
Enrichment of Tumor-Infiltrating B Cells in Group 4 Medulloblastoma in Children

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Patient Cohort Clinical Data
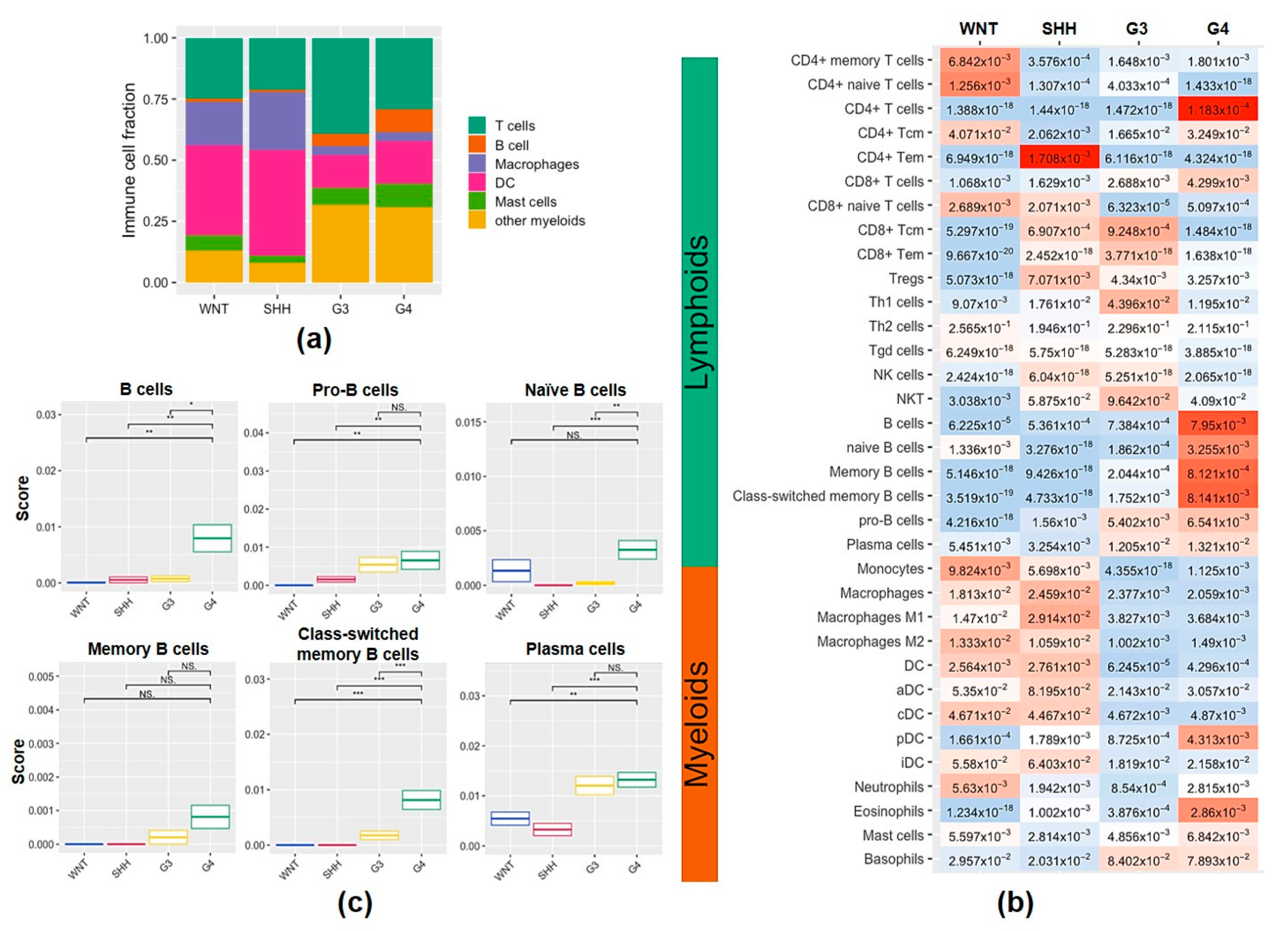
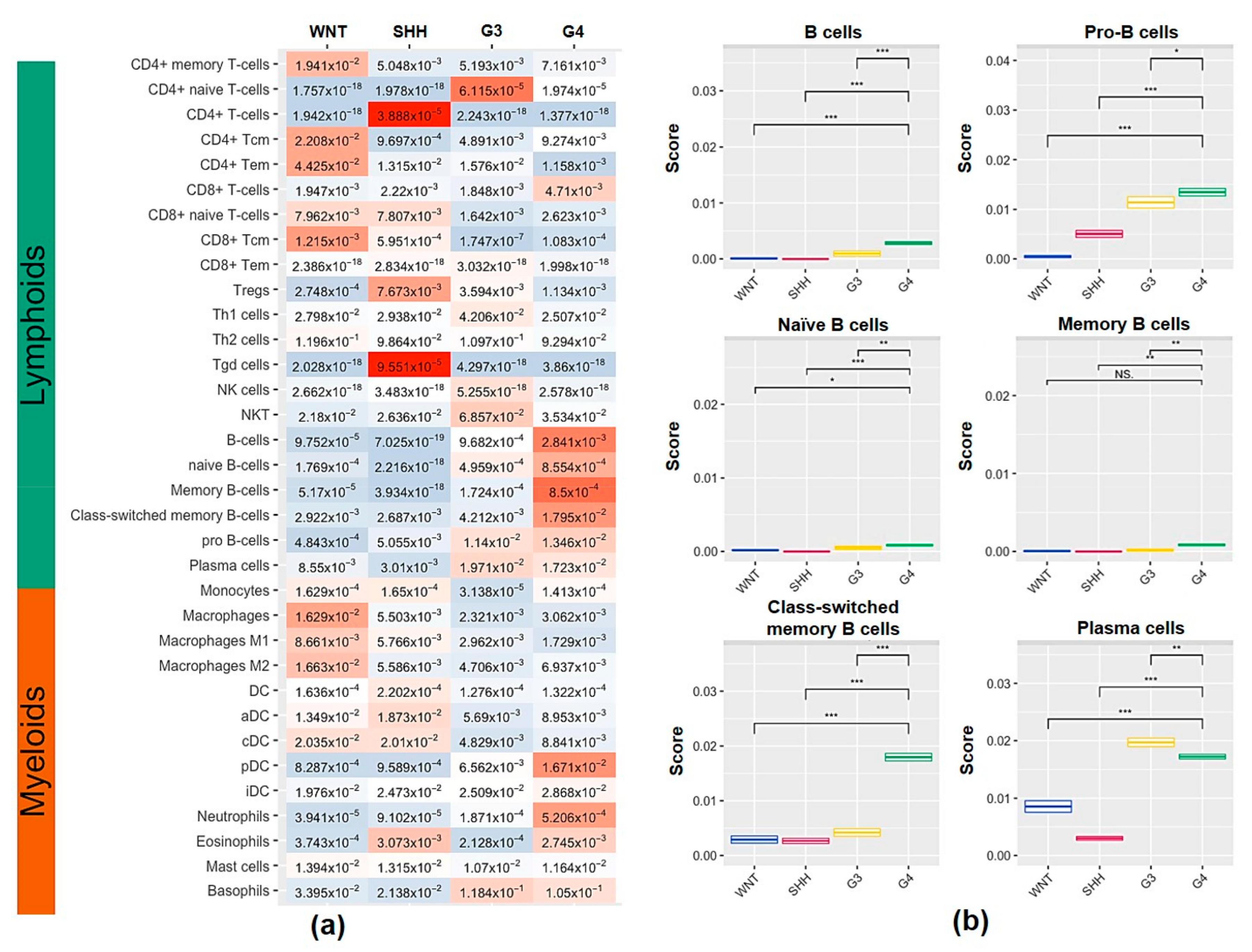
2.2. Characterization of Tumor-Infiltrating Immune Cell Population and Subtypes of Tumor-Infiltrating B Cells in the MB Subgroups
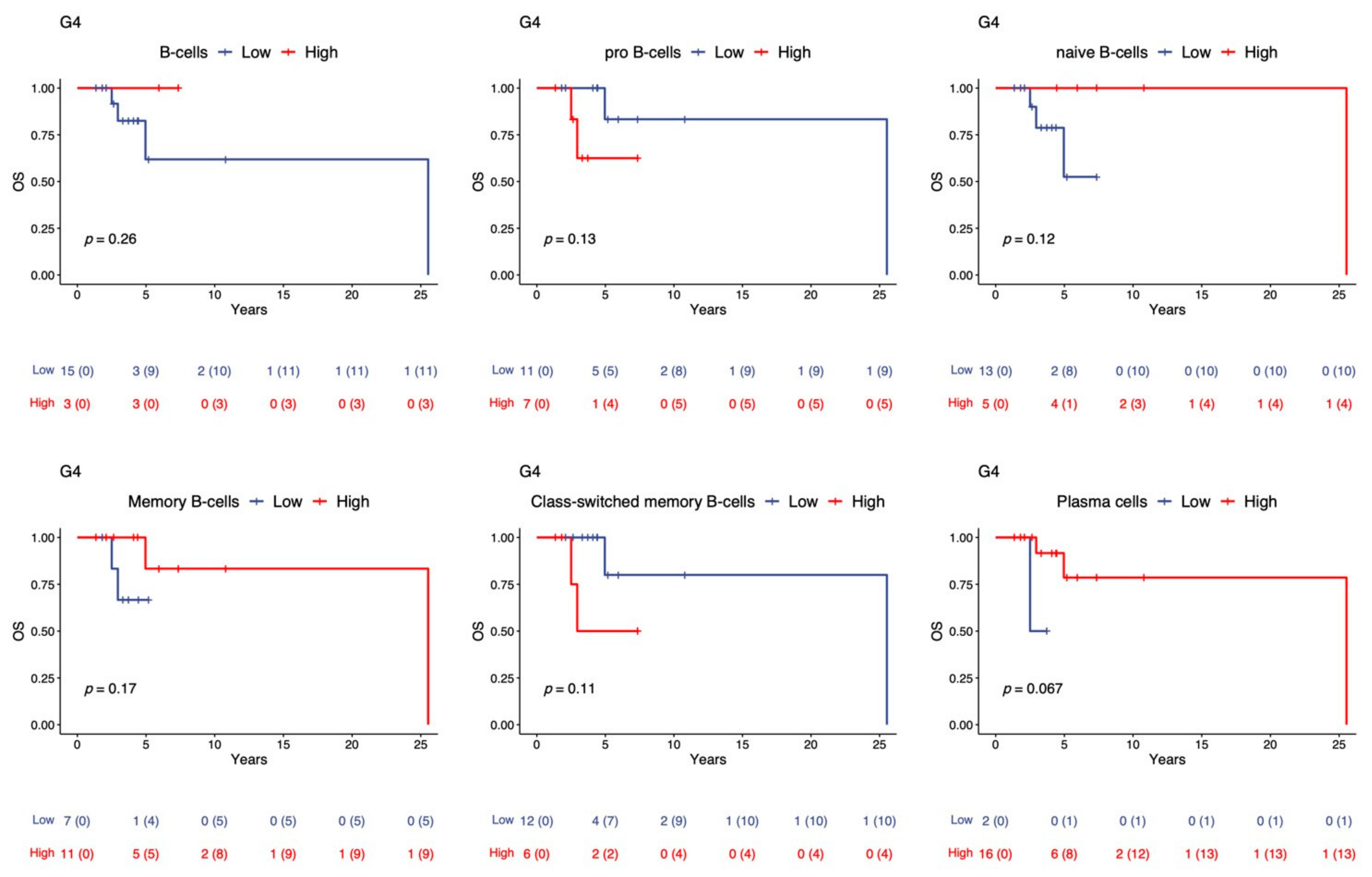
2.3. Overall Survival Analysis of TIL-B in the G4 Subgroups
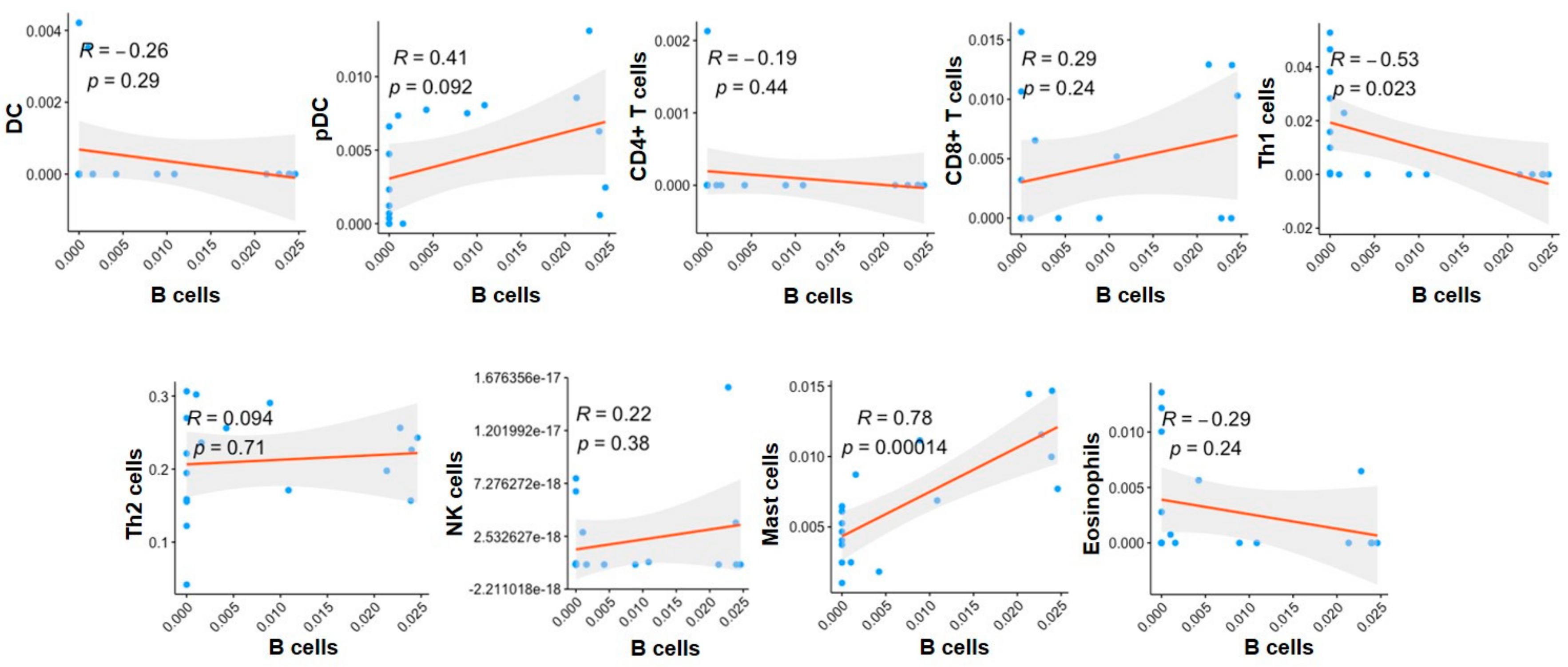
2.4. The Correlation of Immune Cells with B Cells in the G4 Subgroups
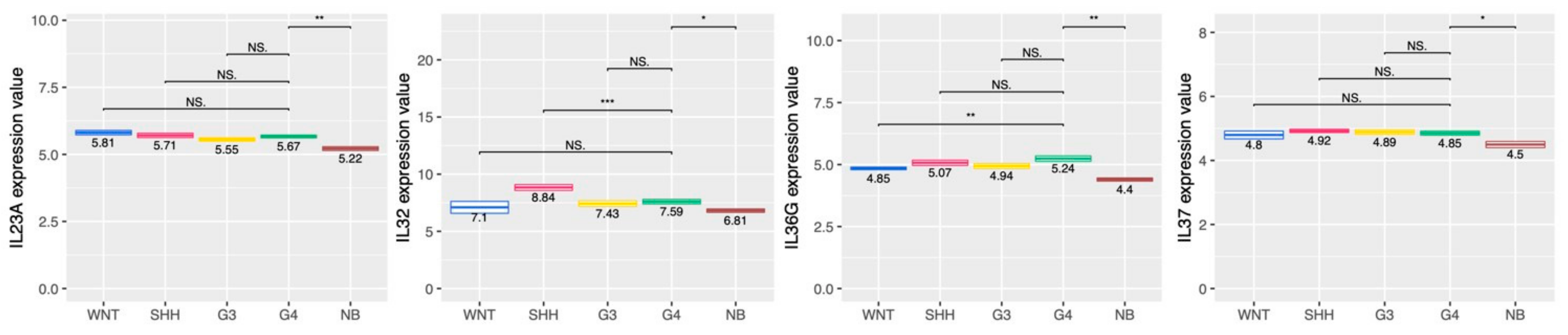
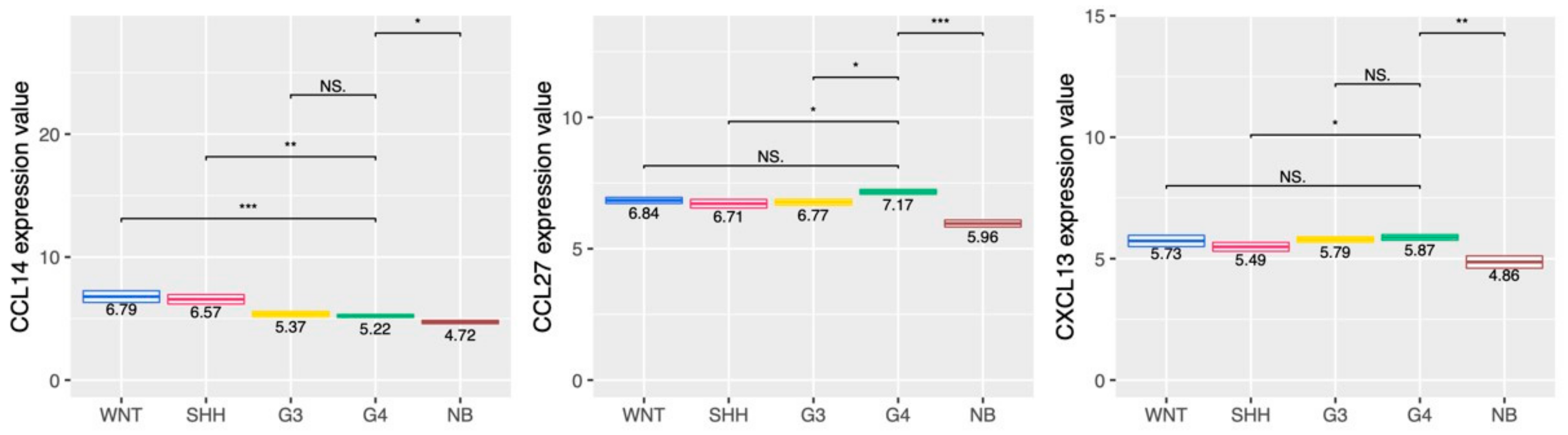
2.5. Cytokine and Chemokine Profiling in the MB Subgroups
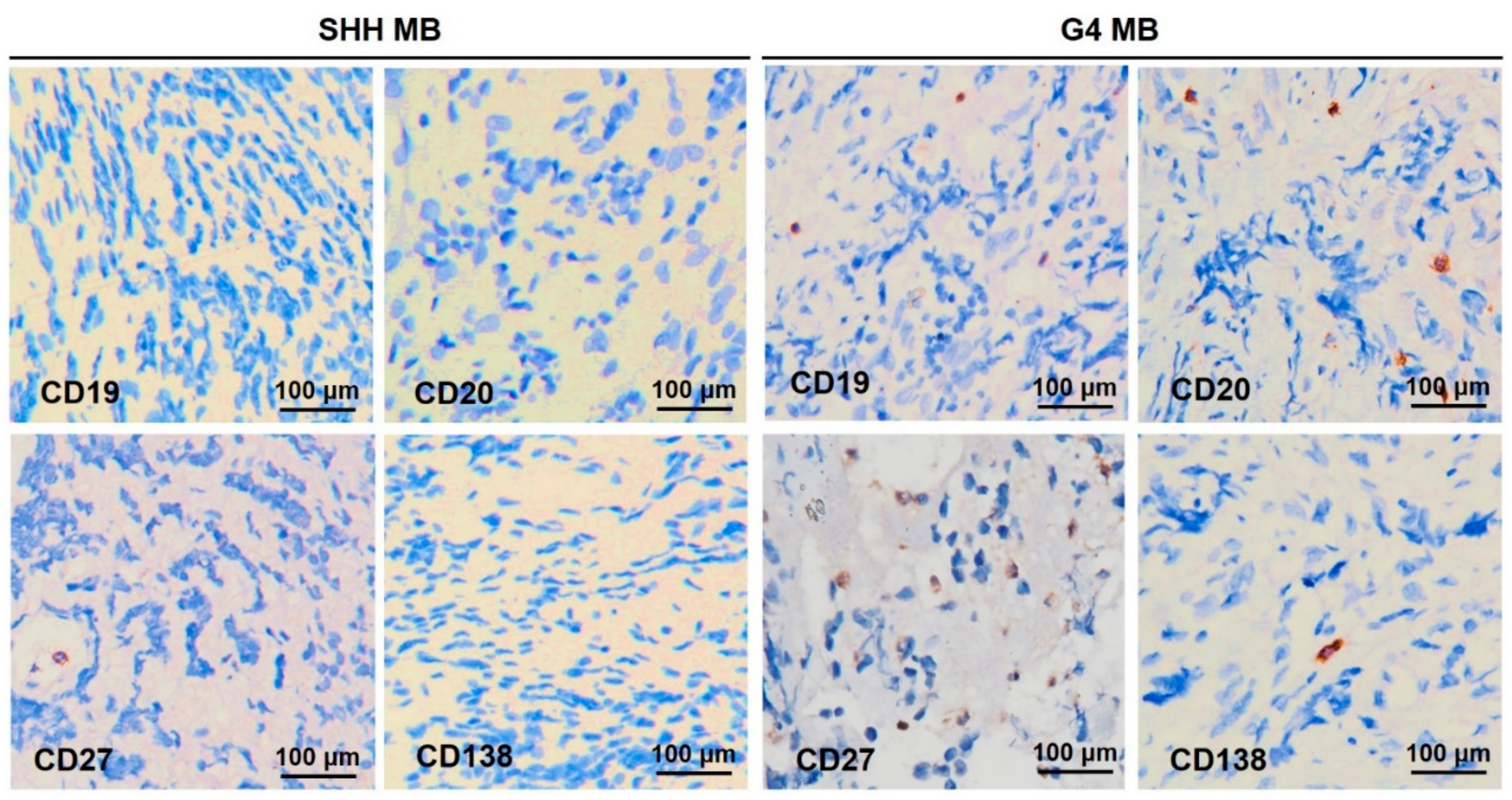
2.6. TIL-B Subsets in the SHH and G4 MB Subgroups
3. Discussion
4. Materials and Methods
4.1. Patient Cohort
4.2. Review of Clinical Data
4.3. Gene Expression Profiles and Subgroup Classification by RNA-Seq
4.4. Immune Cell Enrichment Analysis
4.5. DNA Methylation Array Profiling
4.6. Similarity Network Fusion Analysis for WNT and SHH Subtype Clustering
4.7. Random Forest Classifier for Non-WNT/Non-SHH Subtype Clustering
4.8. Immunohistochemistry (IHC) Analysis
4.9. Survival Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Louis, D.N.; Ohgaki, H.; Wiestler, O.D.; Cavenee, W.K.; Burger, P.C.; Jouvet, A.; Scheithauer, B.W.; Kleihues, P. The 2007 who classification of tumours of the central nervous system. Acta Neuropathol. 2007, 114, 97–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, T.T.; Liu, Y.L.; Ho, D.M.; Chang, K.P.; Liang, M.L.; Chen, H.H.; Lee, Y.Y.; Chang, F.C.; Lin, S.C.; Hsu, T.R.; et al. Factors affecting survival of medulloblastoma in children: The changing concept of management. Child’s Nerv. Syst. ChNS Off. J. Int. Soc. Pediatric Neurosurg. 2015, 31, 1687–1698. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.D.; Northcott, P.A.; Korshunov, A.; Remke, M.; Cho, Y.J.; Clifford, S.C.; Eberhart, C.G.; Parsons, D.W.; Rutkowski, S.; Gajjar, A.; et al. Molecular subgroups of medulloblastoma: The current consensus. Acta Neuropathol. 2012, 123, 465–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kool, M.; Korshunov, A.; Remke, M.; Jones, D.T.; Schlanstein, M.; Northcott, P.A.; Cho, Y.J.; Koster, J.; Schouten-van Meeteren, A.; van Vuurden, D.; et al. Molecular subgroups of medulloblastoma: An international meta-analysis of transcriptome, genetic aberrations, and clinical data of wnt, shh, group 3, and group 4 medulloblastomas. Acta Neuropathol. 2012, 123, 473–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Northcott, P.A.; Korshunov, A.; Witt, H.; Hielscher, T.; Eberhart, C.G.; Mack, S.; Bouffet, E.; Clifford, S.C.; Hawkins, C.E.; French, P.; et al. Medulloblastoma comprises four distinct molecular variants. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2011, 29, 1408–1414. [Google Scholar] [CrossRef] [PubMed]
- Zeltzer, P.M.; Boyett, J.M.; Finlay, J.L.; Albright, A.L.; Rorke, L.B.; Milstein, J.M.; Allen, J.C.; Stevens, K.R.; Stanley, P.; Li, H.; et al. Metastasis stage, adjuvant treatment, and residual tumor are prognostic factors for medulloblastoma in children: Conclusions from the children’s cancer group 921 randomized phase iii study. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 1999, 17, 832–845. [Google Scholar] [CrossRef]
- Ramaswamy, V.; Remke, M.; Bouffet, E.; Bailey, S.; Clifford, S.C.; Doz, F.; Kool, M.; Dufour, C.; Vassal, G.; Milde, T.; et al. Risk stratification of childhood medulloblastoma in the molecular era: The current consensus. Acta Neuropathol. 2016, 131, 821–831. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.S.; Ho, D.M.; Jou, S.T.; Yu, A.L.; Tran, H.M.; Liang, M.L.; Chen, H.H.; Lee, Y.Y.; Chen, Y.W.; Lin, S.C.; et al. Molecular-clinical correlation in pediatric medulloblastoma: A cohort series study of 52 cases in taiwan. Cancers 2020, 12, 653. [Google Scholar] [CrossRef] [Green Version]
- Zhukova, N.; Ramaswamy, V.; Remke, M.; Pfaff, E.; Shih, D.J.; Martin, D.C.; Castelo-Branco, P.; Baskin, B.; Ray, P.N.; Bouffet, E.; et al. Subgroup-specific prognostic implications of tp53 mutation in medulloblastoma. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2013, 31, 2927–2935. [Google Scholar] [CrossRef] [Green Version]
- Shih, D.J.; Northcott, P.A.; Remke, M.; Korshunov, A.; Ramaswamy, V.; Kool, M.; Luu, B.; Yao, Y.; Wang, X.; Dubuc, A.M.; et al. Cytogenetic prognostication within medulloblastoma subgroups. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2014, 32, 886–896. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.; Lee, J.; Choi, S.A.; Kim, S.K.; Wang, K.C.; Park, S.H.; Kim, S.H.; Lee, J.Y.; Phi, J.H. M1 macrophage recruitment correlates with worse outcome in shh medulloblastomas. BMC Cancer 2018, 18, 535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diao, S.; Gu, C.; Zhang, H.; Yu, C. Immune cell infiltration and cytokine secretion analysis reveal a non-inflammatory microenvironment of medulloblastoma. Oncol. Lett. 2020, 20, 397. [Google Scholar] [CrossRef] [PubMed]
- Low, S.Y.Y.; Bte Syed Sulaiman, N.; Tan, E.E.K.; Ng, L.P.; Kuick, C.H.; Chang, K.T.E.; Tang, P.H.; Wong, R.X.; Looi, W.S.; Low, D.C.Y.; et al. Cerebrospinal fluid cytokines in metastatic group 3 and 4 medulloblastoma. BMC Cancer 2020, 20, 554. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.M.; Nirschl, C.J.; Polanczyk, M.J.; Bell, W.R.; Nirschl, T.R.; Harris-Bookman, S.; Phallen, J.; Hicks, J.; Martinez, D.; Ogurtsova, A.; et al. Pd-l1 expression in medulloblastoma: An evaluation by subgroup. Oncotarget 2018, 9, 19177–19191. [Google Scholar] [CrossRef] [Green Version]
- Borst, J.; Ahrends, T.; Bąbała, N.; Melief, C.J.M.; Kastenmüller, W. Cd4+ t cell help in cancer immunology and immunotherapy. Nat. Rev. Immunol. 2018, 18, 635–647. [Google Scholar] [CrossRef]
- Bremnes, R.M.; Busund, L.T.; Kilvær, T.L.; Andersen, S.; Richardsen, E.; Paulsen, E.E.; Hald, S.; Khanehkenari, M.R.; Cooper, W.A.; Kao, S.C.; et al. The role of tumor-infiltrating lymphocytes in development, progression, and prognosis of non-small cell lung cancer. J. Thorac. Oncol. Off. Publ. Int. Assoc. Study Lung Cancer 2016, 11, 789–800. [Google Scholar] [CrossRef] [Green Version]
- Schalper, K.A.; Brown, J.; Carvajal-Hausdorf, D.; McLaughlin, J.; Velcheti, V.; Syrigos, K.N.; Herbst, R.S.; Rimm, D.L. Objective measurement and clinical significance of tils in non-small cell lung cancer. J. Natl. Cancer Inst. 2015, 107, dju435. [Google Scholar] [CrossRef] [Green Version]
- Bruno, T.C.; Ebner, P.J.; Moore, B.L.; Squalls, O.G.; Waugh, K.A.; Eruslanov, E.B.; Singhal, S.; Mitchell, J.D.; Franklin, W.A.; Merrick, D.T.; et al. Antigen-presenting intratumoral b cells affect cd4+ til phenotypes in non-small cell lung cancer patients. Cancer Immunol. Res. 2017, 5, 898–907. [Google Scholar] [CrossRef] [Green Version]
- Jones, H.P.; Wang, Y.C.; Aldridge, B.; Weiss, J.M. Lung and splenic b cells facilitate diverse effects on in vitro measures of antitumor immune responses. Cancer Immun. 2008, 8, 4. [Google Scholar]
- Shi, J.Y.; Gao, Q.; Wang, Z.C.; Zhou, J.; Wang, X.Y.; Min, Z.H.; Shi, Y.H.; Shi, G.M.; Ding, Z.B.; Ke, A.W.; et al. Margin-infiltrating cd20(+) b cells display an atypical memory phenotype and correlate with favorable prognosis in hepatocellular carcinoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2013, 19, 5994–6005. [Google Scholar] [CrossRef] [Green Version]
- Kroeger, D.R.; Milne, K.; Nelson, B.H. Tumor-infiltrating plasma cells are associated with tertiary lymphoid structures, cytolytic t-cell responses, and superior prognosis in ovarian cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2016, 22, 3005–3015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Germain, C.; Gnjatic, S.; Tamzalit, F.; Knockaert, S.; Remark, R.; Goc, J.; Lepelley, A.; Becht, E.; Katsahian, S.; Bizouard, G.; et al. Presence of b cells in tertiary lymphoid structures is associated with a protective immunity in patients with lung cancer. Am. J. Respir. Crit. Care Med. 2014, 189, 832–844. [Google Scholar] [CrossRef] [PubMed]
- Longerich, S.; Basu, U.; Alt, F.; Storb, U. Aid in somatic hypermutation and class switch recombination. Curr. Opin. Immunol. 2006, 18, 164–174. [Google Scholar] [CrossRef]
- De Visser, K.E.; Korets, L.V.; Coussens, L.M. De novo carcinogenesis promoted by chronic inflammation is b lymphocyte dependent. Cancer Cell 2005, 7, 411–423. [Google Scholar] [CrossRef] [Green Version]
- Ou, Z.; Wang, Y.; Liu, L.; Li, L.; Yeh, S.; Qi, L.; Chang, C. Tumor microenvironment b cells increase bladder cancer metastasis via modulation of the il-8/androgen receptor (ar)/mmps signals. Oncotarget 2015, 6, 26065–26078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.E.; Spata, M.; Bayne, L.J.; Buza, E.L.; Durham, A.C.; Allman, D.; Vonderheide, R.H.; Simon, M.C. Hif1a deletion reveals pro-neoplastic function of b cells in pancreatic neoplasia. Cancer Discov. 2016, 6, 256–269. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Jin, Y.; Tian, Y.; Zhang, H.; Wu, J.; Lu, W.; Lu, X. Regulatory b cells contribute to the impaired antitumor immunity in ovarian cancer patients. Tumour Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2016, 37, 6581–6588. [Google Scholar] [CrossRef]
- Xiao, X.; Lao, X.M.; Chen, M.M.; Liu, R.X.; Wei, Y.; Ouyang, F.Z.; Chen, D.P.; Zhao, X.Y.; Zhao, Q.; Li, X.F.; et al. Pd-1hi identifies a novel regulatory b-cell population in human hepatoma that promotes disease progression. Cancer Discov. 2016, 6, 546–559. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Gallastegui, N.; Rosenblatt, J.D. Regulatory b cells in anti-tumor immunity. Int. Immunol. 2015, 27, 521–530. [Google Scholar] [CrossRef]
- Olkhanud, P.B.; Damdinsuren, B.; Bodogai, M.; Gress, R.E.; Sen, R.; Wejksza, K.; Malchinkhuu, E.; Wersto, R.P.; Biragyn, A. Tumor-evoked regulatory b cells promote breast cancer metastasis by converting resting cd4⁺ t cells to t-regulatory cells. Cancer Res. 2011, 71, 3505–3515. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.W.; Yuan, X.L.; Chen, H.; Xie, G.H.; Ma, Y.H.; Zheng, Y.X.; Zhou, Y.L.; Shen, L.S. Cd19+ cd24hicd38hibregs involved in downregulate helper t cells and upregulate regulatory t cells in gastric cancer. Oncotarget 2015, 6, 33486–33499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Su, Y.X.; Lao, X.M.; Liang, Y.J.; Liao, G.Q. Cd19+il-10+ regulatory b cells affect survival of tongue squamous cell carcinoma patients and induce resting cd4+ t cells to cd4+foxp3+ regulatory t cells. Oral Oncol. 2016, 53, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.; Böhm, D.; von Törne, C.; Steiner, E.; Puhl, A.; Pilch, H.; Lehr, H.A.; Hengstler, J.G.; Kölbl, H.; Gehrmann, M. The humoral immune system has a key prognostic impact in node-negative breast cancer. Cancer Res. 2008, 68, 5405–5413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iglesia, M.D.; Parker, J.S.; Hoadley, K.A.; Serody, J.S.; Perou, C.M.; Vincent, B.G. Genomic analysis of immune cell infiltrates across 11 tumor types. J. Natl. Cancer Inst. 2016, 108, djw144. [Google Scholar] [CrossRef]
- Cabrita, R.; Lauss, M.; Sanna, A.; Donia, M.; Skaarup Larsen, M.; Mitra, S.; Johansson, I.; Phung, B.; Harbst, K.; Vallon-Christersson, J.; et al. Tertiary lymphoid structures improve immunotherapy and survival in melanoma. Nature 2020, 577, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Helmink, B.A.; Reddy, S.M.; Gao, J.; Zhang, S.; Basar, R.; Thakur, R.; Yizhak, K.; Sade-Feldman, M.; Blando, J.; Han, G.; et al. B cells and tertiary lymphoid structures promote immunotherapy response. Nature 2020, 577, 549–555. [Google Scholar] [CrossRef]
- Petitprez, F.; de Reyniès, A.; Keung, E.Z.; Chen, T.W.; Sun, C.M.; Calderaro, J.; Jeng, Y.M.; Hsiao, L.P.; Lacroix, L.; Bougoüin, A.; et al. B cells are associated with survival and immunotherapy response in sarcoma. Nature 2020, 577, 556–560. [Google Scholar] [CrossRef]
- Mahmoud, S.M.; Lee, A.H.; Paish, E.C.; Macmillan, R.D.; Ellis, I.O.; Green, A.R. The prognostic significance of b lymphocytes in invasive carcinoma of the breast. Breast Cancer Res. Treat. 2012, 132, 545–553. [Google Scholar] [CrossRef]
- Berntsson, J.; Nodin, B.; Eberhard, J.; Micke, P.; Jirström, K. Prognostic impact of tumour-infiltrating b cells and plasma cells in colorectal cancer. Int. J. Cancer 2016, 139, 1129–1139. [Google Scholar] [CrossRef]
- Edin, S.; Kaprio, T.; Hagström, J.; Larsson, P.; Mustonen, H.; Böckelman, C.; Strigård, K.; Gunnarsson, U.; Haglund, C.; Palmqvist, R. The prognostic importance of cd20+ b lymphocytes in colorectal cancer and the relation to other immune cell subsets. Sci. Rep. 2019, 9, 19997. [Google Scholar] [CrossRef]
- Garnelo, M.; Tan, A.; Her, Z.; Yeong, J.; Lim, C.J.; Chen, J.; Lim, K.H.; Weber, A.; Chow, P.; Chung, A.; et al. Interaction between tumour-infiltrating b cells and t cells controls the progression of hepatocellular carcinoma. Gut 2017, 66, 342–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Herpen, C.M.; van der Voort, R.; van der Laak, J.A.; Klasen, I.S.; de Graaf, A.O.; van Kempen, L.C.; de Vries, I.J.; Boer, T.D.; Dolstra, H.; Torensma, R.; et al. Intratumoral rhil-12 administration in head and neck squamous cell carcinoma patients induces b cell activation. Int. J. Cancer 2008, 123, 2354–2361. [Google Scholar] [CrossRef] [PubMed]
- Santoiemma, P.P.; Reyes, C.; Wang, L.P.; McLane, M.W.; Feldman, M.D.; Tanyi, J.L.; Powell, D.J., Jr. Systematic evaluation of multiple immune markers reveals prognostic factors in ovarian cancer. Gynecol. Oncol. 2016, 143, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Palma, A.M.; Hanes, M.R.; Marshall, J.S. Mast cell modulation of b cell responses: An under-appreciated partnership in host defence. Front. Immunol. 2021, 12, 718499. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.G.; McAlpine, S.M.; Huang, Y.Y.; Haidl, I.D.; Al-Afif, A.; Marshall, J.S.; Anderson, R. Rna sensors enable human mast cell anti-viral chemokine production and ifn-mediated protection in response to antibody-enhanced dengue virus infection. PLoS ONE 2012, 7, e34055. [Google Scholar] [CrossRef]
- Oldford, S.A.; Haidl, I.D.; Howatt, M.A.; Leiva, C.A.; Johnston, B.; Marshall, J.S. A critical role for mast cells and mast cell-derived il-6 in tlr2-mediated inhibition of tumor growth. J. Immunol. 2010, 185, 7067–7076. [Google Scholar] [CrossRef]
- Lichterman, J.N.; Reddy, S.M. Mast cells: A new frontier for cancer immunotherapy. Cells 2021, 10, 1270. [Google Scholar] [CrossRef]
- Gunn, M.D.; Ngo, V.N.; Ansel, K.M.; Ekland, E.H.; Cyster, J.G.; Williams, L.T. A b-cell-homing chemokine made in lymphoid follicles activates burkitt’s lymphoma receptor-1. Nature 1998, 391, 799–803. [Google Scholar] [CrossRef]
- Gu-Trantien, C.; Migliori, E.; Buisseret, L.; de Wind, A.; Brohée, S.; Garaud, S.; Noël, G.; Dang Chi, V.L.; Lodewyckx, J.N.; Naveaux, C.; et al. Cxcl13-producing tfh cells link immune suppression and adaptive memory in human breast cancer. JCI Insight 2017, 2, e91487. [Google Scholar] [CrossRef] [Green Version]
- Bansal, R.R.; Mackay, C.R.; Moser, B.; Eberl, M. Il-21 enhances the potential of human γδ t cells to provide b-cell help. Eur. J. Immunol. 2012, 42, 110–119. [Google Scholar] [CrossRef]
- Lian, L.H.; Milora, K.A.; Manupipatpong, K.K.; Jensen, L.E. The double-stranded rna analogue polyinosinic-polycytidylic acid induces keratinocyte pyroptosis and release of il-36γ. J. Investig. Dermatol. 2012, 132, 1346–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhao, X.; Feng, C.; Weinstein, A.; Xia, R.; Wen, W.; Lv, Q.; Zuo, S.; Tang, P.; Yang, X.; et al. Il-36γ transforms the tumor microenvironment and promotes type 1 lymphocyte-mediated antitumor immune responses. Cancer Cell 2015, 28, 296–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinstein, A.M.; Giraldo, N.A.; Petitprez, F.; Julie, C.; Lacroix, L.; Peschaud, F.; Emile, J.F.; Marisa, L.; Fridman, W.H.; Storkus, W.J.; et al. Association of il-36γ with tertiary lymphoid structures and inflammatory immune infiltrates in human colorectal cancer. Cancer Immunol. Immunother. CII 2019, 68, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Tong, Y.; Fang, H.; Tang, J.; Liu, L.; Hu, Y.; Li, J.; Zhong, L. Il36 indicating good prognosis in human hepatocellular carcinoma. J. Cancer 2020, 11, 6248–6255. [Google Scholar] [CrossRef]
- Morales, J.; Homey, B.; Vicari, A.P.; Hudak, S.; Oldham, E.; Hedrick, J.; Orozco, R.; Copeland, N.G.; Jenkins, N.A.; McEvoy, L.M.; et al. Ctack, a skin-associated chemokine that preferentially attracts skin-homing memory t cells. Proc. Natl. Acad. Sci. USA 1999, 96, 14470–14475. [Google Scholar] [CrossRef] [Green Version]
- Inokuma, D.; Abe, R.; Fujita, Y.; Sasaki, M.; Shibaki, A.; Nakamura, H.; McMillan, J.R.; Shimizu, T.; Shimizu, H. Ctack/ccl27 accelerates skin regeneration via accumulation of bone marrow-derived keratinocytes. Stem Cells 2006, 24, 2810–2816. [Google Scholar] [CrossRef]
- Kunkel, E.J.; Kim, C.H.; Lazarus, N.H.; Vierra, M.A.; Soler, D.; Bowman, E.P.; Butcher, E.C. Ccr10 expression is a common feature of circulating and mucosal epithelial tissue iga ab-secreting cells. J. Clin. Investig. 2003, 111, 1001–1010. [Google Scholar] [CrossRef]
- Keir, M.; Yi, Y.; Lu, T.; Ghilardi, N. The role of il-22 in intestinal health and disease. J. Exp. Med. 2020, 217, e20192195. [Google Scholar] [CrossRef]
- Dmitrieva-Posocco, O.; Dzutsev, A.; Posocco, D.F.; Hou, V.; Yuan, W.; Thovarai, V.; Mufazalov, I.A.; Gunzer, M.; Shilovskiy, I.P.; Khaitov, M.R.; et al. Cell-type-specific responses to interleukin-1 control microbial invasion and tumor-elicited inflammation in colorectal cancer. Immunity 2019, 50, 166–180.e167. [Google Scholar] [CrossRef] [Green Version]
- Barone, F.; Nayar, S.; Campos, J.; Cloake, T.; Withers, D.R.; Toellner, K.M.; Zhang, Y.; Fouser, L.; Fisher, B.; Bowman, S.; et al. Il-22 regulates lymphoid chemokine production and assembly of tertiary lymphoid organs. Proc. Natl. Acad. Sci. USA 2015, 112, 11024–11029. [Google Scholar] [CrossRef] [Green Version]
- Watford, W.T.; Moriguchi, M.; Morinobu, A.; O’Shea, J.J. The biology of il-12: Coordinating innate and adaptive immune responses. Cytokine Growth Factor Rev. 2003, 14, 361–368. [Google Scholar] [CrossRef]
- Robinson, D.; Shibuya, K.; Mui, A.; Zonin, F.; Murphy, E.; Sana, T.; Hartley, S.B.; Menon, S.; Kastelein, R.; Bazan, F.; et al. Igif does not drive th1 development but synergizes with il-12 for interferon-gamma production and activates irak and nfkappab. Immunity 1997, 7, 571–581. [Google Scholar] [CrossRef] [Green Version]
- Tominaga, K.; Yoshimoto, T.; Torigoe, K.; Kurimoto, M.; Matsui, K.; Hada, T.; Okamura, H.; Nakanishi, K. Il-12 synergizes with il-18 or il-1beta for ifn-gamma production from human t cells. Int. Immunol. 2000, 12, 151–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steding, C.E.; Wu, S.T.; Zhang, Y.; Jeng, M.H.; Elzey, B.D.; Kao, C. The role of interleukin-12 on modulating myeloid-derived suppressor cells, increasing overall survival and reducing metastasis. Immunology 2011, 133, 221–238. [Google Scholar] [CrossRef]
- Smith, K.M.; Pottage, L.; Thomas, E.R.; Leishman, A.J.; Doig, T.N.; Xu, D.; Liew, F.Y.; Garside, P. Th1 and th2 cd4+ t cells provide help for b cell clonal expansion and antibody synthesis in a similar manner in vivo. J. Immunol. 2000, 165, 3136–3144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.H.; Housepian, E.M.; Herbert, C., Jr. An operative staging system and a megavoltage radiotherapeutic technic for cerebellar medulloblastomas. Radiology 1969, 93, 1351–1359. [Google Scholar] [CrossRef]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic rna-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- Soneson, C.; Love, M.I.; Robinson, M.D. Differential analyses for rna-seq: Transcript-level estimates improve gene-level inferences. F1000Res 2015, 4, 1521. [Google Scholar] [CrossRef]
- Northcott, P.A.; Shih, D.J.; Remke, M.; Cho, Y.J.; Kool, M.; Hawkins, C.; Eberhart, C.G.; Dubuc, A.; Guettouche, T.; Cardentey, Y.; et al. Rapid, reliable, and reproducible molecular sub-grouping of clinical medulloblastoma samples. Acta Neuropathol. 2012, 123, 615–626. [Google Scholar] [CrossRef] [Green Version]
- Aran, D.; Hu, Z.; Butte, A.J. Xcell: Digitally portraying the tissue cellular heterogeneity landscape. Genome Biol. 2017, 18, 220. [Google Scholar] [CrossRef] [Green Version]
- Aryee, M.J.; Jaffe, A.E.; Corrada-Bravo, H.; Ladd-Acosta, C.; Feinberg, A.P.; Hansen, K.D.; Irizarry, R.A. Minfi: A flexible and comprehensive bioconductor package for the analysis of infinium DNA methylation microarrays. Bioinformatics 2014, 30, 1363–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Y.; Morris, T.J.; Webster, A.P.; Yang, Z.; Beck, S.; Feber, A.; Teschendorff, A.E. Champ: Updated methylation analysis pipeline for illumina beadchips. Bioinformatics 2017, 33, 3982–3984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez, S.; Garrido-Garcia, A.; Garcia-Gerique, L.; Lemos, I.; Sunol, M.; de Torres, C.; Kulis, M.; Perez-Jaume, S.; Carcaboso, A.M.; Luu, B.; et al. A novel method for rapid molecular subgrouping of medulloblastoma. Clin. Cancer Res. 2018, 24, 1355–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalli, F.M.G.; Remke, M.; Rampasek, L.; Peacock, J.; Shih, D.J.H.; Luu, B.; Garzia, L.; Torchia, J.; Nor, C.; Morrissy, A.S.; et al. Intertumoral heterogeneity within medulloblastoma subgroups. Cancer Cell 2017, 31, 737–754.e736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capper, D.; Jones, D.T.W.; Sill, M.; Hovestadt, V.; Schrimpf, D.; Sturm, D.; Koelsche, C.; Sahm, F.; Chavez, L.; Reuss, D.E.; et al. DNA methylation-based classification of central nervous system tumours. Nature 2018, 555, 469–474. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecular Subgroup Assignment | ||||
|---|---|---|---|---|
| n = 70 | WNT n = 8 (11.4%) | SHH n = 24 (34.3%) | Group 3 n = 20 (28.6%) | Group 4 n = 18 (25.7%) |
| Frozen tissue for molecular profiling | ||||
| Primary tumor | 8 (100%) | 23 (95.8%) | 17 (85.0%) | 17 (94.4%) |
| Recurrent tumor | 0 | 1 (4.2%) | 3 (15.0%) | 1 (5.6%) |
| Age (median, range) at diagnosis (years) | ||||
| 6.4 (0.3–18.3) | 7.9 (3.1–11.4) | 4.1 (0.3–14.3) | 5.1 (1.5–18.2) | 10.1 (5.1–18.3) |
| ≤3 y (n = 14, 20.0%) | 0 | 8 (33.3%) | 6 (30.0%) | 0 |
| >3 y (n = 56, 80.0%) | 8 (100%) | 16 (66.7%) | 14 (70.0%) | 18 (100%) |
| Sex | ||||
| Male, n = 35 (50.0%) | 1 (12.5%) | 12 (50.0%) | 11 (55.0%) | 11 (61.1%) |
| Female, n = 35 (50.0%) | 7 (87.5%) | 12 (50.0%) | 9 (45.0%) | 7 (38.9%) |
| Male/female ratio (1/1) | 0.1/1 | 1/1 | 1.2/1 | 1.6/1 |
| Metastasis stage at diagnosis (M0–1, M2–3), number of cases (percentage) | ||||
| M0–1, n = 50 (71.4%) | 8 (100%) | 20 (83.3%) | 10 (50.0%) | 12 (66.7%) |
| M2–3, n = 20 (28.6%) | 0 | 4 (16.7%) | 10 (50.0%) | 6 (33.3%) |
| Pathology variant, number of cases (percentage) and median age (years) | ||||
| Classic, n = 34 (48.6%), 6.1 | 5 (62.5%), | 12 (50.0%) | 9 (45.0%) | 8 (44.4%) |
| DNMB, n = 11 (15.7%), 3.5 | 1 (12.5%) | 9 (37.5%) | 0 | 1 (5.6%) |
| MBEN, n = 1 (1.4%), 1.0 | 0 | 1 (4.2%) | 0 | 0 |
| LCA, n = 23 (32.9%), 7.3 | 2 (25.0%) | 2 (8.3%) | 10 (50.0%) | 9 (50.0%) |
| MMMB, n = 1 (1.4%), 4.3 | 0 | 0 | 1 (5.0%) | 0 |
| Recurrence, number of cases (percentage) | ||||
| Recurrence, n = 27 (38.6%) | 1 (12.5%) | 11 (45.8%) | 11 (55.0%) | 4 (22.2%) |
| Median follow-up time (range) (years) | ||||
| 5.1 (0.3–25.5) | 4.9 (2.8–11.7) | 6.5 (0.7–17.7) | 6.7 (0.3–12.5). | 4.2 (1.4–25.5) |
| Survivals of molecular subgroup (percentage) | ||||
| 5-year OS rate: 74.1% | 100% | 74.1% | 62.4% | 73.8% |
| 5-year RFS rate: 63.0% | 87.5% | 62.5% | 45.0% | 68.5% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, K.-S.; Jian, T.-Y.; Sung, S.-Y.; Hsieh, C.-L.; Huang, M.-H.; Fang, C.-L.; Wong, T.-T.; Lin, Y.-L. Enrichment of Tumor-Infiltrating B Cells in Group 4 Medulloblastoma in Children. Int. J. Mol. Sci. 2022, 23, 5287. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095287
Wu K-S, Jian T-Y, Sung S-Y, Hsieh C-L, Huang M-H, Fang C-L, Wong T-T, Lin Y-L. Enrichment of Tumor-Infiltrating B Cells in Group 4 Medulloblastoma in Children. International Journal of Molecular Sciences. 2022; 23(9):5287. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095287
Chicago/Turabian StyleWu, Kuo-Sheng, Ting-Yan Jian, Shian-Ying Sung, Chia-Ling Hsieh, Man-Hsu Huang, Chia-Lang Fang, Tai-Tong Wong, and Yu-Ling Lin. 2022. "Enrichment of Tumor-Infiltrating B Cells in Group 4 Medulloblastoma in Children" International Journal of Molecular Sciences 23, no. 9: 5287. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095287