
Identification of Novel Circulating miRNAs in Patients with Acute Ischemic Stroke

 , ,
, ,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
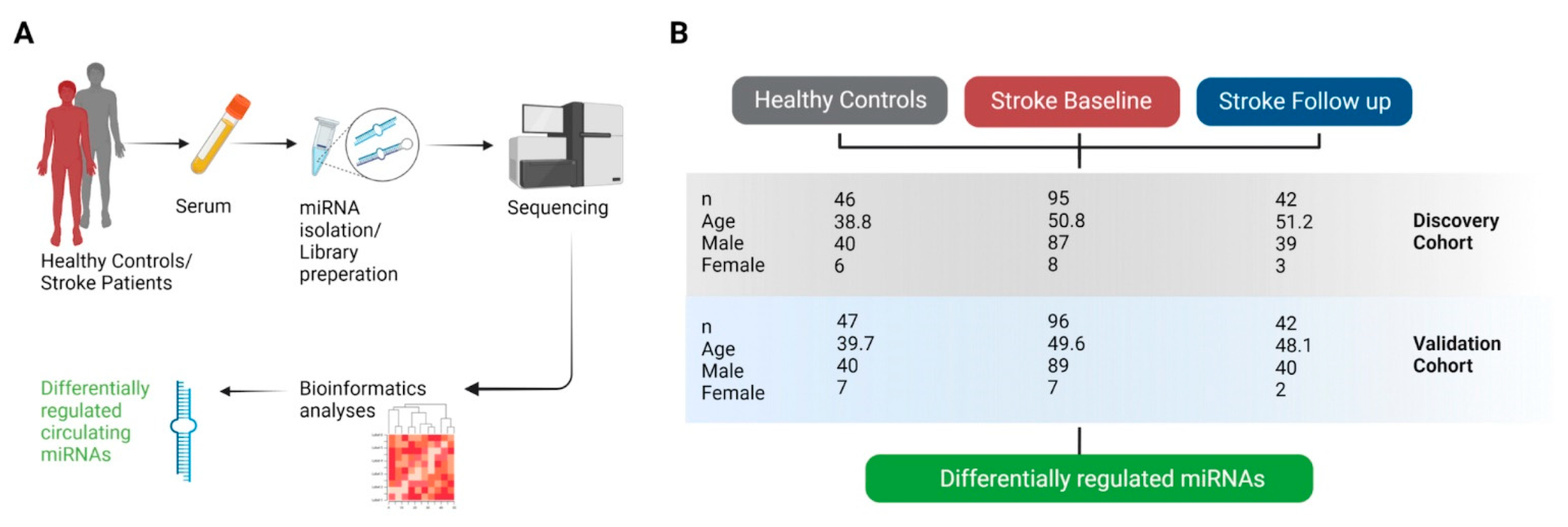
2.1. Study Design
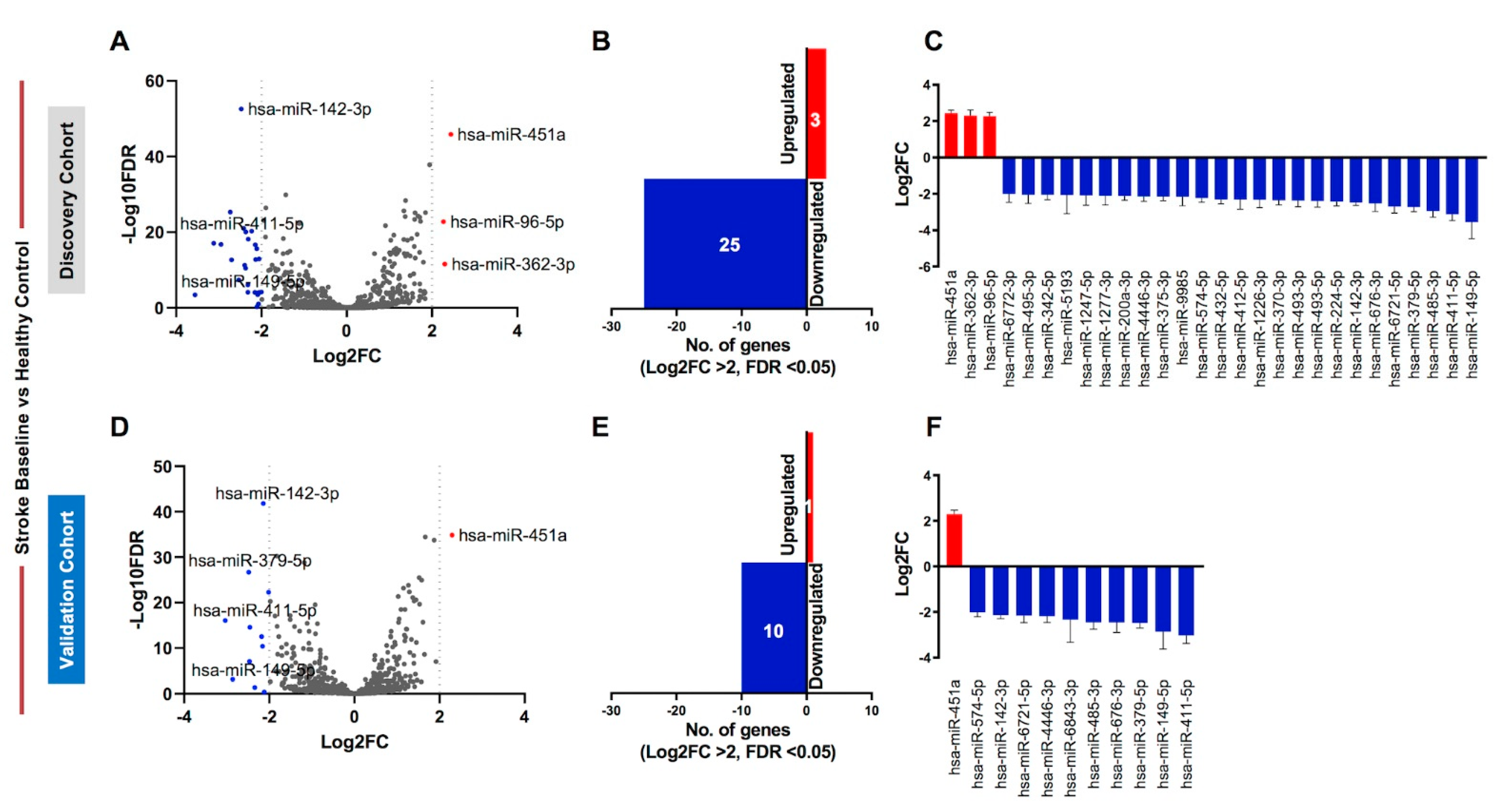
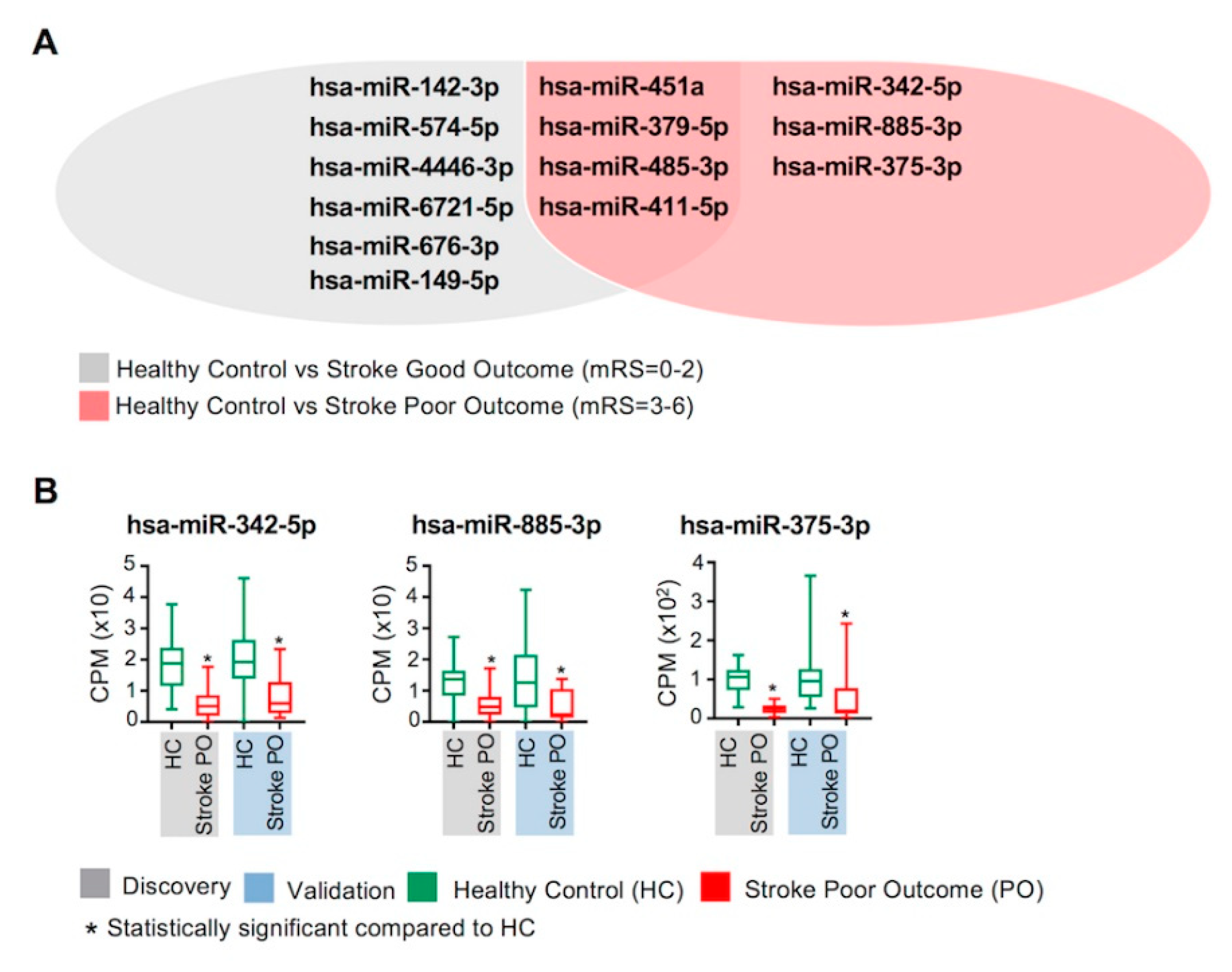
2.2. Identification of Differentially Regulated Circulating miRNAs in Discovery and Validation Cohorts
2.3. Validated Differentially Regulated Circulating miRNAs in Stroke BL Patients Compared with Healthy Controls
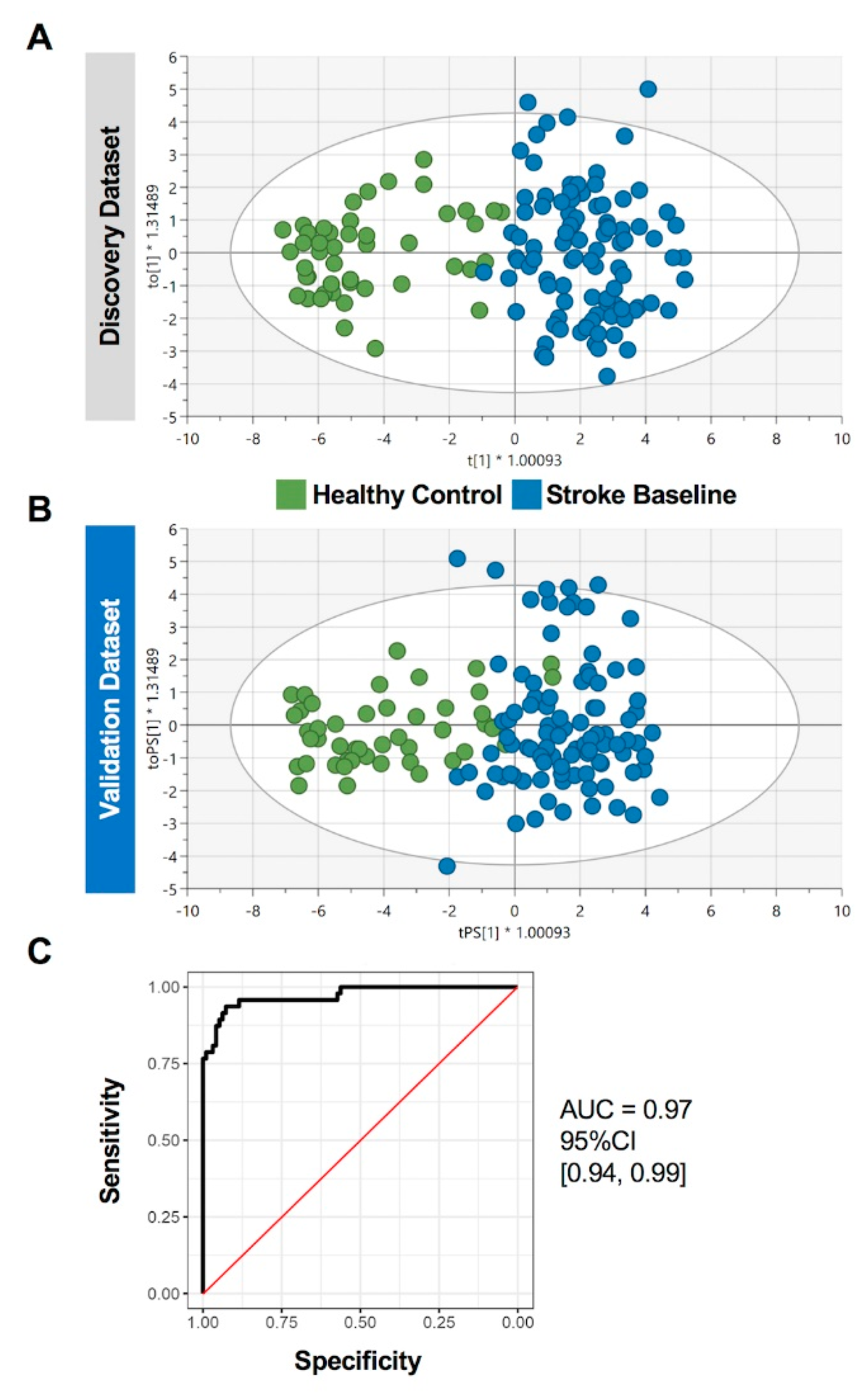
2.4. Prediction Performance of the Identified Circulating miRNAs in Stroke Patients
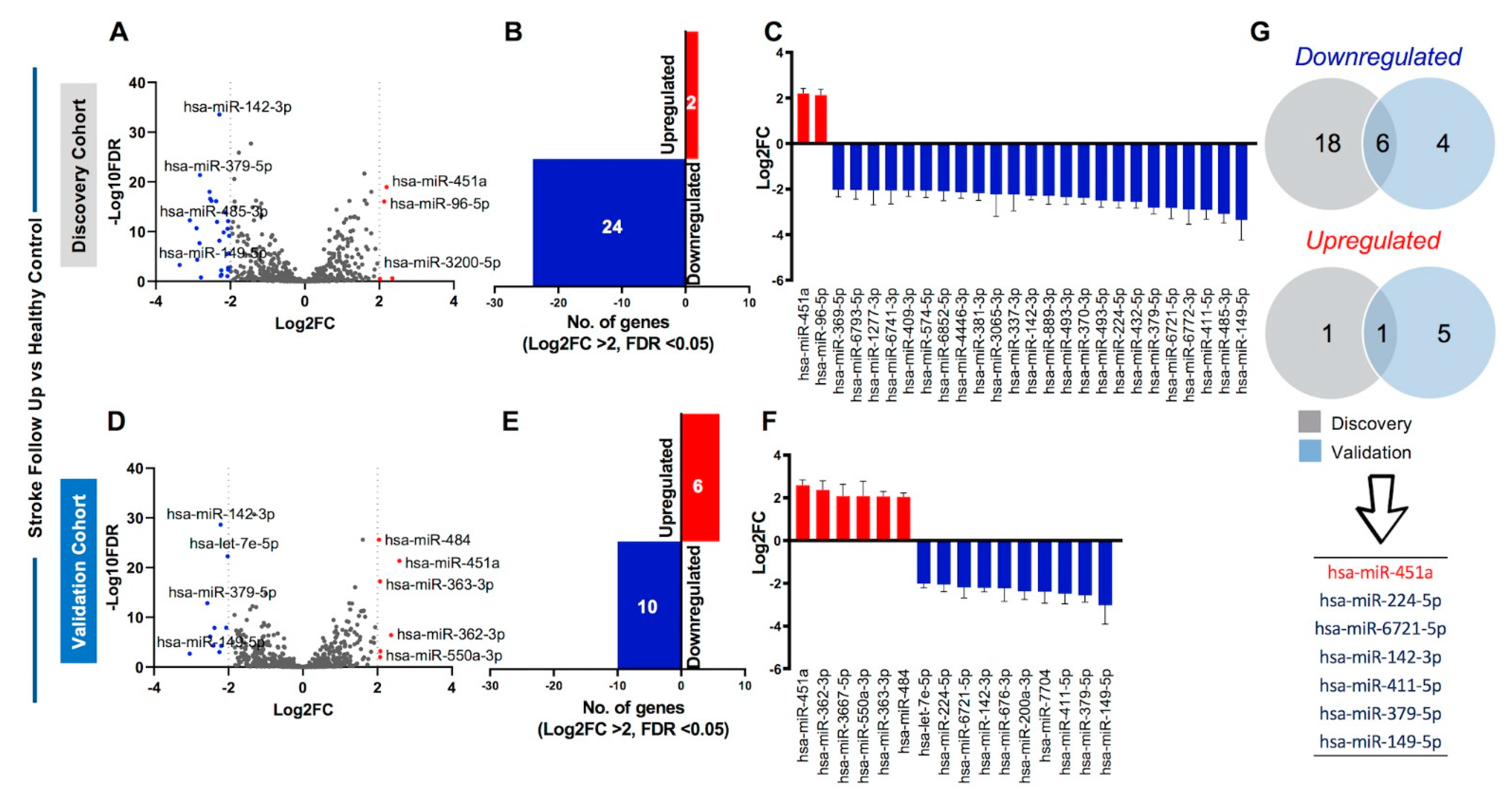
2.5. Identification of Sustained Dysregulation of Circulating miRNAs in Stroke Patients
2.6. Circulating miRNAs and Clinical Outcomes of Stroke
3. Discussion
4. Materials and Methods
4.1. Samples
4.2. Circulating miRNA Purification, Library Preparation, and Sequencing
4.3. RNA-Seq Data Processing and Analyses
4.4. Discriminant Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Katan, M.; Luft, A. Global Burden of Stroke. Semin. Neurol. 2018, 38, 208–211. [Google Scholar] [CrossRef] [Green Version]
- Boehme, A.K.; Esenwa, C.; Elkind, M.S.V. Stroke Risk Factors, Genetics, and Prevention. Circ. Res. 2017, 120, 472–495. [Google Scholar] [CrossRef]
- Sekerdag, E.; Solaroglu, I.; Gürsoy-Ozdemir, Y. Cell Death Mechanisms in Stroke and Novel Molecular and Cellular Treatment Options. Curr. Neuropharmacol. 2018, 16, 1396–1415. [Google Scholar] [CrossRef]
- Adams, H.P., Jr.; Bendixen, B.H.; Kappelle, L.J.; Biller, J.; Love, B.B.; Gordon, D.L.; Marsh, E.E., 3rd. Classification of subtype of acute ischemic stroke. Definitions for use in a multicenter clinical trial. TOAST. Trial of Org 10172 in Acute Stroke Treatment. Stroke 1993, 24, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Chen, J.; Sen, S. MicroRNA as Biomarkers and Diagnostics. J. Cell. Physiol. 2016, 231, 25–30. [Google Scholar] [CrossRef]
- Dagonnier, M.; Donnan, G.A.; Davis, S.M.; Dewey, H.M.; Howells, D.W. Acute Stroke Biomarkers: Are We There Yet? Front. Neurol. 2021, 12, 619721. [Google Scholar] [CrossRef]
- Misra, S.; Montaner, J.; Ramiro, L.; Arora, R.; Talwar, P.; Nath, M.; Kumar, A.; Kumar, P.; Pandit, A.K.; Mohania, D.; et al. Blood biomarkers for the diagnosis and differentiation of stroke: A systematic review and meta-analysis. Int. J. Stroke 2020, 15, 704–721. [Google Scholar] [CrossRef]
- Dewdney, B.; Trollope, A.; Moxon, J.; Manapurathe, D.T.; Biros, E.; Golledge, J. Circulating MicroRNAs as Biomarkers for Acute Ischemic Stroke: A Systematic Review. J. Stroke Cerebrovasc. Dis. 2018, 27, 522–530. [Google Scholar] [CrossRef]
- Hill, M.D.; Yiannakoulias, N.; Jeerakathil, T.; Tu, J.V.; Svenson, L.W.; Schopflocher, D.P. The high risk of stroke immediately after transient ischemic attack: A population-based study. Neurology 2004, 62, 2015–2020. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, X. miRDB: An online database for prediction of functional microRNA targets. Nucleic Acids Res. 2020, 48, D127–D131. [Google Scholar] [CrossRef] [Green Version]
- Giordano, M.; Ciarambino, T.; D’Amico, M.; Trotta, M.C.; Di Sette, A.M.; Marfella, R.; Malatino, L.; Paolisso, G.; Adinolfi, L.E. Circulating MiRNA-195-5p and -451a in Transient and Acute Ischemic Stroke Patients in an Emergency Department. J. Clin. Med. 2019, 8, 130. [Google Scholar] [CrossRef] [Green Version]
- Kong, Y.; Li, S.; Cheng, X.; Ren, H.; Zhang, B.; Ma, H.; Li, M.; Zhang, X.-A. Brain Ischemia Significantly Alters microRNA Expression in Human Peripheral Blood Natural Killer Cells. Front. Immunol. 2020, 11, 759. [Google Scholar] [CrossRef]
- Begum, G.; Yuan, H.; Kahle, K.T.; Li, L.; Wang, S.; Shi, Y.; Shmukler, B.E.; Yang, S.-S.; Lin, S.-H.; Alper, S.L.; et al. Inhibition of WNK3 Kinase Signaling Reduces Brain Damage and Accelerates Neurological Recovery After Stroke. Stroke 2015, 46, 1956–1965. [Google Scholar] [CrossRef] [Green Version]
- Sinner, M.F.; Tucker, N.R.; Lunetta, K.L.; Ozaki, K.; Smith, J.D.; Trompet, S.; Bis, J.C.; Lin, H.; Chung, M.K.; Nielsen, J.B.; et al. Integrating Genetic, Transcriptional, and Functional Analyses to Identify 5 Novel Genes for Atrial Fibrillation. Circulation 2014, 130, 1225–1235. [Google Scholar] [CrossRef] [Green Version]
- Traylor, M.; Farrall, M.; Holliday, E.G.; Sudlow, C.; Hopewell, J.C.; Cheng, Y.-C.; Fornage, M.; Ikram, M.A.; Malik, R.; Bevan, S.; et al. Genetic risk factors for ischaemic stroke and its subtypes (the METASTROKE Collaboration): A meta-analysis of genome-wide association studies. Lancet Neurol. 2012, 11, 951–962. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zhang, X.; Wang, Y.; Lei, H.; Su, H.; Zeng, J.; Pei, Z.; Huang, R. Inhibition of immunoproteasome reduces infarction volume and attenuates inflammatory reaction in a rat model of ischemic stroke. Cell Death Dis. 2015, 6, e1626. [Google Scholar] [CrossRef] [Green Version]
- Sepramaniam, S.; Tan, J.-R.; Tan, K.-S.; DeSilva, D.A.; Tavintharan, S.; Woon, F.-P.; Wang, C.-W.; Yong, F.-L.; Karolina, D.-S.; Kaur, P.; et al. Circulating MicroRNAs as Biomarkers of Acute Stroke. Int. J. Mol. Sci. 2014, 15, 1418–1432. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.-H.; Ko, T.-M.; Chen, C.-H.; Lee, M.-T.M.; Chang, Y.-J.; Chang, C.-H.; Huang, K.-L.; Chang, T.-Y.; Lee, J.-D.; Chang, K.-C.; et al. Identification of PTCSC3 as a Novel Locus for Large--Vessel Ischemic Stroke: A Genome--Wide Association Study. J. Am. Heart Assoc. 2016, 5, e003003. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.S.; Armugam, A.; Sepramaniam, S.; Lim, K.Y.; Setyowati, K.D.; Wang, C.W.; Jeyaseelan, K. Expression Profile of MicroRNAs in Young Stroke Patients. PLoS ONE 2009, 4, e7689. [Google Scholar] [CrossRef] [Green Version]
- Vivinetto, A.L.; Kim, I.-D.; Goldberg, D.C.; Fones, L.; Brown, E.; Tarabykin, V.S.; Hill, C.E.; Cho, S.; Cave, J.W. Zeb2 Is a Regulator of Astrogliosis and Functional Recovery after CNS Injury. Cell Rep. 2020, 31, 107834. [Google Scholar] [CrossRef]
- Xiang, W.; Tian, C.; Lin, J.; Wu, X.; Pang, G.; Zhou, L.; Pan, S.; Deng, Z. Plasma let-7i and miR-15a expression are associated with the effect of recombinant tissue plasminogen activator treatment in acute ischemic stroke patients. Thromb. Res. 2017, 158, 121–125. [Google Scholar] [CrossRef]
- Mo, Y.; Sun, Y.; Yue, E.; Liu, Y.; Liu, K. MicroRNA --379--5p targets MAP3K2 to reduce autophagy and alleviate neuronal injury following cerebral ischemia via the JNK/c--Jun signaling pathway. Kaohsiung J. Med. Sci. 2022, 38, 230–243. [Google Scholar] [CrossRef]
- Yu, Y.L.; Chou, R.H.; Shyu, W.C.; Hsieh, S.C.; Wu, C.S.; Chiang, S.Y.; Chang, W.J.; Chen, J.N.; Tseng, Y.J.; Lin, Y.H.; et al. Smurf2--mediated degradation of EZH2 enhances neuron differentiation and improves functional recovery after ischaemic stroke. EMBO Mol. Med. 2013, 5, 531–547. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Lv, Z.; Guo, Y.; Li, L.; Zhang, Y.; Zhou, L.; Yang, B.; Wu, S.; Zhang, Y.; Xie, C.; et al. Identification of Blood Let-7e-5p as a Biomarker for Ischemic Stroke. PLoS ONE 2016, 11, e0163951. [Google Scholar] [CrossRef] [PubMed]
- Dongsheng, H.; Zhuo, Z.; Jiamin, L.; Hailan, M.; Lijuan, H.; Fan, C.; Dan, Y.; He, Z.; Yun, X. Proteomic Analysis of the Peri-Infarct Area after Human Umbilical Cord Mesenchymal Stem Cell Transplantation in Experimental Stroke. Aging Dis. 2016, 7, 623–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senis, Y.A.; Barr, A.J. Targeting Receptor-Type Protein Tyrosine Phosphatases with Biotherapeutics: Is Outside-in Better than Inside-Out? Molecules 2018, 23, 569. [Google Scholar] [CrossRef] [Green Version]
- Gurnik, S.; Devraj, K.; Macas, J.; Yamaji, M.; Starke, J.; Scholz, A.; Sommer, K.; Di Tacchio, M.; Vutukuri, R.; Beck, H.; et al. Angiopoietin-2-induced blood–brain barrier compromise and increased stroke size are rescued by VE-PTP-dependent restoration of Tie2 signaling. Acta Neuropathol. 2016, 131, 753–773. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Chen, J.; Chen, J.; Liu, X.; Yang, H.; Liu, J.; He, A.; Gao, X.; Xin, Y. KIF 2 mediates the neuroprotection in cerebral ischaemia injury by affecting NF--κB pathway. Clin. Exp. Pharmacol. Physiol. 2020, 47, 274–280. [Google Scholar] [CrossRef]
- Fan, J.-R.; Lee, H.-T.; Lee, W.; Lin, C.-H.; Hsu, C.Y.; Hsieh, C.-H.; Shyu, W.-C. Potential role of CBX7 in regulating pluripotency of adult human pluripotent-like olfactory stem cells in stroke model. Cell Death Dis. 2018, 9, 502. [Google Scholar] [CrossRef]
- Daude, N.; Gapeshina, H.; Dong, B.; Winship, I.; Westaway, D. Neuroprotective properties of the PrP-like Shadoo glycoprotein assessed in the middle cerebral artery occlusion model of ischemia. Prion 2015, 9, 376–393. [Google Scholar] [CrossRef] [Green Version]
- Joladarashi, D.; Garikipati, V.N.S.; Thandavarayan, R.A.; Verma, S.K.; Mackie, A.R.; Khan, M.; Gumpert, A.M.; Bhimaraj, A.; Youker, K.A.; Uribe, C.; et al. Enhanced Cardiac Regenerative Ability of Stem Cells After Ischemia-Reperfusion Injury: Role of Human CD34+ Cells Deficient in MicroRNA-377. J. Am. Coll. Cardiol. 2015, 66, 2214–2226. [Google Scholar] [CrossRef] [Green Version]
- Beck, J.A.; Campbell, T.A.; Adamson, G.; Poulter, M.; Uphill, J.B.; Molou, E.; Collinge, J.; Mead, S. Association of a null allele of SPRN with variant Creutzfeldt-Jakob disease. J. Med. Genet. 2008, 45, 813–817. [Google Scholar] [CrossRef] [PubMed]
- Goyal, P.; Behring, A.; Kumar, A.; Siess, W. STK35L1 Associates with Nuclear Actin and Regulates Cell Cycle and Migration of Endothelial Cells. PLoS ONE 2011, 6, e16249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinciguerra, A.; Cepparulo, P.; Anzilotti, S.; Cuomo, O.; Valsecchi, V.; Amoroso, S.; Annunziato, L.; Pignataro, G. Remote postconditioning ameliorates stroke damage by preventing let-7a and miR-143 up-regulation. Theranostics 2020, 10, 12174–12188. [Google Scholar] [CrossRef] [PubMed]
- Scrutinio, D.; Conserva, F.; Passantino, A.; Iacoviello, M.; Lagioia, R.; Gesualdo, L. Circulating microRNA-150-5p as a novel biomarker for advanced heart failure: A genome-wide prospective study. J. Heart Lung Transplant. 2017, 36, 616–624. [Google Scholar] [CrossRef] [Green Version]
- Riquelme, I.; Tapia, O.; Leal, P.; Sandoval, A.; Varga, M.; Letelier, P.; Buchegger, K.; Bizama, C.; Espinoza, J.; Peek, R.M.; et al. miR-101-2, miR-125b-2 and miR-451a act as potential tumor suppressors in gastric cancer through regulation of the PI3K/AKT/mTOR pathway. Cell. Oncol. 2016, 39, 23–33. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Li, J.; Zhang, G.; Wang, Q.; Wu, C.; Zhang, Q.; Wang, H.; Sun, P.; Xiang, R.; Yang, S. Exosomal miR-451a Functions as a Tumor Suppressor in Hepatocellular Carcinoma by Targeting LPIN1. Cell. Physiol. Biochem. 2019, 53, 19–35. [Google Scholar] [CrossRef] [Green Version]
- Foss, K.; Sima, C.; Ugolini, D.; Neri, M.; Allen, K.E.; Weiss, G.J. miR-1254 and miR-574-5p: Serum-Based microRNA Biomarkers for Early-Stage Non-small Cell Lung Cancer. J. Thorac. Oncol. 2011, 6, 482–488. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Li, Z.; Zhao, M.; Ye, W.; Wu, H.; Liao, Q.; Bu, S.; Zhang, Y. Circulating miRNAs miR-574-5p and miR-3135b are potential metabolic regulators for serum lipids and blood glucose in gestational diabetes mellitus. Gynecol. Endocrinol. 2021, 37, 665–671. [Google Scholar] [CrossRef]
- Lau, P.; Bossers, K.; Janky, R.; Salta, E.; Frigerio, C.S.; Barbash, S.; Rothman, R.; Sierksma, A.S.R.; Thathiah, A.; Greenberg, D.; et al. Alteration of the micro RNA network during the progression of Alzheimer’s disease. EMBO Mol. Med. 2013, 5, 1613–1634. [Google Scholar] [CrossRef] [Green Version]
- Long, G.; Wang, F.; Li, H.; Yin, Z.; Sandip, C.; Lou, Y.; Wang, Y.; Chen, C.; Wang, D.W. Circulating miR-30a, miR-126 and let-7b as biomarker for ischemic stroke in humans. BMC Neurol. 2013, 13, 178. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Chen, L.; Chen, B.; Huang, S.; Zeng, C.; Wu, H.; Chen, C.; Long, F. Increased serum exosomal miR-134 expression in the acute ischemic stroke patients. BMC Neurol. 2018, 18, 198. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, D.-B.; Li, R.-Y.; Zhou, X.; Yu, D.-J.; Lan, X.-Y.; Li, J.-P.; Liu, J.-L. Diagnosis of Hyperacute and Acute Ischaemic Stroke: The Potential Utility of Exosomal MicroRNA-21-5p and MicroRNA-30a-5p. Cerebrovasc. Dis. 2018, 45, 204–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Lv, J.; Chen, D.; Hu, X. Serum miR-599 Serves as a Biomarker for Ischemic Stroke Patients. Clin. Lab. 2019, 65. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Fan, C.L.; Ma, L.J.; Liu, T.; Wang, C.; Song, J.-X.; Lv, Q.-S.; Pan, H.; Zhang, C.; Wang, J.-J. Distinctive expression signatures of serum microRNAs in ischaemic stroke and transient ischaemic attack patients. Thromb. Haemost. 2017, 117, 992–1001. [Google Scholar] [CrossRef]
- Tiedt, S.; Prestel, M.; Malik, R.; Schieferdecker, N.; Duering, M.; Kautzky, V.; Stoycheva, I.; Böck, J.; Northoff, B.; Klein, M.; et al. RNA-Seq Identifies Circulating miR-125a-5p, miR-125b-5p, and miR-143-3p as Potential Biomarkers for Acute Ischemic Stroke. Circ. Res. 2017, 121, 970–980. [Google Scholar] [CrossRef]
- He, X.-W.; Shi, Y.-H.; Liu, Y.-S.; Li, G.-F.; Zhao, R.; Hu, Y.; Lin, C.-C.; Zhuang, M.-T.; Su, J.-J.; Liu, J.-R. Increased plasma levels of miR-124-3p, miR-125b-5p and miR-192-5p are associated with outcomes in acute ischaemic stroke patients receiving thrombolysis. Atherosclerosis 2019, 289, 36–43. [Google Scholar] [CrossRef]
- Ikram, M.A.; Brusselle, G.; Ghanbari, M.; Goedegebure, A.; Ikram, M.K.; Kavousi, M.; Kieboom, B.C.T.; Klaver, C.C.W.; De Knegt, R.J.; Luik, A.I.; et al. Objectives, design and main findings until 2020 from the Rotterdam Study. Eur. J. Epidemiol. 2020, 35, 483–517. [Google Scholar] [CrossRef]
- Mens, M.M.; Heshmatollah, A.; Fani, L.; Ikram, M.A.; Ikram, M.K.; Ghanbari, M. Circulatory MicroRNAs as Potential Biomarkers for Stroke Risk: The Rotterdam Study. Stroke 2021, 52, 945–953. [Google Scholar] [CrossRef]
- Van Rooij, E.; Olson, E.N. MicroRNA therapeutics for cardiovascular disease: Opportunities and obstacles. Nat. Rev. Drug Discov. 2012, 11, 860–872. [Google Scholar] [CrossRef]
- Koturbash, I.; Tolleson, W.H.; Guo, L.; Yu, D.; Chen, S.; Hong, H.; Mattes, W.; Ning, B. microRNAs as pharmacogenomic biomarkers for drug efficacy and drug safety assessment. Biomark. Med. 2015, 9, 1153–1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, S.; Nanhwan, M.; Qian, J.; Kodakandla, M.; Castillo, A.C.; Thomas, B.; Liu, H.; Ye, Y. Modulation of microRNAs in hypertension-induced arterial remodeling through the β1 and β3-adrenoreceptor pathways. J. Mol. Cell. Cardiol. 2013, 65, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Yaribeygi, H.; Katsiki, N.; Behnam, B.; Iranpanah, H.; Sahebkar, A. MicroRNAs and type 2 diabetes mellitus: Molecular mechanisms and the effect of antidiabetic drug treatment. Metabolism 2018, 87, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Zambrano, T.; Hirata, R.D.; Hirata, M.H.; Cerda, A.; Salazar, L.A. Statins differentially modulate microRNAs expression in peripheral cells of hyperlipidemic subjects: A pilot study. Eur. J. Pharm. Sci. 2018, 117, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Willeit, P.; Zampetaki, A.; Dudek, K.; Kaudewitz, D.; King, A.; Kirkby, N.S.; Crosby-Nwaobi, R.; Prokopi, M.; Drozdov, I.; Langley, S.R.; et al. Circulating MicroRNAs as Novel Biomarkers for Platelet Activation. Circ. Res. 2013, 112, 595–600. [Google Scholar] [CrossRef] [Green Version]
- Broderick, J.P.; Adeoye, O.; Elm, J. Evolution of the Modified Rankin Scale and Its Use in Future Stroke Trials. Stroke 2017, 48, 2007–2012. [Google Scholar] [CrossRef]
- Banks, J.L.; Marotta, C.A. Outcomes Validity and Reliability of the Modified Rankin Scale: Implications for Stroke Clinical Trials: A literature review and synthesis. Stroke 2007, 38, 1091–1096. [Google Scholar] [CrossRef] [Green Version]
- Weisscher, N.; Vermeulen, M.; Roos, Y.B.; de Haan, R.J. What should be defined as good outcome in stroke trials; a modified Rankin score of 0–1 or 0–2? J. Neurol. 2008, 255, 867–874. [Google Scholar] [CrossRef]
- Bam, M.; Yang, X.; Sen, S.; Zumbrun, E.E.; Dennis, L.; Zhang, J.; Nagarkatti, P.S.; Nagarkatti, M. Characterization of Dysregulated miRNA in Peripheral Blood Mononuclear Cells from Ischemic Stroke Patients. Mol. Neurobiol. 2018, 55, 1419–1429. [Google Scholar] [CrossRef] [PubMed]
- Ou, J.; Kou, L.; Liang, L.; Tang, C. MiR-375 attenuates injury of cerebral ischemia/reperfusion via targetting Ctgf. Biosci. Rep. 2017, 37, BSR20171242. [Google Scholar] [CrossRef] [Green Version]
- Hunsberger, J.G.; Fessler, E.B.; Wang, Z.; Elkahloun, A.G.; Chuang, D.-M. Post-insult valproic acid-regulated microRNAs: Potential targets for cerebral ischemia. Am. J. Transl. Res. 2012, 4, 316–332. [Google Scholar] [PubMed]
- Risbud, R.M.; Porter, B.E. Changes in MicroRNA Expression in the Whole Hippocampus and Hippocampal Synaptoneurosome Fraction following Pilocarpine Induced Status Epilepticus. PLoS ONE 2013, 8, e53464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worthmann, H.; Tryc, A.; Goldbecker, A.; Ma, Y.T.; Tountopoulou, A.; Hahn, A.; Dengler, R.; Lichtinghagen, R.; Weissenborn, K. The Temporal Profile of Inflammatory Markers and Mediators in Blood after Acute Ischemic Stroke Differs Depending on Stroke Outcome. Cerebrovasc. Dis. 2010, 30, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Glickman, S.W.; Phillips, S.; Anstrom, K.J.; Laskowitz, D.T.; Cairns, C.B. Discriminative Capacity of Biomarkers for Acute Stroke in the Emergency Department. J. Emerg. Med. 2011, 41, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.J.; Emsley, H.C.A.; Gavin, C.M.; Georgiou, R.F.; Vail, A.; Barberan, E.M.; Del Zoppo, G.J.; Hallenbeck, J.M.; Rothwell, N.J.; Hopkins, S.J.; et al. Peak plasma interleukin-6 and other peripheral markers of inflammation in the first week of ischaemic stroke correlate with brain infarct volume, stroke severity and long-term outcome. BMC Neurol. 2004, 4, 2. [Google Scholar] [CrossRef] [PubMed]
- Anand, N.; Stead, L.G. Neuron-Specific Enolase as a Marker for Acute Ischemic Stroke: A Systematic Review. Cerebrovasc. Dis. 2005, 20, 213–219. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Discovery | Validation | Combined | ||||
|---|---|---|---|---|---|---|
| miRNA | Log2FC 1 | FDR 2 | Log2FC 1 | FDR 2 | Log2FC 1 | FDR 2 |
| hsa-miR-451a | 2.4 | 1.28 × 10−46 | 2.3 | 1.36 × 10−35 | 2.3 | 3.78 × 10−85 |
| hsa-miR-574-5p | −2.2 | 5.17 × 10−21 | −2.0 | 5.15 × 10−23 | −2.1 | 7.25 × 10−53 |
| hsa-miR-142-3p | −2.5 | 2.69 × 10−53 | −2.1 | 1.54 × 10−42 | −2.3 | 2.34 × 10−110 |
| hsa-miR-6721-5p | −2.7 | 2.08 × 10−13 | −2.2 | 4.17 × 10−11 | −2.5 | 9.05 × 10−34 |
| hsa-miR-4446-3p | −2.1 | 1.67 × 10−13 | −2.2 | 3.09 × 10−13 | −2.0 | 1.13 × 10−26 |
| hsa-miR-485-3p | −2.9 | 1.71 × 10−17 | −2.5 | 2.82 × 10−15 | −2.4 | 1.33 × 10−30 |
| hsa-miR-676-3p | −2.5 | 4.34 × 10−08 | −2.5 | 8.88 × 10−08 | −2.5 | 3.76 × 10−21 |
| hsa-miR-379-5p | −2.7 | 4.76 × 10−26 | −2.5 | 2.01 × 10−27 | −2.5 | 1.67 × 10−50 |
| hsa-miR-149-5p | −3.6 | 3.57 × 10−04 | −2.9 | 7.29 × 10−04 | −3.3 | 4.24 × 10−16 |
| hsa-miR-411-5p | −3.1 | 8.11 × 10−18 | −3.0 | 8.78 × 10−17 | −3.0 | 1.11 × 10−40 |
| miRNA | Target-1 | Score | Target-2 | Score | Target-3 | Score |
|---|---|---|---|---|---|---|
| hsa-miR-451a | OSR1 | 92 | CUX2 | 90 | PSMB8 | 90 |
| hsa-miR-574-5p | CALCOCO1 | 100 | FOXI2 | 100 | C11ORF96 | 97 |
| hsa-miR-142-3p | ZEB2 | 100 | TASOR2 | 100 | RICTOR | 99 |
| hsa-miR-6721-5p | NECTIN1 | 100 | KIF21B | 100 | SPRN | 100 |
| hsa-miR-4446-3p | DR1 | 98 | CBX7 | 96 | MBNL2 | 96 |
| hsa-miR-485-3p | CREBRF | 100 | ELAVL2 | 99 | PDZD2 | 99 |
| hsa-miR-676-3p | PTPRB | 98 | SMURF2 | 97 | ANP32B | 95 |
| hsa-miR-379-5p | TXLNG | 98 | MTMR2 | 97 | EIF4G2 | 94 |
| hsa-miR-149-5p | CACHD1 | 98 | ELP5 | 98 | VPS53 | 98 |
| hsa-miR-411-5p | MITD1 | 97 | ELFN1 | 97 | RNF149 | 97 |
| Discovery | Validation | Combined | ||||
|---|---|---|---|---|---|---|
| miRNA | Log2FC 1 | FDR 2 | Log2FC 1 | FDR 2 | Log2FC 1 | FDR 2 |
| hsa-miR-451a | 2.2 | 1.09 × 10−19 | 2.6 | 4.46 × 10−22 | 2.3 | 2.03 × 10−45 |
| hsa-miR-224-5p | −2.5 | 2.68 × 10−17 | −2.1 | 1.17 × 10−08 | −2.3 | 2.04 × 10−27 |
| hsa-miR-6721-5p | −2.8 | 2.07 × 10−08 | −2.2 | 5.38 × 10−05 | −2.4 | 1.98 × 10−15 |
| hsa-miR-142-3p | −2.3 | 2.85 × 10−34 | −2.2 | 2.16 × 10−29 | −2.3 | 6.91 × 10−77 |
| hsa-miR-411-5p | −2.9 | 2.13 × 10−11 | −2.5 | 7.66 × 10−07 | −2.8 | 4.52 × 10−25 |
| hsa-miR-379-5p | −2.8 | 4.15 × 10−22 | −2.6 | 1.43 × 10−13 | −2.7 | 1.17 × 10−44 |
| hsa-miR-149-5p | −3.4 | 5.20 × 10−04 | −3.0 | 2.00 × 10−03 | −3.3 | 1.06 × 10−11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aldous, E.K.; Toor, S.M.; Parray, A.; Al-Sarraj, Y.; Diboun, I.; Abdelalim, E.M.; Arredouani, A.; El-Agnaf, O.; Thornalley, P.J.; Akhtar, N.; et al. Identification of Novel Circulating miRNAs in Patients with Acute Ischemic Stroke. Int. J. Mol. Sci. 2022, 23, 3387. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23063387
Aldous EK, Toor SM, Parray A, Al-Sarraj Y, Diboun I, Abdelalim EM, Arredouani A, El-Agnaf O, Thornalley PJ, Akhtar N, et al. Identification of Novel Circulating miRNAs in Patients with Acute Ischemic Stroke. International Journal of Molecular Sciences. 2022; 23(6):3387. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23063387
Chicago/Turabian StyleAldous, Eman K., Salman M. Toor, Aijaz Parray, Yasser Al-Sarraj, Ilhame Diboun, Essam M. Abdelalim, Abdelilah Arredouani, Omar El-Agnaf, Paul J. Thornalley, Naveed Akhtar, and et al. 2022. "Identification of Novel Circulating miRNAs in Patients with Acute Ischemic Stroke" International Journal of Molecular Sciences 23, no. 6: 3387. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23063387