
Effect of Empagliflozin on Sphingolipid Catabolism in Diabetic and Hypertensive Rats

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. General Characteristics of Diabetic Rats
2.2. Characteristics of Angiotensin II-Dependent Hypertension Rats
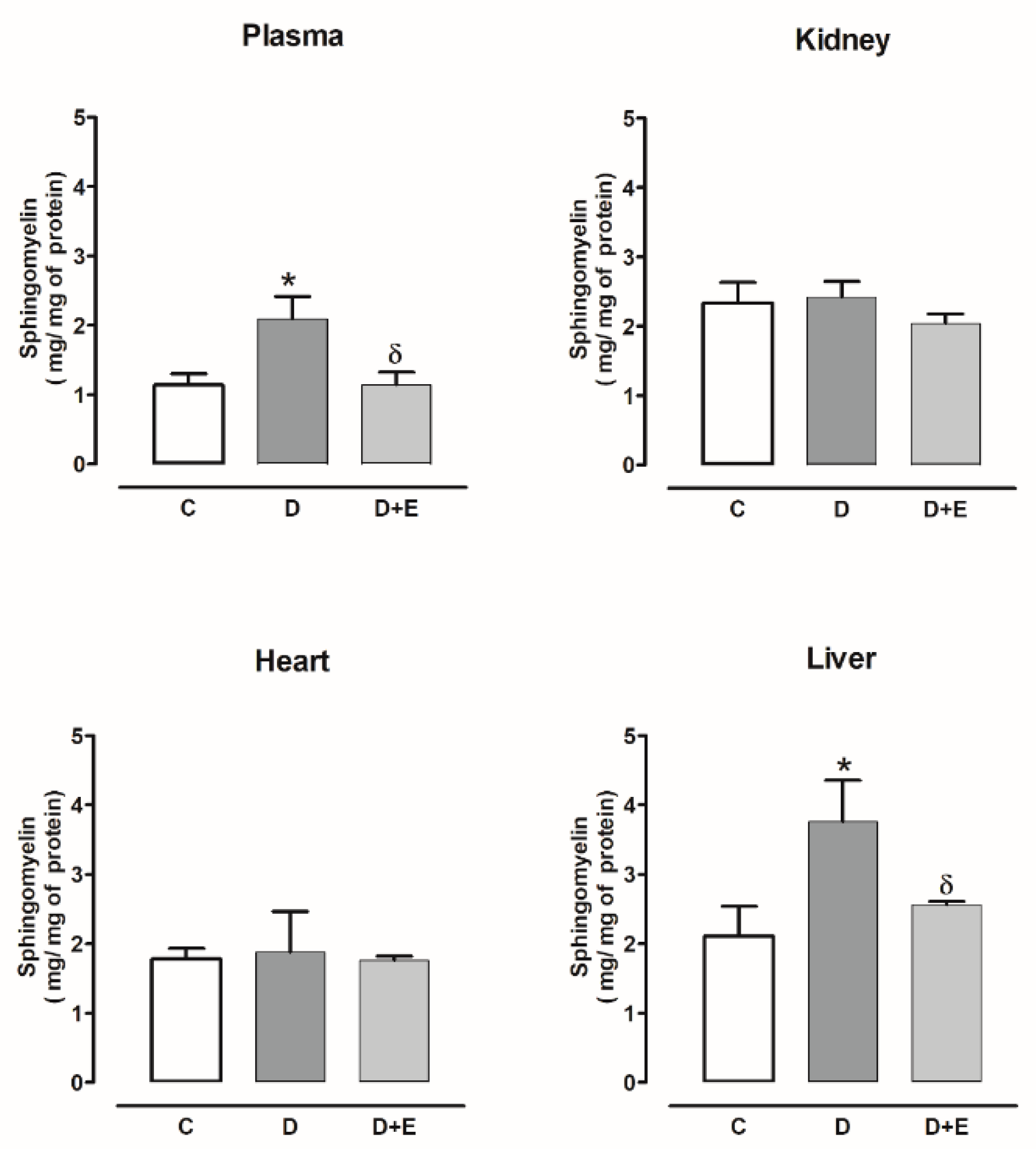
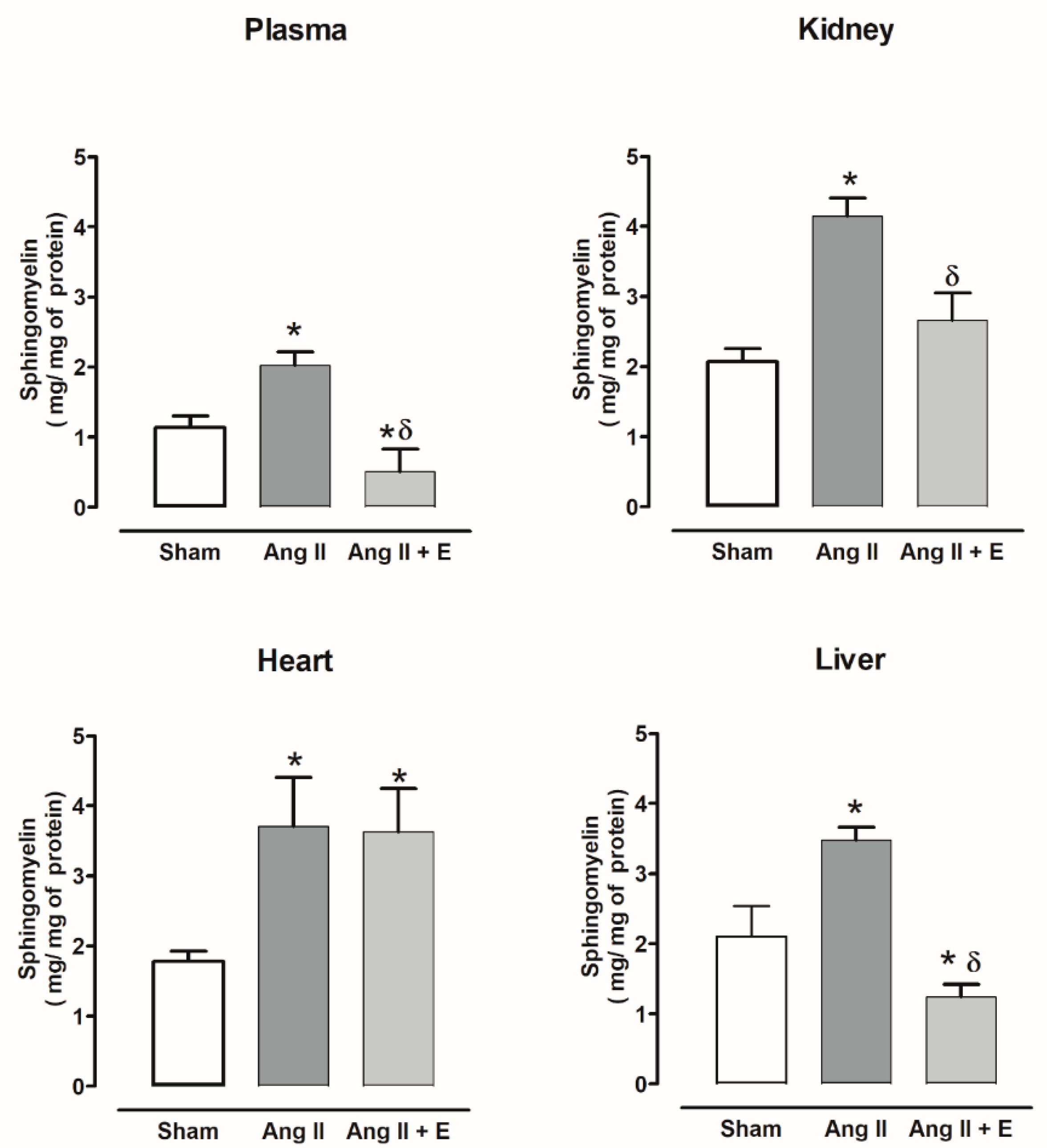
2.3. Sphingomyelin Content in Diabetic and Hypertensive Rats
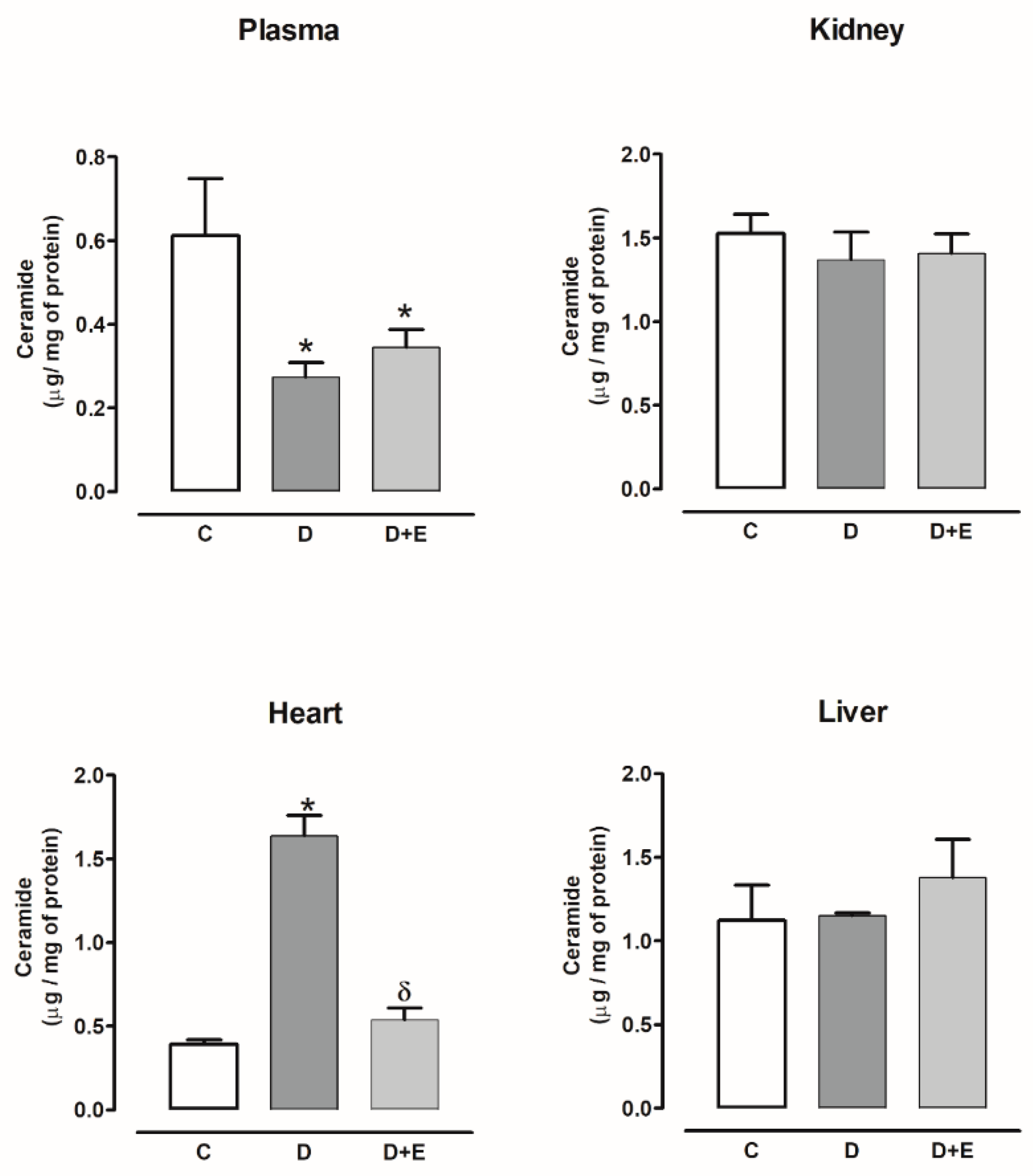
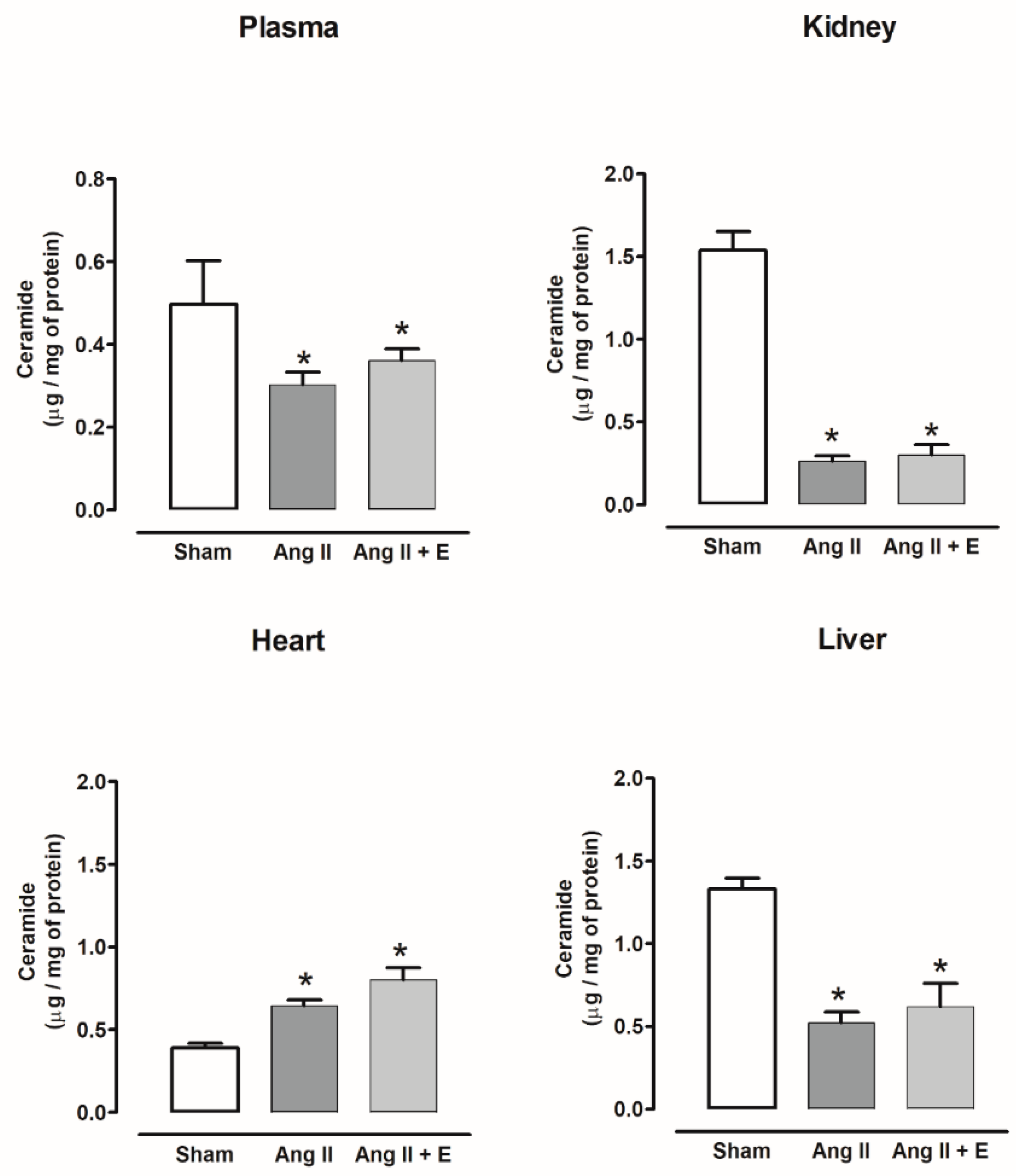
2.4. Ceramide Content in Diabetic and Hypertensive Rats
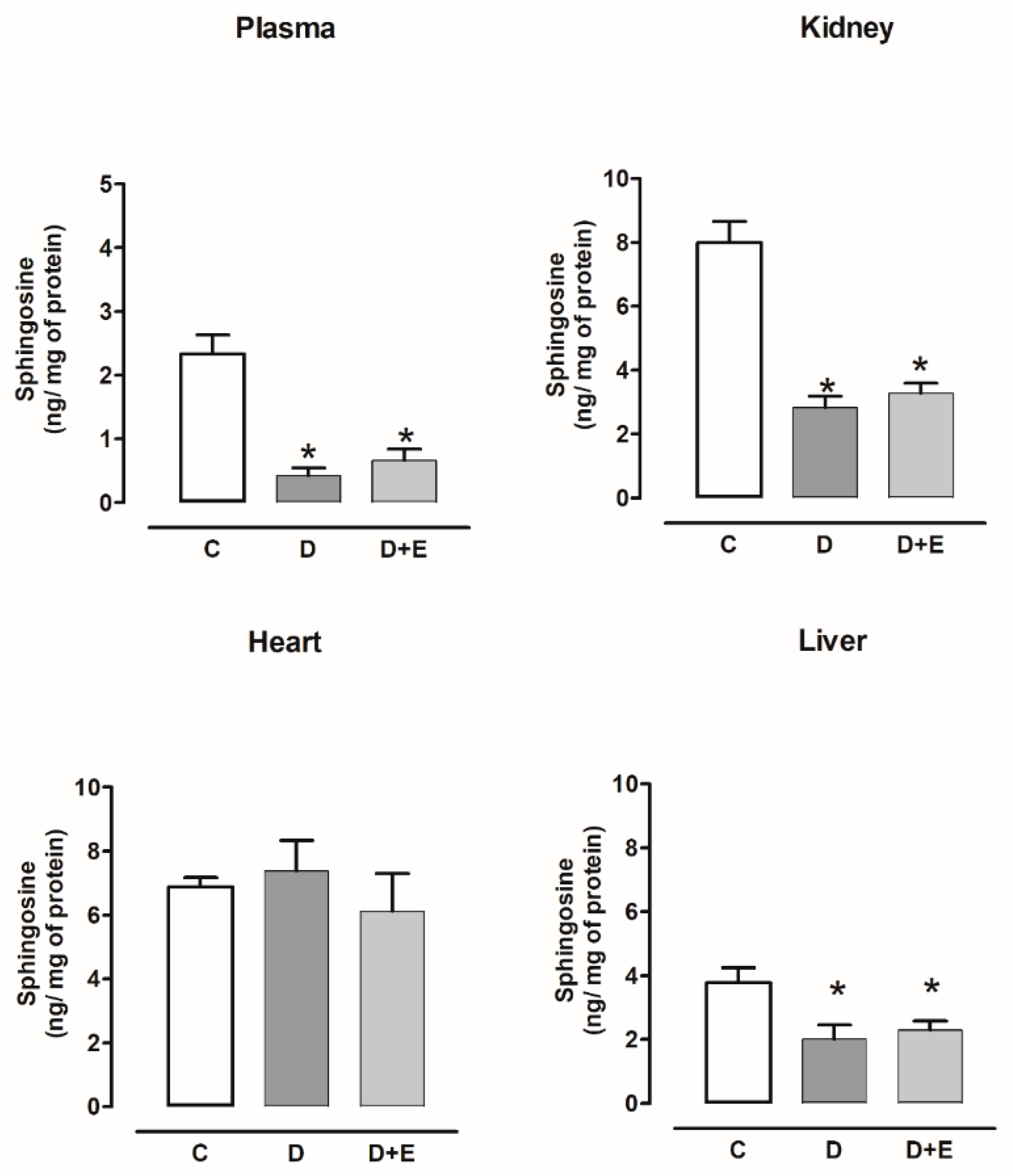
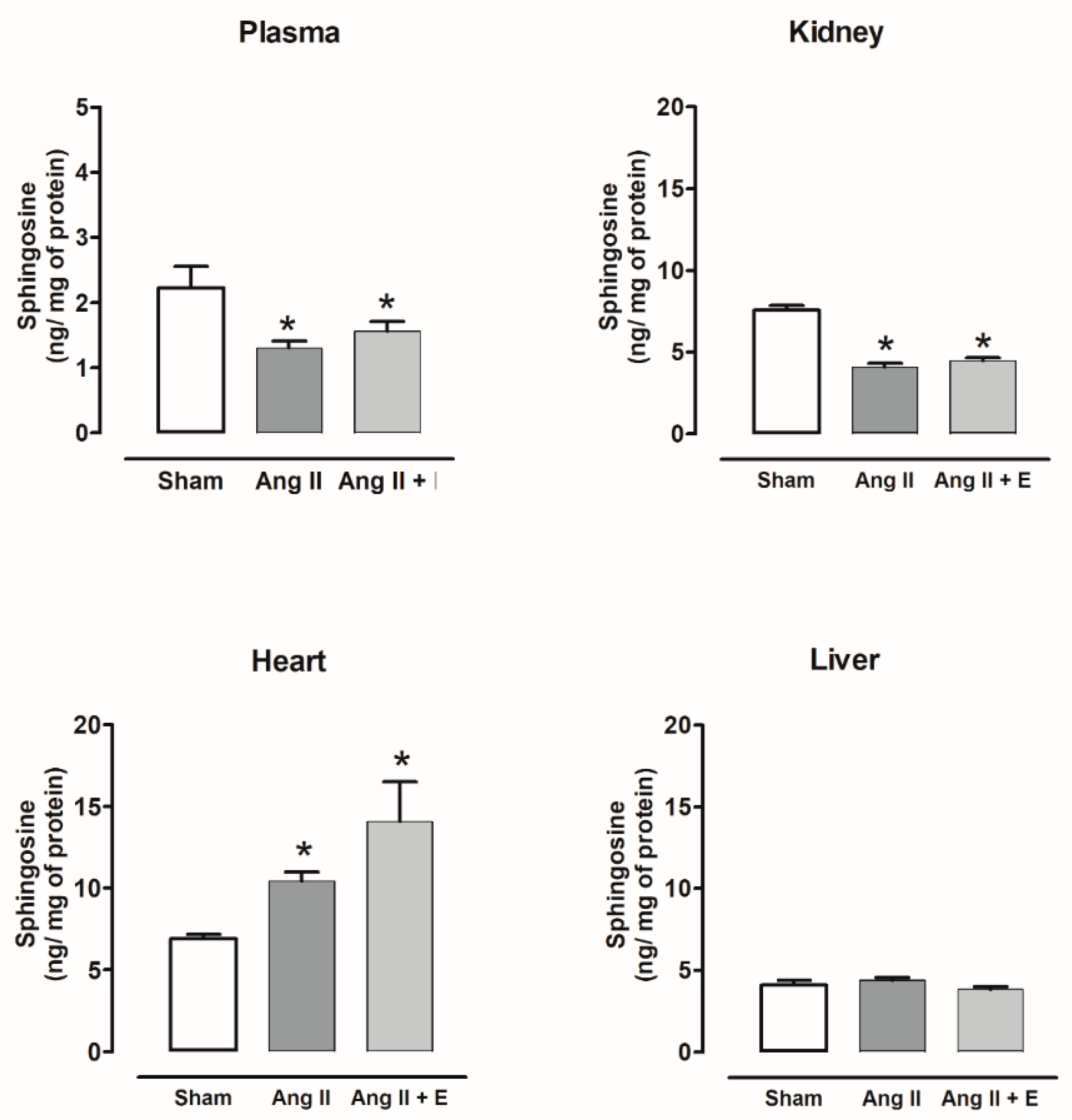
2.5. Sphingosine Content in Diabetic and Hypertensive Rats
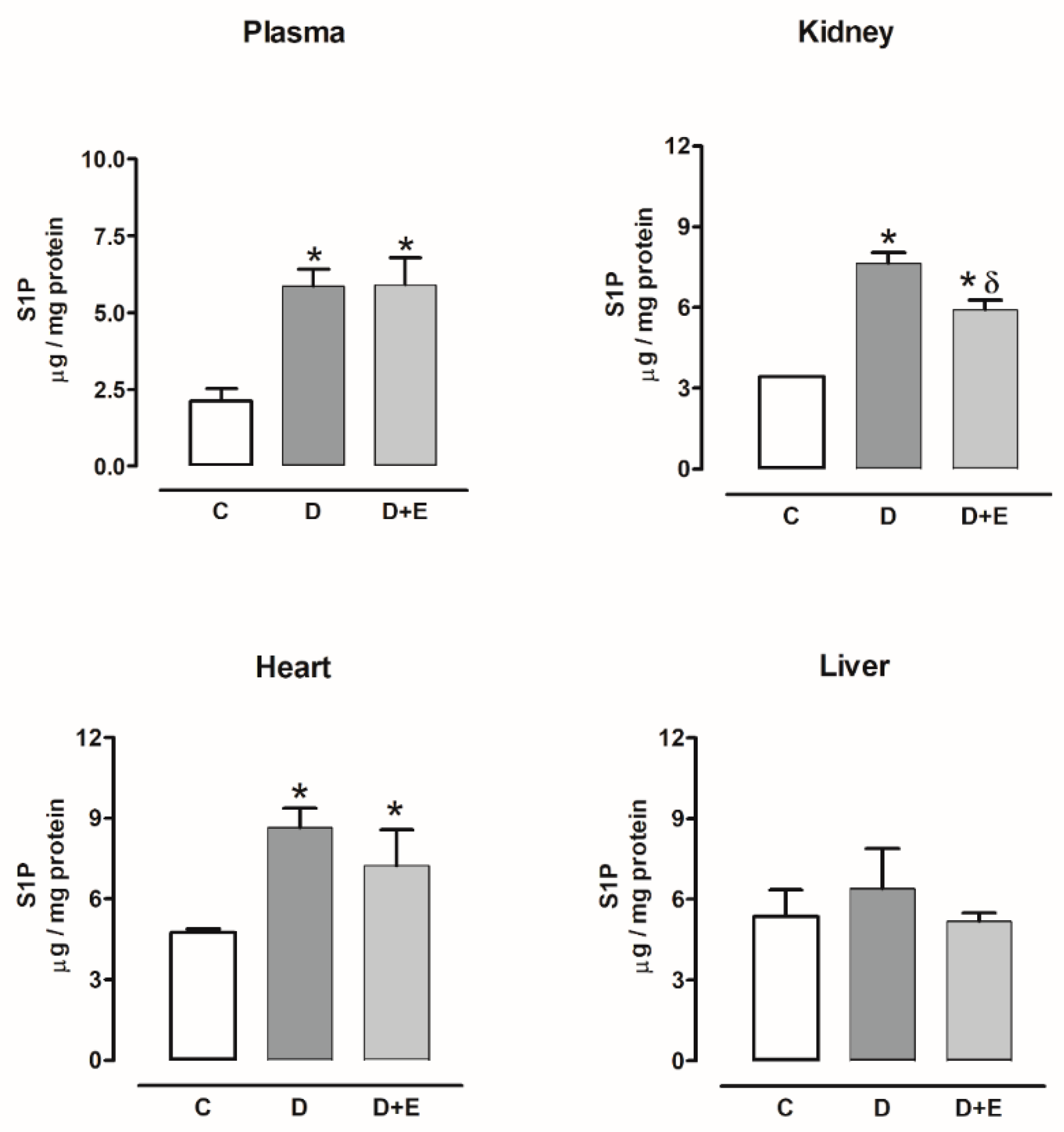
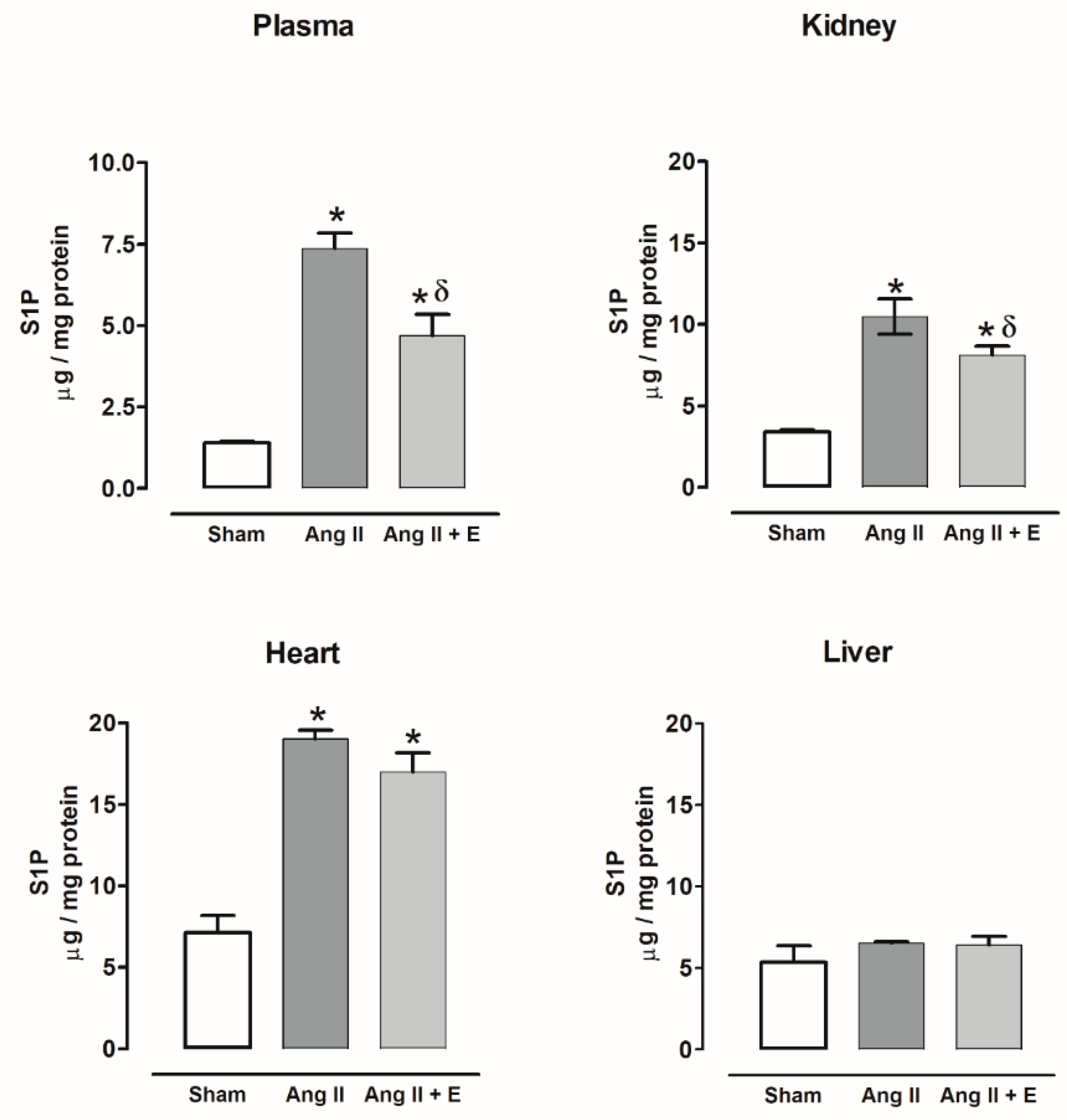
2.6. Sphingosine-1-Phosphate Content in Diabetic and Hypertensive Rats
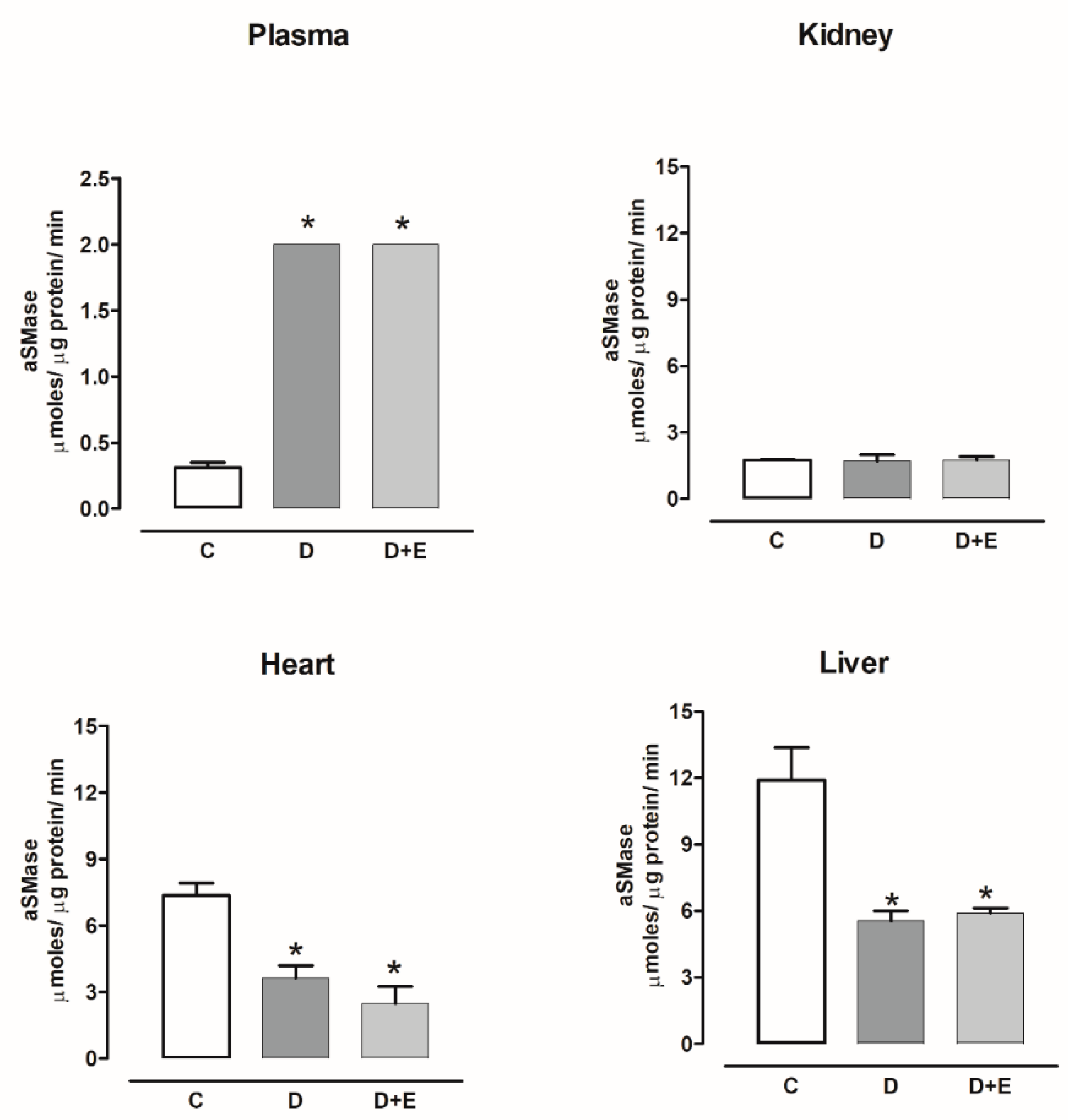
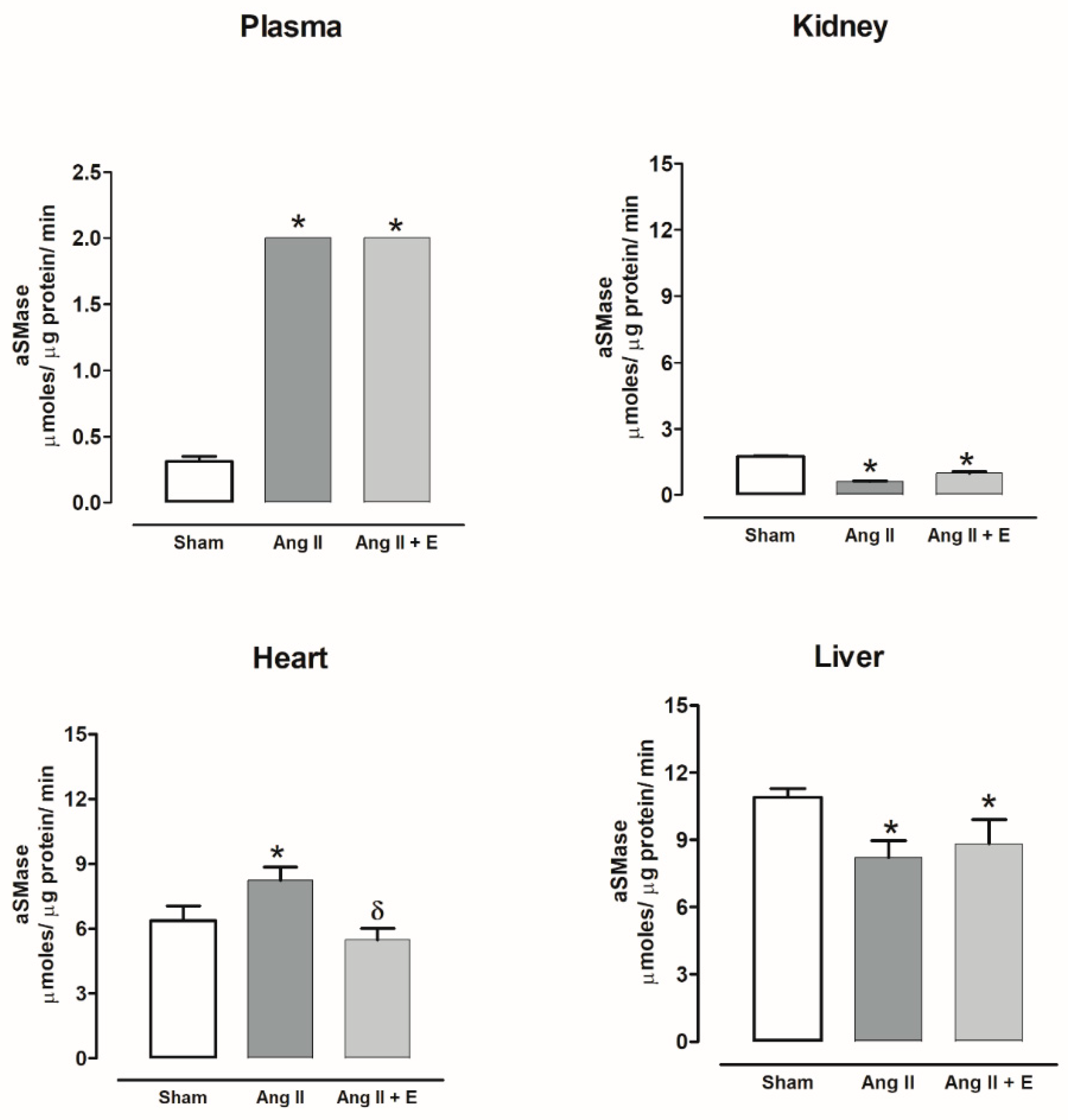
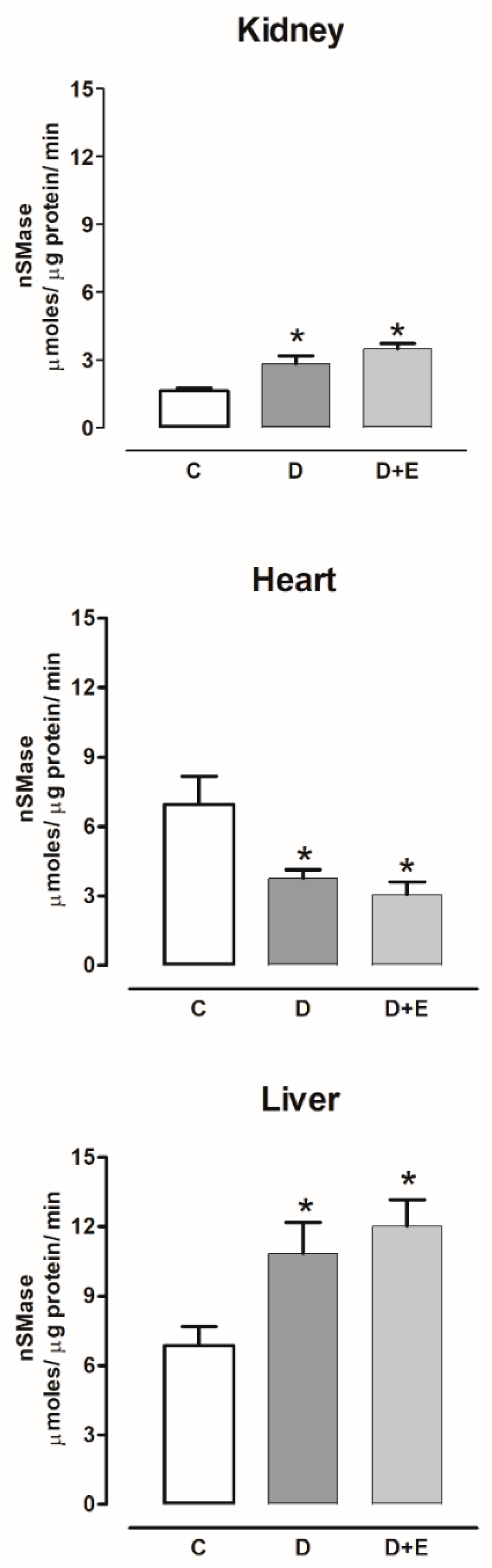
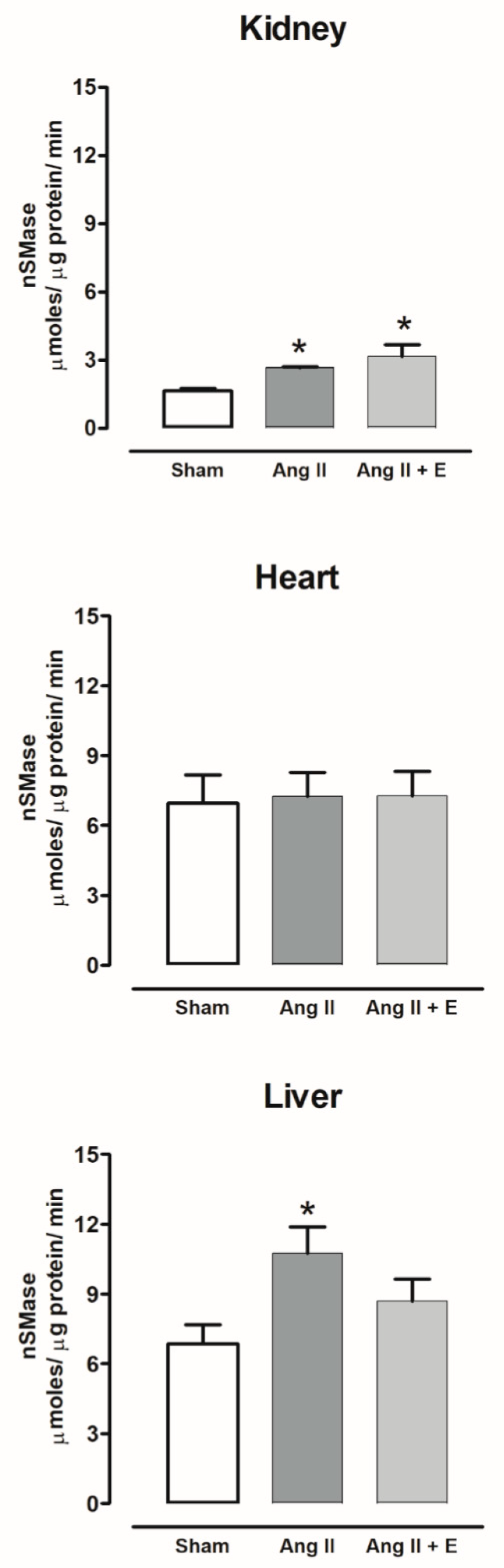
2.7. Sphingomielinases Activity in Diabetic and Hypertensive Rats
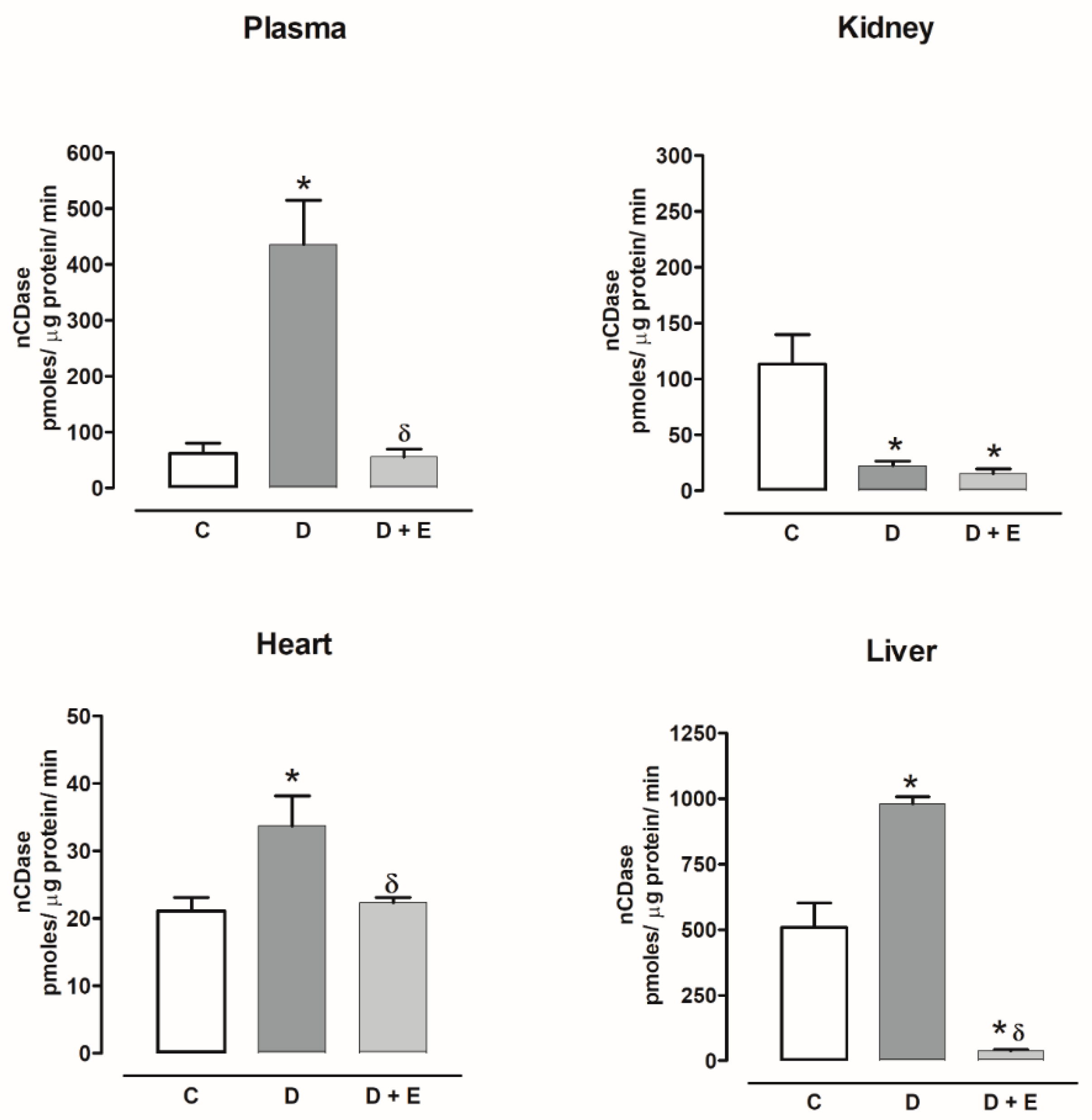
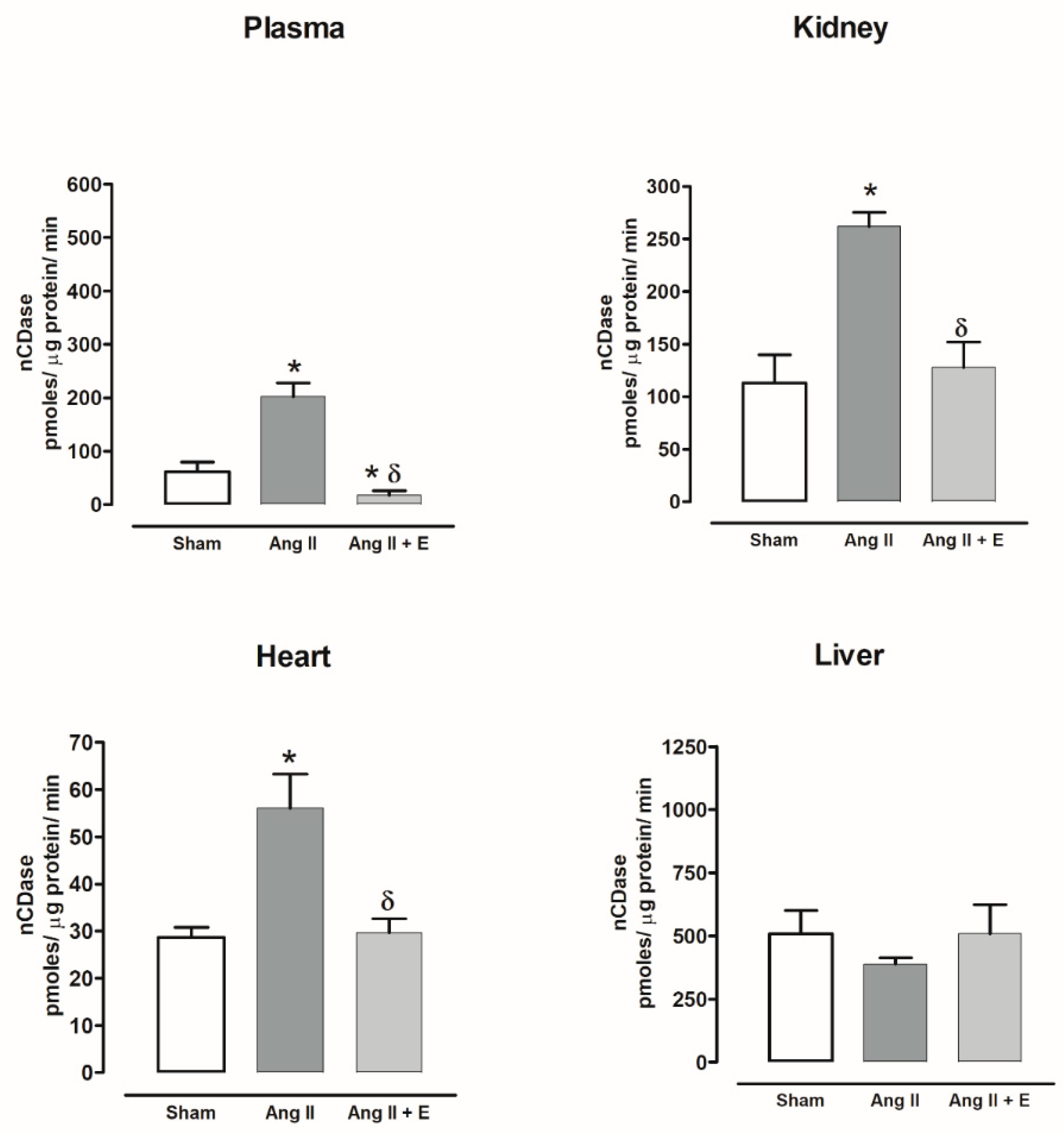
2.8. Ceramidase Activity in Diabetic and Hypertensive Rats
3. Discussion
4. Materials and Methods
4.1. Animal Models
4.1.1. Animal Procedures
4.1.2. Streptozotocin-Induced Diabetic Rats
4.1.3. Angiotensin II-Induced Hypertension Rats
4.1.4. Sample Collection and Preparation
4.2. Sphingomyelin Determination
4.3. Enzyme-Linked Immunosorbent Assay (ELISA) for Ceramide, Sphingosine, and Sphingosine-1-Phosphate Determination
4.4. Sphingomielinase Activity
4.5. Ceramidase Activity
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spijkers, L.J.; Van den Akker, R.F.; Janssen, B.J.; Debets, J.J.; De Mey, J.G.; Stroes, E.S.; Van den Born, B.J.; Wijesinghe, D.S.; Chalfant, C.E.; MacAleese, L.; et al. Hypertension is associated with marked alterations in sphingolipid biology: A potential role for ceramide. PLoS ONE 2011, 6, 217–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blachnio-Zabielska, A.U.; Koutsari, C.; Tchkonia, T.; Jensen, M.D. Sphingolipid Content of Human Adipose Tissue: Relationship to Adiponectin and Insulin Resistance. Obesity 2012, 20, 2341–2347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barlovic, D.P.; Harjutsalo, V.; Sandholm, N.; Forsblom, C.; Groop, P.-H. on behalf of the FinnDiane Study Group Sphingomyelin and progression of renal and coronary heart disease in individuals with type 1 diabetes. Diabetologia 2020, 63, 1847–1856. [Google Scholar]
- Jensen, P.N.; Fretts, A.M.; Hoofnagle, A.N.; Sitlani, C.M.; McKnight, B.; King, I.B.; Siscovick, D.S.; Psaty, B.M.; Heckbert, S.R.; Mozaffarian, D.; et al. Plasma Ceramides and Sphingomyelins in Relation to Atrial Fibrillation Risk: The Cardiovascular Health Study. J. Am. Heart Assoc. 2020, 9, e012853. [Google Scholar] [CrossRef] [PubMed]
- Haus, J.M.; Kashyap, S.R.; Kasumov, T.; Zhang, R.; Kelly, K.R.; Defronzo, R.A.; Kirwan, J.P. Plasma ceramides are elevated in obese subjects with type 2 diabetes and correlate with the severity of insulin resistance. Diabetes 2009, 58, 337–343. [Google Scholar]
- Longato, L.; Tong, M.; Wands, J.R.; de la Monte, S.M. High fat diet induced hepatic steatosis and insulin resistance: Role of dysregulated ceramide metabolism. Hepatol. Res. 2012, 42, 412–427. [Google Scholar] [CrossRef] [Green Version]
- Błachnio-Zabielska, A.; Pułka, M.; Baranowski, M.; Nikołajuk, A.; Zabielski, P.; Górska, M.; Górski, J. Ceramide metabolism is affected by obesity and diabetes in human adipose tissue. J. Cell. Physiol. 2012, 227, 550–557. [Google Scholar] [CrossRef]
- Lopez, X.; Goldfine, A.; Holland, W.L.; Gordillo, R.; Scherer, P.E. Plasma ceramides are elevated in female children and adolescents with type 2 diabetes. J. Pediatr. Endocrinol. Metab. 2013, 26, 995–998. [Google Scholar] [CrossRef]
- Mitsnefes, M.; Scherer, P.E.; Friedman, L.A.; Gordillo, R.; Furth, S.; Warady, B.A.; CKiD Study Group; The CKiD Study Group. Ceramides and cardiac function in children with chronic kidney disease. Pediatr. Nephrol. 2014, 29, 415–422. [Google Scholar] [CrossRef] [Green Version]
- Klein, R.L.; Hammad, S.M.; Baker, N.L.; Hunt, K.J.; Al Gadban, M.M.; Cleary, P.A.; Virella, G.; Lopes-Virella, M.F. Decreased plasma levels of select very long chain ceramide species Are associated with the development of nephropathy in type 1 diabetes. Metabolism 2014, 63, 1287–1295. [Google Scholar] [CrossRef] [Green Version]
- de la Maza, M.P.; Rodriguez, J.M.; Hirsch, S.; Leiva, L.; Barrera, G.; Bunout, D. Skeletal muscle ceramide species in men with abdominal obesity. J. Nutr. Health Aging 2015, 19, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Górska, M.; Dobrzyn, A.; Baranowski, M. Concentrations of sphingosine and sphinganine in plasma of patients with type 2 diabetes. Med. Sci. Monit. 2005, 11, CR35–CR38. [Google Scholar] [PubMed]
- Deutschman, D.H.; Carstens, J.S.; Klepper, R.L.; Smith, W.S.; Page, M.; Young, T.R.; Gleason, L.; Nakajima, N.; Sabbadini, R. Predicting obstructive coronary artery disease with serum sphingosine-1-phosphate. Am. Hear. J. 2003, 146, 62–68. [Google Scholar] [CrossRef]
- Kowalski, G.M.; Carey, A.L.; Selathurai, A.; Kingwell, B.A.; Bruce, C.R. Plasma Sphingosine-1-Phosphate Is Elevated in Obesity. PLoS ONE 2013, 8, e72449. [Google Scholar] [CrossRef] [Green Version]
- Hannun, Y.A.; Luberto, C.; Argraves, K.M. Enzymes of Sphingolipid Metabolism: From Modular to Integrative Signaling. Biochemistry 2001, 40, 4893–4903. [Google Scholar] [CrossRef]
- Baranowski, M.; Blachnio-Zabielska, A.; Hirnle, T.; Harasiuk, D.; Matlak, K.; Knapp, M.; Zabielski, P.; Górski, J. Myocardium of type 2 diabetic and obese patients is characterized by alterations in sphingolipid metabolic enzymes but not by accumulation of ceramide. J. Lipid Res. 2010, 51, 74–80. [Google Scholar] [CrossRef] [Green Version]
- Kolak, M.; Gertow, J.; Westerbacka, J.; Summers, A.S.; Liska, J.; Franco-Cereceda, A.; Orešič, M.; Yki-Järvinen, H.; Eriksson, P.; Fisher, R.M. Expression of ceramide-metabolising enzymes in subcutaneous and intra-abdominal human adipose tissue. Lipids Health Dis. 2012, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Pepe, G.; Cotugno, M.; Marracino, F.; Giova, S.; Capocci, L.; Forte, M.; Stanzione, R.; Bianchi, F.; Marchitti, S.; Di Pardo, A.; et al. Differential Expression of Sphingolipid Metabolizing Enzymes in Spontaneously Hypertensive Rats: A Possible Substrate for Susceptibility to Brain and Kidney Damage. Int. J. Mol. Sci. 2021, 22, 3796. [Google Scholar] [CrossRef]
- Górska, M.; Barańczuk, E.; Dobrzyń, A. Secretory Zn2+-dependent Sphingomyelinase Activity in the Serum of Patients with Type 2 Diabetes is Elevated. Horm. Metab. Res. 2003, 35, 506–507. [Google Scholar]
- Doehner, W.; Bunck, A.C.; Rauchhaus, M.; von Haehling, S.; Brunkhorst, F.M.; Cicoira, M.; Tschope, C.; Ponikowski, P.; Claus, R.A.; Anker, S.D. Secretory sphingomyelinase is upregulated in chronic heart failure: A second messenger system of immune activation relates to body composition, muscular functional capacity, and peripheral blood flow. Eur. Heart J. 2007, 28, 821–828. [Google Scholar] [CrossRef]
- Pan, W.; Yu, J.; Shi, R.; Yan, L.; Yang, T.; Li, Y.; Zhang, Z.; Yu, G.; Bai, Y.; Schuchman, E.H.; et al. Elevation of ceramide and activation of secretory acid sphingomyelinase in patients with acute coronary syndromes. Coron. Artery Dis. 2014, 25, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Heise, T.; Seman, L.J.; Macha, S.; Jones, P.; Marquart, A.; Pinnetti, S.; Woerle, H.J.; Dugi, K. Safety, Tolerability, Pharmacokinetics, and Pharmacodynamics of Multiple Rising Doses of Empagliflozin in Patients with Type 2 Diabetes Mellitus. Diabetes Ther. 2013, 4, 331–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamos, E.M.; Younk, L.M.; Davis, S.N. Empagliflozin, a sodium glucose co-transporter 2 inhibitor, in the treatment of type 1 diabetes. Expert Opin. Investig. Drugs 2014, 23, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Neeland, I.J.; Rocha, N.D.A.; Hughes, C.; Ayers, C.R.; Malloy, C.R.; Jin, E.S. Effects of Empagliflozin Treatment on Glycerol-Derived Hepatic Gluconeogenesis in Adults with Obesity: A Randomized Clinical Trial. Obesity 2020, 28, 1254–1262. [Google Scholar] [CrossRef]
- Pabel, S.; Wagner, S.; Bollenberg, H.; Bengel, P.; Kovács, Á.; Schach, C.; Tirilomis, P.; Mustroph, J.; Renner, A.; Gummert, J.; et al. Empagliflozin directly improves diastolic function in human heart failure. Eur. J. Heart Fail. 2018, 20, 1690–1700. [Google Scholar] [CrossRef]
- Tripolt, N.J.; Kolesnik, E.; Pferschy, P.N.; Verheyen, N.; Ablasser, K.; Sailer, S.; Alber, H.; Berger, R.; Kaulfersch, C.; Leitner, K.; et al. Impact of EMpagliflozin on cardiac function and biomarkers of heart failure in patients with acute MYocardial infarction—The EMMY trial. Am. Hear. J. 2020, 221, 39–47. [Google Scholar] [CrossRef]
- Han, J.H.; Oh, T.J.; Lee, G.; Maeng, H.J.; Lee, D.H.; Kim, K.M.; Choi, S.H.; Jang, H.C.; Lee, H.S.; Park, K.S.; et al. The beneficial effects of empagliflozin, an SGLT2 inhibitor, on atherosclerosis in ApoE -/- mice fed a Western diet. Diabetologia 2017, 60, 364–376. [Google Scholar] [CrossRef] [Green Version]
- Baker, W.; Buckley, L.; Kelly, M.; Bucheit, J.; Parod, E.D.; Brown, R.; Carbone, S.; Abbate, A.; Dixon, D.L. Effects of Sodium-Glucose Cotransporter 2 Inhibitors on 24-Hour Ambulatory Blood Pressure: A Systematic Review and Meta-Analysis. J. Am. Hear. Assoc. 2017, 6, e005686. [Google Scholar] [CrossRef]
- Aragón-Herrera, A.; Feijóo-Bandín, S.; Santiago, M.O.; Barral, L.; Campos-Toimil, M.; Gil-Longo, J.; Pereira, T.M.C.; García-Caballero, T.; Rodríguez-Segade, S.; Rodríguez, J.; et al. Empagliflozin reduces the levels of CD36 and cardiotoxic lipids while improving autophagy in the hearts of Zucker diabetic fatty rats. Biochem. Pharmacol. 2019, 170, 113677. [Google Scholar] [CrossRef]
- Bautista-Pérez, R.; Arellano, A.; Franco, M.; Osorio, H.; Coronel, I. Sphingosine-1-phosphate induced vasoconstriction is increased in the isolated perfused kidneys of diabetic rats. Diabetes Res. Clin. Pr. 2011, 94, e8–e11. [Google Scholar] [CrossRef]
- Bautista-Pérez, R.; Del Valle-Mondragón, L.; Cano-Martínez, A.; Pérez-Méndez, O.; Escalante, B.; Franco, M. Involvement of neutral sphingomyelinase in the angiotensin II signaling pathway. Am. J. Physiol. Physiol. 2015, 308, F1178–F1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bangalore, S.; Fakheri, R.; Wandel, S.; Toklu, B.; Wandel, J.; Messerli, F.H. Renin angiotensin system inhibitors for patients with stable coronary artery disease without heart failure: Systematic review and meta-analysis of randomized trials. BMJ 2017, 356, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Hu, J.; Luo, T.; Wang, Y.; Yang, S.; Qing, H.; Cheng, Q.; Li, Q. Effects of Angiotensin-Converting Enzyme Inhibitors and Angiotensin II Receptor Blockers on All-Cause Mortality and Renal Outcomes in Patients with Diabetes and Albuminuria: A Systematic Review and Meta-Analysis. Kidney Blood Press. Res. 2018, 43, 768–779. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Ghani, M.A.; Norton, L.; DeFronzo, R.A. Role of Sodium-Glucose Cotransporter 2 (SGLT 2) Inhibitors in the Treatment of Type 2 Diabetes. Endocr. Rev. 2011, 32, 515–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, S.J.; Chung, S.; Kim, S.J.; Lee, E.-M.; Yoo, Y.-H.; Kim, J.-W.; Ahn, Y.-B.; Kim, E.-S.; Moon, S.-D.; Kim, M.-J.; et al. Effect of Sodium-Glucose Co-Transporter 2 Inhibitor, Dapagliflozin, on Renal Renin-Angiotensin System in an Animal Model of Type 2 Diabetes. PLoS ONE 2016, 11, e0165703. [Google Scholar] [CrossRef]
- Vallon, V.; Gerasimova, M.; Rose, M.A.; Masuda, T.; Satriano, J.; Mayoux, E.; Koepsell, H.; Thomson, S.C.; Rieg, T. SGLT2 inhibitor empagliflozin reduces renal growth and albuminuria in proportion to hyperglycemia and prevents glomerular hyperfiltration in diabetic Akita mice. Am. J. Physiol. Renal Physiol. 2014, 306, 194–204. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.T.; Chen, L.; Li, S.Y.; Mayoux, E.; Leung, P.S. The Effects of Empagliflozin, an SGLT2 Inhibitor, on Pancreatic β-Cell Mass and Glucose Homeostasis in Type 1 Diabetes. PLoS ONE 2016, 11, e0147391. [Google Scholar] [CrossRef] [Green Version]
- Castoldi, G.; Carletti, R.; Ippolito, S.; Colzani, M.; Barzaghi, F.; Stella, A.; Zerbini, G.; Perseghin, G.; Di Gioia, C.R. Renal Anti-Fibrotic Effect of Sodium Glucose Cotransporter 2 Inhibition in Angiotensin II-Dependent Hypertension. Am. J. Nephrol. 2020, 51, 119–129. [Google Scholar] [CrossRef]
- Fox, T.E.; Bewley, M.C.; Unrath, K.A.; Pedersen, M.M.; Anderson, R.E.; Jung, D.Y.; Jefferson, L.S.; Kim, J.K.; Bronson, S.K.; Flanagan, J.M.; et al. Circulating sphingolipid biomarkers in models of type 1 diabetes. J. Lipid Res. 2011, 52, 509–517. [Google Scholar] [CrossRef] [Green Version]
- Samad, F.; Hester, K.D.; Yang, G.; Hannun, Y.A.; Bielawski, J. Altered Adipose and Plasma Sphingolipid Metabolism in Obesity. Diabetes 2006, 55, 2579–2587. [Google Scholar] [CrossRef] [Green Version]
- Ohanian, J.; Liao, A.; Forman, S.P.; Ohanian, V. Age-related remodeling of small arteries is accompanied by increased sphingomyelinase activity and accumulation of long-chain ceramides. Physiol. Rep. 2014, 2, e12015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babenko, N.A.; Shakhova, E.G. Effects of Chamomilla recutita flavonoids on age-related liver sphingolipid turnover in rats. Exp. Gerontol. 2006, 41, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Babenko, N.A.; Shakhova, E.G. Long-term food restriction prevents aging-associated sphingolipid turnover dysregulation in the brain. Arch. Gerontol. Geriatr. 2014, 58, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Sacket, S.J.; Chung, H.-Y.; Okajima, F.; Im, D.-S. Increase in sphingolipid catabolic enzyme activity during aging. Acta Pharmacol. Sin. 2009, 30, 1454–1461. [Google Scholar] [CrossRef] [Green Version]
- Empinado, H.M.; Deevska, G.M.; Nikolova-Karakashian, M.; Yoo, J.-K.; Christou, D.D.; Ferreira, L.F. Diaphragm dysfunction in heart failure is accompanied by increases in neutral sphingomyelinase activity and ceramide content. Eur. J. Hear. Fail. 2014, 16, 519–525. [Google Scholar] [CrossRef] [Green Version]
- Boini, K.M.; Xia, M.; Koka, S.; Gehr, T.W.; Li, P.-L. Instigation of NLRP3 inflammasome activation and glomerular injury in mice on the high fat diet: Role of acid sphingomyelinase gene. Oncotarget 2016, 7, 19031–19044. [Google Scholar] [CrossRef] [Green Version]
- Geoffroy, K.; Troncy, L.; Wiernsperger, N.; Lagarde, M.; El Bawab, S. Glomerular proliferation during early stages of diabetic nephropathy is associated with local increase of sphingosine-1-phosphate levels. FEBS Lett. 2005, 579, 1249–1254. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhang, H.; Li, Z.; Hailemariam, T.K.; Chakraborty, M.; Jiang, K.; Qiu, D.; Bui, H.H.; Peake, D.A.; Kuo, M.S.; et al. Sphingomyelin synthase 2 is one of the determinants for plasma and liver sphingomyelin levels in mice. Arterioscler Thromb Vasc Biol. 2009, 29, 850–856. [Google Scholar] [CrossRef] [Green Version]
- Becker, K.P.; Kitatani, K.; Idkowiak-Baldys, J.; Bielawski, J.; Hannun, Y.A. Selective inhibition of juxtanuclear translocation of protein kinase C beta II by a negative feedback mechanism involving ceramide formed from the salvage pathway. J. Biol. Chem. 2005, 280, 2606–2612. [Google Scholar] [CrossRef] [Green Version]
- Inoue, T.; Okino, N.; Kakuta, Y.; Hijikata, A.; Okano, H.; Goda, H.M.; Tani, M.; Sueyoshi, N.; Kambayashi, K.; Matsumura, H.; et al. Mechanistic Insights into the Hydrolysis and Synthesis of Ceramide by Neutral Ceramidase. J. Biol. Chem. 2009, 284, 9566–9577. [Google Scholar] [CrossRef] [Green Version]
- Novgorodov, S.A.; Wu, B.X.; Gudz, T.I.; Bielawski, J.; Ovchinnikova, T.V.; Hannun, Y.A.; Obeid, L.M. Novel pathway of ceramide production in mitochondria: Thioesterase and neutral ceramidase produce ceramide from sphingosine and acyl-CoA. J. Biol. Chem. 2011, 286, 25352–25362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobczak, A.I.; Pitt, S.J.; Smith, T.K.; Ajjan, R.A.; Stewart, A.J. Lipidomic profiling of plasma free fatty acids in type-1 diabetes highlights specific changes in lipid metabolism. Biochim. et Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2021, 1866, 158823. [Google Scholar] [CrossRef] [PubMed]
- Spijkers, L.J.A.; Janssen, B.J.A.; Nelissen, J.; Meens, M.J.P.M.T.; Wijesinghe, D.; Chalfant, C.E.; De Mey, J.G.R.; Alewijnse, A.E.; Peters, S.L.M. Antihypertensive Treatment Differentially Affects Vascular Sphingolipid Biology in Spontaneously Hypertensive Rats. PLoS ONE 2011, 6, e29222. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Pardo, H.; Bautista, R.; Vargas-Robles, H.; Rios, A.; Sanchez, D.; Escalante, B. Role of sodium/glucose cotransporter inhibition on a rat model of angiotensin II–dependent kidney damage. BMC Nephrol. 2019, 20, 1–9. [Google Scholar] [CrossRef]
- Ozawa, Y.; Kobori, H.; Suzaki, Y.; Navar, L.G. Sustained renal interstitial macrophage infiltration following chronic angiotensin II infusions. Am. J. Physiol. Physiol. 2007, 292, F330–F339. [Google Scholar] [CrossRef]
- Ruiz-Ortega, M.; Ruperez, M.; Lorenzo, O.; Esteban, V.; Blanco, J.; Mezzano, S.; Egido, J. Angiotensin II regulates the synthesis of proinflammatory cytokines and chemokines in the kidney. Kidney Int. 2002, 62, S12–S22. [Google Scholar] [CrossRef] [Green Version]
- Zhuo, J.L. Monocyte chemoattractant protein-1: A key mediator of angiotensin II-induced target organ damage in hypertensive heart disease? J. Hypertens. 2004, 22, 451–454. [Google Scholar] [CrossRef]
- Bautista-Pérez, R.; Pérez-Méndez, O.; Cano-Martínez, A.; Pacheco, U.; Santamaría, J.; Rodríguez-Iturbe, F.R.B.; Navar, L.G.; Franco, M. The Role of P2X7 Purinergic Receptors in the Renal Inflammation Associated with Angiotensin II-induced Hypertension. Int. J. Mol. Sci. 2020, 21, 4041. [Google Scholar] [CrossRef]
- Devaraj, S.; Glaser, N.; Griffen, S.; Wang-Polagruto, J.; Miguelino, E.; Jialal, I. Increased Monocytic Activity and Biomarkers of Inflammation in Patients with Type 1 Diabetes. Diabetes 2006, 55, 774–779. [Google Scholar] [CrossRef] [Green Version]
- Hofmeister, R.; Wiegmann, K.; Korherr, C.; Bernardo, K.; Krönke, M.; Falk, W. Activation of Acid Sphingomyelinase by Interleukin-1 (IL-1) Requires the IL-1 Receptor Accessory Protein. J. Biol. Chem. 1997, 272, 27730–27736. [Google Scholar] [CrossRef] [Green Version]
- Wiegmann, K.; Schütze, S.; Machleidt, T.; Witte, D.; Krönke, M. Functional dichotomy of neutral and acidic sphingomyelinases in tumor necrosis factor signaling. Cell 1994, 78, 1005–1015. [Google Scholar] [CrossRef]
- Coroneos, E.; Martinez, M.; McKenna, S.; Kester, M. Differential regulation of sphingomyelinase and ceramidase activities by growth factors and cytokines. Implications for cellular proliferation and differentiation. J. Biol. Chem. 1995, 270, 23305–23309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oelze, M.; Kröller-Schön, S.; Welschof, P.; Jansen, T.; Hausding, M.; Mikhed, Y.; Stamm, P.; Mader, M.; Zinßius, E.; Agdauletova, S.; et al. The Sodium-Glucose Co-Transporter 2 Inhibitor Empagliflozin Improves Diabetes-Induced Vascular Dysfunction in the Streptozotocin Diabetes Rat Model by Interfering with Oxidative Stress and Glucotoxicity. PLoS ONE 2014, 9, e112394. [Google Scholar] [CrossRef] [PubMed]
- Hojjati, M.R.; Jiang, X.-C. Rapid, specific, and sensitive measurements of plasma sphingomyelin and phosphatidylcholine. J. Lipid Res. 2006, 47, 673–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez-Camacho, I.; Bautista-Pérez, R.; Correa, F.; Buelna-Chontal, M.; Román-Anguiano, N.G.; Medel-Franco, M.; Medina-Campos, O.N.; Pedraza-Chaverri, J.; Cano-Martínez, A.; Zazueta, C. Role of sphingomyelinase in mitocondrial ceramide accumulation during reperfusion. Biochim. Biophys. Acta 2016, 1862, 1955–1963. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, N.; Jones, S.T.; Williams, D.G.; Cunningham, H.B.; Moreno, K.; Visentin, B.; Gentile, A.; Vekich, J.; Shestowsky, W.; Hiraiwa, M.; et al. Production and characterization of monoclonal anti-sphingosine-1-phosphate antibodies. J Lipid Res. 2009, 50, 2245–2257. [Google Scholar] [CrossRef] [Green Version]
- Cai, B.B.; Lu, Y.N.; Xu, M. Acid sphingomyelinase downregulation alleviates vascular endothelial leptin resistance in rats. Acta Pharmacol. Sin. 2020, 41, 650–660. [Google Scholar] [CrossRef]
- Casasampere, M.; Camacho, L.; Cingolani, F.; Casas, J.; Egido-Gabás, M.; Abad, J.L.; Bedia, C.; Xu, R.; Wang, K.; Canals, D.; et al. Activity of neutral and alkaline ceramidases on fluorogenic N-acylated coumarin-containing aminodiols. J. Lipid Res. 2015, 56, 2019–2028. [Google Scholar] [CrossRef] [Green Version]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Diabetic | Diabetic + Empagliflozin | ||||
|---|---|---|---|---|---|---|
| Basal | Final | Basal | Final | Basal | Final | |
| Body weight (g) | 315 ± 15 | 330 ± 10 | 325 ± 8 | 250 ± 18 * | 330 ± 10 | 294 ± 6.5 * |
| Blood glucose (mg/dL) | 102 ± 8 | 113 ± 2.5 | 106 ± 4 | 589 ± 34 * | 104 ± 6 | 206 ± 43 * |
| Urine volume (mL/24 h) | 12 ± 3 | 12 ± 8 | 15 ± 2 | 141 ± 4.2 * | 14 ± 3 | 50.3 ± 0.7 * |
| Sham | Ang II | Ang II + Empagliflozin | ||||
|---|---|---|---|---|---|---|
| Basal | Final | Basal | Final | Basal | Final | |
| Body weight (g) | 324 ± 23 | 332 ± 20 | 352 ± 7 | 303 ± 6 * | 341 ± 18 | 321 ± 13 |
| Blood glucose (mg/dL) | 114 ± 5 | 120 ± 7 | 116 ± 4 | 122 ± 11 | 108 ± 8 | 118 ± 18 |
| Urine volume (mL/24 hrs) | 12.5 ± 4 | 13.3 ± 9.2 | 14.4 ± 5.11 | 45 ± 8.1 * | 45 ± 12 | 40 ± 14 |
| Blood pressure | 120 ± 5 | 122 ± 6.6 | 125 ± 4.51 | 195 ± 22 * | 119 ± 3 | 195 ± 12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Villavicencio, R.; Flores-Estrada, J.; Franco, M.; Escalante, B.; Pérez-Méndez, O.; Mercado, A.; Bautista-Pérez, R. Effect of Empagliflozin on Sphingolipid Catabolism in Diabetic and Hypertensive Rats. Int. J. Mol. Sci. 2022, 23, 2883. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052883
Pérez-Villavicencio R, Flores-Estrada J, Franco M, Escalante B, Pérez-Méndez O, Mercado A, Bautista-Pérez R. Effect of Empagliflozin on Sphingolipid Catabolism in Diabetic and Hypertensive Rats. International Journal of Molecular Sciences. 2022; 23(5):2883. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052883
Chicago/Turabian StylePérez-Villavicencio, Roxana, Javier Flores-Estrada, Martha Franco, Bruno Escalante, Oscar Pérez-Méndez, Adriana Mercado, and Rocio Bautista-Pérez. 2022. "Effect of Empagliflozin on Sphingolipid Catabolism in Diabetic and Hypertensive Rats" International Journal of Molecular Sciences 23, no. 5: 2883. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052883