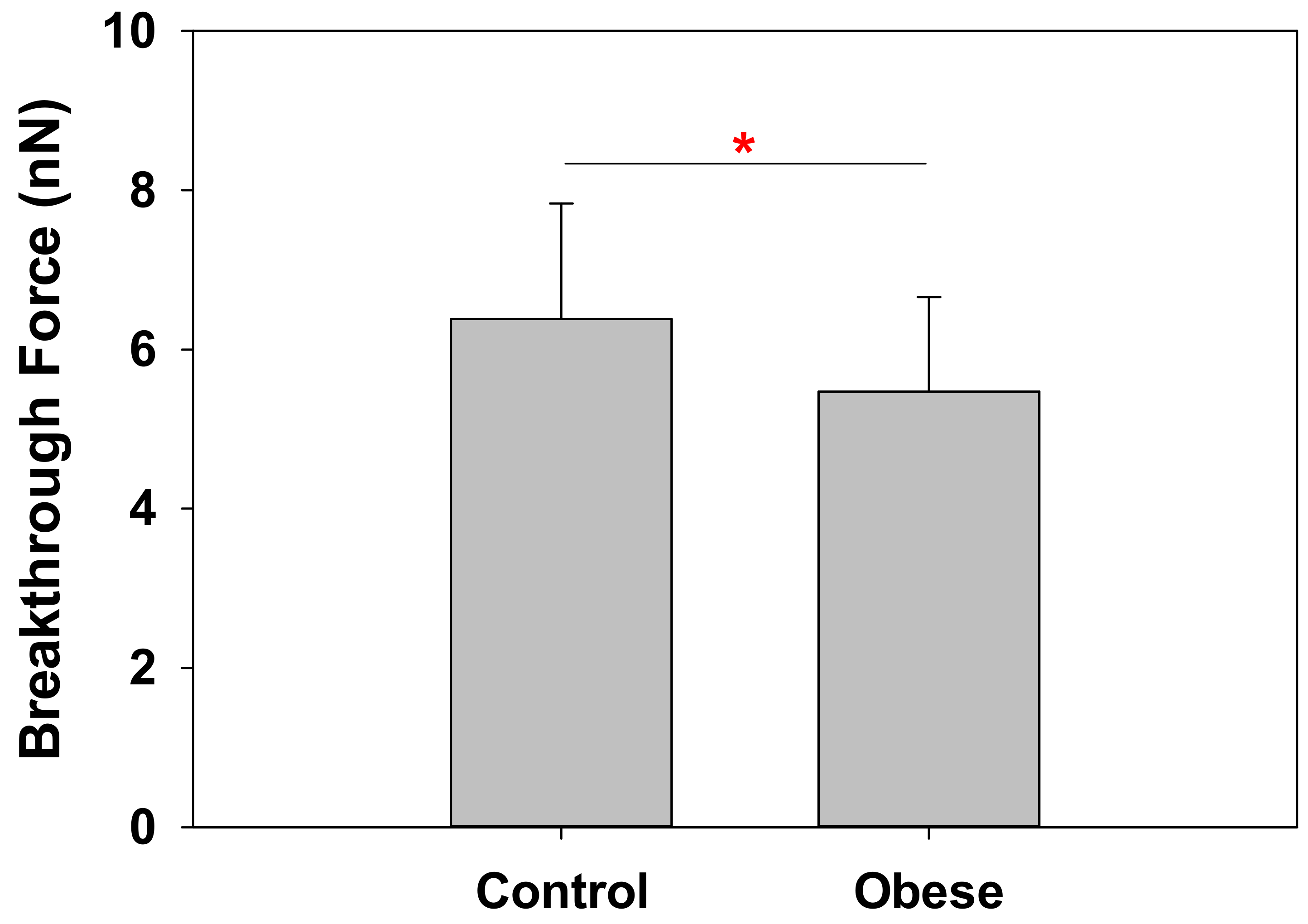
2.2. Obese Patient RBC Membranes Exhibit Lower Nanomechanical Rigidity under the Atomic Force Microscope
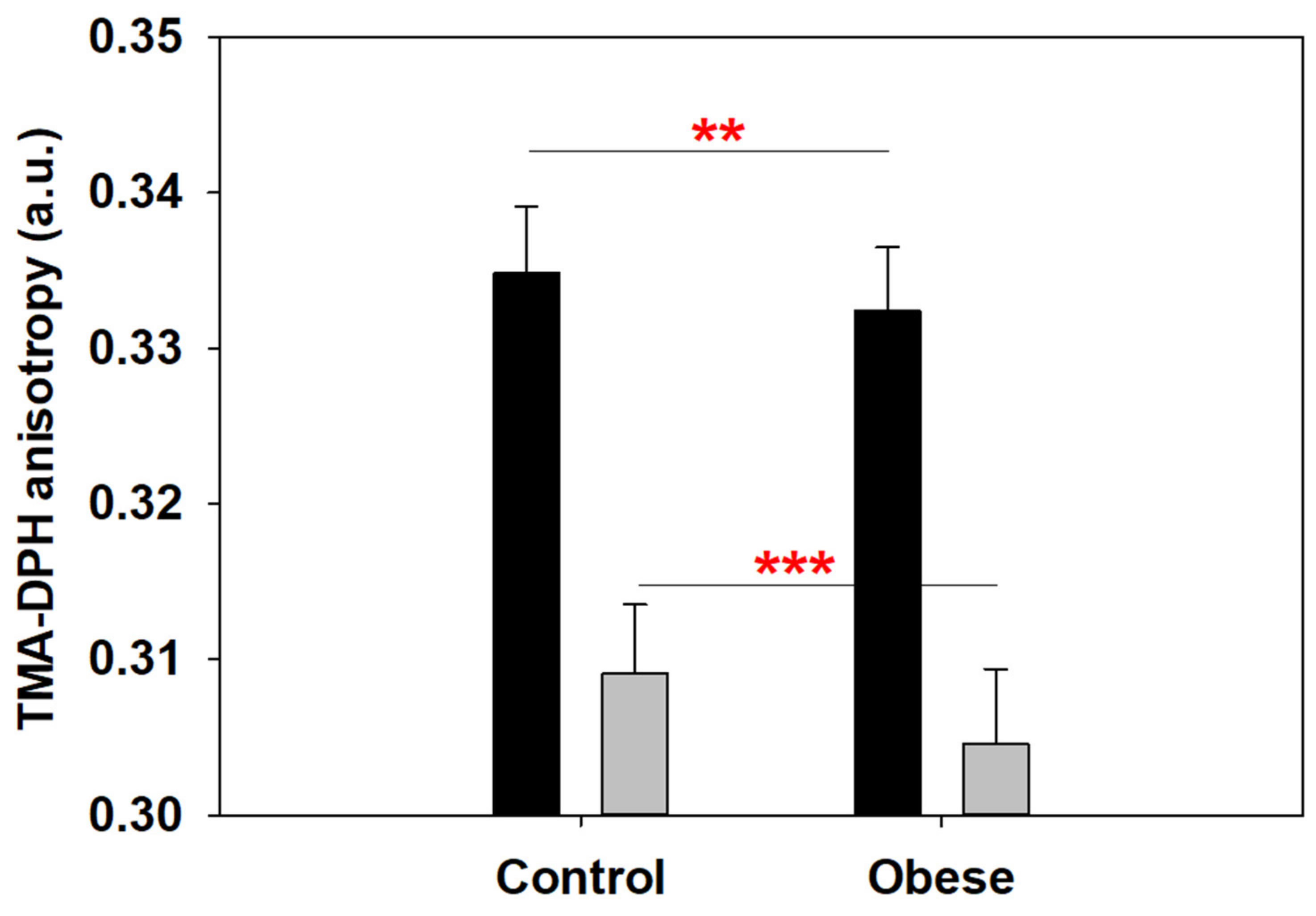
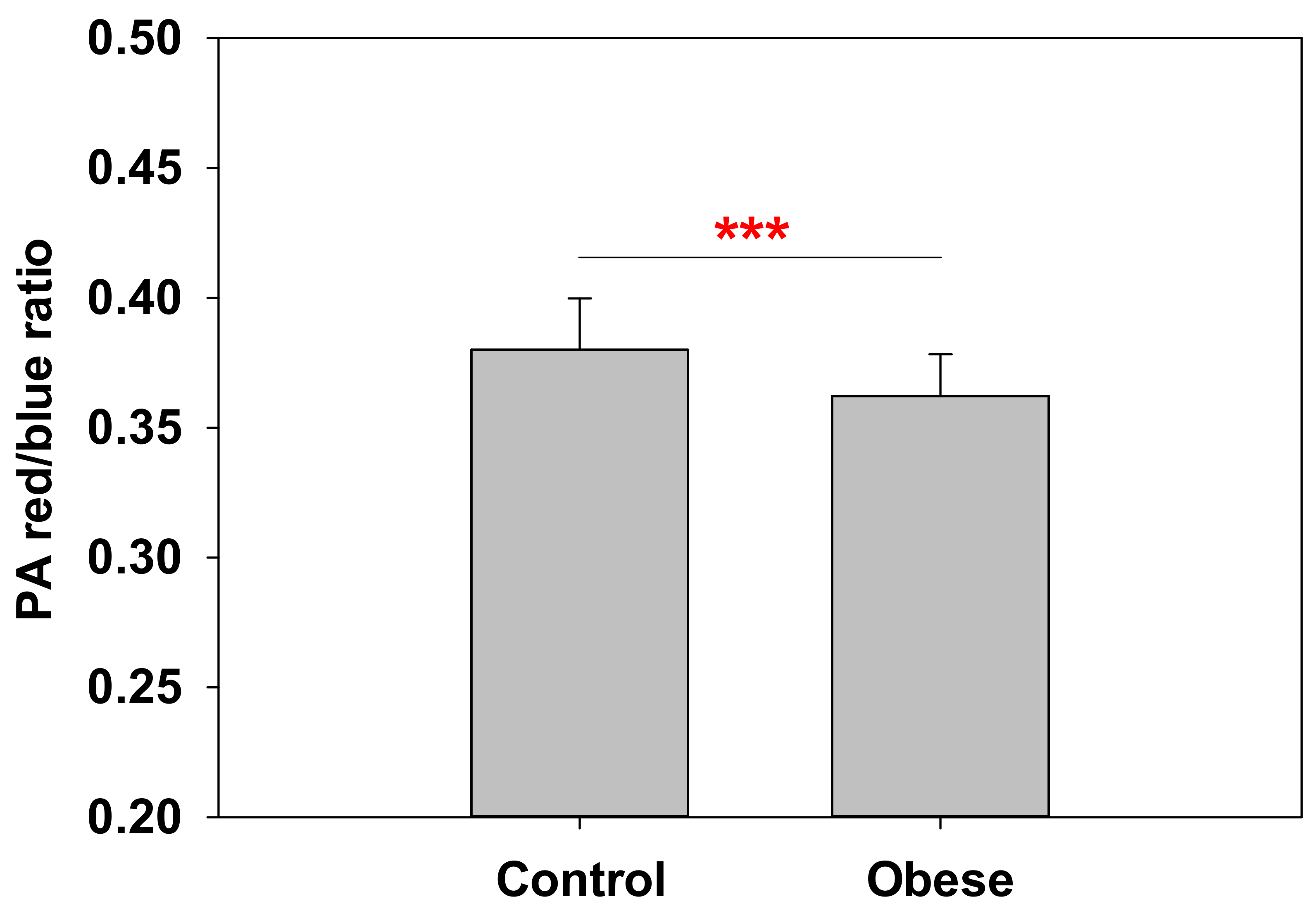
2.3. Anisotropy at the Polar–Nonpolar Interface Is Decreased in Obese Patient Erythrocyte Membranes
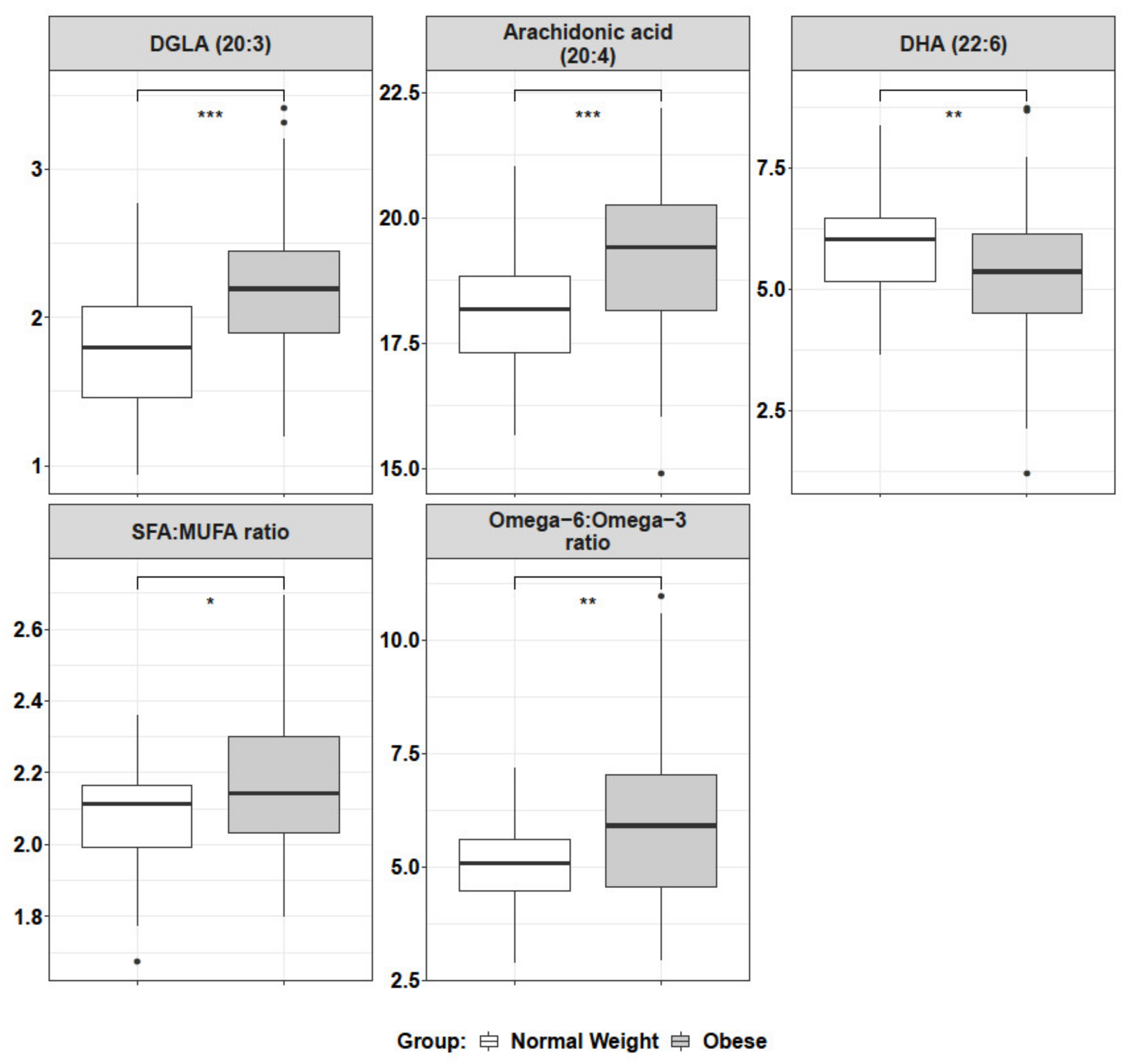
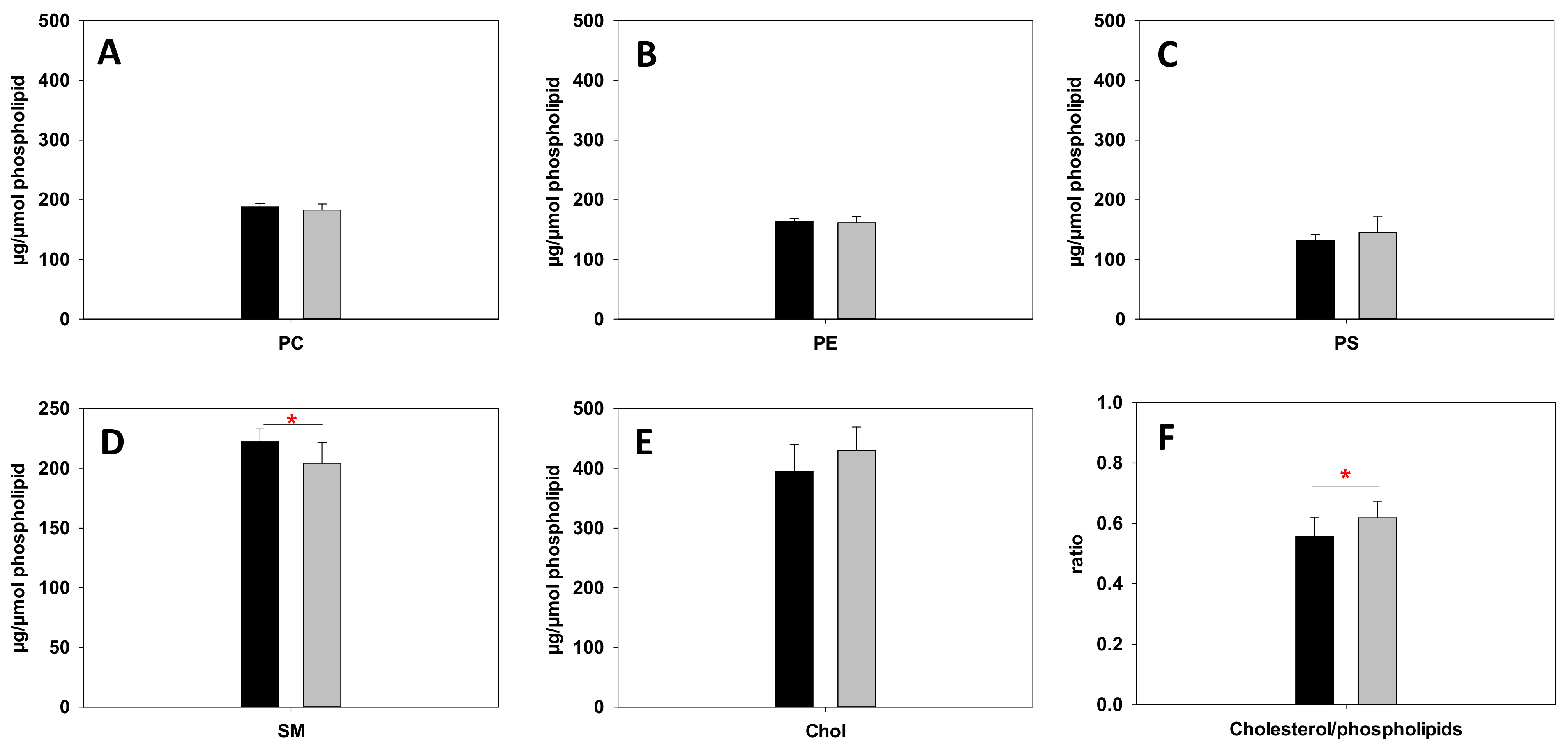
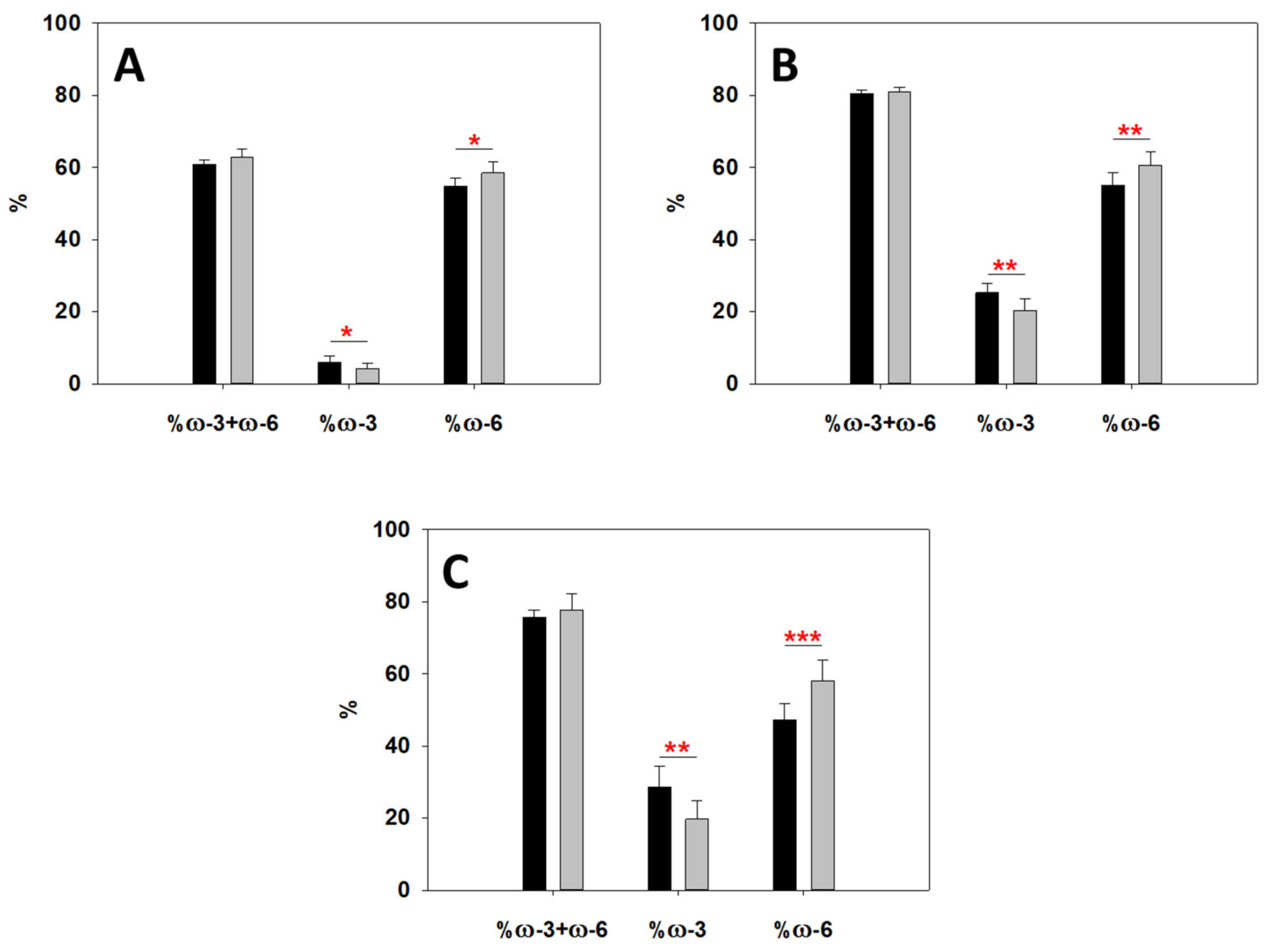
2.4. Lipidomics Reveal an Altered Metabolism of Sphingomyelin (SM), ω-6 and ω-3 Fatty Acids
2.5. Differences in Blood Plasma between Obese Patients and Control Blood Samples
3. Discussion
4. Materials and Methods
4.1. Subjects and Study Design and Anthropometric Measurements
4.2. Blood Management and RBC Isolation
4.3. Fluorescence Anisotropy of TMA-DPH in Erythrocyte Membrane
4.4. Sample Preparation for AFM Measurements
4.5. AFM Force Measurements
4.6. PA Ratio in Blood Plasma Using Fluorescence Spectroscopy
4.7. Mature RBC Lipidomics of Global Fatty Acids
4.8. Mature Red Blood Cell Membrane Fatty Acid Cluster
4.9. RBC Lipidomics of Specific Lipid Species
4.9.1. Sample Preparation
4.9.2. Analysis of the Samples
>4.9.3. Data Processing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bernotienė, G.; Dulskienė, V.; Klumbienė, J.; Kučienė, R.; Medžionienė, J.; Petkevičienė, J.; Petrauskienė, A.; Radišauskas, R.; Tamošiūnas, A.; Vaitkevičiūtė, J. Heterogeneous contributions of change in population distribution of body mass index to change in obesity and underweight. Elife 2021, 10, e60060. [Google Scholar]
- Schwartz, M.W.; Seeley, R.J.; Zeltser, L.M.; Drewnowski, A.; Ravussin, E.; Redman, L.M.; Leibel, R.L. Obesity pathogenesis: An endocrine society scientific statement. Endocr. Rev. 2017, 38, 267–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flier, J.S. Obesity wars: Molecular progress confronts an expanding epidemic. Cell 2004, 116, 337–350. [Google Scholar] [CrossRef] [Green Version]
- Perona, J.S. Membrane lipid alterations in the metabolic syndrome and the role of dietary oils. Biochim. Et Biophys. Acta (BBA) Biomembr. 2017, 1859, 1690–1703. [Google Scholar] [CrossRef]
- Cox, M.M.; Nelson, D.L. Lehninger Principles of Biochemistry; WH Freeman: New York, NY, USA, 2008; Volume 5. [Google Scholar]
- Lecerf, J.-M.; De Lorgeril, M. Dietary cholesterol: From physiology to cardiovascular risk. Br. J. Nutr. 2011, 106, 6–14. [Google Scholar] [CrossRef] [Green Version]
- Ohvo-Rekilä, H.; Ramstedt, B.; Leppimäki, P.; Peter Slotte, J. Cholesterol interactions with phospholipids in membranes. Prog. Lipid Res. 2002, 41, 66–97. [Google Scholar] [CrossRef]
- Goñi, F.M. The basic structure and dynamics of cell membranes: An update of the Singer-Nicolson model. Biochim. Et Biophys. Acta 2014, 1838, 1467–1476. [Google Scholar] [CrossRef] [Green Version]
- Harayama, T.; Riezman, H. Understanding the diversity of membrane lipid composition. Nat. Rev. Mol. Cell Biol. 2018, 19, 281–296. [Google Scholar] [CrossRef]
- Ferreri, C.; Masi, A.; Sansone, A.; Giacometti, G.; Larocca, A.V.; Menounou, G.; Scanferlato, R.; Tortorella, S.; Rota, D.; Conti, M. Fatty acids in membranes as homeostatic, metabolic and nutritional biomarkers: Recent advancements in analytics and diagnostics. Diagnostics 2017, 7, 1. [Google Scholar] [CrossRef]
- Torres, M.; Parets, S.; Fernández-Díaz, J.; Beteta-Göbel, R.; Rodríguez-Lorca, R.; Román, R.; Lladó, V.; Rosselló, C.A.; Fernández-García, P.; Escribá, P.V. Lipids in Pathophysiology and Development of the Membrane Lipid Therapy: New Bioactive Lipids. Membranes 2021, 11, 919. [Google Scholar] [CrossRef]
- Alsharari, Z.D.; Risérus, U.; Leander, K.; Sjögren, P.; Carlsson, A.C.; Vikström, M.; Laguzzi, F.; Gigante, B.; Cederholm, T.; De Faire, U. Serum fatty acids, desaturase activities and abdominal obesity–a population-based study of 60-year old men and women. PLoS ONE 2017, 12, e0170684. [Google Scholar] [CrossRef]
- Del Pozo, M.D.P.; Lope, V.; Criado-Navarro, I.; Pastor-Barriuso, R.; Fernández de Larrea, N.; Ruiz, E.; Castelló, A.; Lucas, P.; Sierra, Á.; Romieu, I. Serum Phospholipid Fatty Acids Levels, Anthropometric Variables and Adiposity in Spanish Premenopausal Women. Nutrients 2020, 12, 1895. [Google Scholar] [CrossRef]
- Vinknes, K.J.; Elshorbagy, A.K.; Drevon, C.A.; Nurk, E.; Tell, G.S.; Nygård, O.; Vollset, S.E.; Refsum, H. Associations between plasma polyunsaturated fatty acids, plasma stearoyl-CoA desaturase indices and body fat. Obesity 2013, 21, E512–E519. [Google Scholar] [CrossRef]
- Pan, D.A.; Hulbert, A.; Storlien, L. Dietary fats, membrane phospholipids and obesity. J. Nutr. 1994, 124, 1555–1565. [Google Scholar] [CrossRef]
- Min, Y.; Ghebremeskel, K.; Lowy, C.; Thomas, B.; Crawford, M. Adverse effect of obesity on red cell membrane arachidonic and docosahexaenoic acids in gestational diabetes. Diabetologia 2004, 47, 75–81. [Google Scholar] [CrossRef] [Green Version]
- Cazzola, R.; Rondanelli, M.; Russo-Volpe, S.; Ferrari, E.; Cestaro, B. Decreased membrane fluidity and altered susceptibility to peroxidation and lipid composition in overweight and obese female erythrocytes. J. Lipid Res. 2004, 45, 1846–1851. [Google Scholar] [CrossRef] [Green Version]
- Pietiläinen, K.; Róg, T.; Seppänen-Laakso, T.; Virtue, S.; Tang, J.; Vattulainen, I.; Vidal-Puig, A.; Oresic, M. Remodeling of Adipose Tissue Lipidome as Adaptation to Acquired Obesity: Benefits and Costs. In Proceedings of the Metabolomics 2010: Breakthroughs in Plant, Microbial and Human Biology, Clinical and Nutritional Research, and Biomarker Discovery, Amsterdam, The Netherlands, 27 June–1 July 2010; p. 164. Available online: http://fiehnlab.ucdavis.edu/downloads/staff/kind/metabolomics_2010_abstract_book (accessed on 1 July 2010).
- Arranz, S.; Jauregibeitia, I.; Tueros, I.; Amezaga, J.; Uriarte, M. Lipidomic membrane as a molecular basis for precision nutrition in childhood obesity. Proc. Nutr. Soc. 2019, 78. [Google Scholar] [CrossRef] [Green Version]
- Jauregibeitia, I.; Portune, K.; Rica, I.; Tueros, I.; Velasco, O.; Grau, G.; Trebolazabala, N.; Castaño, L.; Larocca, A.V.; Ferreri, C. Fatty Acid Profile of Mature Red Blood Cell Membranes and Dietary Intake as a New Approach to Characterize Children with Overweight and Obesity. Nutrients 2020, 12, 3446. [Google Scholar] [CrossRef]
- Maulucci, G.; Cohen, O.; Daniel, B.; Sansone, A.; Petropoulou, P.; Filou, S.; Spyridonidis, A.; Pani, G.; De Spirito, M.; Chatgilialoglu, C. Fatty acid-related modulations of membrane fluidity in cells: Detection and implications. Free. Radic. Res. 2016, 50 (Suppl. 1), S40–S50. [Google Scholar] [CrossRef] [Green Version]
- Romero, L.O.; Massey, A.E.; Mata-Daboin, A.D.; Sierra-Valdez, F.J.; Chauhan, S.C.; Cordero-Morales, J.F.; Vásquez, V. Dietary fatty acids fine-tune Piezo1 mechanical response. Nat. Commun. 2019, 10, 1200. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Sheng, W.; Sun, G.Y.; Lee, J.C.-M. Effects of fatty acid unsaturation numbers on membrane fluidity and α-secretase-dependent amyloid precursor protein processing. Neurochem. Int. 2011, 58, 321–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Arribas, A.B.; Goni, F.M.; Alonso, A. Lipid Self-Assemblies under the Atomic Force Microscope. Int. J. Mol. Sci. 2021, 22, 85. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Manyes, S.; Redondo-Morata, L.; Oncins, G.; Sanz, F. Nanomechanics of lipid bilayers: Heads or tails? J. Am Chem. Soc. 2010, 132, 12874–12886. [Google Scholar] [CrossRef] [PubMed]
- Lakowicz, J.; Prendergast, F. Detection of hindered rotations of 1, 6-diphenyl-1, 3, 5-hexatriene in lipid bilayers by differential polarized phase fluorometry. Biophys. J. 1978, 24, 213–231. [Google Scholar] [CrossRef] [Green Version]
- Kuhry, J.-G.; Fonteneau, P.; Duportail, G.; Maechling, C.; Laustriat, G. TMA-DPH: A suitable fluorescence polarization probe for specific plasma membrane fluidity studies in intact living cells. Cell Biophys. 1983, 5, 129–140. [Google Scholar] [CrossRef]
- Poojari, C.; Wilkosz, N.; Lira, R.B.; Dimova, R.; Jurkiewicz, P.; Petka, R.; Kepczynski, M.; Rog, T. Behavior of the DPH fluorescence probe in membranes perturbed by drugs. Chem. Phys. Lipids 2019, 223, 104784. [Google Scholar] [CrossRef]
- Niko, Y.; Didier, P.; Mely, Y.; Konishi, G.; Klymchenko, A.S. Bright and photostable push-pull pyrene dye visualizes lipid order variation between plasma and intracellular membranes. Sci. Rep. 2016, 6, 18870. [Google Scholar] [CrossRef] [Green Version]
- Sot, J.; Esnal, I.; Monasterio, B.G.; Leon-Irra, R.; Niko, Y.; Goni, F.M.; Klymchenko, A.; Alonso, A. Phase-selective staining of model and cell membranes, lipid droplets and lipoproteins with fluorescent solvatochromic pyrene probes. Biochim. Et Biophys. Acta. Biomembr. 2021, 1863, 183470. [Google Scholar] [CrossRef]
- Smith, W.L.; Song, I. The enzymology of prostaglandin endoperoxide H synthases-1 and -2. Prostaglandins Other Lipid Mediat. 2002, 68-69, 115–128. [Google Scholar] [CrossRef]
- Belch, J.J.; Hill, A. Evening primrose oil and borage oil in rheumatologic conditions. Am. J. Clin. Nutr. 2000, 71 (Suppl. S1), 352S–356S. [Google Scholar] [CrossRef] [Green Version]
- Ferreri, C.; Chatgilialoglu, C. Role of fatty acid-based functional lipidomics in the development of molecular diagnostic tools. Expert Rev. Mol. Diagn. 2012, 12, 767–780. [Google Scholar] [CrossRef]
- Innes, J.K.; Calder, P.C. Marine Omega-3 (N-3) Fatty Acids for Cardiovascular Health: An Update for 2020. Int. J. Mol. Sci. 2020, 21, 1362. [Google Scholar] [CrossRef] [Green Version]
- Mandviwala, T.; Khalid, U.; Deswal, A. Obesity and cardiovascular disease: A risk factor or a risk marker? Curr. Atheroscler. Rep. 2016, 18, 21. [Google Scholar] [CrossRef]
- Simons, K.; Sampaio, J.L. Membrane organization and lipid rafts. Cold Spring Harb. Perspect. Biol. 2011, 3, a004697. [Google Scholar] [CrossRef]
- Ahyayauch, H.; García-Arribas, A.B.; Sot, J.; González-Ramírez, E.J.; Busto, J.V.; Monasterio, B.G.; Jiménez-Rojo, N.; Contreras, F.X.; Rendón-Ramírez, A.; Martin, C.; et al. Pb(II) Induces Scramblase Activation and Ceramide-Domain Generation in Red Blood Cells. Sci. Rep. 2018, 8, 7456. [Google Scholar] [CrossRef] [Green Version]
- Carquin, M.; Pollet, H.; Veiga-da-Cunha, M.; Cominelli, A.; Van Der Smissen, P.; N’Kuli, F.; Emonard, H.; Henriet, P.; Mizuno, H.; Courtoy, P.J.; et al. Endogenous sphingomyelin segregates into submicrometric domains in the living erythrocyte membrane. J. Lipid Res. 2014, 55, 1331–1342. [Google Scholar] [CrossRef] [Green Version]
- Montes, L.R.; Lopez, D.J.; Sot, J.; Bagatolli, L.A.; Stonehouse, M.J.; Vasil, M.L.; Wu, B.X.; Hannun, Y.A.; Goñi, F.M.; Alonso, A. Ceramide-enriched membrane domains in red blood cells and the mechanism of sphingomyelinase-induced hot-cold hemolysis. Biochemistry 2008, 47, 11222–11230. [Google Scholar] [CrossRef] [Green Version]
- D’Auria, L.; Fenaux, M.; Aleksandrowicz, P.; Van Der Smissen, P.; Chantrain, C.; Vermylen, C.; Vikkula, M.; Courtoy, P.J.; Tyteca, D. Micrometric segregation of fluorescent membrane lipids: Relevance for endogenous lipids and biogenesis in erythrocytes. J. Lipid Res. 2013, 54, 1066–1076. [Google Scholar] [CrossRef] [Green Version]
- García-Arribas, A.B.; Ahyayauch, H.; Sot, J.; López-González, P.L.; Alonso, A.; Goñi, F.M. Ceramide-induced lamellar gel phases in fluid cell lipid extracts. Langmuir 2016, 32, 9053–9063. [Google Scholar] [CrossRef]
- Redondo-Morata, L.; Giannotti, M.I.; Sanz, F. Influence of cholesterol on the phase transition of lipid bilayers: A temperature-controlled force spectroscopy study. Langmuir 2012, 28, 12851–12860. [Google Scholar] [CrossRef]
- Sullan, R.M.A.; Li, J.K.; Hao, C.; Walker, G.C.; Zou, S. Cholesterol-dependent nanomechanical stability of phase-segregated multicomponent lipid bilayers. Biophys. J. 2010, 99, 507–516. [Google Scholar] [CrossRef] [Green Version]
- Rajamoorthi, K.; Petrache, H.I.; McIntosh, T.J.; Brown, M.F. Packing and viscoelasticity of polyunsaturated omega-3 and omega-6 lipid bilayers as seen by (2)H NMR and X-ray diffraction. J. Am Chem. Soc. 2005, 127, 1576–1588. [Google Scholar] [CrossRef]
- Murthy, A.V.R.; Guyomarc’h, F.; Lopez, C. Palmitoyl ceramide promotes milk sphingomyelin gel phase domains formation and affects the mechanical properties of the fluid phase in milk-SM/DOPC supported membranes. Biochim. Biophys. Acta Biomembr. 2018, 1860, 635–644. [Google Scholar] [CrossRef]
- García-Arribas, A.B.; Busto, J.V.; Alonso, A.; Goñi, F.M. Atomic force microscopy characterization of palmitoylceramide and cholesterol effects on phospholipid bilayers: A topographic and nanomechanical study. Langmuir 2015, 31, 3135–3145. [Google Scholar] [CrossRef]
- Chiantia, S.; Kahya, N.; Ries, J.; Schwille, P. Effects of ceramide on liquid-ordered domains investigated by simultaneous AFM and FCS. Biophys. J. 2006, 90, 4500–4508. [Google Scholar] [CrossRef] [Green Version]
- Alessandrini, A.; Facci, P. Nanoscale mechanical properties of lipid bilayers and their relevance in biomembrane organization and function. Micron 2012, 43, 1212–1223. [Google Scholar] [CrossRef] [Green Version]
- Alessandrini, A.; Seeger, H.M.; Caramaschi, T.; Facci, P. Dynamic force spectroscopy on supported lipid bilayers: Effect of temperature and sample preparation. Biophys. J. 2012, 103, 38–47. [Google Scholar] [CrossRef] [Green Version]
- Seeger, H.M.; Di Cerbo, A.; Alessandrini, A.; Facci, P. Supported lipid bilayers on mica and silicon oxide: Comparison of the main phase transition behavior. J. Phys. Chem. B 2010, 114, 8926–8933. [Google Scholar] [CrossRef]
- Relat-Goberna, J.; Beedle, A.E.M.; Garcia-Manyes, S. The Nanomechanics of Lipid Multibilayer Stacks Exhibits Complex Dynamics. Small 2017, 13, 24. [Google Scholar] [CrossRef] [Green Version]
- Behbodikhah, J.; Ahmed, S.; Elyasi, A.; Kasselman, L.J.; De Leon, J.; Glass, A.D.; Reiss, A.B. Apolipoprotein B and Cardiovascular Disease: Biomarker and Potential Therapeutic Target. Metabolites 2021, 11, 690. [Google Scholar] [CrossRef]
- Klein-Szanto, A.J.; Bassi, D.E. Keep recycling going: New approaches to reduce LDL-C. Biochem. Pharmacol. 2019, 164, 336–341. [Google Scholar] [CrossRef]
- Julve, J.; Martin-Campos, J.M.; Escola-Gil, J.C.; Blanco-Vaca, F. Chylomicrons: Advances in biology, pathology, laboratory testing, and therapeutics. Clin. Chim. Acta Int. J. Clin. Chem. 2016, 455, 134–148. [Google Scholar] [CrossRef] [PubMed]
- Jauregibeitia, I.; Portune, K.; Gaztambide, S.; Rica, I.; Tueros, I.; Velasco, O.; Grau, G.; Martín, A.; Castaño, L.; Larocca, A.V. Molecular Differences Based on Erythrocyte Fatty Acid Profile to Personalize Dietary Strategies between Adults and Children with Obesity. Metabolites 2021, 11, 43. [Google Scholar] [CrossRef] [PubMed]
- Monasterio, B.G.; Alonso, B.; Sot, J.; Garcia-Arribas, A.B.; Gil-Carton, D.; Valle, M.; Zurutuza, A.; Goni, F.M. Coating Graphene Oxide with Lipid Bilayers Greatly Decreases Its Hemolytic Properties. Langmuir 2017, 33, 8181–8191. [Google Scholar] [CrossRef] [PubMed]
- Ghezzo, A.; Visconti, P.; Abruzzo, P.M.; Bolotta, A.; Ferreri, C.; Gobbi, G.; Malisardi, G.; Manfredini, S.; Marini, M.; Nanetti, L. Oxidative stress and erythrocyte membrane alterations in children with autism: Correlation with clinical features. PLoS ONE 2013, 8, e66418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sansone, A.; Tolika, E.; Louka, M.; Sunda, V.; Deplano, S.; Melchiorre, M.; Anagnostopoulos, D.; Chatgilialoglu, C.; Formisano, C.; Di Micco, R. Hexadecenoic fatty acid isomers in human blood lipids and their relevance for the interpretation of lipidomic profiles. PLoS ONE 2016, 11, e0152378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Vegt, S.G.L.; Ruben, A.T.; Werre, J.; Palsma, D.; Verhoef, C.; De Gier, J.; Staal, G. Counterflow centrifugation of red cell populations: A cell age related separation technique. Br. J. Haematol. 1985, 61, 393–403. [Google Scholar] [CrossRef]
- Breil, C.; Abert Vian, M.; Zemb, T.; Kunz, W.; Chemat, F. “Bligh and Dyer” and Folch methods for solid–liquid–liquid extraction of lipids from microorganisms. Comprehension of solvatation mechanisms and towards substitution with alternative solvents. Int. J. Mol. Sci. 2017, 18, 708. [Google Scholar] [CrossRef] [Green Version]
- Ferreri, C.; Mennella, M.R.F.; Formisano, C.; Landi, L.; Chatgilialoglu, C. Arachidonate geometrical isomers generated by thiyl Radicals: The relationship with trans lipids detected in biological samples. Free. Radic. Biol. Med. 2002, 33, 1516–1526. [Google Scholar] [CrossRef]
- Bottcher, C. A rapid and sensitive sub-micro phosphorus determination. Anal. Chim. Acta 1961, 24, 203. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex | Age (yr) | BMI (kg × m−2) | |
|---|---|---|---|
| Control (49) | 16 M | 30.2 ± 13.2 | 21.8 ± 5.6 |
| 33 F | 34.5 ± 14.5 | 21.5 ± 4.2 | |
| Obese (52) | 16 M | 48.0 ± 13.0 | 38.2 ± 11.0 |
| 36 F | 45.8 ± 12.6 | 40.7 ± 8.7 |
| Internal Standard Mix | Compound Name | Exact Mass | Chemical Formule | Conc. (µg/mL) |
|---|---|---|---|---|
| Splash LipidoMix | 15:0–18:1(d7) PC | 752.6061 | C41H73D7NO8P | 150.6 |
| 15:0–18:1(d7) PE | 710.5591 | C38H67D7NO8P | 5.3 | |
| 15:0–18:1(d7) PS (Na Salt) | 776.5309 | C39H66D7NNaO10P | 3.9 | |
| 15:0–18:1(d7) PG (Na Salt) | 763.5357 | C39H67D7NaO10P | 26.7 | |
| 15:0–18:1(d7) PI (NH4 Salt) | 846.5963 | C42H75D7NO13P | 8.5 | |
| 15:0–18:1(d7) PA (Na Salt) | 689.4994 | C36H61D7NaO8P | 6.9 | |
| 18:1(d7) Lyso PC | 528.3921 | C26H45D7NO7P | 23.8 | |
| 18:1(d7) Lyso PE | 486.3451 | C23H39D7NO7P | 4.9 | |
| 18:1(d7) Chol Ester | 657.6441 | C45H71D7O2 | 329.1 | |
| 18:1(d7) MG | 363.3366 | C21H33D7O4 | 1.8 | |
| 15:0–18:1(d7) DG | 587.5506 | C36H61D7O5 | 8.8 | |
| 15:0–18:1(d7)-15:0 TG | 811.7646 | C51H89D7O6 | 52.8 | |
| D18:1-18:1(d9) SM | 737.6397 | C41H72D9N2O6P | 29.6 | |
| Cholesterol (d7) | 393.3988 | C27H39D7O | 98.4 | |
| Ceramide/Sphingoid Internal Standard Mixture I | Sphingosine (C17) | 285.2668 | C17H35NO2 | 6.9 |
| Sphinganine (C17) | 287.2824 | C17H37NO2 | 7.1 | |
| Sphingosine-1-P (C17) | 365.2331 | C17H36NO5P | 9.3 | |
| Sphinganine-1-P (C17) | 367.2488 | C17H38NO5P | 9.2 | |
| Ceramide (C12) | 481.4495 | C30H59NO3 | 11.6 | |
| Ceramide (C25) | 663.6529 | C43H85NO3 | 16.2 | |
| Ceramide-1-P (C12) | 561.4158 | C30H60NO6P | 13.4 | |
| Sphingomyelin (C12) | 646.5050 | C35H71N2O6P | 15.7 | |
| Glucosyl(β) C12 Ceramide | 643.5023 | C36H69NO8 | 15.6 | |
| Lactosyl(β) C12 Ceramide | 805.5550 | C42H79NO13 | 19.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sot, J.; García-Arribas, A.B.; Abad, B.; Arranz, S.; Portune, K.; Andrade, F.; Martín-Nieto, A.; Velasco, O.; Arana, E.; Tueros, I.; et al. Erythrocyte Membrane Nanomechanical Rigidity Is Decreased in Obese Patients. Int. J. Mol. Sci. 2022, 23, 1920. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031920
Sot J, García-Arribas AB, Abad B, Arranz S, Portune K, Andrade F, Martín-Nieto A, Velasco O, Arana E, Tueros I, et al. Erythrocyte Membrane Nanomechanical Rigidity Is Decreased in Obese Patients. International Journal of Molecular Sciences. 2022; 23(3):1920. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031920
Chicago/Turabian StyleSot, Jesús, Aritz B. García-Arribas, Beatriz Abad, Sara Arranz, Kevin Portune, Fernando Andrade, Alicia Martín-Nieto, Olaia Velasco, Eunate Arana, Itziar Tueros, and et al. 2022. "Erythrocyte Membrane Nanomechanical Rigidity Is Decreased in Obese Patients" International Journal of Molecular Sciences 23, no. 3: 1920. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031920