
Comparison of the Biochemical Properties and Roles in the Xyloglucan-Rich Biomass Degradation of a GH74 Xyloglucanase and Its CBM-Deleted Variant from Thielavia terrestris


Abstract
:1. Introduction
2. Results and Discussion
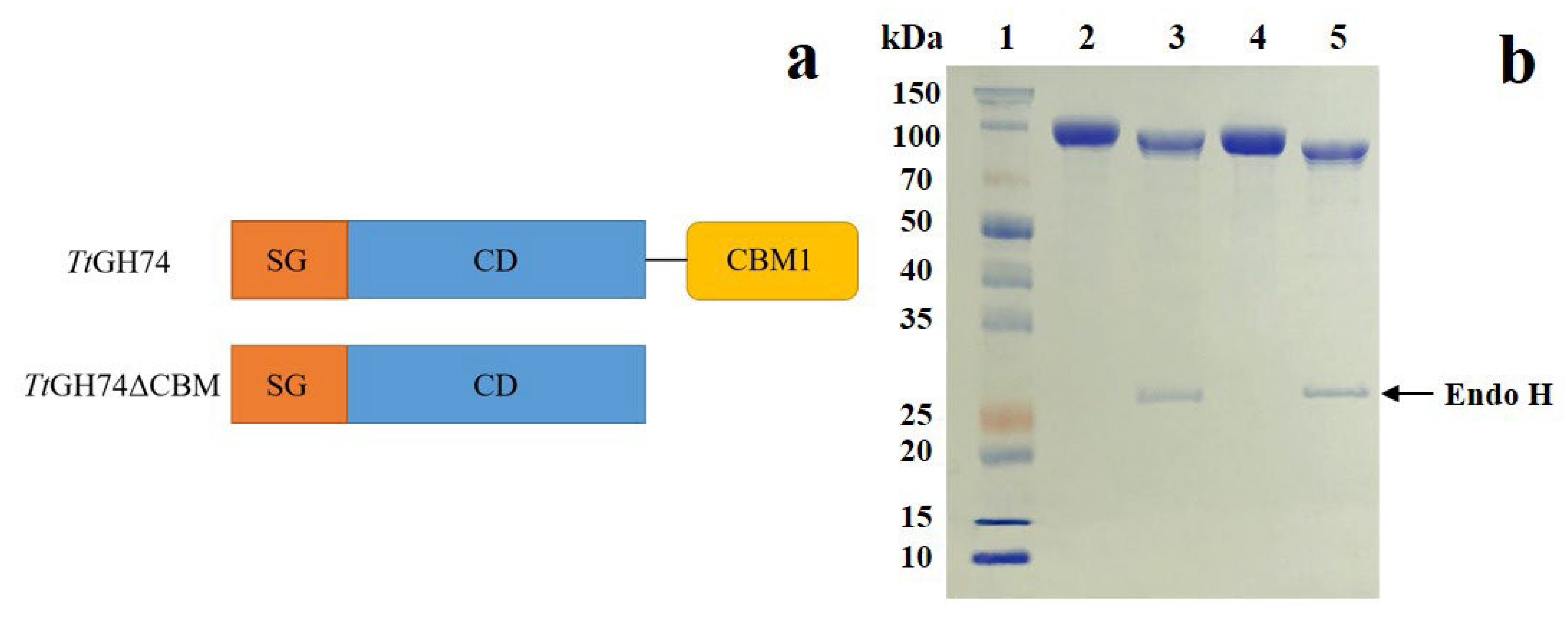
2.1. Expression and Purification of TtGH74 and TtGH74ΔCBM
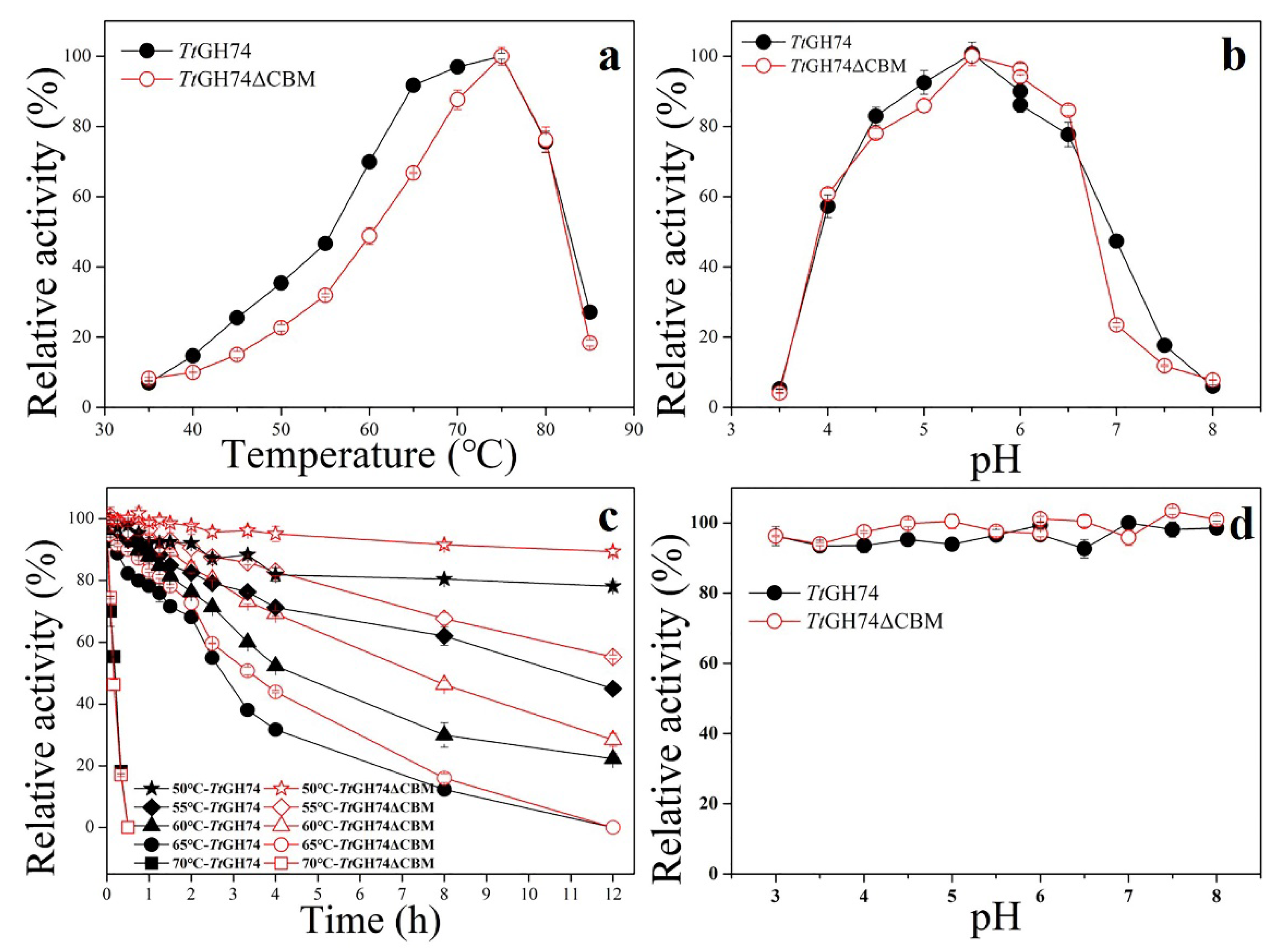
2.2. Properties of Recombinant TtGH74 and TtGH74ΔCBM from P. pastoris
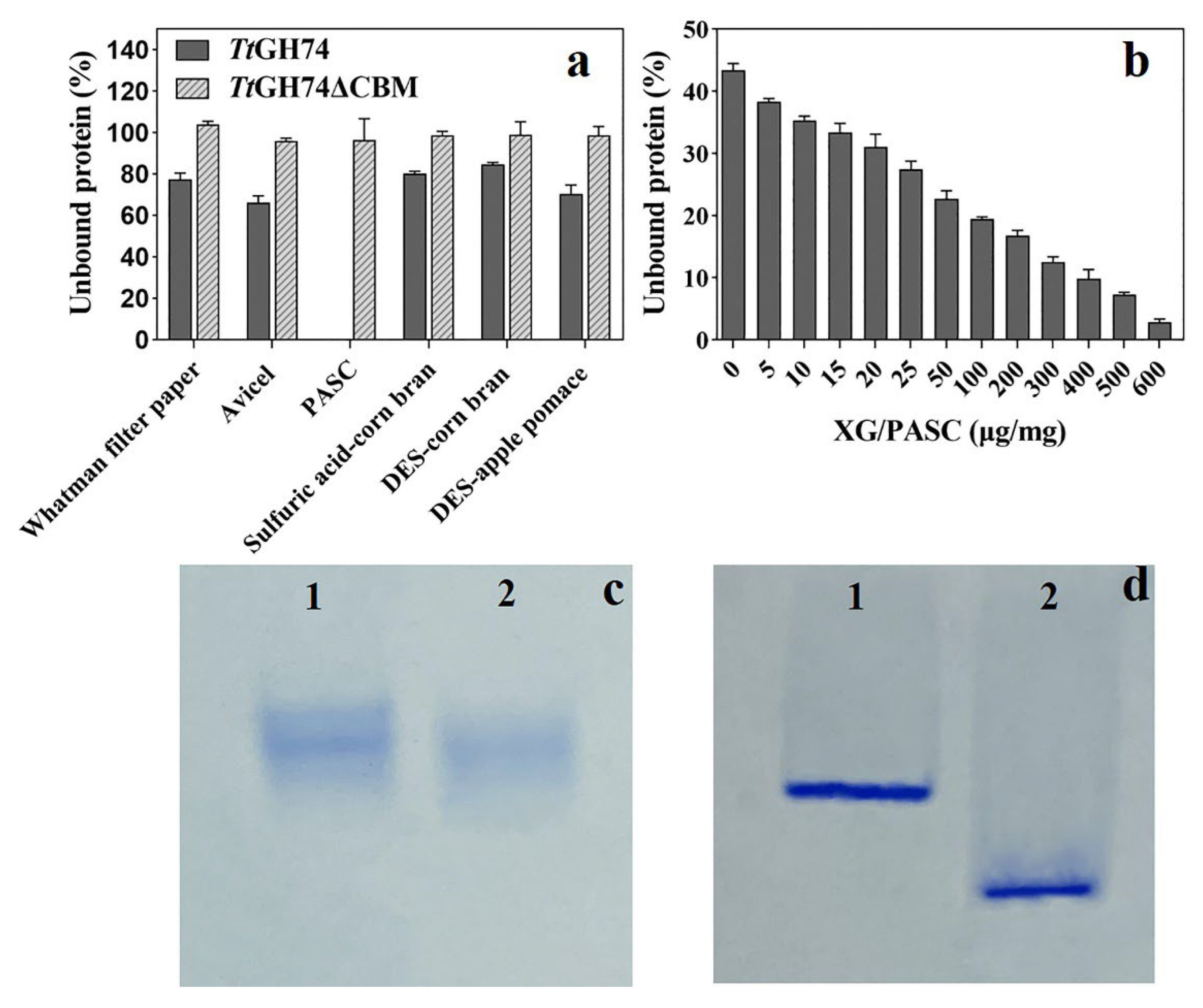
2.3. Adsorption of TtGH74 and TtGH74ΔCBM on Different Substrates
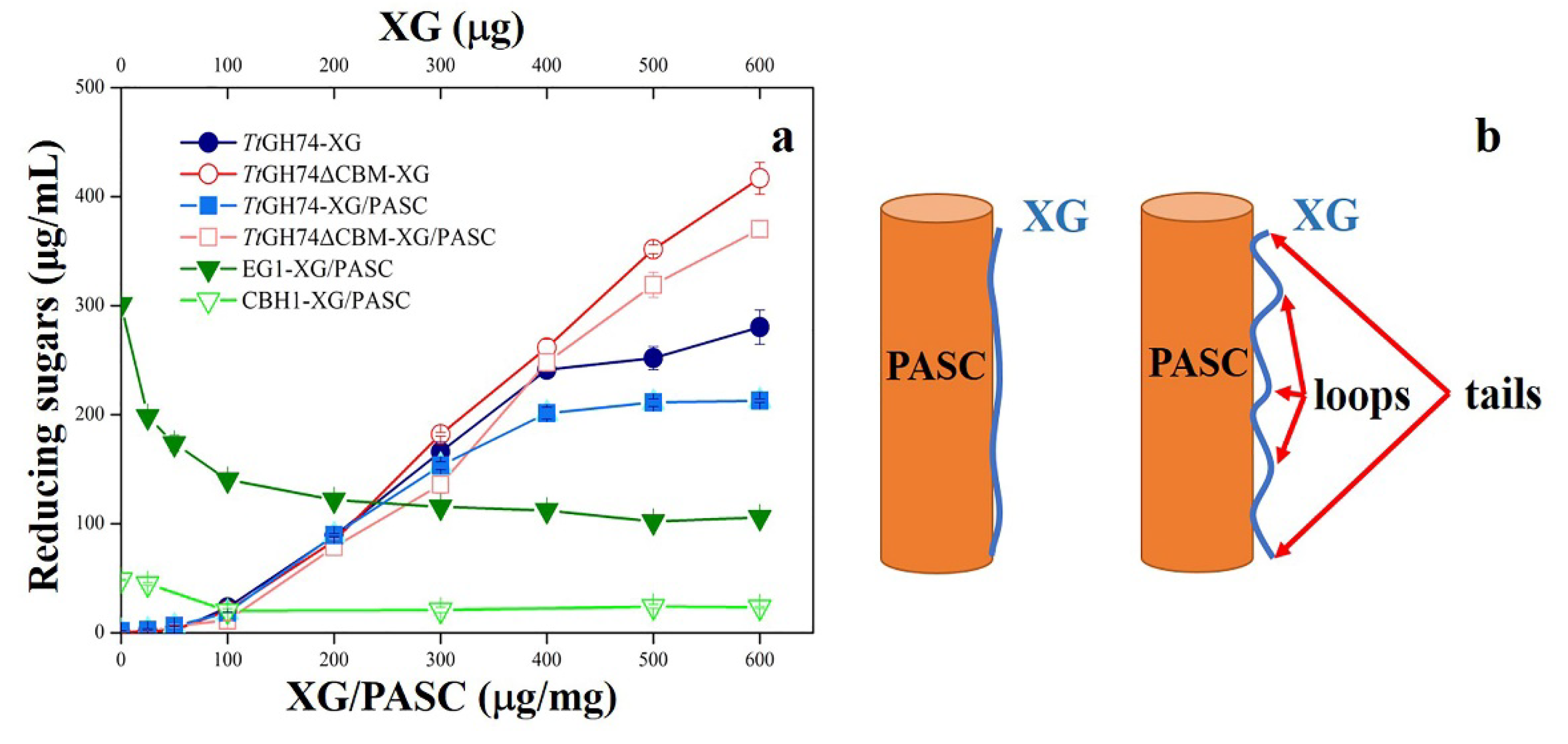
2.4. Hydrolysis Action of TtGH74 and TtGH74ΔCBM on XG and XG-Coated PASC
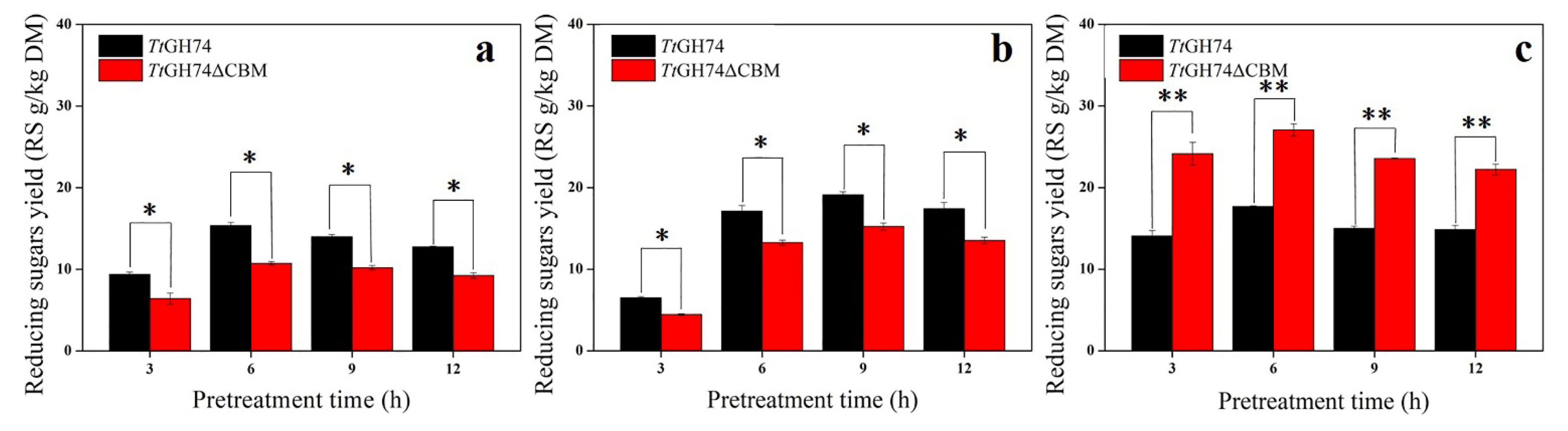
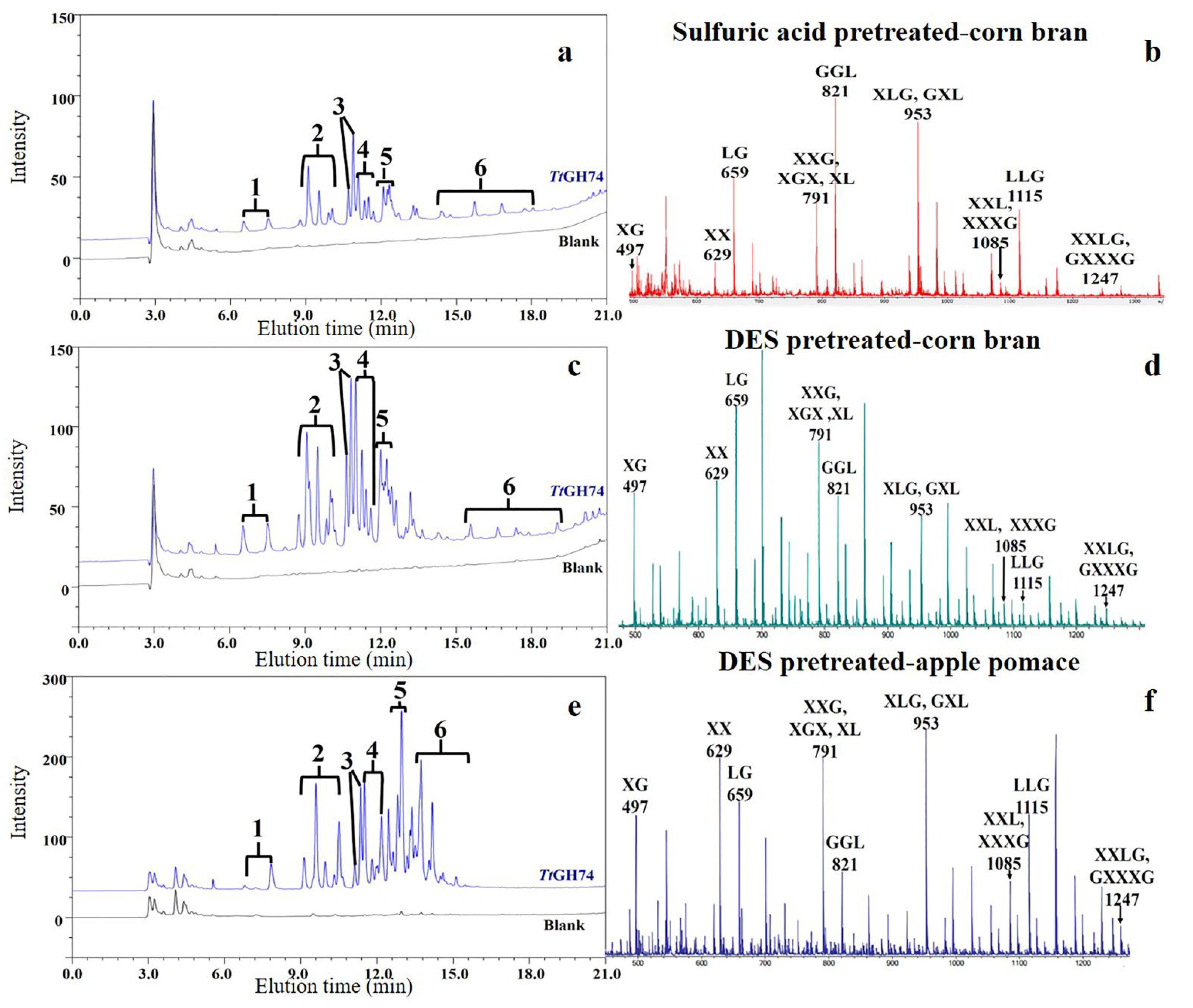
2.5. Evaluation of the Presence of XG and Hydrolysis Action of TtGH74 and TtGH74ΔCBM on Pretreated Corn Bran and Apple Pomace
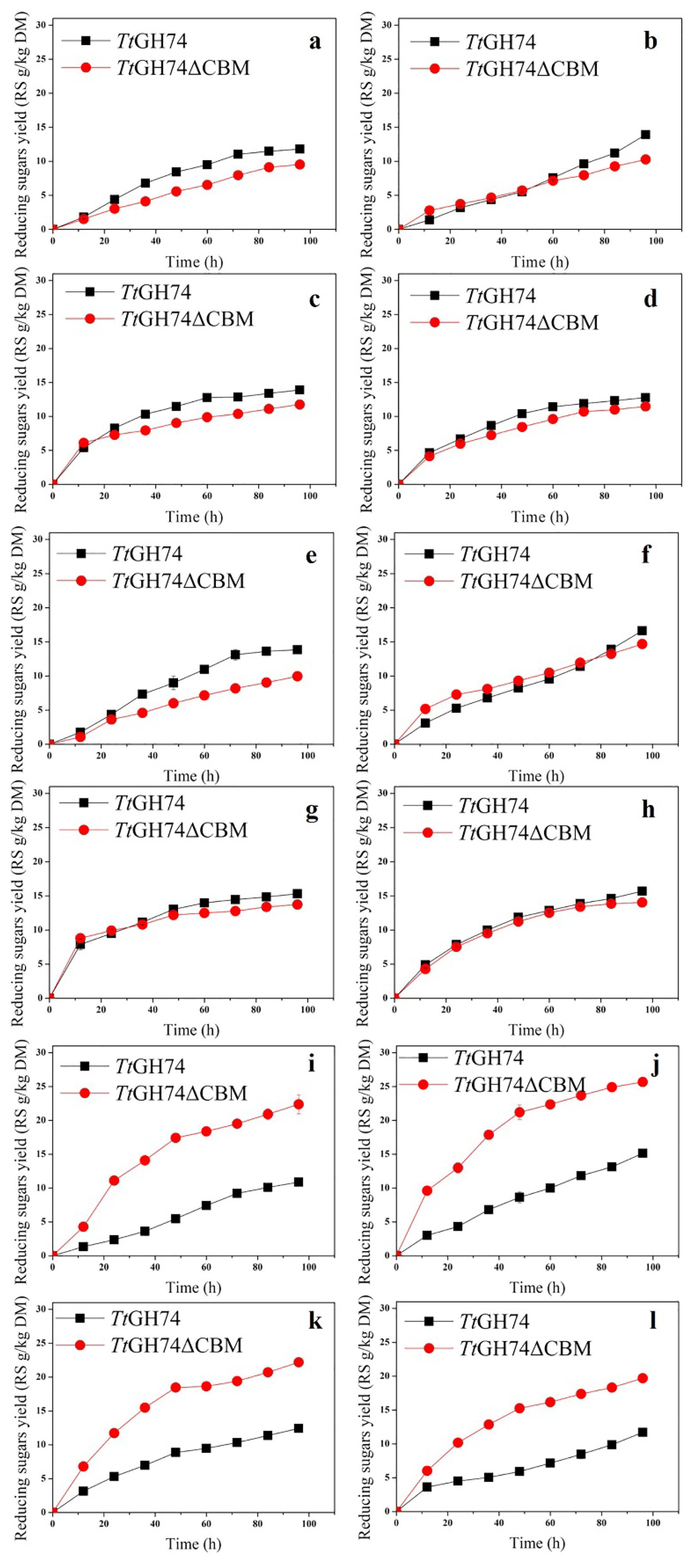
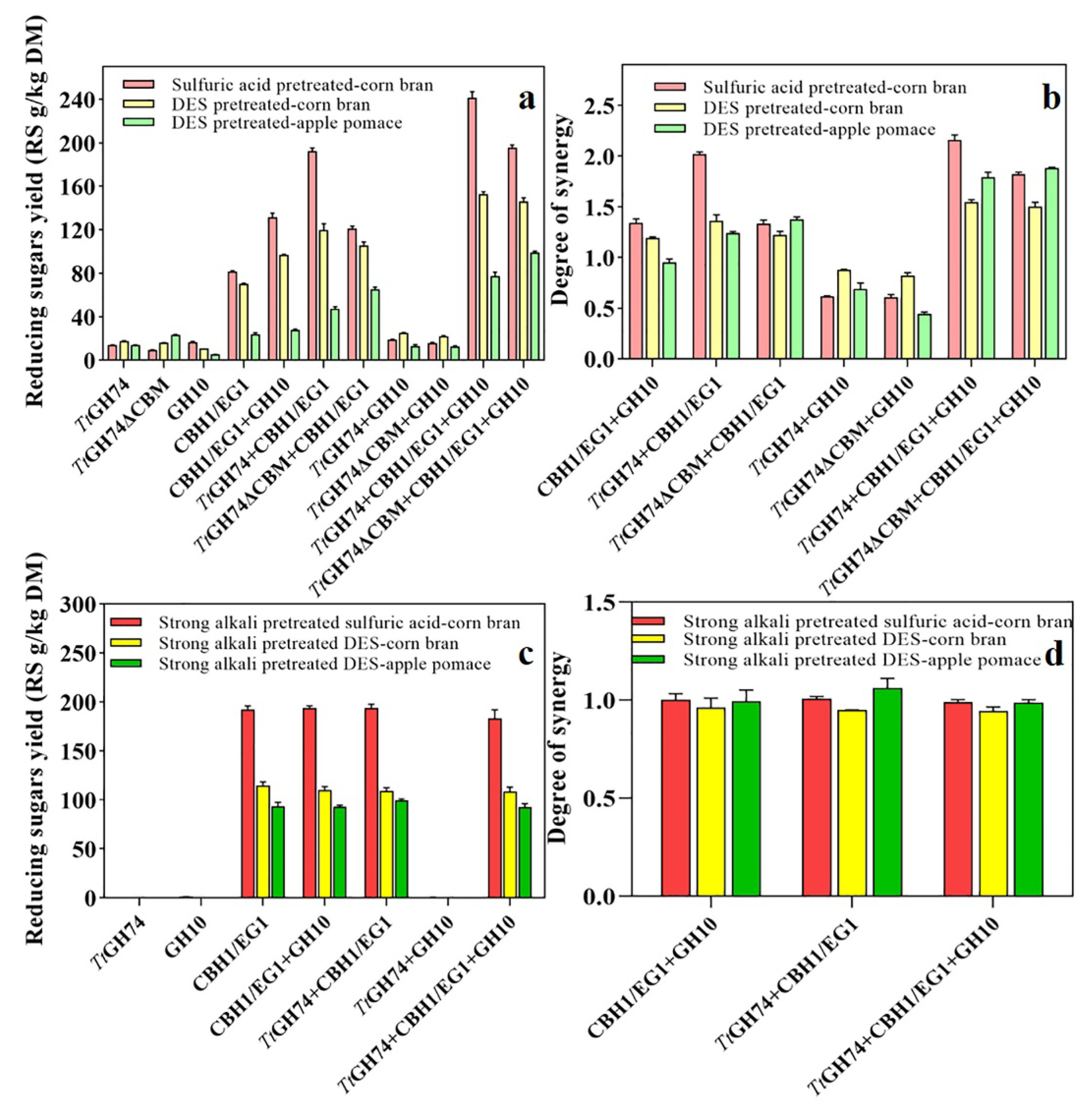
2.6. Synergistic Action of TtGH74 or TtGH74ΔCBM with CBH1/EG1 Mixture and Xylanase on Pretreated Corn Bran and Apple Pomace
3. Materials and Methods
3.1. Materials
3.2. Construction of TtGH74 and TtGH74ΔCBM
3.3. Expression and Purification of TtGH74 and TtGH74ΔCBM
3.4. Properties of Recombinant TtGH74 and TtGH74ΔCBM from P. pastoris
3.5. Adsorption of TtGH74 and TtGH74ΔCBM on Different Substrates
3.6. Hydrolysis Action of TtGH74 and TtGH74ΔCBM on XG and XG-coated PASC
3.7. Analysis of the Presence of XG and Hydrolysis Action of TtGH74 or TtGH74ΔCBM on Pretreated Residues
3.8. Synergistic Action of TtGH74 or TtGH74ΔCBM with CBH1/EG1 Mixture and Xylanase on Pretreated Lignocellulose
3.9. Test Method
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Elyamny, S.; Hamdy, A.; Ali, R.; Hamad, H. Role of Combined Na2HPO4 and ZnCl2 in the Unprecedented Catalysis of the Sequential Pretreatment of Sustainable Agricultural and Agro-Industrial Wastes in Boosting Bioethanol Production. Int. J. Mol. Sci. 2022, 23, 1777. [Google Scholar] [CrossRef] [PubMed]
- Huber, G.W.; Iborra, S.; Corma, A. Synthesis of Transportation Fuels from Biomass: Chemistry, Catalysts, and Engineering. Chem. Rev. 2006, 106, 4044–4098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauly, M.; Keegstra, K. Biosynthesis of the Plant Cell Wall Matrix Polysaccharide Xyloglucan. Annu. Rev. Plant Biol. 2016, 67, 235–259. [Google Scholar] [CrossRef] [PubMed]
- Somerville, C.; Bauer, S.; Brininstool, G.; Facette, M.; Hamann, T.; Milne, J.; Osborne, E.; Paredez, A.; Persson, S.; Raab, T.; et al. Toward a systems approach to understanding plant cell walls. Science 2004, 306, 2206–2211. [Google Scholar] [CrossRef] [Green Version]
- Ebringerová, A. Structural Diversity and Application Potential of Hemicelluloses. Macromol. Symp. 2005, 232, 1–12. [Google Scholar] [CrossRef]
- McNeil, M.; Darvill, A.G.; Fry, S.C.; Albersheim, P. Structure and function of the primary cell walls of plants. Annu. Rev. Biochem. 1984, 53, 625–663. [Google Scholar] [CrossRef]
- Fry, S.C. The Structure and Functions of Xyloglucan. J. Exp. Bot. 1989, 40, 1–11. [Google Scholar] [CrossRef]
- Fry, S.C.; York, W.S.; Albersheim, P.; Darvill, A.; Hayashi, T.; Joseleau, J.-P.; Kato, Y.; Lorences, E.P.; Maclachlan, G.A.; McNeil, M.; et al. An unambiguous nomenclature for xyloglucan-derived oligosaccharides. Physiol. Plant. 1993, 89, 1–3. [Google Scholar] [CrossRef]
- Zabotina, O.A. Xyloglucan and Its Biosynthesis. Front. Plant Sci. 2012, 3, 134. [Google Scholar] [CrossRef] [Green Version]
- Østby, H.; Hansen, L.D.; Horn, S.J.; Eijsink, V.G.H.; Várnai, A. Enzymatic processing of lignocellulosic biomass: Principles, recent advances and perspectives. J. Ind. Microbiol. Biotechnol. 2020, 47, 623–657. [Google Scholar] [CrossRef]
- Zoghlami, A.; Paës, G. Lignocellulosic Biomass: Understanding Recalcitrance and Predicting Hydrolysis. Front. Chem. 2019, 7, 874. [Google Scholar] [CrossRef] [Green Version]
- Kothari, N.; Bhagia, S.; Pu, Y.; Yoo, C.G.; Li, M.; Venketachalam, S.; Pattathil, S.; Kumar, R.; Cai, C.M.; Hahn, M.G.; et al. The effects of switchgrass plant cell wall properties on its deconstruction by thermochemical pretreatments coupled with fungal enzymatic hydrolysis or Clostridium thermocellum consolidated bioprocessing. Green Chem. 2020, 22, 7924–7945. [Google Scholar] [CrossRef]
- Shi, Y.X.; Chen, K.X.; Long, L.K.; Ding, S.J. A highly xyloglucan active lytic polysaccharide monooxygenase EpLPMO9A from Eupenicillium parvum 4-14 shows boosting effect on hydrolysis of complex lignocellulosic substrates. Int. J. Biol. Macromol. 2021, 167, 202–213. [Google Scholar] [CrossRef]
- Pauly, M.; Andersen, L.N.; Kauppinen, S.; Kofod, L.V.; York, W.S.; Albersheim, P.; Darvill, A. A xyloglucan-specific endo-beta-1,4-glucanase from Aspergillus aculeatus: Expression cloning in yeast, purification and characterization of the recombinant enzyme. Glycobiology 1999, 9, 93–100. [Google Scholar] [CrossRef]
- Gusakov, A.V.; Uporov, I.V.; Sinitsyna, O.A. Molecular dynamics simulations of two GH74 endo-processive xyloglucanases and the mutated variants to understand better the mechanism of the enzyme action. Biochim. Biophys. Acta-Gen. Subj. 2020, 1864, 129721. [Google Scholar] [CrossRef]
- Matsuzawa, T.; Kameyama, A.; Nakamichi, Y.; Yaoi, K. Identification and characterization of two xyloglucan-specific endo-1,4-glucanases in Aspergillus oryzae. Appl. Microbiol. Biotechnol. 2020, 104, 8761–8773. [Google Scholar] [CrossRef]
- Matsuzawa, T.; Yaoi, K. GH74 Xyloglucanases: Structures and Modes of Activity. Trends Glycosci. Glycotechnol. 2016, 28, E63–E70. [Google Scholar] [CrossRef] [Green Version]
- Arnal, G.; Stogios, P.J.; Asohan, J.; Attia, M.A.; Skarina, T.; Viborg, A.H.; Henrissat, B.; Savchenko, A.; Brumer, H. Substrate specificity, regiospecificity, and processivity in glycoside hydrolase family 74. J. Biol. Chem. 2019, 294, 13233–13247. [Google Scholar] [CrossRef] [Green Version]
- Gunnarsson, L.C.; Dexlin, L.; Karlsson, E.N.; Holst, O.; Ohlin, M. Evolution of a carbohydrate binding module into a protein-specific binder. Biomol. Eng. 2006, 23, 111–117. [Google Scholar] [CrossRef]
- Abbott, D.W.; van Bueren, A.L. Using structure to inform carbohydrate binding module function. Curr. Opin. Struct. Biol. 2014, 28, 32–40. [Google Scholar] [CrossRef]
- Boraston, A.B.; Bolam, D.N.; Gilbert, H.J.; Davies, G.J. Carbohydrate-binding modules: Fine-tuning polysaccharide recognition. Biochem. J. 2004, 382, 769–781. [Google Scholar] [CrossRef] [PubMed]
- Inoue, H.; Kishishita, S.; Kumagai, A.; Kataoka, M.; Fujii, T.; Ishikawa, K. Contribution of a family 1 carbohydrate-binding module in thermostable glycoside hydrolase 10 xylanase from Talaromyces cellulolyticus toward synergistic enzymatic hydrolysis of lignocellulose. Biotechnol. Biofuels 2015, 8, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernardes, A.; Pellegrini, V.O.A.; Curtolo, F.; Camilo, C.M.; Mello, B.L.; Johns, M.A.; Scott, J.L.; Guimaraes, F.E.C.; Polikarpov, I. Carbohydrate binding modules enhance cellulose enzymatic hydrolysis by increasing access of cellulases to the substrate. Carbohydr. Polym. 2019, 211, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Hori, C.; Gaskell, J.; Igarashi, K.; Samejima, M.; Hibbett, D.; Henrissat, B.; Cullen, D. Genomewide analysis of polysaccharides degrading enzymes in 11 white- and brown-rot Polyporales provides insight into mechanisms of wood decay. Mycologia 2013, 105, 1412–1427. [Google Scholar] [CrossRef] [Green Version]
- Borin, G.P.; Sanchez, C.C.; Souza, A.P.; Santana, E.S.; Souza, A.T.; Leme, A.F.P.; Squina, F.M.; Buckeridge, M.; Goldman, G.H.; Oliveira, J.V.C. Comparative secretome analysis of Trichoderma reesei and Aspergillus niger during growth on sugarcane biomass. PLoS ONE 2015, 10, e0129275. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, N.; Pearce, R.; Yi, S.; Zhao, X. Comparative secretomics analysis reveals the major components of Penicillium oxalicum 16 and Trichoderma reesei RUT-C30. Microorganisms 2021, 9, 2042. [Google Scholar] [CrossRef]
- Vélez-Mercado, M.I.; Talavera-Caro, A.G.; Escobedo-Uribe, K.M.; Sánchez-Muñoz, S.; Luévanos-Escareño, M.P.; Hernández-Terán, F.; Alvarado, A.; Balagurusamy, N. Bioconversion of lignocellulosic biomass into value added products under anaerobic conditions: Insight into proteomic studies. Int. J. Mol. Sci. 2021, 22, 12249. [Google Scholar] [CrossRef]
- Lopes, D.C.B.; Carraro, C.B.; Silva, R.N.; Paula, R.G. Molecular characterization of xyloglucanase cel74a from Trichoderma reesei. Int. J. Mol. Sci. 2021, 22, 4545. [Google Scholar] [CrossRef]
- Hu, J.G.; Arantes, V.; Pribowo, A.; Saddler, J.N. The synergistic action of accessory enzymes enhances the hydrolytic potential of a “cellulase mixture” but is highly substrate specific. Biotechnol. Biofuels 2013, 6, 112. [Google Scholar] [CrossRef] [Green Version]
- Benko, Z.; Siika-aho, M.; Viikari, L.; Reczey, K. Evaluation of the role of xyloglucanase in the enzymatic hydrolysis of lignocellulosic substrates. Enzym. Microb. Technol. 2008, 43, 109–114. [Google Scholar] [CrossRef]
- Hu, J.G.; Arantes, V.; Saddler, J.N. The enhancement of enzymatic hydrolysis of lignocellulosic substrates by the addition of accessory enzymes such as xylanase: Is it an additive or synergistic effect? Biotechnol. Biofuels 2011, 4, 36. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, F.L.; Azevedo, G.O.; Coelho, L.F.; Pagnocca, F.C.; Brienzo, M. Minimum lignin and xylan removal to improve cellulose accessibility. BioEnergy Res. 2020, 13, 775–785. [Google Scholar] [CrossRef]
- Novy, V.; Nielsen, F.; Cullen, D.; Sabat, G.; Houtman, C.J.; Hunt, C.G. The characteristics of insoluble softwood substrates affect fungal morphology, secretome composition, and hydrolytic efficiency of enzymes produced by Trichoderma reesei. Biotechnol. Biofuels 2021, 14, 105. [Google Scholar] [CrossRef]
- Freiesleben, P.; Spodsberg, N.; Stenbaek, A.; Stalbrand, H.; Krogh, K.; Meyer, A.S. Boosting of enzymatic softwood saccharification by fungal GH5 and GH26 endomannanases. Biotechnol. Biofuels 2018, 11, 194. [Google Scholar] [CrossRef]
- Berka, R.M.; Grigoriev, I.V.; Otillar, R.; Salamov, A.; Grimwood, J.; Reid, I.; Ishmael, N.; John, T.; Darmond, C.; Moisan, M.C.; et al. Comparative genomic analysis of the thermophilic biomass-degrading fungi Myceliophthora thermophila and Thielavia terrestris. Nat. Biotechnol. 2011, 29, 922–927. [Google Scholar] [CrossRef]
- Maheshwari, R.; Bharadwaj, G.; Bhat, M.K. Thermophilic fungi: Their physiology and enzymes. Microbiol. Mol. Biol. Rev. 2000, 64, 461–488. [Google Scholar] [CrossRef] [Green Version]
- Cereghino, J.L.; Cregg, J.M. Heterologous protein expression in the methylotrophic yeast Pichia pastoris. FEMS Microbiol. Rev. 2000, 24, 45–66. [Google Scholar] [CrossRef]
- Kristjansson, M.M.; Kinsella, J.E. Protein and enzyme stability: Structural, thermodynamic, and experimental aspects. Adv. Food Nutr. Res. 1991, 35, 237–316. [Google Scholar] [CrossRef]
- Voutilainen, S.P.; Nurmi-Rantala, S.; Penttila, M.; Aoivula, K. Engineering chimeric thermostable GH7 cellobiohydrolases in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2014, 98, 2991–3001. [Google Scholar] [CrossRef]
- Couturier, M.; Feliu, J.; Haon, M.; Navarro, D.; Lesage-Meessen, L.; Coutinho, P.M.; Berrin, J. A thermostable GH45 endoglucanase from yeast: Impact of its atypical multimodularity on activity. Microb. Cell Factories 2011, 10, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Ishida, T.; Yaoi, K.; Hiyoshi, A.; Igarashi, K.; Samejima, M. Substrate recognition by glycoside hydrolase family 74 xyloglucanase from the basidiomycete Phanerochaete chrysosporium. FEBS J. 2007, 274, 5727–5736. [Google Scholar] [CrossRef] [PubMed]
- Berezina, O.V.; Herlet, J.; Rykov, S.V.; Kornberger, P.; Zavyalov, A.; Kozlov, D.; Sakhibgaraeva, L.; Krestyanova, I.; Schwarz, W.H.; Zverlov, V.V.; et al. Thermostable multifunctional GH74 xyloglucanase from Myceliophthora thermophila: High-level expression in Pichia pastoris and characterization of the recombinant protein. Appl. Microbiol. Biotechnol. 2017, 101, 5653–5666. [Google Scholar] [CrossRef] [PubMed]
- Damasio, A.R.L.; Rubio, M.V.; Goncalves, T.A.; Persinoti, G.F.; Segato, F.; Prade, R.A.; Contesini, F.J.; Souza, A.P.; Buckeridge, M.S.; Squina, F.M. Xyloglucan breakdown by endo-xyloglucanase family 74 from Aspergillus fumigatus. Appl. Microbiol. Biotechnol. 2017, 101, 2893–2903. [Google Scholar] [CrossRef] [PubMed]
- Matsuzawa, T.; Saito, Y.; Yaoi, K. Key amino acid residues for the endo-processive activity of GH74 xyloglucanase. FEBS Lett. 2014, 588, 1731–1738. [Google Scholar] [CrossRef] [Green Version]
- Arnal, G.; Stogios, P.J.; Asohan, J.; Skarina, T.; Savchenko, A.; Hrumer, B. Structural enzymology reveals the molecular basis of substrate regiospecificity and processivity of an exemplar bacterial glycoside hydrolase family 74 endo-xyloglucanase. Biochem. J. 2018, 475, 3963–3978. [Google Scholar] [CrossRef] [Green Version]
- Sun, P.; Li, X.; Dilokpimol, A.; Henrissat, B.; Vries, R.P.; Kabel, M.A.; Makela, M.R. Fungal glycoside hydrolase family 44 xyloglucanases are restricted to the phylum basidiomycota and show a distinct xyloglucan cleavage pattern. iScience 2022, 25, 103666. [Google Scholar] [CrossRef]
- Lehtio, J.; Sugiyama, J.; Gustavsson, M.; Fransson, L.; Linder, M.; Teeri, T.T. The binding specificity and affinity determinants of family 1 and family 3 cellulose binding modules. Proc. Natl. Acad. Sci. USA 2003, 100, 484–489. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yang, F.; Hu, F.H.; Song, J.L.; Wu, S.F.; Jin, Y.C. Binding preference of family 1 carbohydrate binding module on nanocrystalline cellulose and nanofibrillar cellulose films assessed by quartz crystal microbalance. Cellulose 2018, 25, 3327–3337. [Google Scholar] [CrossRef]
- Kraulis, J.P.; Clore, G.M.; Nilges, M.; Jones, T.A.; Pettersson, G.; Knowles, J.; Gronenborn, A.M. Determination of the three-dimensional solution structure of the C-terminal domain of cellobiohydrolase 1 from Trichoderma reesei. A study using nuclear magnetic resonance and hybrid distance geometry-dynamical simulated annealing. Biochemistry 1989, 28, 7241–7257. [Google Scholar] [CrossRef]
- Dammak, A.; Quemener, B.; Bonnin, E.; Alvarado, C.; Bouchet, B.; Villares, A.; Moreau, C.; Cathala, B. Exploring architecture of xyloglucan cellulose nanocrystal complexes through enzyme susceptibility at different adsorption regimes. Biomacromolecules 2015, 16, 589–596. [Google Scholar] [CrossRef]
- Zheng, F.; Ding, S.J. Processivity and enzymatic mode of a glycoside hydrolase family 5 endoglucanase from Volvariella volvacea. Appl. Environ. Microbiol. 2013, 79, 989–996. [Google Scholar] [CrossRef] [Green Version]
- Kurasin, M.; Valjamae, P. Processivity of cellobiohydrolases is limited by the substrate. J. Biol. Chem. 2011, 286, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Nikolic, M.V.; Mojovic, L. Characterization and degradation of pectin derived from Budimka apple. J. Serb. Chem. Soc. 2008, 73, 157–167. [Google Scholar] [CrossRef]
- Jiang, K.K.; Li, L.L.; Long, L.K.; Ding, S.J. Comprehensive evaluation of combining hydrothermal pretreatment (autohydrolysis) with enzymatic hydrolysis for efficient release of monosaccharides and ferulic acid from corn bran. Ind. Crops Prod. 2018, 113, 348–357. [Google Scholar] [CrossRef]
- Gao, L.; He, X.; Guo, Y.; Wu, Z.; Zhao, J.; Liu, G.; Qu, Y. Combinatorial engineering of transcriptional activators in Penicillium oxalicum for improved production of corn-fiber-degrading enzymes. J. Agric. Food Chem. 2021, 69, 2539–2548. [Google Scholar] [CrossRef]
- Schmitz, E.; Leontakianakou, S.; Norlander, S.; Karlsson, E.N.; Adlercreutz, P. Lignocellulose degradation for the bioeconomy: The potential of enzyme synergies between xylanases, ferulic acid esterase and laccase for the production of arabinoxylo-oligosaccharides. Bioresour. Technol. 2022, 343, 126114. [Google Scholar] [CrossRef]
- Valério, R.; Serra, A.T.; Baixinho, J.; Cardeira, M.; Fernández, N.; Bronze, M.R.; Duarte, L.C.; Tavares, M.L.; Crespo, J.G.; Brazinha, C. Combined hydrothermal pre-treatment and enzymatic hydrolysis of corn fibre: Production of ferulic acid extracts and assessment of their antioxidant and antiproliferative properties. Ind. Crops Prod. 2021, 170, 113731. [Google Scholar] [CrossRef]
- Appeldoorn, M.M.; Kabel, M.A.; Eylen, D.V.; Gruppen, H.; Schols, H.A. Characterization of oligomeric xylan structures from corn fiber resistant to pretreatment and simultaneous saccharification and fermentation. J. Agric. Food Chem. 2010, 58, 11294–11301. [Google Scholar] [CrossRef]
- Skinner, R.C.; Gigliotti, J.C.; MKu, K.; Tou, J.C. A comprehensive analysis of the composition, health benefits, and safety of apple pomace. Nutr. Rev. 2018, 76, 893–909. [Google Scholar] [CrossRef]
- Curci, N.; Strazzulli, A.; Iacono, R.; Lise, F.D.; Maurelli, L.; Fenza, M.D.; Cobucci-Ponzano, B.; Moracci, M. Xyloglucan oligosaccharides hydrolysis by exo-acting glycoside hydrolases from hyperthermophilic microorganism Saccharolobus solfataricus. Int. J. Mol. Sci. 2021, 22, 3325. [Google Scholar] [CrossRef]
- Zhang, Y.H.P.; Cui, J.B.; Lynd, L.R.; Kuang, L.R. A transition from cellulose swelling to cellulose dissolution by o-phosphoric acid: Evidence from enzymatic hydrolysis and supramolecular structure. Biomacromolecules 2006, 7, 644–648. [Google Scholar] [CrossRef]
- Rose, D.J.; Inglett, G.E. Production of feruloylated arabinoxylo-oligosaccharides from maize (Zea mays) bran by microwave-assisted autohydrolysis. Food Chem. 2010, 119, 1613–1618. [Google Scholar] [CrossRef]
- Long, L.K.; Xu, M.J.; Shi, Y.X.; Lin, Q.Y.; Wang, J.; Ding, S.J. Characterization of two new endo-beta-1,4-xylanases from Eupenicillium parvum 4-14 and their applications for production of feruloylated oligosaccharides. Appl. Biochem. Biotechnol. 2018, 186, 816–833. [Google Scholar] [CrossRef]
- Wood, T.M.; Bhat, K.M. Methods for measuring cellulase activities. Methods Enzymol. 1988, 160, 87–112. [Google Scholar] [CrossRef]
- Furtado, G.P.; Lourenzoni, M.R.; Fuzo, C.A.; Fonseca-Maldonado, R.; Guazzaroni, M.E.; Ribeiro, L.F.; Ward, R.J. Engineering the affinity of a family 11 carbohydrate binding module to improve binding of branched over unbranched polysaccharides. Int. J. Biol. Macromol. 2018, 120, 2509–2516. [Google Scholar] [CrossRef]
- Chapla, D.; Pandit, P.; Shah, A. Production of xylooligosaccharides from corncob xylan by fungal xylanase and their utilization by probiotics. Bioresour. Technol. 2012, 115, 215–221. [Google Scholar] [CrossRef]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D.; Crocker, D. Determination of structural carbohydrates and lignin in biomass. Lab. Anal. Proced. 2008, 1617, 1–16. [Google Scholar]
- Chen, K.X.; Liu, X.C.; Long, L.K.; Ding, S.J. Cellobiose dehydrogenase from Volvariella volvacea and its effect on the saccharification of cellulose. Process. Biochem. 2017, 60, 52–58. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Glycosidic Bond | TtGH74 Activity |
|---|---|---|
| β-Glucan (barley) | β-1,3 and β-1,4 | ++ |
| Glucomannan (konjac) | β-1,4 | ++ |
| Lichenan (lichen of iceland) | β-1,3 and β-1,4 | + |
| Xyloglucan (tamarind seed) | β-1,4 α-1,6 and β-1,6 | +++++ |
| Laminarin (laminaria) | β-1,6-endo-β-1,3 | |
| Soluble starch (potato) | α-1,4 | |
| Xylan (birch) | β-1,4 | |
| Arabinoxylan (wheat) | β-1,4 | |
| Pectin | α-1,4 | |
| Chitin | β-1,4 | |
| CMC-Na | β-1,4 | |
| Avicel | β-1,4 | |
| PASC | β-1,4 | + |
| Entry Name | Strain | Temperature (°C) | pH | Vmax (U/mg) | Km (mg/mL) | Kcat (s−1) | References |
|---|---|---|---|---|---|---|---|
| TtGH74 | Thielavia terrestris | 75 | 5.5 | 193.2 | 0.3225 | 283.36 | This paper |
| TtGH74ΔCBM | 168.5 | 0.2671 | 233.09 | ||||
| PcGH74 | Phanerochaete chrysosporium | 55 | 6.0 | 0.25 | 28.1 | [41] | |
| PcGH74ΔCBM | 0.28 | 31.9 | |||||
| MtGH74 | Myceliophthora thermophila VKPM | 70–75 | 6.5 | 0.57 | [42] | ||
| AfGH74 | Aspergillus fumigatus | 50 | 5.5 | 11.9 | 1.5 | 16.4 | [43] |
| XEG74 | Paenibacillus sp KM21 | 45 | 6.0 | 36.8 | 0.96 | 49.2 | [44] |
| PoGH74 | Paenibacillus odorifer | 50 | 6.0 | 0.05 | 39.8 | [45] |
| Lignin | Glucose | Xylose | Galactose | Mannose | Arabinose | |
|---|---|---|---|---|---|---|
| Sulfuric-acid-pretreated corn bran | 20.73 ± 1.34% | 66.04 ± 0.41% | 8.10 ± 0.99% | 0.18 ± 0.07% | 1.82 ± 0.32% | 0.74 ± 0.17% |
| DES-pretreated corn bran | 11.10 ± 0.97% | 70.04 ± 0.65% | 7.78 ± 1.03% | 0.09 ± 0.02% | 0.98 ± 0.07% | 0.43 ± 0.09% |
| DES-pretreated apple pomace | 21.69 ± 1.83% | 43.22 ± 0.32% | 8.43 ± 1.32% | 2.59 ± 0.15% | 2.34 ± 0.18% | 0.22 ± 0.06% |
| Primer | Sequence (5′ to 3′) |
|---|---|
| 5′AOX | GACTGGTTCCAATTGACAAG |
| TtGH74ΔCBM-R | ATAGTTTAGCGGCCGCTTAGTGATGGTGATGGTGATGGTGAGATTGAGTAGCTTGAGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, B.; Chen, K.; Zhang, P.; Long, L.; Ding, S. Comparison of the Biochemical Properties and Roles in the Xyloglucan-Rich Biomass Degradation of a GH74 Xyloglucanase and Its CBM-Deleted Variant from Thielavia terrestris. Int. J. Mol. Sci. 2022, 23, 5276. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095276
Wang B, Chen K, Zhang P, Long L, Ding S. Comparison of the Biochemical Properties and Roles in the Xyloglucan-Rich Biomass Degradation of a GH74 Xyloglucanase and Its CBM-Deleted Variant from Thielavia terrestris. International Journal of Molecular Sciences. 2022; 23(9):5276. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095276
Chicago/Turabian StyleWang, Beibei, Kaixiang Chen, Peiyu Zhang, Liangkun Long, and Shaojun Ding. 2022. "Comparison of the Biochemical Properties and Roles in the Xyloglucan-Rich Biomass Degradation of a GH74 Xyloglucanase and Its CBM-Deleted Variant from Thielavia terrestris" International Journal of Molecular Sciences 23, no. 9: 5276. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095276