
Disulfiram Exerts Antifibrotic and Anti-Inflammatory Therapeutic Effects on Perimysial Orbital Fibroblasts in Graves’ Orbitopathy

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
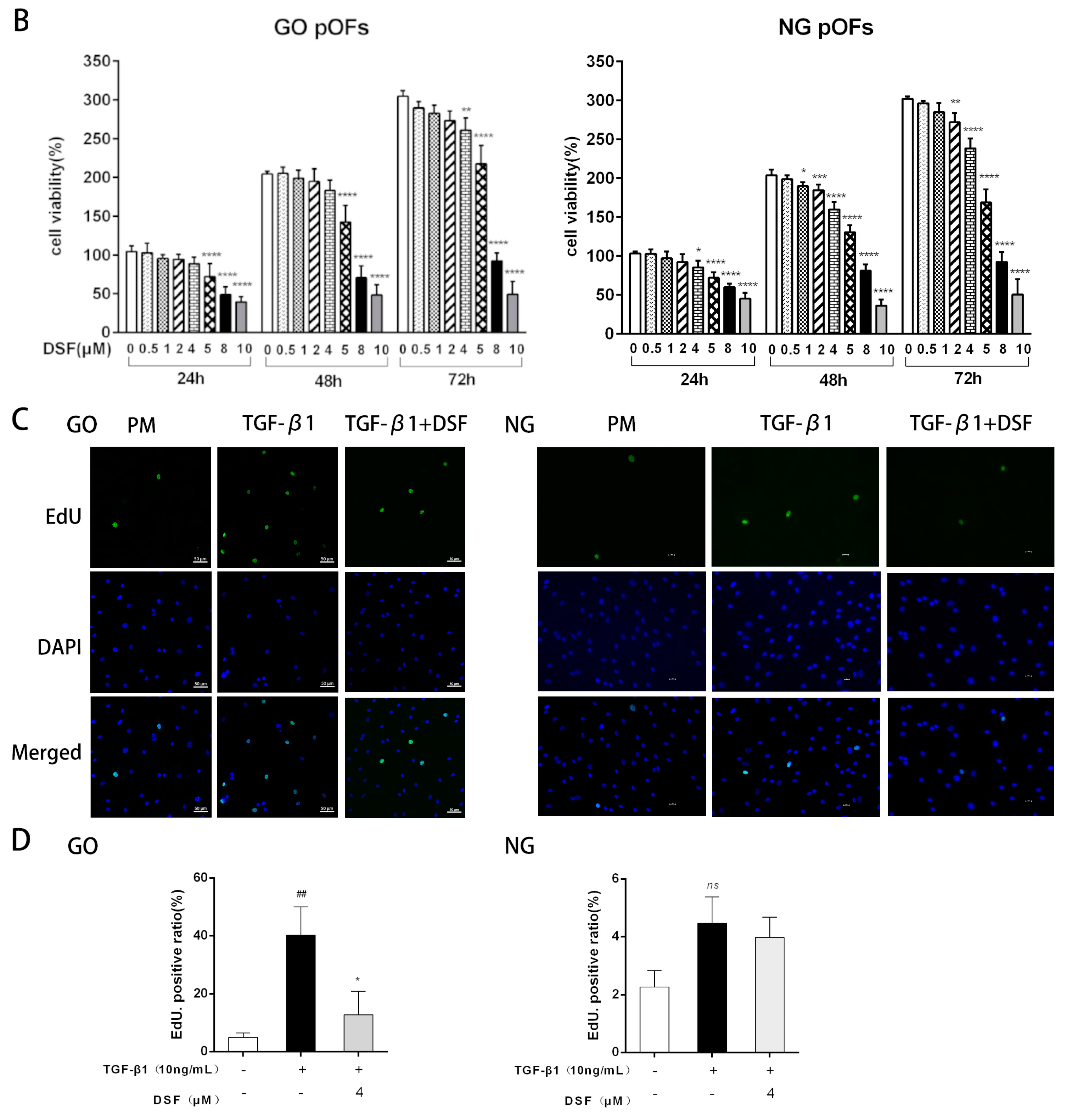
2.1. DSF Inhibits TGF-β1-Induced Proliferation of GO pOFs
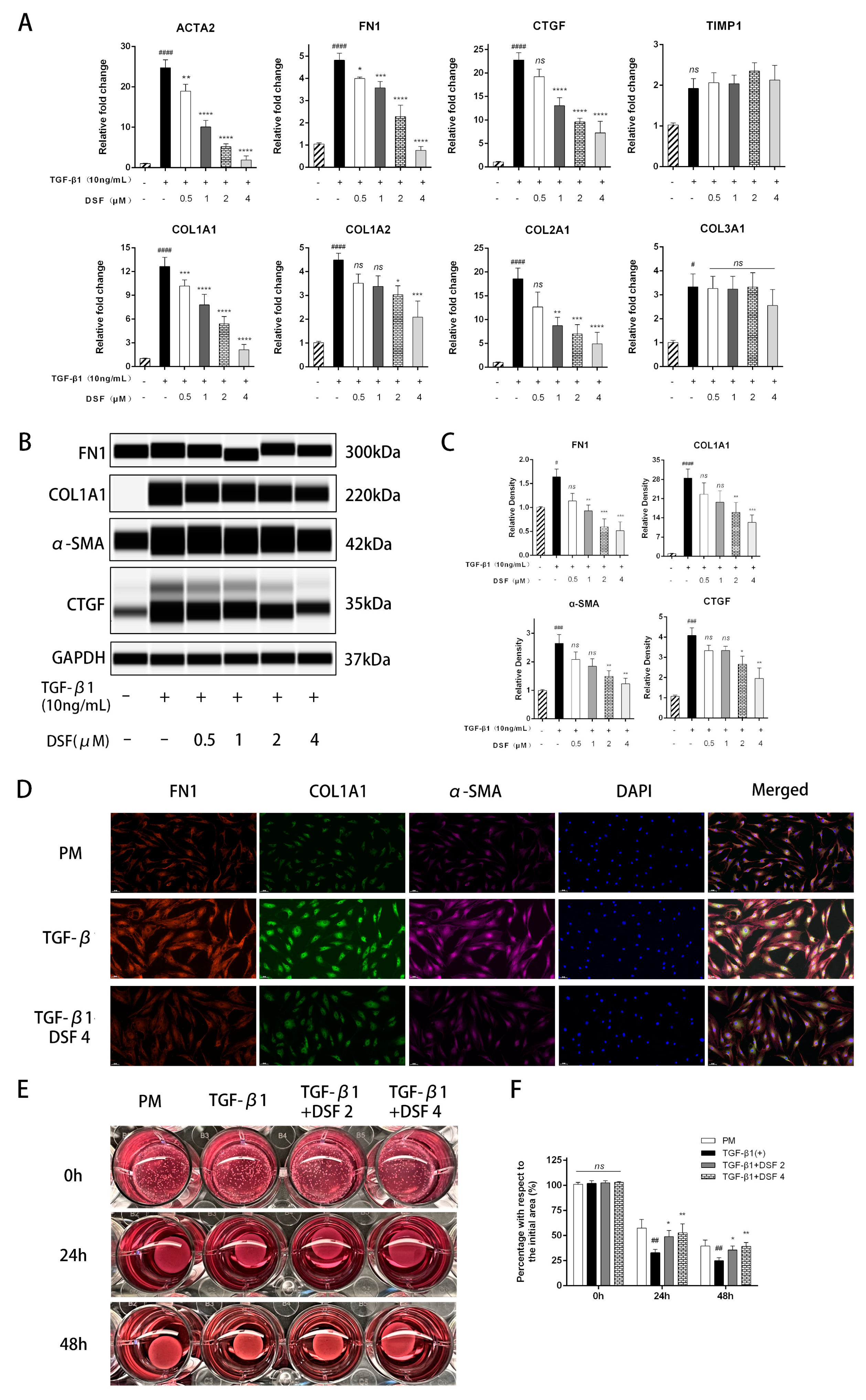
2.2. DSF Exerts Antifibrotic Effects on pOFs in the GO Group
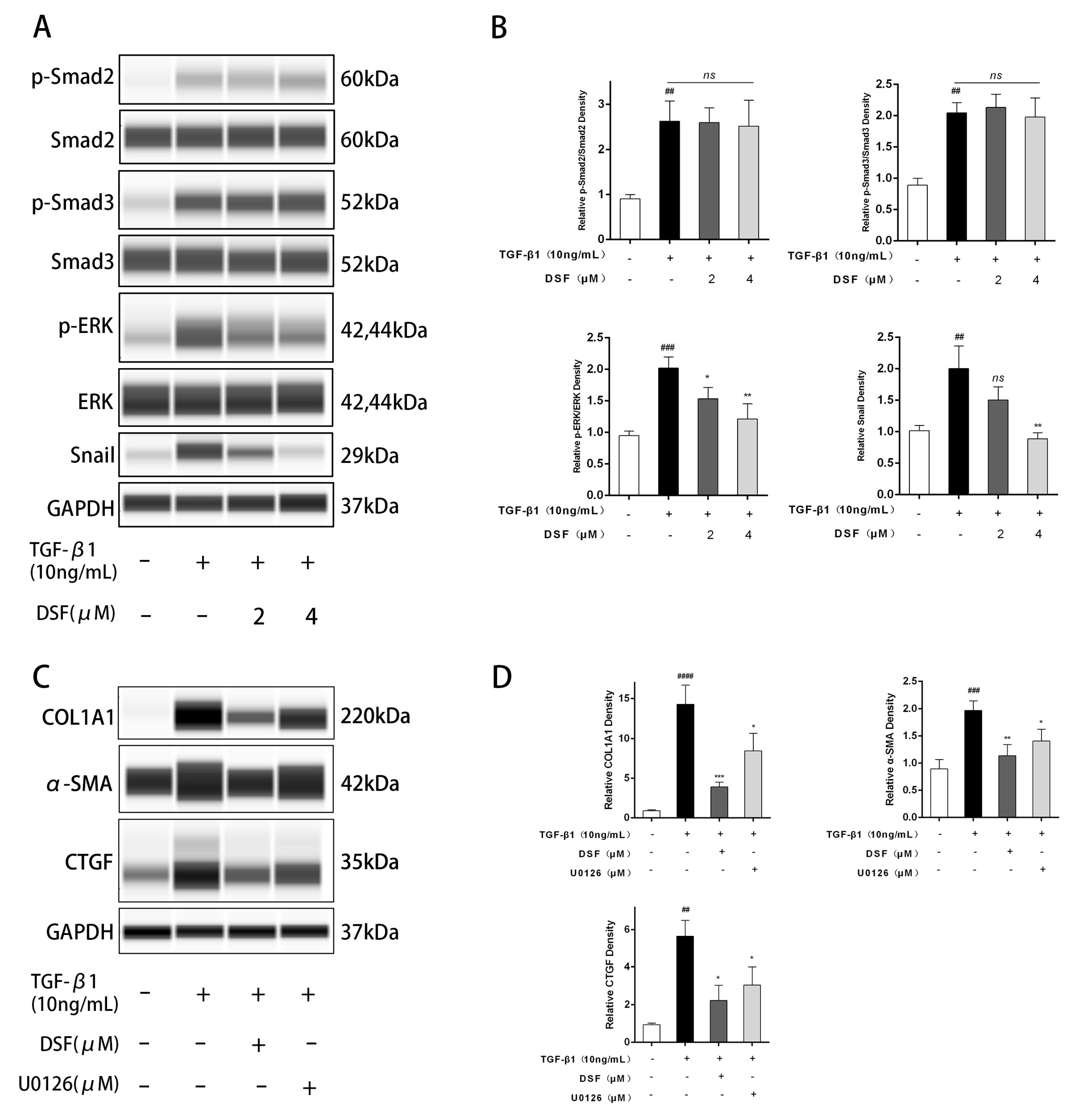
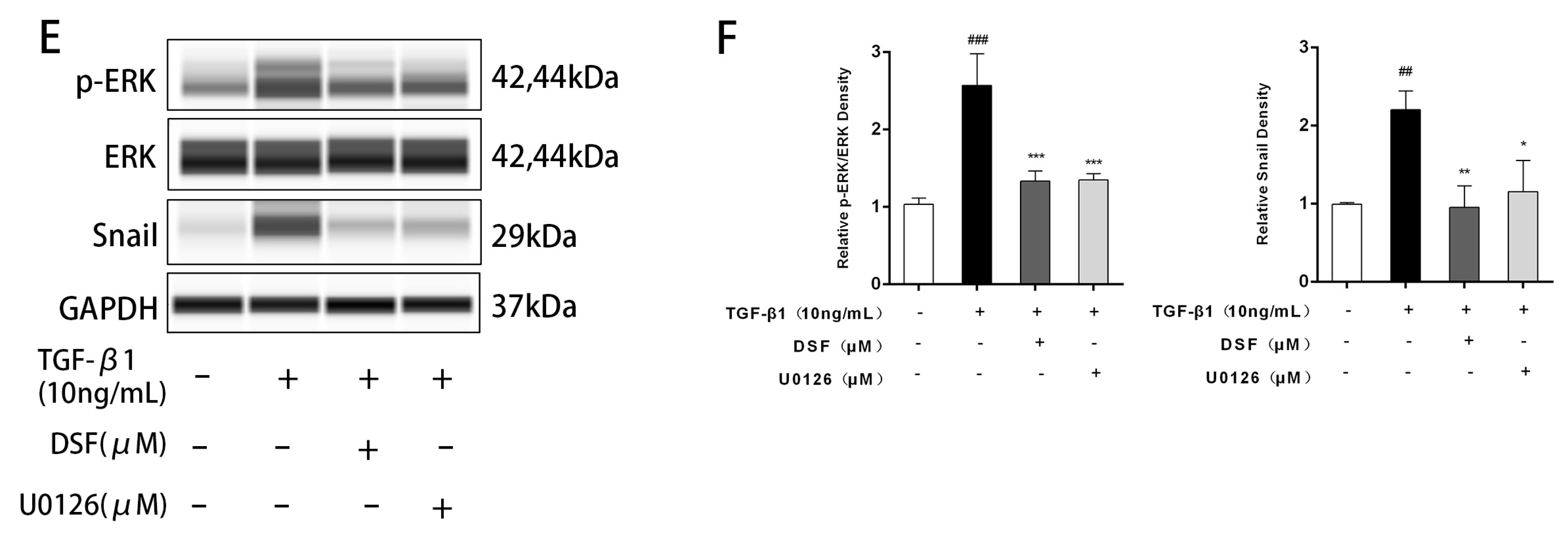
2.3. DSF Regulates the ERK-Snail Pathway during Fibrosis
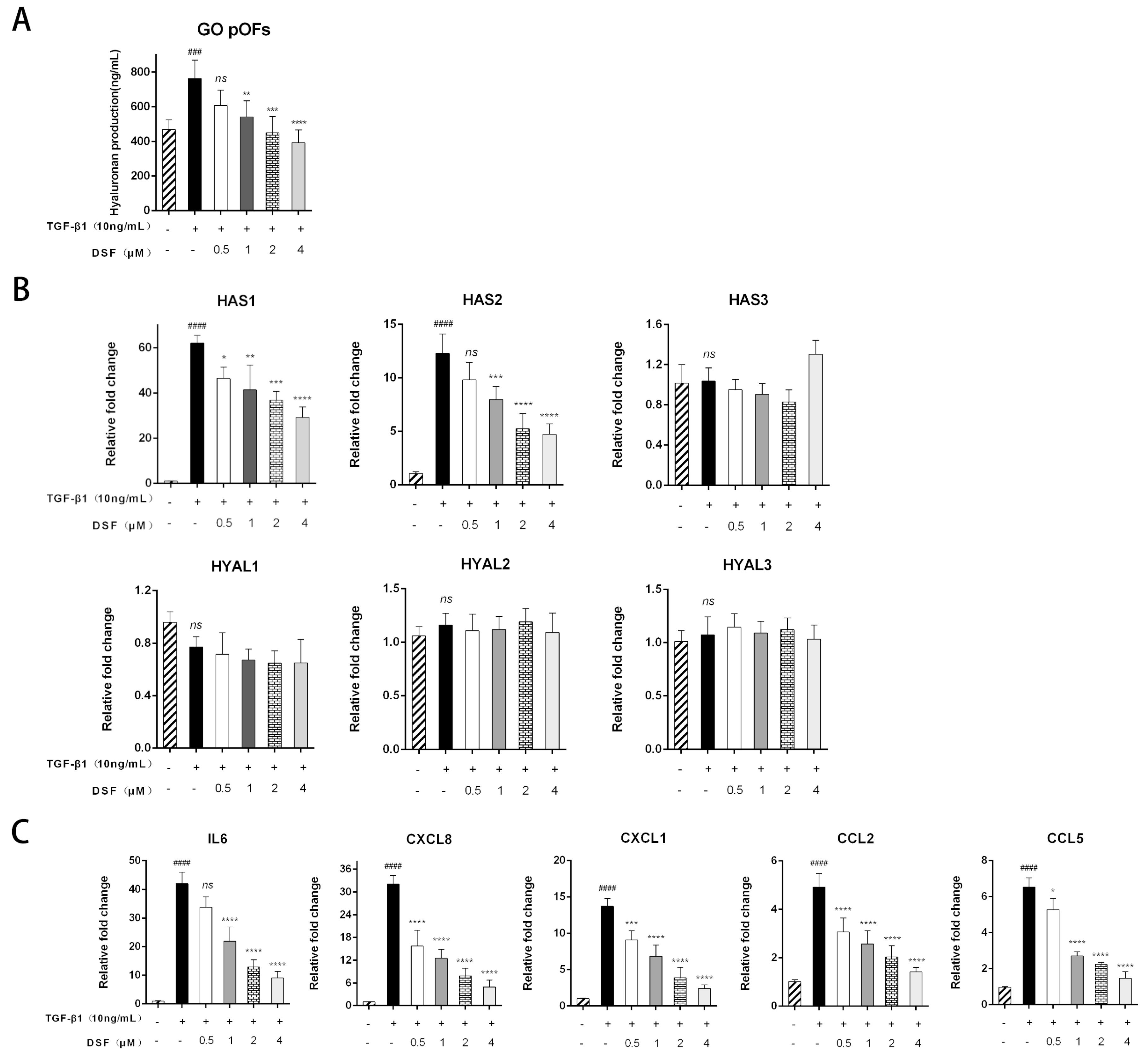
2.4. DSF Attenuates HA Production and Suppresses Inflammatory and Chemokine Expression
3. Discussion
Study Limitations
4. Materials and Methods
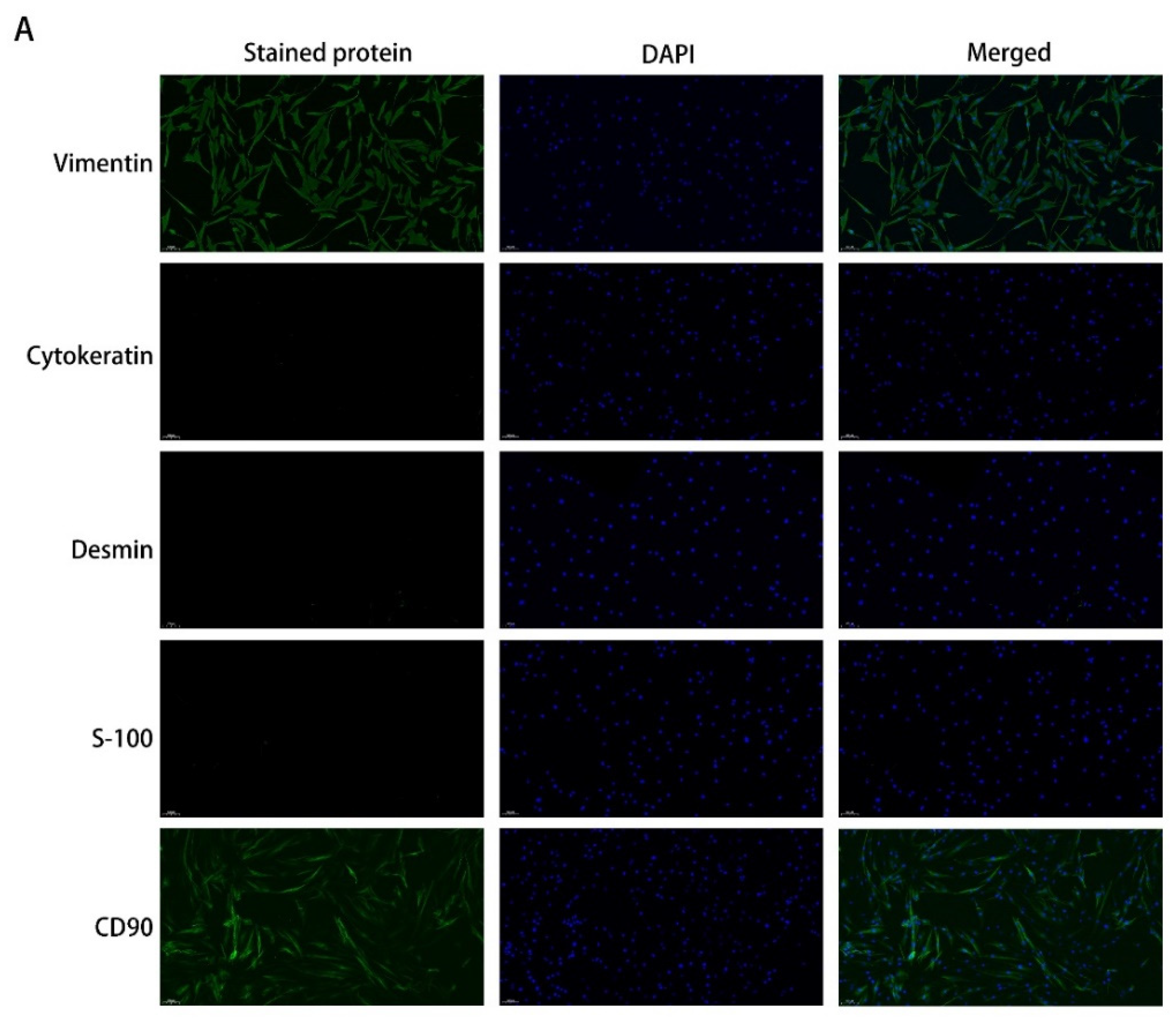
4.1. Primary Cultures of pOFs
4.2. Immunofluorescence (IF) Analysis
4.3. Cell Viability Assays
4.4. EdU
4.5. Collagen Gel Contraction Assay
4.6. Wound-Healing Assay
4.7. RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction (qPCR)
4.8. Automated Western Blotting
4.9. Hyaluronan (HA) Measurement
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Wakelkamp, I.M.; Baldeschi, L.; Saeed, P.; Mourits, M.; Prummel, M.F.; Wiersinga, W.M. Surgical or medical decompression as a first-line treatment of optic neuropathy in Graves’ ophthalmopathy? A randomized controlled trial. Clin. Endocrinol. 2005, 63, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.J.; Koumas, L.; Gagnon, A.; Bell, A.; Sempowski, G.D.; Phipps, R.P.; Sorisky, A. Orbital fibroblast heterogeneity may determine the clinical presentation of thyroid-associated ophthalmopathy. J. Clin. Endocrinol. Metab. 2002, 87, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Port, M.; Landi, S.; Gemignani, F.; Cipollini, M.; Elisei, R.; Goudeva, L.; Müller, J.A.; Nerlich, K.; Pellegrini, G.; et al. Evidence for enhanced Thy-1 (CD90) expression in orbital fibroblasts of patients with Graves’ ophthalmopathy. Thyroid 2008, 18, 1291–1296. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Fernandez, F.; Bevilacqua, T.; Lee, K.I.; Chandrashekar, R.; Hsu, L.; Garlipp, M.A.; Griswold, J.B.; Crouch, R.K.; Ghosh, D. Imatinib mesylate and AMN107 inhibit PDGF-signaling in orbital fibroblasts: A potential treatment for Graves’ ophthalmopathy. Investig. Ophthalmol. Vis. Sci. 2009, 50, 3091–3098. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.C.; Wu, S.B.; Kao, S.C.; Kau, H.C.; Lee, F.L.; Wei, Y.H. The protective effect of antioxidants on orbital fibroblasts from patients with Graves’ ophthalmopathy in response to oxidative stress. Mol. Vis. 2013, 19, 927–934. [Google Scholar]
- Bell, R.G.; Smith, H.W. Preliminary report on clinical trials of antabuse. Can. Med. Assoc. J. 1949, 60, 286–288. [Google Scholar]
- Liu, T.; Wang, P.; Cong, M.; Zhao, X.; Zhang, D.; Xu, H.; Liu, L.; Jia, J.; You, H. Diethyldithiocarbamate, an anti-abuse drug, alleviates steatohepatitis and fibrosis in rodents through modulating lipid metabolism and oxidative stress. Br. J. Pharmacol. 2018, 175, 4480–4495. [Google Scholar] [CrossRef]
- Li, Y.; Chen, F.; Chen, J.; Chan, S.; He, Y.; Liu, W.; Zhang, G. Disulfiram/copper induces antitumor activity against both nasopharyngeal cancer cells and cancer-associated fibroblasts through ROS/MAPK and ferroptosis pathways. Cancers 2020, 12, 138. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, R.; Han, X. Disulfiram inhibits inflammation and fibrosis in a rat unilateral ureteral obstruction model by inhibiting gasdermin D cleavage and pyroptosis. Inflamm. Res. 2021, 70, 543–552. [Google Scholar] [CrossRef]
- Fillmore, N.; Bell, S.; Shen, C.; Nguyen, V.; La, J.; Dubreuil, M.; Strymish, J.; Brophy, M.; Mehta, G.; Wu, H.; et al. Disulfiram use is associated with lower risk of COVID-19: A retrospective cohort study. PLoS ONE 2021, 16, e0259061. [Google Scholar] [CrossRef]
- Adrover, J.M.; Carrau, L.; Daßler-Plenker, J.; Bram, Y.; Chandar, V.; Houghton, S.; Redmond, D.; Merrill, J.R.; Shevik, M. Disulfiram inhibits neutrophil extracellular trap formation protecting rodents from acute lung injury and SARS-CoV-2 infection. JCI Insight 2022, 7, e157342. [Google Scholar] [CrossRef] [PubMed]
- Ahadome, S.D.; Abraham, D.J.; Rayapureddi, S.; Saw, V.P.; Saban, D.R.; Calder, V.L.; Norman, J.T.; Ponticos, M.; Daniels, J.T.; Dart, J.K. Aldehyde dehydrogenase inhibition blocks mucosal fibrosis in human and mouse ocular scarring. JCI Insight 2016, 1, e87001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Li, H.; Chen, X.; Yang, H.; Guan, H.; He, X.; Chen, Y.; Pokharel, S.; Xiao, H.; Li, Y. Novel roles of chloroquine and hydroxychloroquine in graves’ orbitopathy therapy by targeting orbital fibroblasts. J. Clin. Endocrinol. Metab. 2020, 105, 1906–1917. [Google Scholar] [CrossRef] [Green Version]
- El Agha, E.; Kramann, R.; Schneider, R.K.; Li, X.; Seeger, W.; Humphreys, B.D.; Bellusci, S. Mesenchymal stem cells in fibrotic disease. Cell Stem Cell 2017, 21, 166–177. [Google Scholar] [CrossRef] [PubMed]
- Koumas, L.; Smith, T.J.; Feldon, S.; Blumberg, N.; Phipps, R.P. Thy-1 expression in human fibroblast subsets defines myofibroblastic or lipofibroblastic phenotypes. Am. J. Pathol. 2003, 163, 1291–1300. [Google Scholar] [CrossRef] [Green Version]
- Force, T.; Bonventre, J.V. Growth factors and mitogen-activated protein kinases. Hypertension 1998, 31, 152–161. [Google Scholar] [CrossRef]
- Liu, K.; Song, Y.; Xu, G.; Ye, J.; Wu, Z.; Liu, X.; Dong, X.; Zhang, M.; Xing, Y.; Zhu, S.; et al. Conbercept for treatment of neovascular age-related macular degeneration: Results of the randomized phase 3 PHOENIX study. Am. J. Ophthalmol. 2019, 197, 156–167. [Google Scholar] [CrossRef]
- Smith, T.; Bahn, R.; Gorman, C. Connective tissue, glycosaminoglycans, and diseases of the thyroid. Endocr. Rev. 1989, 10, 366–391. [Google Scholar] [CrossRef]
- Smith, T.J.; Kahaly, G.J.; Ezra, D.G.; Fleming, J.C.; Dailey, R.A.; Tang, R.A.; Harris, G.J.; Antonelli, A.; Salvi, M.; Goldberg, R.A.; et al. Teprotumumab for thyroid-associated ophthalmopathy. N. Engl. J. Med. 2017, 376, 1748–1761. [Google Scholar] [CrossRef]
- Douglas, R.S.; Kahaly, G.J.; Patel, A.; Sile, S.; Thompson, E.H.; Perdok, R.; Fleming, J.C.; Fowler, B.T.; Marcocci, C.; Marinò, M.; et al. Teprotumumab for the treatment of active thyroid eye disease. N. Engl. J. Med. 2020, 382, 341–352. [Google Scholar] [CrossRef]
- Vannucchi, G.; Campi, I.; Covelli, D.; Currò, N.; Lazzaroni, E.; Palomba, A.; Soranna, D.; Zambon, A.; Fugazzola, L.; Muller, I.; et al. Efficacy profile and safety of very low-dose rituximab in patients with graves’ orbitopathy. Thyroid 2021, 31, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Ayabe, R.; Rootman, D.B.; Hwang, C.J.; Ben-Artzi, A.; Goldberg, R. Adalimumab as steroid-sparing treatment of inflammatory-stage thyroid eye disease. Ophthalmic Plast. Reconstr. Surg. 2014, 30, 415–419. [Google Scholar] [CrossRef] [PubMed]
- Durrani, O.M.; Reuser, T.Q.; Murray, P.I. Infliximab: A novel treatment for sight-threatening thyroid associated ophthalmopathy. Orbit 2005, 24, 117–119. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Bilbao, L.; Martínez-López, D.; Revenga, M.; López-Vázquez, Á.; Valls-Pascual, E.; Atienza-Mateo, B.; Valls-Espinosa, B.; Maiz-Alonso, O.; Blanco, A.; Torre-Salaberri, I.; et al. Anti-IL-6 receptor tocilizumab in refractory graves’ orbitopathy: National multicenter observational study of 48 patients. J. Clin. Med. 2020, 9, 2816. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Moreiras, J.V.; Varela-Agra, M.; Prada-Sánchez, M.C.; Prada-Ramallal, G. Steroid-resistant graves’ orbitopathy treated with tocilizumab in real-world clinical practice: A 9-year single-center experience. J. Clin. Med. 2021, 10, 706. [Google Scholar] [CrossRef]
- Reich, K.; Warren, R.B.; Lebwohl, M.; Gooderham, M.; Strober, B.; Langley, R.G.; Paul, C.; De Cuyper, D.; Vanvoorden, V.; Madden, C.; et al. Bimekizumab versus Secukinumab in plaque psoriasis. N. Engl. J. Med. 2021, 385, 142–152. [Google Scholar] [CrossRef]
- Virakul, S.; van Steensel, L.; Dalm, V.A.; Paridaens, D.; van Hagen, P.M.; Dik, W.A. Platelet-derived growth factor: A key factor in the pathogenesis of graves’ ophthalmopathy and potential target for treatment. Eur. Thyroid J. 2014, 3, 217–226. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.B.; Hou, T.Y.; Kau, H.C.; Tsai, C.C. Effect of pirfenidone on TGF-beta1-induced myofibroblast differentiation and extracellular matrix homeostasis of human orbital fibroblasts in graves’ ophthalmopathy. Biomolecules 2021, 11, 1424. [Google Scholar] [CrossRef]
- Yoon, J.S.; Chae, M.K.; Jang, S.Y.; Lee, S.Y.; Lee, E.J. Antifibrotic effects of quercetin in primary orbital fibroblasts and orbital fat tissue cultures of Graves’ orbitopathy. Investig. Ophthalmol. Vis. Sci. 2012, 53, 5921–5929. [Google Scholar] [CrossRef]
- Wei, Y.H.; Liao, S.L.; Wang, S.H.; Wang, C.C.; Yang, C.H. Simvastatin and ROCK inhibitor Y-27632 inhibit myofibroblast differentiation of graves’ ophthalmopathy-derived orbital fibroblasts via RhoA-mediated ERK and p38 signaling pathways. Front. Endocrinol. 2020, 11, 607968. [Google Scholar] [CrossRef]
- Yu, W.K.; Hwang, W.L.; Wang, Y.C.; Tsai, C.C.; Wei, Y.H. Curcumin suppresses TGF-beta1-induced myofibroblast differentiation and attenuates angiogenic activity of orbital fibroblasts. Int. J. Mol. Sci. 2021, 22, 6829. [Google Scholar] [CrossRef]
- Ma, R.; Ren, H.; Xu, B.; Cheng, Y.; Gan, L.; Zhang, R.; Wu, J.; Qian, J. PH20 inhibits TGFbeta1-induced differentiation of perimysial orbital fibroblasts via hyaluronan-CD44 pathway in thyroid-associated ophthalmopathy. Investig. Ophthalmol. Vis. Sci. 2019, 60, 1431–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burch, H.B.; Lahiri, S.; Bahn, R.S.; Barnes, S. Superoxide radical production stimulates retroocular fibroblast proliferation in Graves’ ophthalmopathy. Exp. Eye Res. 1997, 65, 311–316. [Google Scholar] [CrossRef] [PubMed]
- van Steensel, L.; Paridaens, D.; van Meurs, M.; van Hagen, P.M.; van den Bosch, W.A.; Kuijpers, R.W.; Drexhage, H.A.; Hooijkaas, H.; Dik, W.A. Orbit-infiltrating mast cells, monocytes, and macrophages produce PDGF isoforms that orchestrate orbital fibroblast activation in Graves’ ophthalmopathy. J. Clin. Endocrinol. Metab. 2012, 97, E400–E408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.E. Non-smad signaling pathways of the TGF-beta family. Cold Spring Harb. Perspect. Biol. 2017, 9, a022129. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Law, B.K.; Chytil, A.M.; Brown, K.A.; Aakre, M.E.; Moses, H.L. Activation of the Erk pathway is required for TGF-beta1-induced EMT in vitro. Neoplasia 2004, 6, 603–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbera, M.J.; Puig, I.; Domínguez, D.; Julien-Grille, S.; Guaita-Esteruelas, S.; Peiro, S.; Baulida, J.; Francí, C.; Dedhar, S.; Larue, L.; et al. Regulation of Snail transcription during epithelial to mesenchymal transition of tumor cells. Oncogene 2004, 23, 7345–7354. [Google Scholar] [CrossRef] [Green Version]
- Jiang, F.; Chen, Y.; Ren, S.; Li, Z.; Sun, K.; Xing, Y.; Zhu, Y.; Piao, D. Cyclovirobuxine D inhibits colorectal cancer tumorigenesis via the CTHRC1AKT/ERKSnail signaling pathway. Int. J. Oncol. 2020, 57, 183–196. [Google Scholar] [CrossRef] [Green Version]
- Saifi, M.A.; Shaikh, A.S.; Kaki, V.R.; Godugu, C. Disulfiram prevents collagen crosslinking and inhibits renal fibrosis by inhibiting lysyl oxidase enzymes. J. Cell Physiol. 2022. online ahead of print. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, J.; Zhao, J.; Chen, S.; Zhou, T.; Xu, J. Treatment of severe acute pancreatitis and related lung injury by targeting gasdermin D-mediated pyroptosis. Front. Cell Dev. Biol. 2021, 9, 780142. [Google Scholar] [CrossRef]
- Bartalena, L.; Baldeschi, L.; Dickinson, A.; Eckstein, A.; Kendall-Taylor, P.; Marcocci, C.; Mourits, M.; Perros, P.; Boboridis, K.; Boschi, A.; et al. Consensus statement of the European group on Graves’ orbitopathy (EUGOGO) on management of Graves’ orbitopathy. Thyroid 2008, 18, 333–346. [Google Scholar] [CrossRef] [PubMed]
- Bartalena, L.; Baldeschi, L.; Boboridis, K.; Eckstein, A.; Kahaly, G.J.; Marcocci, C.; Perros, P.; Salvi, M.; Wiersinga, W.M. The 2016 European Thyroid Association/European Group on Graves’ Orbitopathy Guidelines for the Management of Graves’ Orbitopathy. Eur. Thyroid J. 2016, 5, 9–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartalena, L.; Kahaly, G.J.; Baldeschi, L.; Dayan, C.M.; Eckstein, A.; Marcocci, C.; Marino, M.; Vaidya, B.; Wiersinga, W.M. The 2021 European Group on Graves’ orbitopathy (EUGOGO) clinical practice guidelines for the medical management of Graves’ orbitopathy. Eur. J. Endocrinol. 2021, 185, G43–G67. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Wu, Y.; Huang, Y.; Fang, S.; Li, Y.; Sun, J.; Zhou, H. Immunological features of paranasal sinus mucosa in patients with graves’ orbitopathy. Front. Endocrinol. 2020, 11, 621321. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age (Years) | Gender (M/F) | Smoking (Y/N) | Duration of GO (Years) | Previous GO Treatment | Proptosis (R/L, mm) | CAS | GO Severity Assessment | Surgery Performed |
|---|---|---|---|---|---|---|---|---|
| GO patients | ||||||||
| 64 | M | Y | 2.25 | GCs | 20/18 | 3/7 | VI | Decompression |
| 58 | F | N | 2.5 | GCs | 19/20 | 0/7 | VI | Decompression |
| 68 | M | N | 3 | None | 20/18 | 1/7 | VI | Decompression |
| 47 | M | Y | 1.75 | GCs | 18/19 | 2/7 | VI | Decompression |
| 43 | M | N | 1.25 | GCs | 20/21 | 0/7 | VI | Decompression |
| 50 | F | N | 1.5 | None | 22/22 | 2/7 | IV | Decompression |
| 42 | F | N | 1 | None | 19/20 | 0/7 | IV | Decompression |
| 54 | M | N | 2 | GCs | 19/19 | 2/7 | VI | Decompression |
| Non-GO control patients | ||||||||
| 45 | F | N | - | - | - | - | - | Enucleation |
| 55 | M | Y | - | - | - | - | - | Enucleation |
| 51 | F | N | - | - | - | - | - | Enucleation |
| 39 | F | N | - | - | - | - | - | Enucleation |
| 56 | F | N | - | - | - | - | - | Enucleation |
| 67 | M | N | - | - | - | - | - | Enucleation |
| Genes | Sequences (5′-3′) |
|---|---|
| ACTA2 | F: GAACCCTAAGGCCAACCGGGAGAAA |
| R: CCACATACATGGCGGGGACATTGA | |
| FN1 | F: ACAAGCATGTCTCTCTGCCAA |
| R: GCAATGTGCAGCCCTCATTT | |
| CTGF | F: AGCTGACCTGGAAGAGAACATT |
| R: GCTCGGTATGTCTTCATGCTG | |
| TIMP1 | F: CATCACTACCTGCAGTTTTGTG |
| R: TGGATAAACAGGGAAACACTGT | |
| COL1A1 | F: AAAGATGGACTCAACGGTCTC |
| R: CATCGTGAGCCTTCTCTTGAG | |
| COL1A2 | F: CTCCATGGTGAGTTTGGTCTC |
| R: CTTCCAATAGGACCAGTAGGAC | |
| COL2A1 | F: CCAGATGACCTTCCTACGCC |
| R: TTCAGGGCAGTGTACGTGAAC | |
| COL3A1 | F: CTTCTCTCCAGCCGAGCTTC |
| R: CCAGTGTGTTTCGTGCAACC | |
| HAS1 | F: GCGGGCTTGTCAGAGCTAC |
| R: ACTGCTGCAAGAGGTTATTCC | |
| HAS2 | F: CCTCCTGGGTGGTGTGATTT |
| R: GCGTCAAAAGCATGACCCAA | |
| HAS3 | F: TTATACAGCTTTTCTACCGGGG |
| R: CAGAAGGCTGGACATATAGAGG | |
| HYAL1 | F: TTCCCTGACTGCTACAACTATG |
| R: CATGTAGATGCTGGGATAGAGG | |
| HYAL2 | F: CATGATTATGTGCAGAACTGGG |
| R: GTCGTGTGAAGACGTAGACTG | |
| HYAL3 | F: CAGCTCTACAAGGCCTATACTG |
| R:TAGTTGGAAGCCATACTATGCC | |
| IL6 | F: CACTGGTCTTTTGGAGTTTGAG |
| R: GGACTTTTGTACTCATCTGCAC | |
| CXCL8 | F: CCACCGGAAGGAACCATCTC |
| R: GGGGTGGAAAGGTTTGGAGT | |
| CXCL1 | F: TTCACAGTGTGTGGTCAACAT |
| R: AAGCCCCTTTGTTCTAAGCCA | |
| CCL2 | F: CCTTCATTCCCCAAGGGCTC |
| R: CTTCTTTGGGACACTTGCTGC | |
| CCL5 | F: GCAAGCTTTGTCACCCGAAA |
| R: CCCAAGCTAGGACAAGAGCA | |
| GAPDH | F: TTGCCATCAATGACCCCTT |
| R: CGCCCCACTTGATTTTGGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Ye, H.; Yang, S.; Sha, X.; Wang, X.; Zhang, T.; Chen, R.; Xiao, W.; Yang, H. Disulfiram Exerts Antifibrotic and Anti-Inflammatory Therapeutic Effects on Perimysial Orbital Fibroblasts in Graves’ Orbitopathy. Int. J. Mol. Sci. 2022, 23, 5261. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095261
Wang X, Ye H, Yang S, Sha X, Wang X, Zhang T, Chen R, Xiao W, Yang H. Disulfiram Exerts Antifibrotic and Anti-Inflammatory Therapeutic Effects on Perimysial Orbital Fibroblasts in Graves’ Orbitopathy. International Journal of Molecular Sciences. 2022; 23(9):5261. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095261
Chicago/Turabian StyleWang, Xing, Huijing Ye, Shenglan Yang, Xiaotong Sha, Xiandai Wang, Te Zhang, Rongxin Chen, Wei Xiao, and Huasheng Yang. 2022. "Disulfiram Exerts Antifibrotic and Anti-Inflammatory Therapeutic Effects on Perimysial Orbital Fibroblasts in Graves’ Orbitopathy" International Journal of Molecular Sciences 23, no. 9: 5261. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095261