
Nitroxyl Delivered by Angeli’s Salt Causes Short-Lasting Activation Followed by Long-Lasting Deactivation of Meningeal Afferents in Models of Headache Generation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
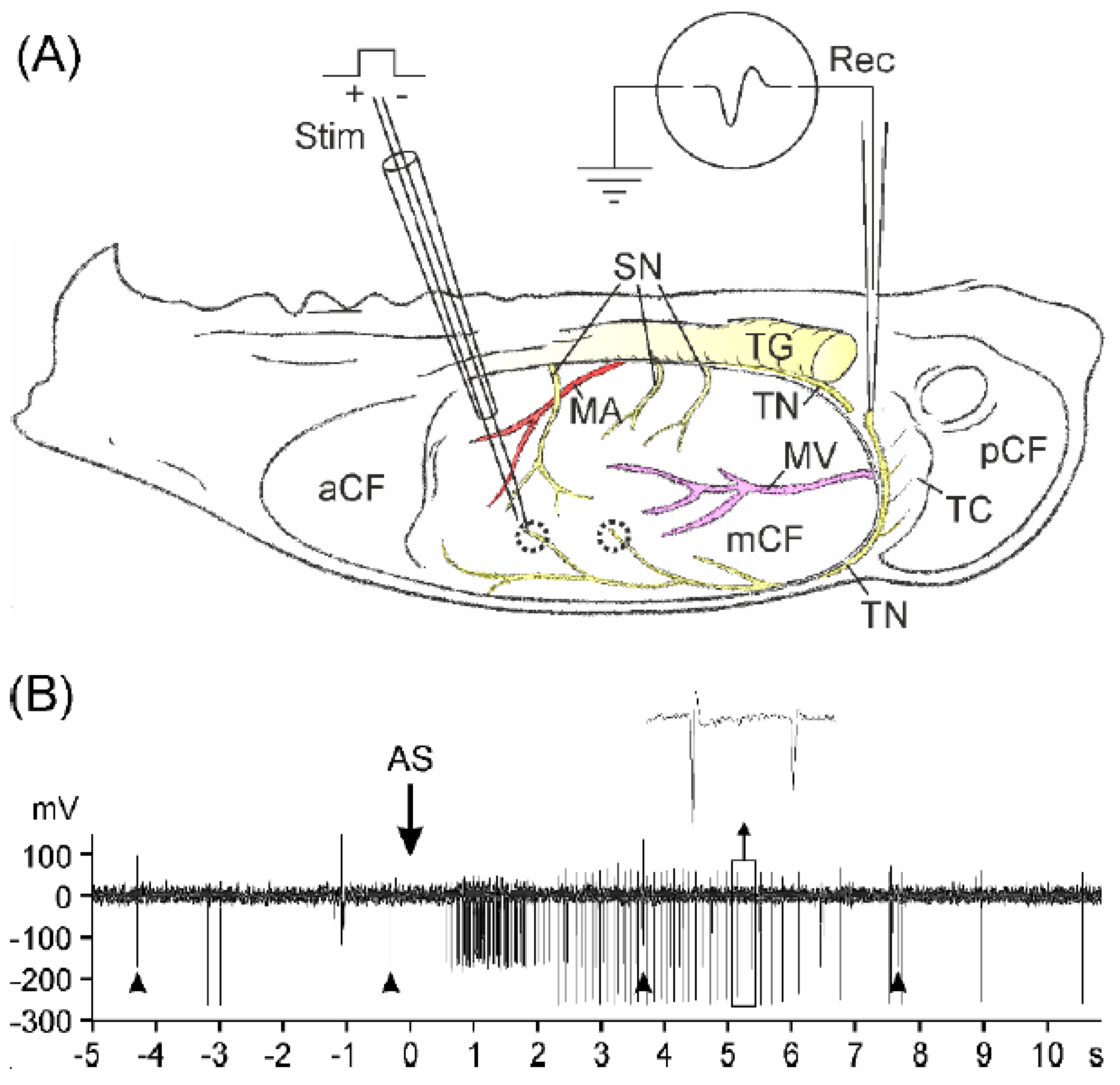
2.1. Primary Afferent Recordings In Vitro
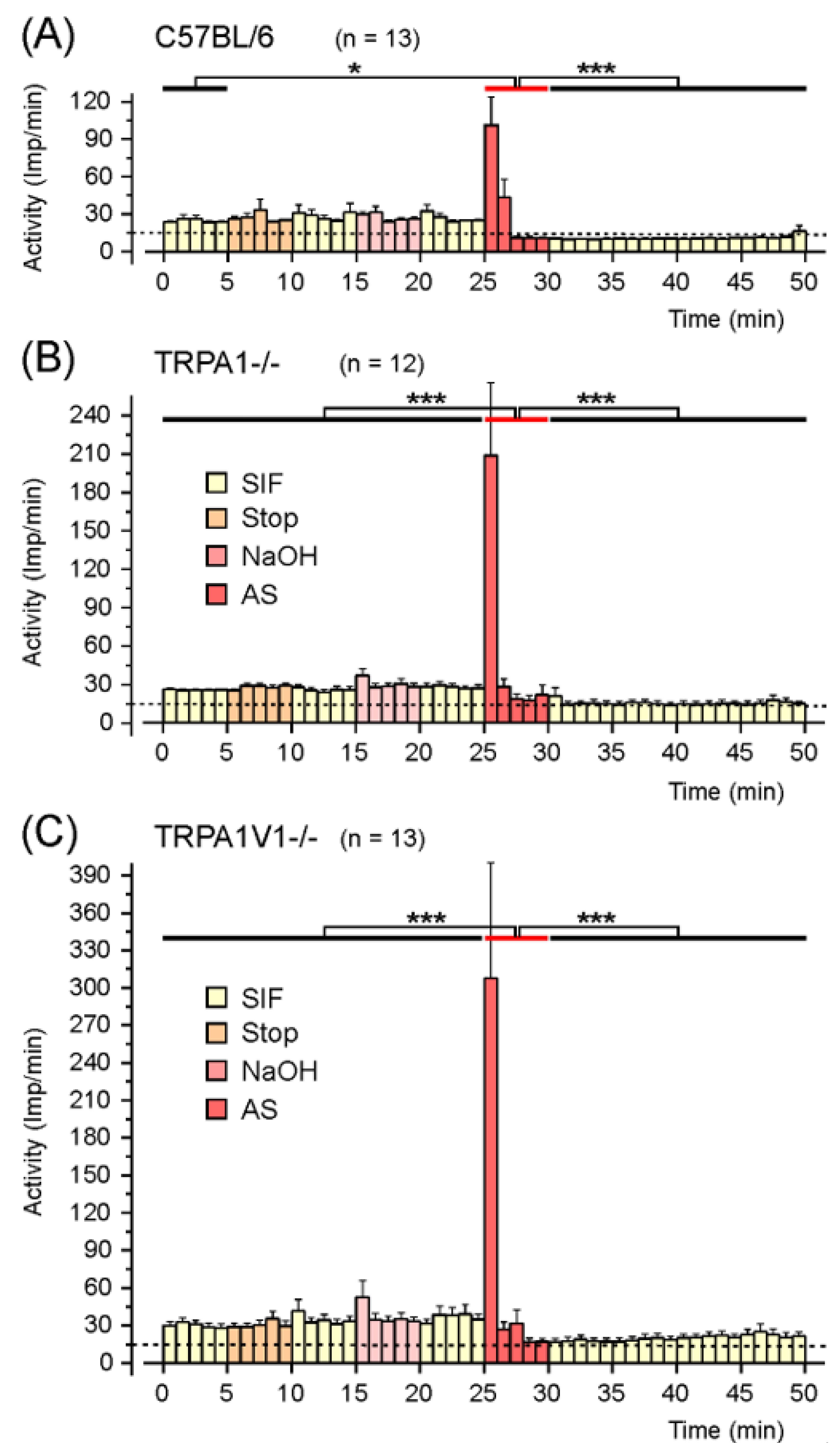
2.1.1. Responses to AS in C57BL/6 Wild-Type Mice
2.1.2. Responses to AS in TRPA1−/− Mice
2.1.3. Responses to AS in TRPA1/V1−/− Mice
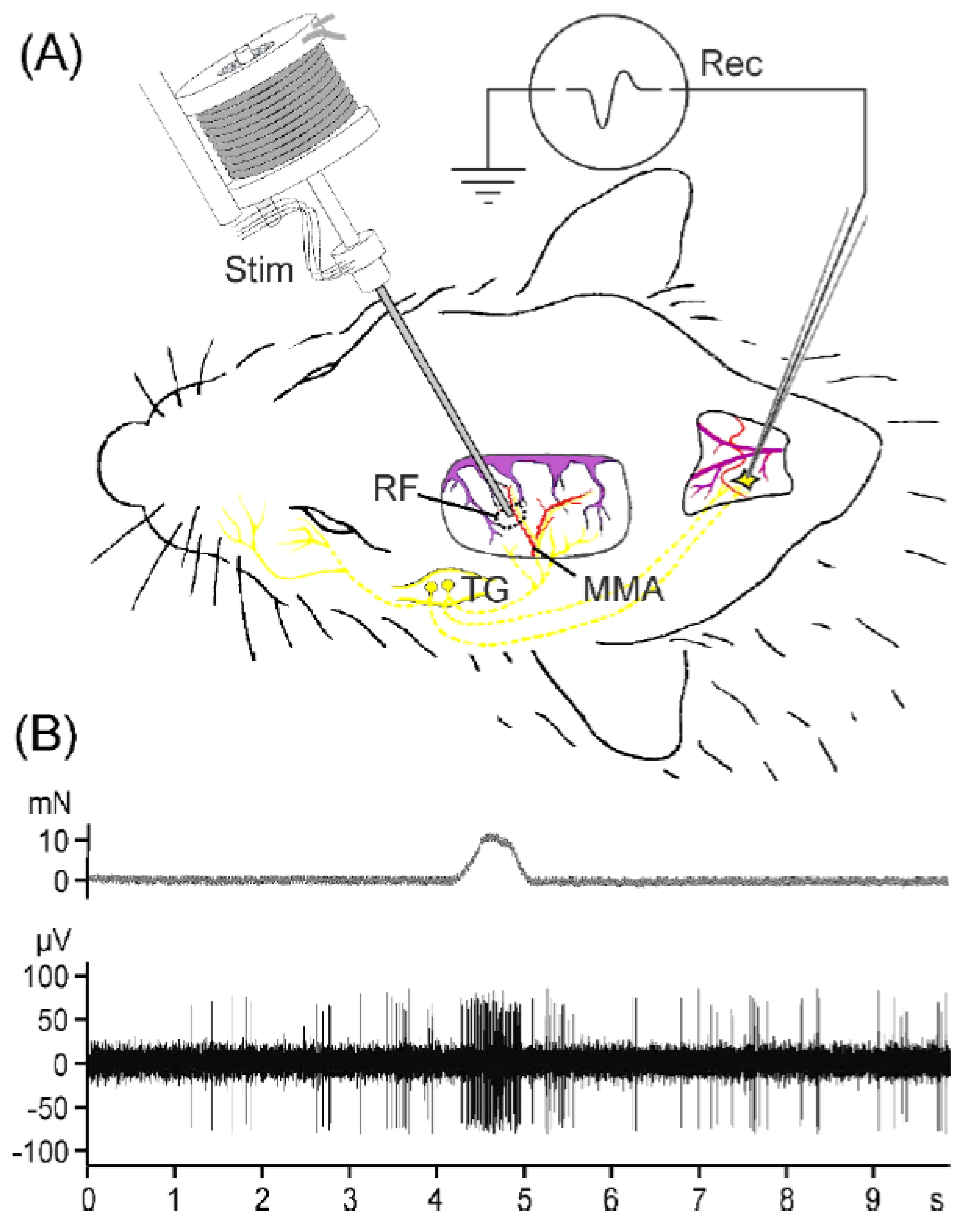
2.2. Second-Order Neuron Recordings In Vivo
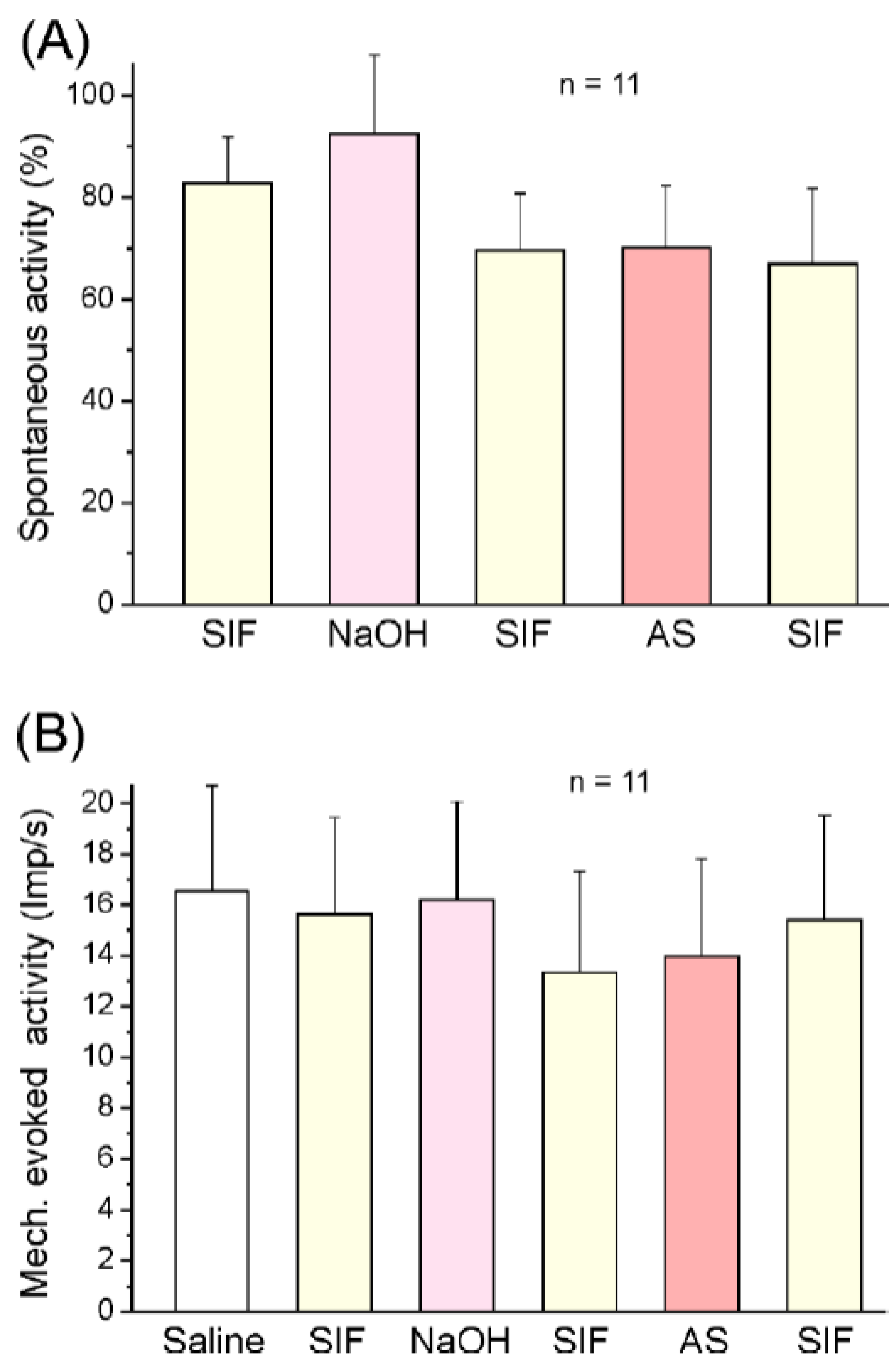
2.2.1. Responses of Spinal Trigeminal Neurons to Angeli’s Salt
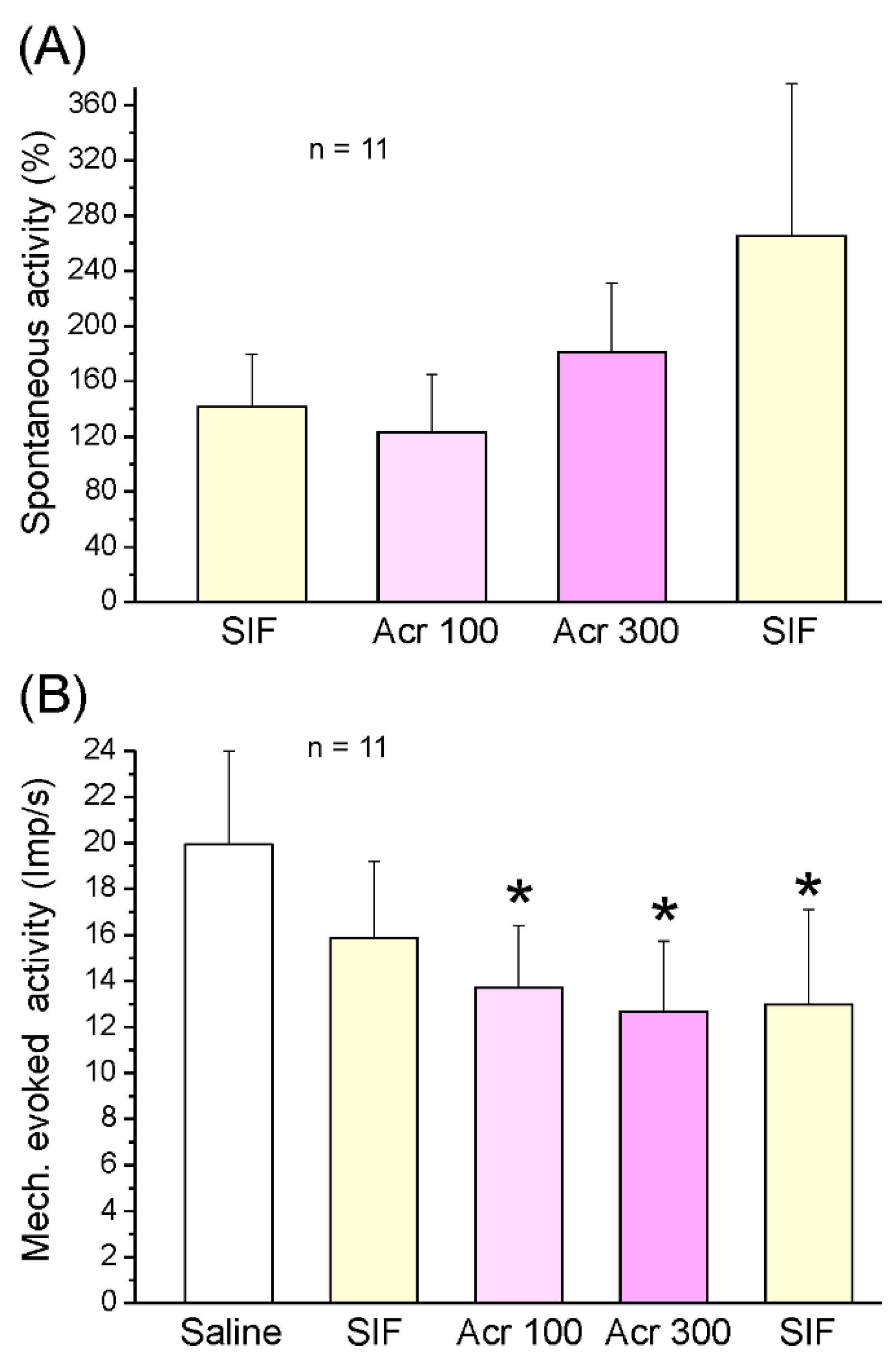
2.2.2. Responses of Spinal Trigeminal Neurons to Acrolein
3. Discussion
4. Materials and Methods
4.1. Hemisected Head Preparation and Experimental Setup
4.2. Recordings of Meningeal Afferent Activity and Chemical Stimulation
4.3. Preparation for Recordings In Vivo and Experimental Setup
4.4. Recordings of Spinal Trigeminal Activity and Chemical Stimulation
4.5. Data Processing and Statistics
4.5.1. Primary Afferent Activity
4.5.2. Second Order Neuron Activity
4.5.3. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ramsey, I.S.; Delling, M.; Clapham, D.E. An Introduction to TRP Channels. Annu. Rev. Physiol. 2006, 68, 619–647. [Google Scholar] [CrossRef] [Green Version]
- Julius, D. TRP Channels and Pain. Annu. Rev. Cell Dev. Biol. 2013, 29, 355–384. [Google Scholar] [CrossRef] [Green Version]
- Caterina, M.J. How Do You Feel? A Warm and Touching 2021 Nobel Tribute. J. Clin. Investig. 2021, 131, e156587. [Google Scholar] [CrossRef]
- Huang, D.; Li, S.; Dhaka, A.; Story, G.M.; Cao, Y.-Q. Expression of the Transient Receptor Potential Channels TRPV1, TRPA1 and TRPM8 in Mouse Trigeminal Primary Afferent Neurons Innervating the Dura. Mol. Pain 2012, 8, 66. [Google Scholar] [CrossRef] [Green Version]
- Benemei, S.; De Cesaris, F.; Fusi, C.; Rossi, E.; Lupi, C.; Geppetti, P. TRPA1 and Other TRP Channels in Migraine. J. Headache Pain 2013, 14, 71. [Google Scholar] [CrossRef] [Green Version]
- Dux, M.; Rosta, J.; Messlinger, K. TRP Channels in the Focus of Trigeminal Nociceptor Sensitization Contributing to Primary Headaches. Int. J. Mol. Sci. 2020, 21, 342. [Google Scholar] [CrossRef] [Green Version]
- Hinman, A.; Chuang, H.-H.; Bautista, D.M.; Julius, D. TRP Channel Activation by Reversible Covalent Modification. Proc. Natl. Acad. Sci. USA 2006, 103, 19564–19568. [Google Scholar] [CrossRef] [Green Version]
- Macpherson, L.J.; Dubin, A.E.; Evans, M.J.; Marr, F.; Schultz, P.G.; Cravatt, B.F.; Patapoutian, A. Noxious Compounds Activate TRPA1 Ion Channels through Covalent Modification of Cysteines. Nature 2007, 445, 541–545. [Google Scholar] [CrossRef]
- Andersson, D.A.; Gentry, C.; Bevan, S. TRPA1 Has a Key Role in the Somatic Pro-Nociceptive Actions of Hydrogen Sulfide. PLoS ONE 2012, 7, e46917. [Google Scholar] [CrossRef]
- Nassini, R.; Materazzi, S.; Vriens, J.; Prenen, J.; Benemei, S.; De Siena, G.; la Marca, G.; Andrè, E.; Preti, D.; Avonto, C.; et al. The “headache Tree” via Umbellulone and TRPA1 Activates the Trigeminovascular System. Brain 2012, 135, 376–390. [Google Scholar] [CrossRef] [Green Version]
- Eberhardt, M.; Dux, M.; Namer, B.; Miljkovic, J.; Cordasic, N.; Will, C.; Kichko, T.I.; de la Roche, J.; Fischer, M.; Suárez, S.A.; et al. H2S and NO Cooperatively Regulate Vascular Tone by Activating a Neuroendocrine HNO-TRPA1-CGRP Signalling Pathway. Nat. Commun. 2014, 5, 4381. [Google Scholar] [CrossRef] [PubMed]
- Filipovic, M.R. Chapter 6: HNO/Thiol relationship. In The Chemistry and Biology of Nitroxyl (HNO); Doctorovic, F., Farmer, P., Marti, M.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Amatore, C.; Arbault, S.; Ducrocq, C.; Hu, S.; Tapsoba, I. Angeli’s Salt (Na2N2O3) Is a Precursor of HNO and NO: A Voltammetric Study of the Reactive Intermediates Released by Angeli’s Salt Decomposition. ChemMedChem 2007, 2, 898–903. [Google Scholar] [CrossRef] [PubMed]
- Dux, M.; Will, C.; Vogler, B.; Filipovic, M.R.; Messlinger, K. Meningeal Blood Flow Is Controlled by H2S-NO Crosstalk Activating a HNO-TRPA1-CGRP Signalling Pathway. Br. J. Pharmacol. 2016, 173, 431–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teicher, C.; De Col, R.; Messlinger, K. Hydrogen Sulfide Mediating Both Excitatory and Inhibitory Effects in a Rat Model of Meningeal Nociception and Headache Generation. Front. Neurol. 2017, 8, 336. [Google Scholar] [CrossRef] [Green Version]
- Fukuto, J.M.; Switzer, C.H.; Miranda, K.M.; Wink, D.A. Nitroxyl (HNO): Chemistry, Biochemistry, and Pharmacology. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 335–355. [Google Scholar] [CrossRef]
- Denner, A.C.; Vogler, B.; Messlinger, K.; De Col, R. Role of Transient Receptor Potential Ankyrin 1 Receptors in Rodent Models of Meningeal Nociception—Experiments In Vitro. Eur. J. Pain 2016, 21, 843–854. [Google Scholar] [CrossRef]
- Roch, M.; Messlinger, K.; Kulchitsky, V.; Tichonovich, O.; Azev, O.; Koulchitsky, S. Ongoing Activity in Trigeminal Wide-Dynamic Range Neurons Is Driven from the Periphery. Neuroscience 2007, 150, 681–691. [Google Scholar] [CrossRef]
- Pouokam, E.; Bell, A.; Diener, M. Actions of Angeli’s Salt, a Nitroxyl (HNO) Donor, on Ion Transport across Mucosa-Submucosa Preparations from Rat Distal Colon. Eur. J. Pharmacol. 2013, 715, 133–141. [Google Scholar] [CrossRef]
- Gastreich-Seelig, M.; Jimenez, M.; Pouokam, E. Mechanisms Associated to Nitroxyl (HNO)-Induced Relaxation in the Intestinal Smooth Muscle. Front. Physiol. 2020, 11, 438. [Google Scholar] [CrossRef]
- Pouokam, E.; Vallejo, A.; Martínez, E.; Traserra, S.; Jimenez, M. Complementary Mechanisms of Modulation of Spontaneous Phasic Contractions by the Gaseous Signalling Molecules NO, H2S, HNO and the Polysulfide Na2S3 in the Rat Colon. J. Basic Clin. Physiol. Pharmacol. 2021, 11, 181. [Google Scholar] [CrossRef]
- Irvine, J.C.; Favaloro, J.L.; Kemp-Harper, B.K. NO− Activates Soluble Guanylate Cyclase and Kv Channels to Vasodilate Resistance Arteries. Hypertension 2003, 41, 1301–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemp-Harper, B.K.; Velagic, A.; Paolocci, N.; Horowitz, J.D.; Ritchie, R.H. Cardiovascular Therapeutic Potential of the Redox Siblings, Nitric Oxide (NO•) and Nitroxyl (HNO), in the Setting of Reactive Oxygen Species Dysregulation. Handb. Exp. Pharmacol. 2021, 264, 311–337. [Google Scholar] [CrossRef] [PubMed]
- Zarpelon, A.C.; Souza, G.R.; Cunha, T.M.; Schivo, I.R.S.; Marchesi, M.; Casagrande, R.; Pinge-Filho, P.; Cunha, F.Q.; Ferreira, S.H.; Miranda, K.M.; et al. The Nitroxyl Donor, Angeli’s Salt, Inhibits Inflammatory Hyperalgesia in Rats. Neuropharmacology 2013, 71, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Staurengo-Ferrari, L.; Zarpelon, A.C.; Longhi-Balbinot, D.T.; Marchesi, M.; Cunha, T.M.; Alves-Filho, J.C.; Cunha, F.Q.; Ferreira, S.H.; Casagrande, R.; Miranda, K.M.; et al. Nitroxyl Inhibits Overt Pain-like Behavior in Mice: Role of CGMP/PKG/ATP-Sensitive Potassium Channel Signaling Pathway. Pharmacol. Rep. 2014, 66, 691–698. [Google Scholar] [CrossRef] [Green Version]
- Wulf-Johansson, H.; Hay-Schmidt, A.; Poulsen, A.N.; Klaerke, D.A.; Olesen, J.; Jansen-Olesen, I. Expression of BK Ca Channels and the Modulatory Beta-Subunits in the Rat and Porcine Trigeminal Ganglion. Brain Res. 2009, 1292, 1–13. [Google Scholar] [CrossRef]
- Ploug, K.B.; Baun, M.; Hay-Schmidt, A.; Olesen, J.; Jansen-Olesen, I. Presence and Vascular Pharmacology of KATP Channel Subtypes in Rat Central and Peripheral Tissues. Eur. J. Pharmacol. 2010, 637, 109–117. [Google Scholar] [CrossRef]
- Gozalov, A.; Jansen-Olesen, I.; Klaerke, D.; Olesen, J. Role of BK(Ca) Channels in Cephalic Vasodilation Induced by CGRP, NO and Transcranial Electrical Stimulation in the Rat. Cephalalgia 2007, 27, 1120–1127. [Google Scholar] [CrossRef]
- Christensen, S.L.; Munro, G.; Petersen, S.; Shabir, A.; Jansen-Olesen, I.; Kristensen, D.M.; Olesen, J. ATP Sensitive Potassium (KATP) Channel Inhibition: A Promising New Drug Target for Migraine. Cephalalgia 2020, 40, 650–664. [Google Scholar] [CrossRef]
- Al-Karagholi, M.A.-M.; Hansen, J.M.; Guo, S.; Olesen, J.; Ashina, M. Opening of ATP-Sensitive Potassium Channels Causes Migraine Attacks: A New Target for the Treatment of Migraine. Brain 2019, 142, 2644–2654. [Google Scholar] [CrossRef]
- Al-Karagholi, M.A.-M.; Ghanizada, H.; Waldorff Nielsen, C.A.; Skandarioon, C.; Snellman, J.; Lopez-Lopez, C.; Hansen, J.M.; Ashina, M. Opening of BKCa Channels Causes Migraine Attacks: A New Downstream Target for the Treatment of Migraine. Pain 2021, 162, 2512–2520. [Google Scholar] [CrossRef]
- Al-Karagholi, M.A.-M.; Olesen, J.; Ashina, M. Reply: Hyperpolarization through ATP-Sensitive Potassium Channels; Relevance to Migraine Pathology. Brain 2020, 143, e14. [Google Scholar] [CrossRef] [PubMed]
- Haanes, K.A.; Edvinsson, L. Hyperpolarization through ATP-Sensitive Potassium Channels; Relevance to Migraine Pathology. Brain 2020, 143, e13. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.B.; Gray, J.; Gunthorpe, M.J.; Hatcher, J.P.; Davey, P.T.; Overend, P.; Harries, M.H.; Latcham, J.; Clapham, C.; Atkinson, K.; et al. Vanilloid Receptor-1 Is Essential for Inflammatory Thermal Hyperalgesia. Nature 2000, 405, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Kwan, K.Y.; Allchorne, A.J.; Vollrath, M.A.; Christensen, A.P.; Zhang, D.-S.; Woolf, C.J.; Corey, D.P. TRPA1 Contributes to Cold, Mechanical, and Chemical Nociception but Is Not Essential for Hair-Cell Transduction. Neuron 2006, 50, 277–289. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stöckl, S.K.; de Col, R.; Filipovic, M.R.; Messlinger, K. Nitroxyl Delivered by Angeli’s Salt Causes Short-Lasting Activation Followed by Long-Lasting Deactivation of Meningeal Afferents in Models of Headache Generation. Int. J. Mol. Sci. 2022, 23, 2330. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042330
Stöckl SK, de Col R, Filipovic MR, Messlinger K. Nitroxyl Delivered by Angeli’s Salt Causes Short-Lasting Activation Followed by Long-Lasting Deactivation of Meningeal Afferents in Models of Headache Generation. International Journal of Molecular Sciences. 2022; 23(4):2330. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042330
Chicago/Turabian StyleStöckl, Stephanie K., Roberto de Col, Milos R. Filipovic, and Karl Messlinger. 2022. "Nitroxyl Delivered by Angeli’s Salt Causes Short-Lasting Activation Followed by Long-Lasting Deactivation of Meningeal Afferents in Models of Headache Generation" International Journal of Molecular Sciences 23, no. 4: 2330. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23042330