
Visualization of the Crossroads between a Nascent Infection Thread and the First Cell Division Event in Phaseolus vulgaris Nodulation

 ,
,  , and
, and 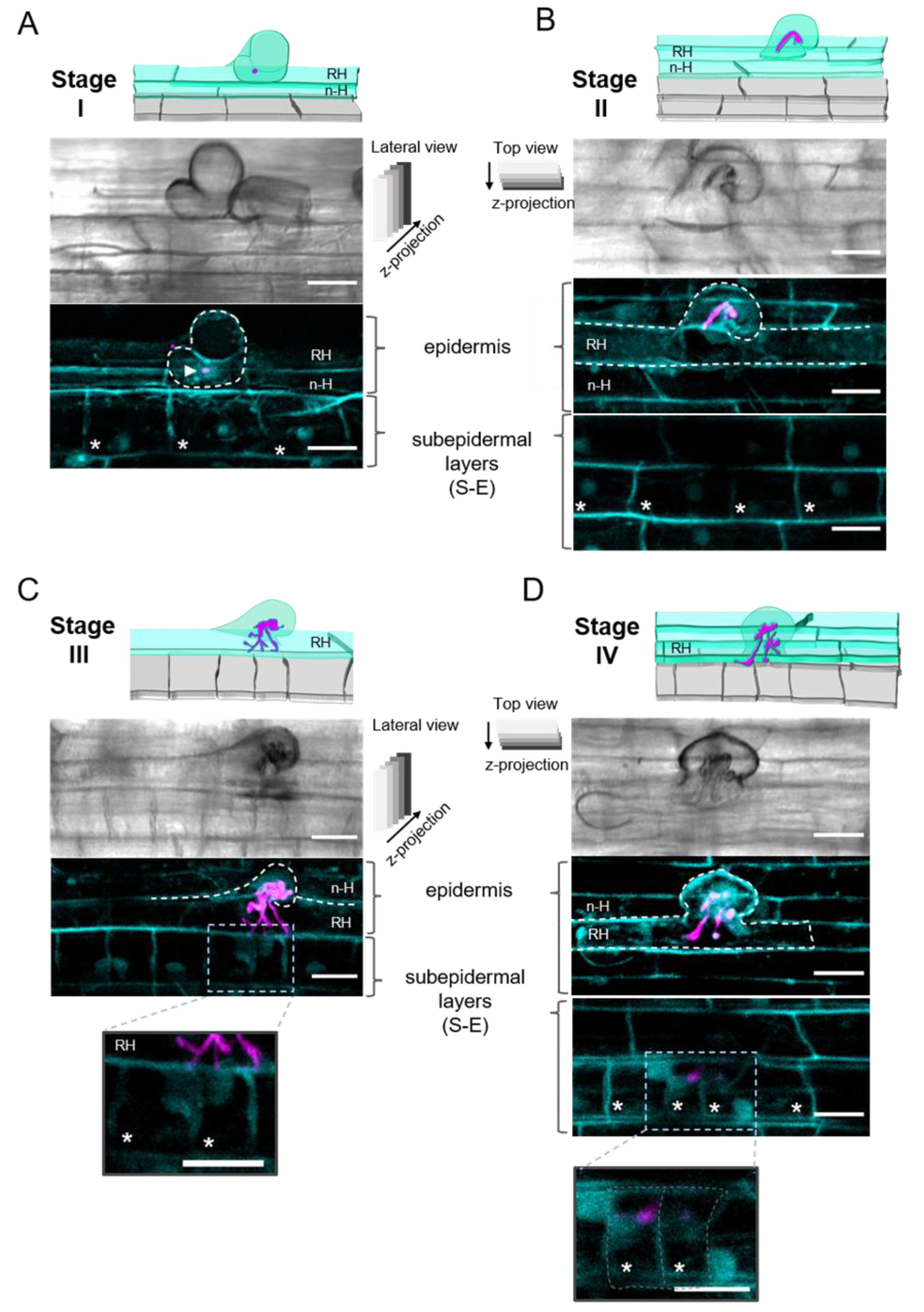{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
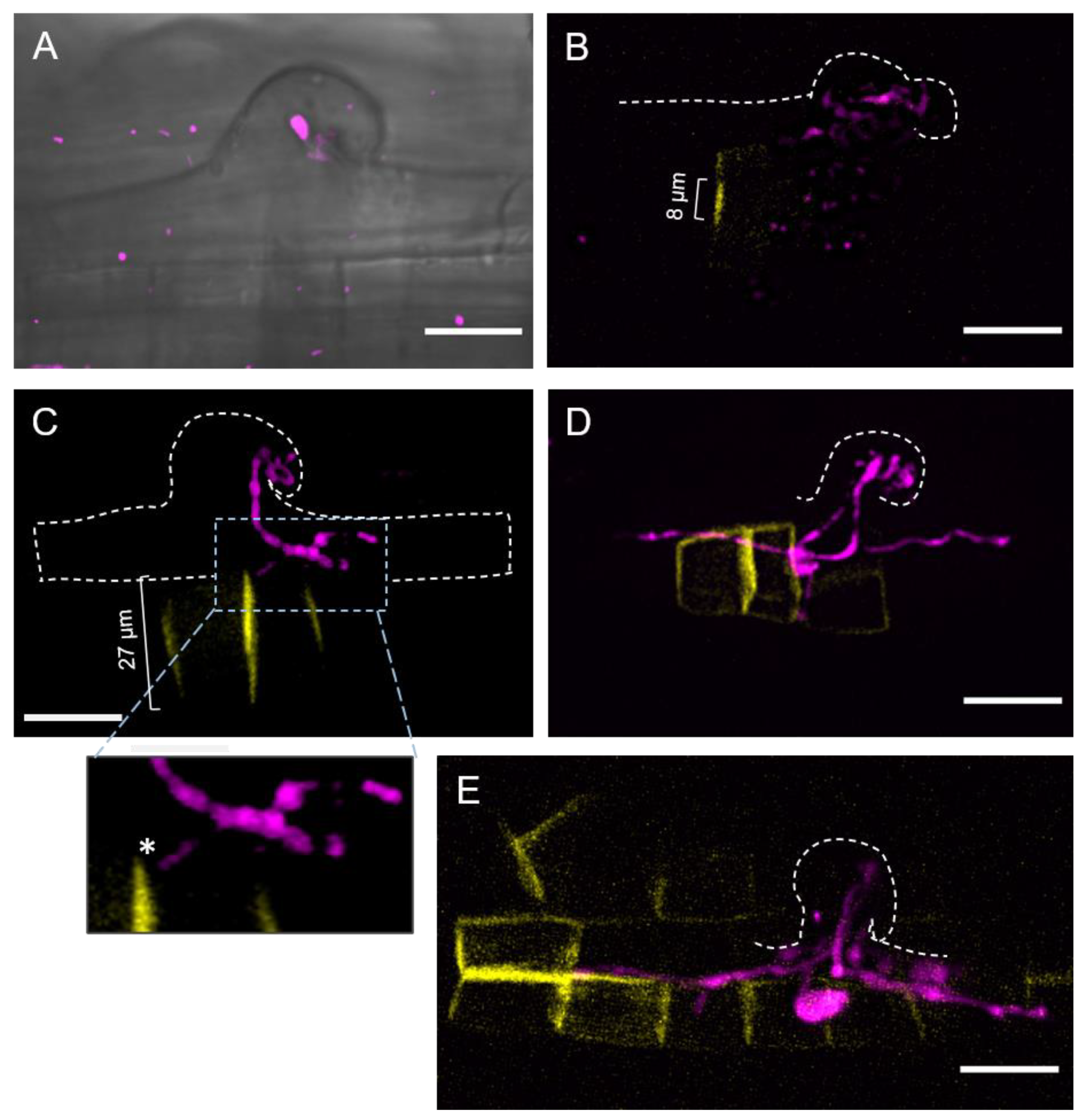
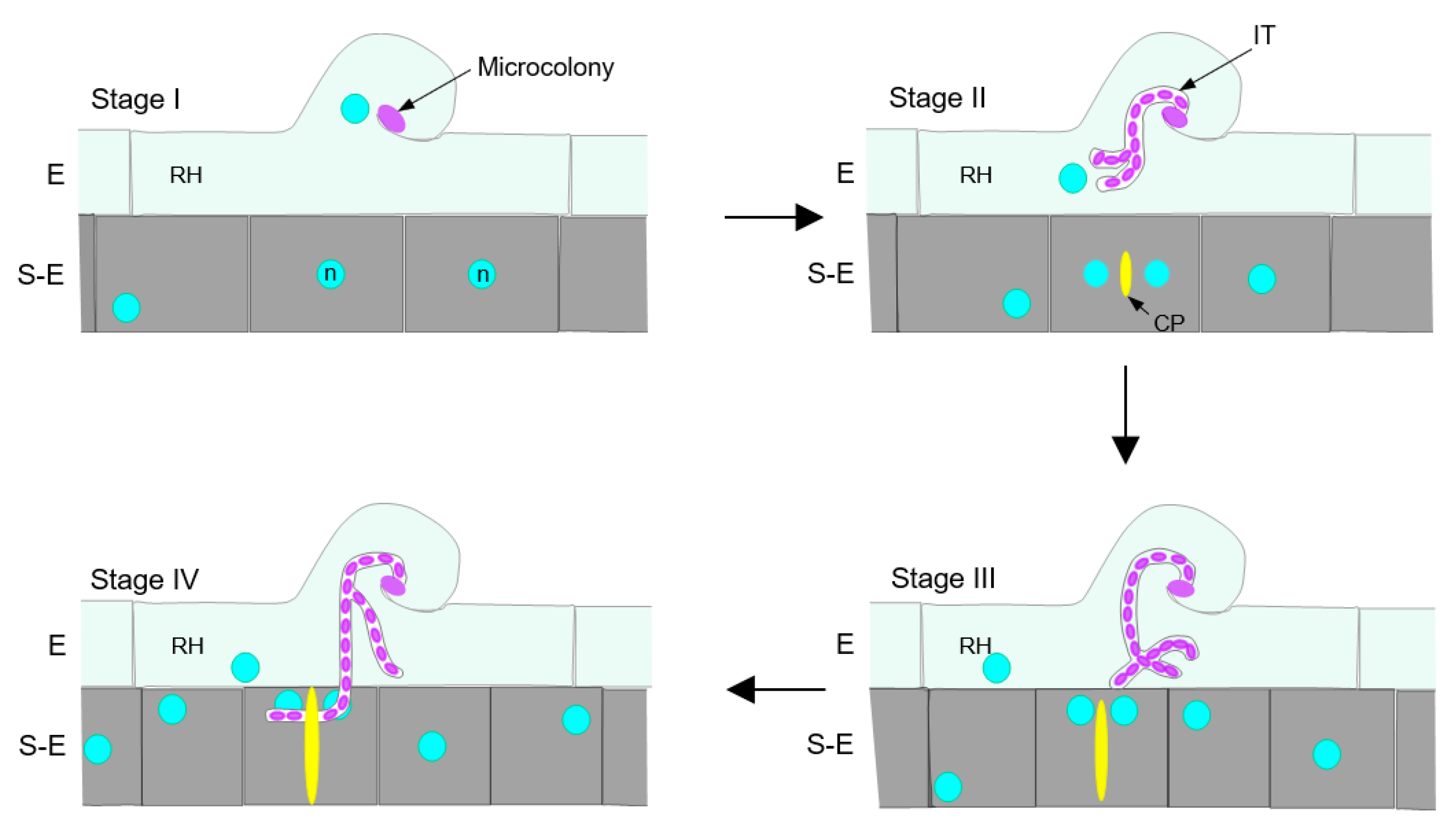
2.1. Epidermal Infection Progression Can Be Described in Four Stages (I–IV)
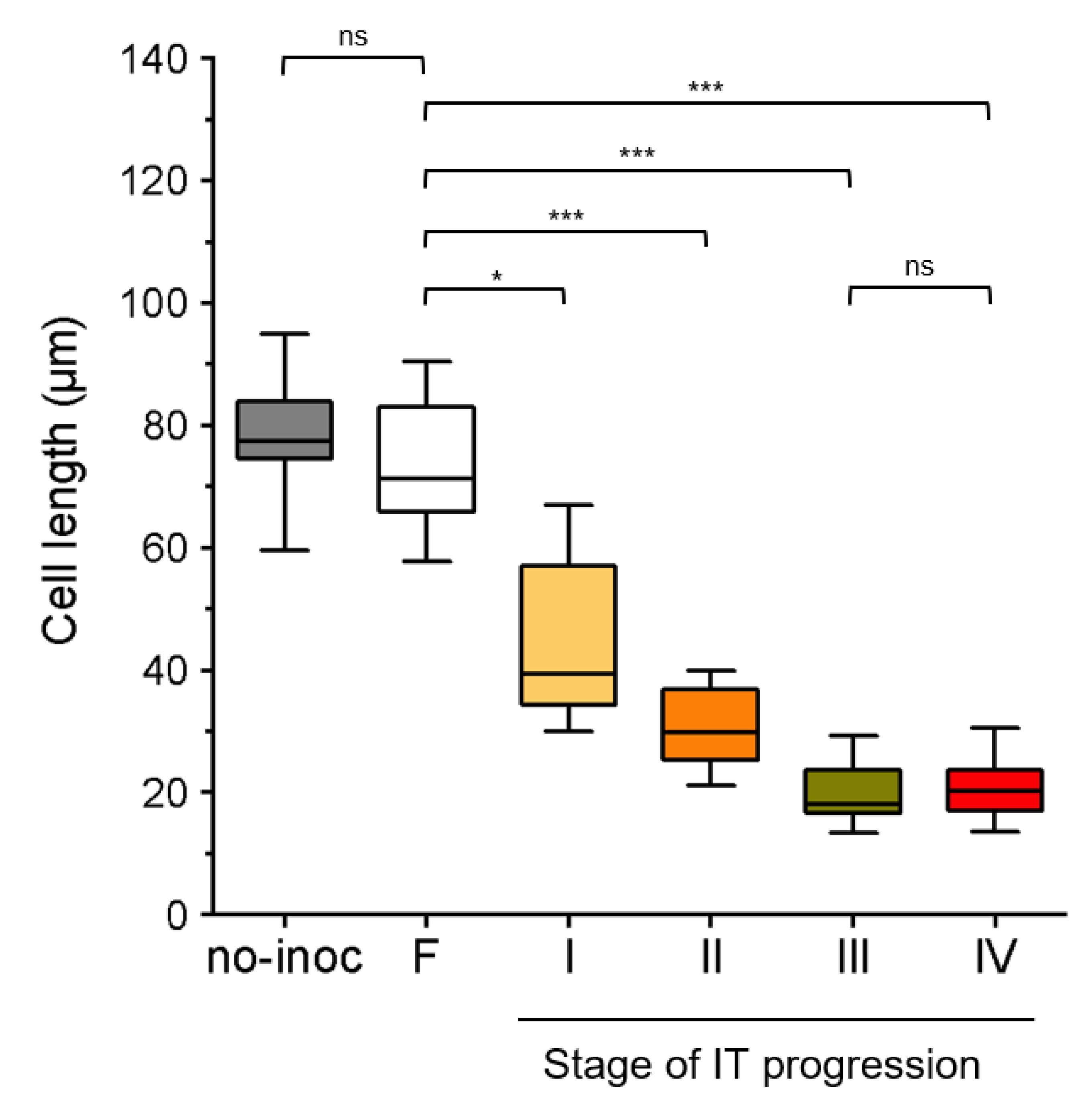
2.2. Subepidermal Cells (S-E) Underneath the Infection Site Become Shorter as the Infection Thread (IT) Progresses
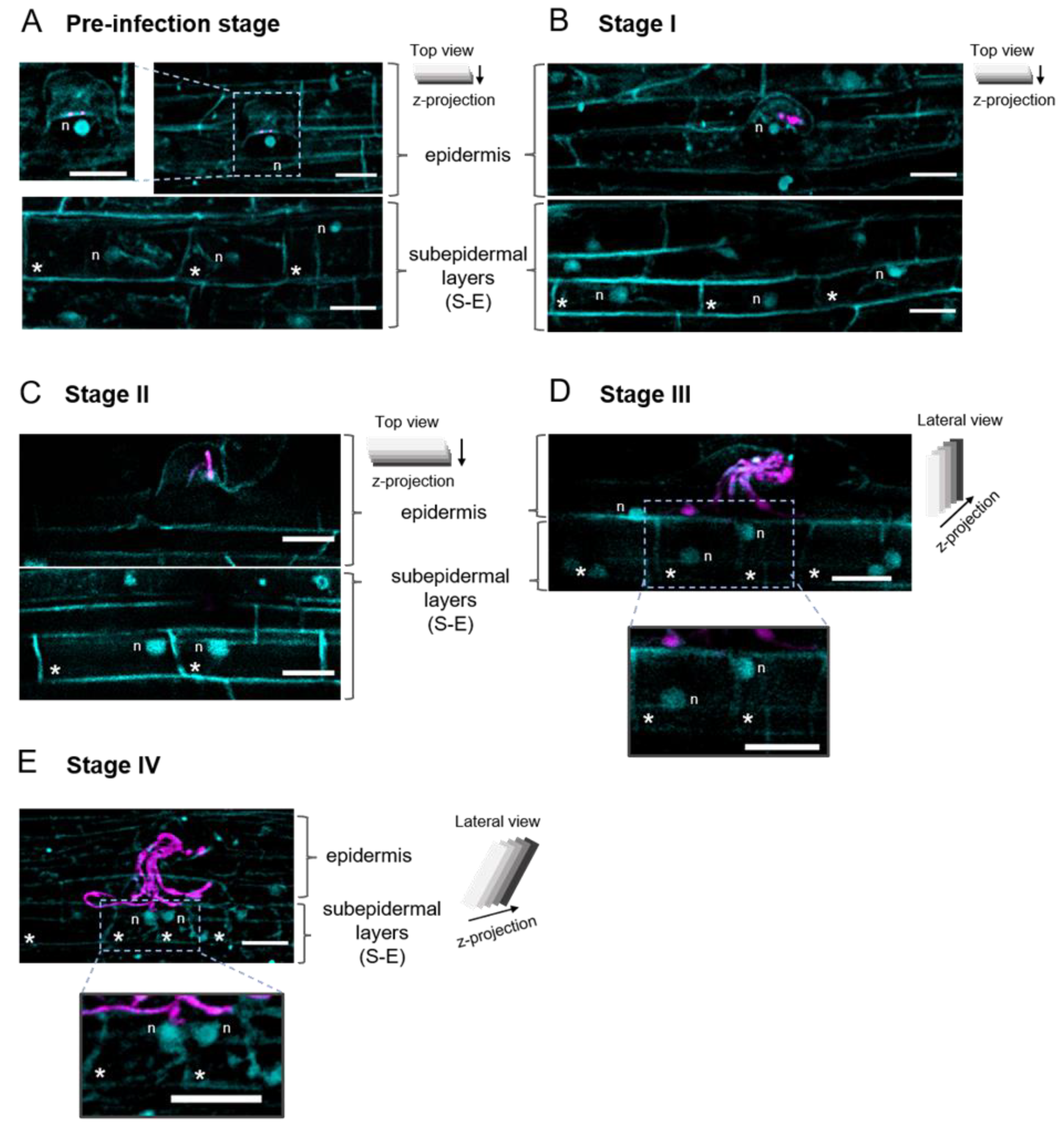
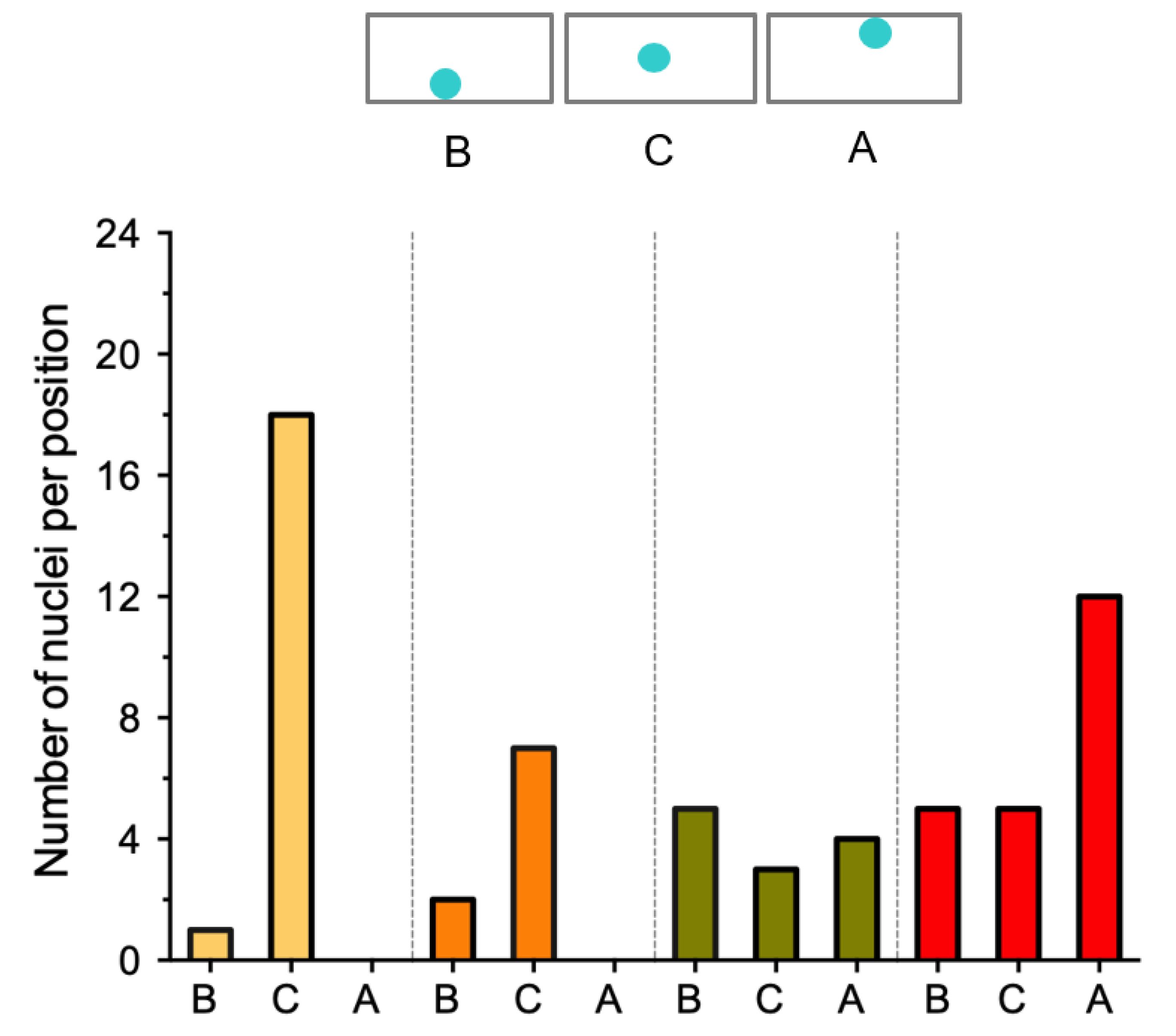
2.3. The Position of the Nucleus in the Subepidermal Cells Underlying the Epidermal Infection Site Correlates with the Infection Thread (IT) Progression
2.4. Cell Plate-Labeling with YFP-PvKNOLLE Indicates That the Infection Thread (IT) Penetrates the Subepidermal Cell (S-E) at Late Cytokinesis
3. Discussion
4. Materials and Methods
4.1. Plants and Bacteria Growth Conditions
4.2. Plasmid Constructions
4.3. Fixation and DAPI Staining Conditions
4.4. Confocal Imaging
4.5. Image Processing and 3D Visualization
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Patriarca, E.J.; Tatè, R.; Ferraioli, S.; Iaccarino, M. Organogenesis of Legume Root Nodules. Int. Rev. Cytol. 2004, 234, 201–262. [Google Scholar] [CrossRef] [PubMed]
- Guinel, F.C. Getting around the legume nodule: II. Molecular biology of its peripheral zone and approaches to study its vasculature. Botany 2009, 87, 1139–1166. [Google Scholar] [CrossRef]
- Xiao, T.T.; Schilderink, S.; Moling, S.; Deinum, E.E.; Kondorosi, E.; Franssen, H.; Kulikova, O.; Niebel, A.; Bisseling, T. Fate map of Medicago truncatula root nodules. Development 2014, 141, 3517–3528. [Google Scholar] [CrossRef] [Green Version]
- Van Spronsen, P.C.; Grønlund, M.; Bras, C.P.; Spaink, H.P.; Kijne, J.W. Cell biological changes of outer cortical root cells in early determinate nodulation. Mol. Plant-Microbe Interact. 2001, 14, 839–847. [Google Scholar] [CrossRef]
- Ferguson, B.J.; Indrasumunar, A.; Hayashi, S.; Lin, M.G.; Lin, Y.H.; Reid, D.E.; Gresshoff, P.M. Molecular analysis of legume nodule development and autoregulation. J. Integr. Plant. Biol. 2010, 52, 61–76. [Google Scholar] [CrossRef]
- Oldroyd, G.E.D.; Downie, J.A. Coordinating nodule morphogenesis with rhizobial infection in legumes. Annu. Rev. Plant Biol. 2008, 59, 519–546. [Google Scholar] [CrossRef]
- Madsen, L.H.; Tirichine, L.; Jurkiewicz, A.; Sullivan, J.T.; Heckmann, A.B.; Bek, A.S.; Ronson, C.W.; James, E.K.; Stougaard, J. The molecular network governing nodule organogenesis and infection in the model legume Lotus japonicus. Nat. Commun. 2010, 1, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lebedeva, M.; Azarakhsh, M.; Sadikova, D.; Lutova, L. At the root of nodule organogenesis: Conserved regulatory pathways recruited by rhizobia. Plants 2021, 10, 2654. [Google Scholar] [CrossRef]
- Murray, J.D. Invasion by invitation: Rhizobial infection in legumes. Mol. Plant-Microbe Interact. 2011, 24, 631–639. [Google Scholar] [CrossRef] [Green Version]
- Tsyganova, A.V.; Brewin, N.J.; Tsyganov, V.E. Structure and development of the legume-rhizobial symbiotic interface in infection threads. Cells 2021, 10, 1050. [Google Scholar] [CrossRef] [PubMed]
- White, J.; Prell, J.; James, E.K.; Poole, P. Nutrient sharing between symbionts. Plant Physiol. 2007, 144, 604–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzaki, T.; Ito, M.; Kawaguchi, M. Genetic basis of cytokinin and auxin functions during root nodule development. Front. Plant Sci. 2013, 4, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franssen, H.J.; Xiao, T.T.; Kulikova, O.; Wan, X.; Bisseling, T.; Scheres, B.; Heidstra, R. Root developmental programs shape the Medicago truncatula nodule meristem. Developement 2015, 142, 2941–2950. [Google Scholar] [CrossRef] [Green Version]
- Guinel, F.C. Ethylene, a hormone at the center-stage of nodulation. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, J.L.P.; Hassan, S.; Truong, T.T.; Hocart, C.H.; Laffont, C.; Frugier, F.; Mathesius, U. Flavonoids and auxin transport inhibitors rescue symbiotic nodulation in the Medicago truncatula cytokinin perception mutant cre1. Plant Cell. 2015, 27, 2210–2226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamas, P.; Brault, M.; Jardinaud, M.F.; Frugier, F. Cytokinins in Symbiotic Nodulation: When, Where, What For? Trends Plant Sci. 2017, 22, 792–802. [Google Scholar] [CrossRef] [PubMed]
- Kohlen, W.; Ng, J.L.P.; Deinum, E.E.; Mathesius, U. Auxin transport, metabolism, and signalling during nodule initiation: Indeterminate and determinate nodules. J. Exp. Bot. 2018, 69, 229–244. [Google Scholar] [CrossRef]
- Mergaert, P.; Kereszt, A.; Kondorosi, E. Gene Expression in Nitrogen-Fixing Symbiotic Nodule Cells in Medicago truncatula and Other Nodulating Plants. Plant Cell. 2020, 32, 42–68. [Google Scholar] [CrossRef]
- Suzaki, T.; Yano, K.; Ito, M.; Umehara, Y.; Suganuma, N.; Kawaguchi, M. Positive and negative regulation of cortical cell division during root nodule development in Lotus japonicus is accompanied by auxin response. Developement 2012, 139, 3997–4006. [Google Scholar] [CrossRef] [Green Version]
- Tan, S.; Sanchez, M.; Laffont, C.; Boivin, S.; Signor, C.L.; Thompson, R.; Frugier, F.; Brault, M. A cytokinin signaling type-b response regulator transcription factor acting in early nodulation1. Plant Physiol. 2020, 183, 1319–1330. [Google Scholar] [CrossRef]
- Catoira, R.; Galera, C.; de Billy, F.; Penmetsa, R.V.; Journet, E.P.; Maillet, F.; Rosenberg, C.; Cook, D.; Gough, C.; Dénarié, J. Four Genes of Medicago truncatula Controlling Components of a Nod Factor Transduction Pathway. Plant Cell. 2000, 12, 1647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endre, G.; Kereszt, A.; Kevei, Z.; Mihacea, S.; Kaló, P.; Kiss, G.B. A receptor kinase gene regulating symbiotic nodule development. Nature 2002, 417, 962–966. [Google Scholar] [CrossRef] [PubMed]
- Stracke, S.; Kistner, C.; Yoshida, S.; Mulder, L.; Sato, S.; Kaneko, T.; Tabata, S.; Sandal, N.; Stougaard, J.; Szczyglowski, K.; et al. A plant receptor-like kinase required for both bacterial and fungal symbiosis. Nature 2002, 417, 959–962. [Google Scholar] [CrossRef] [PubMed]
- Radutoiu, S.; Madsen, L.H.; Madsen, E.B.; Felle, H.H.; Umehara, Y.; Grønlund, M.; Sato, S.; Nakamura, Y.; Tabata, S.; Niels Sandal, N.; et al. Plant recognition of symbiotic bacteria requires two LysM receptor-like kinases. Nature 2003, 425, 585–592. [Google Scholar] [CrossRef]
- Sánchez-López, R.; Jáuregui, D.; Nava, N.; Alvarado-Affantranger, X.; Montiel, J.; Santana, O.; Sanchez, F.; Quinto, C. Down-regulation of SymRK correlates with a deficiency in vascular bundle development in Phaseolus vulgaris nodules. Plant Cell Environ. 2011, 34, 2109–2121. [Google Scholar] [CrossRef]
- Ke, D.; Fang, Q.; Chen, C.; Zhu, H.; Chen, T.; Chang, X.; Yuan, S.; Kang, H.; Ma, L.; Hong, Z.; et al. The small GTPase ROP6 interacts with NFR5 and is involved in nodule formation in Lotus japonicus. Plant Physiol. 2012, 159, 131–143. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Zhu, M.; Duan, L.; Alvarado-Affantranger, X.; Montiel, J.; Santana, O.; Sanchez, F.; Quinto, C. Lotus japonicus clathrin heavy chain1 is associated with rho-like GTPase ROP6 and involved in nodule formation. Plant Physiol. 2015, 167, 1497–1510. [Google Scholar] [CrossRef] [Green Version]
- Lei, M.J.; Wang, Q.; Li, X.; Chen, A.; Luo, L.; Xie, Y.; Li, G.; Luo, D.; Mysore, K.S.; Wen, J.; et al. The small GTPase ROP10 of Medicago truncatula is required for both tip growth of root hairs and nod factor-induced root hair deformation. Plant Cell. 2015, 27, 806–822. [Google Scholar] [CrossRef] [Green Version]
- Blanco, F.A.; Meschini, E.P.; Zanetti, M.E.; Aguilar, O.M. A small GTPase of the rab family is required for root hair formation and preinfection stages of the common bean-rhizobium symbiotic association. Plant Cell. 2009, 21, 2797–2810. [Google Scholar] [CrossRef] [Green Version]
- Via, D.V.; Traubenik, S.; Rivero, C.; Aguilar, O.M.; Zanetti, M.E.; Blanco, F.A. The monomeric GTPase RabA2 is required for progression and maintenance of membrane integrity of infection threads during root nodule symbiosis. Plant Mol. Biol. 2017, 93, 549–562. [Google Scholar] [CrossRef]
- García-Soto, I.; Boussageon, R.; Cruz-Farfán, Y.M.; Castro-Chilpa, J.D.; Hernández-Cerezo, L.X.; Bustos-Zagal, V.; Leija-Salas, A.; Hernández, G.; Torres, M.; Formey, D.; et al. The Lotus japonicus ROP3 Is Involved in the Establishment of the Nitrogen-Fixing Symbiosis but Not of the Arbuscular Mycorrhizal Symbiosis. Front. Plant Sci. 2021, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Fournier, J.; Teillet, A.; Chabaud, M.; Ivanov, S.; Genre, A.; Limpens, E.; Carvalho-Niebel, F.; Barker, D.G. Remodeling of the infection chamber before infection thread formation reveals a two-step mechanism for rhizobial entry into the host legume root hair. Plant Physiol. 2015, 167, 1233–1242. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Murray, J.D.; Kim, J.; Jurkiewicz, A.; Rueda, P.; Radutoiu, S.; Held, M.; Hossain, M.S.; Szczyglowski, K.; Morieri, G.; et al. Legume pectate lyase required for root infection by rhizobia. Proc. Natl. Acad. Sci. USA 2012, 109, 633–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokota, K.; Fukai, E.; Madsen, L.H.; Jurkiewicz, A.; Rueda, P.; Radutoiu, S.; Held, M.; Hossain, M.S.; Szczyglowski, K.; Morieri, G.; et al. Rearrangement of actin cytoskeleton mediates invasion of lotus japonicus roots by Mesorhizobium loti. Plant Cell. 2009, 21, 267–284. [Google Scholar] [CrossRef] [Green Version]
- Haney, C.H.; Long, S.R. Plant flotillins are required for infection by nitrogen-fixing bacteria. Proc. Natl. Acad. Sci. USA 2010, 107, 478–483. [Google Scholar] [CrossRef] [Green Version]
- Miyahara, A.; Richens, J.; Starker, C.; Morieri, G.; Smith, L.; Long, S.; Downie, J.A.; Oldroyd, G.E.D. Conservation in Function of a SCAR/WAVE Component during Infection Thread and Root Hair Growth in Medicago truncatula. Mol. Plant-Microbe Interact. 2010, 23, 1553–1562. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.S.; Liao, J.; James, E.K.; Sato, S.; Tabata, S.; Jurkiewicz, A.; Madsen, L.H.; Stougaard, S.; Ross, L.; Szczyglowski, K. Lotus japonicus ARPC1 is required for rhizobial infection. Plant Physiol. 2012, 160, 917–928. [Google Scholar] [CrossRef] [Green Version]
- Perrine-Walker, F.M.; Lartaud, M.; Kouchi, H.; Ridge, R.W. Microtubule array formation during root hair infection thread initiation and elongation in the Mesorhizobium-Lotus symbiosis. Protoplasma 2014, 251, 1099–1111. [Google Scholar] [CrossRef]
- Qiu, L.; Lin, J.S.; Xu, J.; Sato, S.; Parniske, M.; Wang, T.L.; Downie, J.A.; Xie, F. SCARN a Novel Class of SCAR Protein That Is Required for Root-Hair Infection during Legume Nodulation. PLoS Genet. 2015, 11, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Liang, P.; Schmitz, C.; Lace, B.; Ditengou, F.A.; Su, C.; Schulze, E.; Knerr, J.; Grosse, R.; Keller, J.; Libourel, C.; et al. Formin-mediated bridging of cell wall, plasma membrane, and cytoskeleton in symbiotic infections of Medicago truncatula. Curr. Biol. 2021, 31, 2712–2719.e5. [Google Scholar] [CrossRef]
- Den Herder, G.; De Keyser, A.; De Rycke, R.; Rombauts, S.; Van de Velde, W.; Clemente, M.R.; Verplancke, C.; Mergaert, P.; Kondorosi, E.; Holsters, M.; et al. Seven in absentia proteins affect plant growth and nodulation in Medicago truncatula. Plant Physiol. 2008, 148, 369–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yano, K.; Shibata, S.; Chen, W.L.; Sato, S.; Kaneko, T.; Jurkiewicz, A.; Sandal, N.; Banba, M.; Imaizumi-Anraku, H.; Kojima, T.; et al. CERBERUS, a novel U-box protein containing WD-40 repeats, is required for formation of the infection thread and nodule development in the legume-Rhizobium symbiosis. Plant J. 2009, 60, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Mbengue, M.; Camut, S.; de Carvalho-Niebel, F.; Deslandes, L.; Froidure, S.; Klaus-Heisen, D.; Moreau, S.; Rivas, S.; Timmers, T.; Hervé, C.; et al. The Medicago truncatula E3 ubiquitin ligase PUB1 interacts with the LYK3 symbiotic receptor and negatively regulates infection and nodulation. Plant Cell. 2010, 22, 3474–3488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsh, J.F.; Rakocevic, A.; Mitra, R.M.; Brocard, L.; Sun, J.; Eschstruth, A.; Long, S.R.; Schultze, M.; Ratet, P.; Oldroyd, G.E.D. Medicago truncatula NIN is essential for rhizobial-independent nodule organogenesis induced by autoactive calcium/calmodulin-dependent protein kinase. Plant Physiol. 2007, 144, 324–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yano, K.; Yoshida, S.; Müller, J.; Singh, S.; Banba, M.; Vickers, K.; Markmann, K.; White, C.; Schuller, B.; Sato, S.; et al. CYCLOPS, a mediator of symbiotic intracellular accommodation. Proc. Natl. Acad. Sci. USA 2008, 105, 20540–20545. [Google Scholar] [CrossRef] [Green Version]
- Zanetti, M.E.; Blanco, F.A.; Beker, M.P.; Battaglia, M.; Aguilar, O.M. A C subunit of the plant nuclear factor NF-Y required for rhizobial infection and nodule development affects partner selection in the common bean-Rhizobium etli Symbiosis. Plant Cell. 2010, 22, 4142–4157. [Google Scholar] [CrossRef] [Green Version]
- Laporte, P.; Lepage, A.; Fournier, J.; Catrice, O.; Moreau, S.; Jardinaud, M.F.; Mun, J.H.; Larrainzar, E.; Cook, D.R.; Gamas, P.; et al. The CCAAT box-binding transcription factor NF-YA1 controls rhizobial infection. J. Exp. Bot. 2014, 65, 481–494. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Katzer, K.; Lambert, J.; Cerri, M.; Parniske, M. CYCLOPS, a DNA-binding transcriptional activator, orchestrates symbiotic root nodule development. Cell Host Microbe 2014, 15, 139–152. [Google Scholar] [CrossRef] [Green Version]
- Vernié, T.; Kim, J.; Frances, L.; Ding, Y.; Sun, J.; Guan, D.; Niebel, A.; Gifford, M.L.; Carvalho-Niebel, F.; Oldroyd, G.E.D. The NIN transcription factor coordinates diverse nodulation programs in different tissues of the Medicago truncatula root. Plant Cell. 2015, 27, 3410–3424. [Google Scholar] [CrossRef] [Green Version]
- Cerri, M.R.; Frances, L.; Kelner, A.; Fournier, J.; Middleton, P.H.; Auriac, M.H.; Mysore, K.S.; Wen, J.; Erard, M.; Barker, D.G.; et al. The symbiosis-related ERN transcription factors act in concert to coordinate rhizobial host root infection. Plant Physiol. 2016, 171, 1037–1054. [Google Scholar] [CrossRef]
- Kawaharada, Y.; James, E.K.; Kelly, S.; Sandal, N.; Stougaard, J. The ethylene responsive factor required for nodulation 1 (ERN1) transcription factor is required for infection-thread formation in Lotus japonicus. Mol. Plant-Microbe Interact. 2017, 30, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Soyano, T.; Kouchi, H.; Hirota, A.; Hayashi, M. NODULE INCEPTION Directly Targets NF-Y Subunit Genes to Regulate Essential Processes of Root Nodule Development in Lotus japonicus. PLOS Genet. 2013, 9, e1003352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breakspear, A.; Liu, C.; Roy, S.; Stacey, N.; Rogers, C.; Trick, M.; Morieri, G.; Mysore, K.S.; Wen, J.; Oldroyd, G.E.D.; et al. The root hair “infectome” of Medicago truncatula uncovers changes in cell cycle genes and reveals a requirement for auxin signaling in rhizobial infectionw. Plant Cell. 2014, 26, 4680–4701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maekawa, T.; Maekawa-Yoshikawa, M.; Takeda, N.; Imaizumi-Anraku, H.; Murooka, Y.; Hayashi, M. Gibberellin controls the nodulation signaling pathway in Lotus japonicus. Plant J. 2009, 58, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Liu, H.; Luo, D.; Yu, N.; Dong, W.; Wang, C.; Zhang, X.; Dai, H.; Yang, J.; Wang, E. DELLA proteins are common components of symbiotic rhizobial and mycorrhizal signalling pathways. Nat. Commun. 2016, 7, 1–14. [Google Scholar] [CrossRef]
- Dong, W.; Zhu, Y.; Chang, H.; Wang, C.; Yang, J.; Shi, J.; Gao, J.; Yang, W.; Lan, L.; Wang, Y.; et al. An SHR–SCR module specifies legume cortical cell fate to enable nodulation. Nature 2021, 589, 586–590. [Google Scholar] [CrossRef]
- Heidstra, R.; Yang, W.C.; Yalcin, Y.; Peck, S.; Emons, A.; Kammen, A.; Bisseling, T. Ethylene provides positional information on cortical cell division but is not involved in Nod factor-induced root hair tip growth in Rhizobium-legume interaction. Development 1997, 124, 1781–1787. [Google Scholar] [CrossRef]
- Pacios-Bras, C.; Schlaman, H.R.M.; Boot, K.; Admiraal, P.; Langerak, J.L.; Stougaard, J.; Spaink, H.P. Auxin distribution in Lotus japonicus during root nodule development. Plant Mol. Biol. 2003, 52, 1169–1180. [Google Scholar] [CrossRef]
- Roudier, F.; Fedorova, E.; Lebris, M.; Lecomte, P.; Györgyey, J.; Vaubert, D.; Horvath, G.; Abad, P.; Kondorosi, A.; Kondorosi, E. The Medicago species A2-type cyclin is auxin regulated and involved in meristem formation but dispensable for endoreduplication-associated developmental programs. Plant Physiol. 2003, 131, 1091–1103. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Rizzo, S.; Crespi, M.; Frugier, F. The Medicago truncatula CRE1 cytokinin receptor regulates lateral root development and early symbiotic interaction with Sinorhizobium meliloti. Plant Cell. 2006, 18, 2680–2693. [Google Scholar] [CrossRef] [Green Version]
- Prayitno, J.; Rolfe, B.G.; Mathesius, U. The ethylene-insensitive sickle mutant of Medicago truncatula shows altered auxin transport regulation during nodulation. Plant Physiol. 2006, 142, 168–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, J.D.; Karas, B.J.; Sato, S.; Tabata, S.; Amyot, L.; Szczyglowski, K. A cytokinin perception mutant colonized by Rhizobium in the absence of nodule organogenesis. Science 2007, 315, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.C.; De Blank, C.; Meskiene, I.; Hirt, H.; Bakker, J.; van Kammen, A.; Franssen, H.; Bisseling, T. Rhizobium nod factors reactivate the cell cycle during infection and nodule primordium formation, but the cycle is only completed in primordium formation. Plant Cell. 1994, 6, 1415–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Libbenga, K.R.; van Iren, F.; Bogers, R.J.; Schraag-Lamers, M.F. The role of hormones and gradients in the initiation of cortex proliferation and nodule formation in Pisum sativum L. Planta 1973, 114, 29–39. [Google Scholar] [CrossRef]
- Mathesius, U.; Schlaman, H.R.M.; Spaink, H.P.; Sautter, C.; Rolfe, B.G.; Djordjevic, M.A. Auxin transport inhibition precedes root nodule formation in white clover roots and is regulated by flavonoids and derivatives of chitin oligosaccharides. Plant J. 1998, 14, 23–34. [Google Scholar] [CrossRef]
- Nizampatnam, N.R.; Schreier, S.J.; Damodaran, S.; Adhikari, S.; Subramanian, S. MicroRNA160 dictates stage-specific auxin and cytokinin sensitivities and directs soybean nodule development. Plant J. 2015, 84, 140–153. [Google Scholar] [CrossRef]
- Tirichine, L.; Sandal, N.; Madsen, L.H.; Radutoiu, S.; Albrektsen, A.S.; Sato, S.; Asamizu, E.; Tabata, S.; Stougaard, J. A gain-of-function mutation in a cytokinin receptor triggers spontaneous root nodule organogenesis. Science 2007, 315, 104–107. [Google Scholar] [CrossRef] [Green Version]
- Tate, R.; Patriarca, E.J.; Riccio, A.; Defez, R.; Iaccarino, M. Development of Phaseolus vulgaris root nodules. Mol. Plant-Microbe Interact. 1994, 7, 582–589. [Google Scholar] [CrossRef]
- Jürgens, G. Cytokinesis in higher plants. Annu. Rev. Plant Biol. 2005, 56, 281–299. [Google Scholar] [CrossRef]
- Lauber, M.H.; Waizenegger, I.; Steinmann, T.; Schwarz, H.; Mayer, U.; Hwang, I.; Lukowitz, W.; Jürgens, G. The Arabidopsis KNOLLE protein is a cytokinesis-specific syntaxin. J. Cell Biol. 1997, 139, 1485–1493. [Google Scholar] [CrossRef] [Green Version]
- Touihri, S.; Knöll, C.; Stierhof, Y.D.; Müller, I.; Mayer, U.; Jürgens, G. Functional anatomy of the Arabidopsis cytokinesis-specific syntaxin KNOLLE. Plant J. 2011, 68, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Lukowitz, W.; Mayer, U.; Jürgens, G. Cytokinesis in the Arabidopsis embryo involves the syntaxin-related KNOLLE gene product. Cell 1996, 84, 61–71. [Google Scholar] [CrossRef] [Green Version]
- Menges, M.; De Jager, S.M.; Gruissem, W.; Murray, J.A.H. Global analysis of the core cell cycle regulators of Arabidopsis identifies novel genes, reveals multiple and highly specific profiles of expression and provides a coherent model for plant cell cycle control. Plant J. 2005, 41, 546–566. [Google Scholar] [CrossRef]
- Reichardt, I.; Stierhof, Y.D.; Mayer, U.; Richter, S.; Schwarz, H.; Schumacher, K.; Jürgens, G. Plant Cytokinesis Requires De Novo Secretory Trafficking but Not Endocytosis. Curr. Biol. 2007, 17, 2047–2053. [Google Scholar] [CrossRef] [Green Version]
- Dávila-Delgado, R.; Bañuelos-Vazquez, L.A.; Monroy-Morales, E.; Sánchez-López, R. Rhizobium etli CE3-DsRed pMP604: A useful biological tool to study initial infection steps in Phaseolus vulgaris nodulation. Planta 2020, 252, 1–6. [Google Scholar] [CrossRef]
- Charon, C.; Johansson, C.; Kondorosi, E.; Kondorosi, A.; Crespi, M. Enod40 Induces Dedifferentiation and Division of Root Cortical Cells in Legumes. Proc. Natl. Acad. Sci. USA 1997, 94, 8901–8906. [Google Scholar] [CrossRef] [Green Version]
- Nadzieja, M.; Stougaard, J.; Reid, D. A Toolkit for High Resolution Imaging of Cell Division and Phytohormone Signaling in Legume Roots and Root Nodules. Front. Plant Sci. 2019, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.W.; Breakspear, A.; Stacey, N.; Findlay, K.; Nakashima, J.; Ramakrishnan, K.; Liu, M.; Xie, F.; Endre, G.; de Carvalho-Niebel, F.; et al. A protein complex required for polar growth of rhizobial infection threads. Nat. Commun. 2019, 10, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Fournier, J.; Timmers, A.C.J.; Sieberer, B.J.; Jauneau, A.; Chabaud, M.; Barker, D.G. Mechanism of infection thread elongation in root hairs of Medicago truncatula and dynamic interplay with associated rhizobial colonization. Plant Physiol. 2008, 148, 1985–1995. [Google Scholar] [CrossRef] [Green Version]
- Ito, M.; Araki, S.; Matsunaga, S.; Itoh, T.; Nishihama, R.; Machida, Y.; Doonan, J.H.; Watanabe, A. G2/M-phase-specific transcription during the plant cell cycle is mediated by c-Myb-like transcription factors. Plant Cell. 2001, 13, 1891–1905. [Google Scholar] [CrossRef]
- Schütz, L.M.; Louveaux, M.; Vilches Barro, A.; Bouziri, S.; Cerrone, L.; Wolny, A.; Kreshuk, A.; Hamprecht, F.A.; Maize, A. Integration of Cell Growth and Asymmetric Division during Lateral Root Initiation in Arabidopsis thaliana. Plant Cell Physiol. 2021, 62, 1269–1279. [Google Scholar] [CrossRef] [PubMed]
- van Oostende-Triplet, C.; Guillet, D.; Triplet, T.; Pandzic, E.; Wiseman, P.W.; Geitmann, A. Vesicle dynamics during plant cell cytokinesis reveals distinct developmental phases. Plant Physiol. 2017, 174, 1544–1558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gage, D.J. Infection and Invasion of Roots by Symbiotic, Nitrogen-Fixing Rhizobia during Nodulation of Temperate Legumes. Microbiol. Mol. Biol. Rev. 2004, 68, 280–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrine-Walker, F.M.; Kouchi, H.; Ridge, R.W. Endoplasmic reticulum-targeted GFP reveals ER remodeling in Mesorhizobium-treated Lotus japonicus root hairs during root hair curling and infection thread formation. Protoplasma 2014, 251, 817–826. [Google Scholar] [CrossRef] [PubMed]
- Timmers, A.C.J.; Auriac, M.C.; Truchet, G. Refined analysis of early symbiotic steps of the Rhizobium-Medicago interaction in relationship with microtubular cytoskeleton rearrangements. Development 1999, 126, 3617–3628. [Google Scholar] [CrossRef]
- Van Brussel, A.A.N.; Bakhuizen, R.; Van Spronsen, P.C.; Spaink, H.P.; Tak, T.; Lugtenberg, B.J.J.; Kijne, J.W. Induction of pre-infection thread structures in the leguminous host plant by mitogenic lipo-oligosaccharides of rhizobium. Science 1992, 257, 70–72. [Google Scholar] [CrossRef]
- Esau, K.; Gill, R.H. Distribution of Vacuoles and Some Other Organelles in Dividing Cells. Bot. Gaz. 1991, 152, 397–407. [Google Scholar] [CrossRef]
- Bakhuizen, R.; van Spronsen, P.C.; Sluiman-den Hertog, F.A.J.; Venverloo, C.J.; Goosen-de Roo, L. Nuclear envelope radiating microtubules in plant cells during interphase mitosis transition. Protoplasma 1985, 128, 43–51. [Google Scholar] [CrossRef]
- Kutsuna, N.; Hasezawa, S. Dynamic organization of vacuolar and microtubule structures during cell cycle progression in synchronized tobacco BY-2 cells. Plant Cell Physiol. 2002, 43, 965–973. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.J.; Smith, L.G. Division plane orientation in plant cells. Plant Cell Monogr. 2007, 9, 33–57. [Google Scholar] [CrossRef]
- Tsyganov, V.E.; Voroshilova, V.A.; Priefer, U.B.; Borisov, A.Y.; Tikhonovich, I.A. Genetic dissection of the initiation of the infection process and nodule tissue development in the Rhizobium-pea (Pisum sativum L.) symbiosis. Ann. Bot. 2002, 89, 357–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, P.; Ratet, P.; Crespi, M.D.; Schultze, M.; Kondorosi, A. Nod factors and cytokinins induce similar cortical cell division, amyloplast deposition and MsEnoD12A expression patterns in alfalfa roots. Plant J. 1996, 10, 91–105. [Google Scholar] [CrossRef]
- Reid, D.; Nadzieja, M.; Novák, O.; Heckmann, A.B.; Sandal, N.; Stougaard, J. Cytokinin biosynthesis promotes cortical cell responses during nodule development. Plant Physiol. 2017, 175, 361–375. [Google Scholar] [CrossRef] [PubMed]
- Heckmann, A.B.; Sandal, N.; Bek, A.S.; Madsen, L.H.; Jurkiewicz, A.; Nielsen, M.W.; Tirichine, L.; Stougaard, J. Cytokinin induction of root nodule primordia in Lotus japonicus is regulated by a mechanism operating in the root cortex. Mol. Plant-Microbe Interact. 2011, 24, 1385–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasternak, T.; Kircher, S.; Palme, K. Estimation of differential cell cycle kinetics in higher plant root meristem with cellular fate and positional resolution. bioRxiv 2021. [Google Scholar] [CrossRef]
- Fåhraeus, G. The infection of clover root hairs by nodule bacteria studied by a simple glass slide technique. J. Gen. Microbiol. 1957, 16, 374–381. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.; Ruan, Z.; Long, F.; Simpson, J.H.; Myers, E.W. V3D enables real-time 3D visualization and quantitative analysis of large-scale biological image data sets. Nat. Biotechnol. 2010, 28, 348–353. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monroy-Morales, E.; Dávila-Delgado, R.; Ayala-Guzmán, E.; Gamboa-deBuen, A.; Sánchez-López, R. Visualization of the Crossroads between a Nascent Infection Thread and the First Cell Division Event in Phaseolus vulgaris Nodulation. Int. J. Mol. Sci. 2022, 23, 5267. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095267
Monroy-Morales E, Dávila-Delgado R, Ayala-Guzmán E, Gamboa-deBuen A, Sánchez-López R. Visualization of the Crossroads between a Nascent Infection Thread and the First Cell Division Event in Phaseolus vulgaris Nodulation. International Journal of Molecular Sciences. 2022; 23(9):5267. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095267
Chicago/Turabian StyleMonroy-Morales, Elizabeth, Raúl Dávila-Delgado, Emmanuel Ayala-Guzmán, Alicia Gamboa-deBuen, and Rosana Sánchez-López. 2022. "Visualization of the Crossroads between a Nascent Infection Thread and the First Cell Division Event in Phaseolus vulgaris Nodulation" International Journal of Molecular Sciences 23, no. 9: 5267. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095267