Zooxanthellate, Sclerite-Free, and Pseudopinnuled Octocoral Hadaka nudidomus gen. nov. et sp. nov. (Anthozoa, Octocorallia) from Mesophotic Reefs of the Southern Ryukyus Islands


Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimen Collection and Morphological Examinations
2.2. DNA Extraction, Amplification, and Sequencing
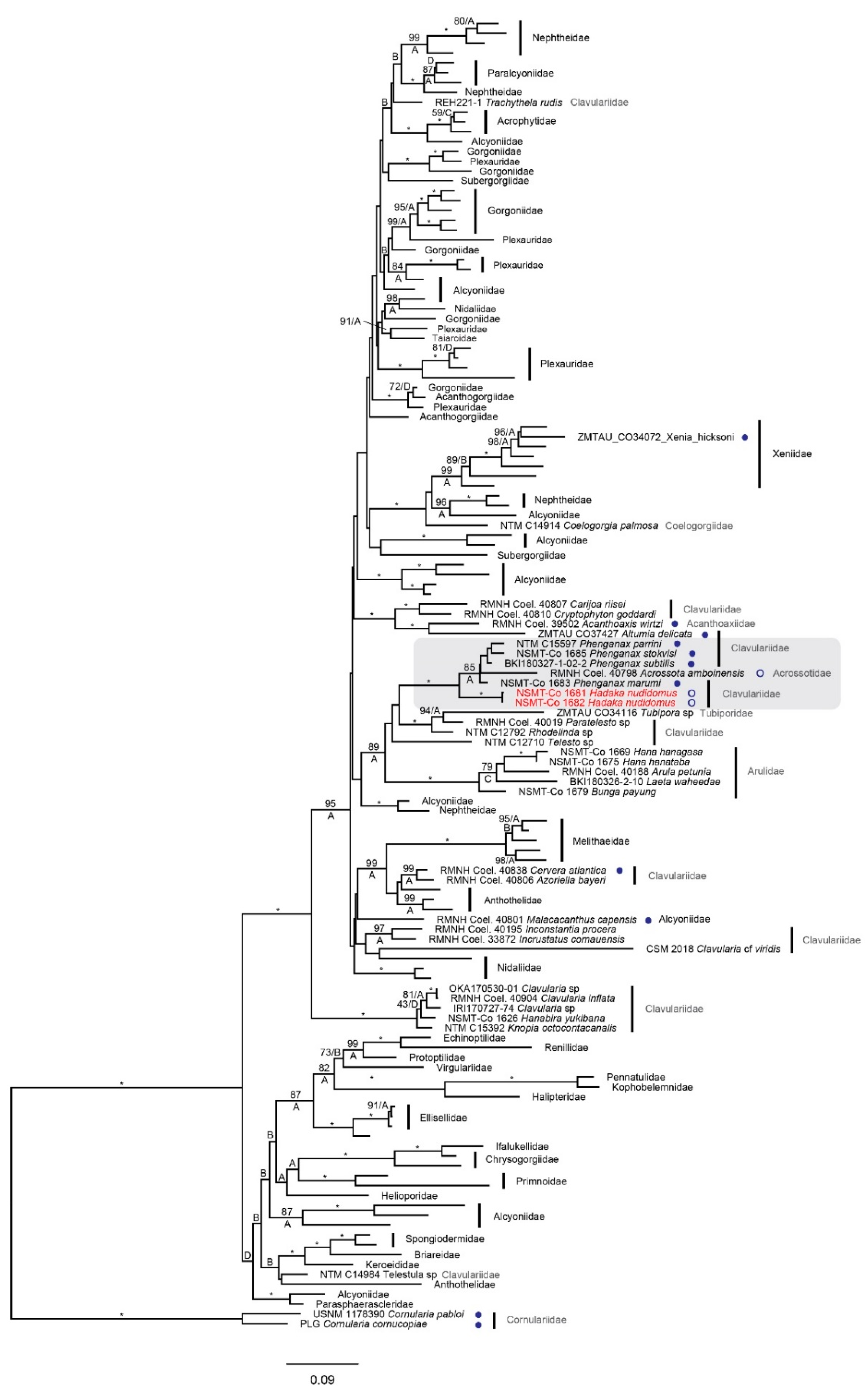
2.3. Molecular Phylogenetic Analyses
3. Systematic Account
3.1. Genus Hadaka gen. nov.
3.2. Hadaka nudidomus sp. nov.
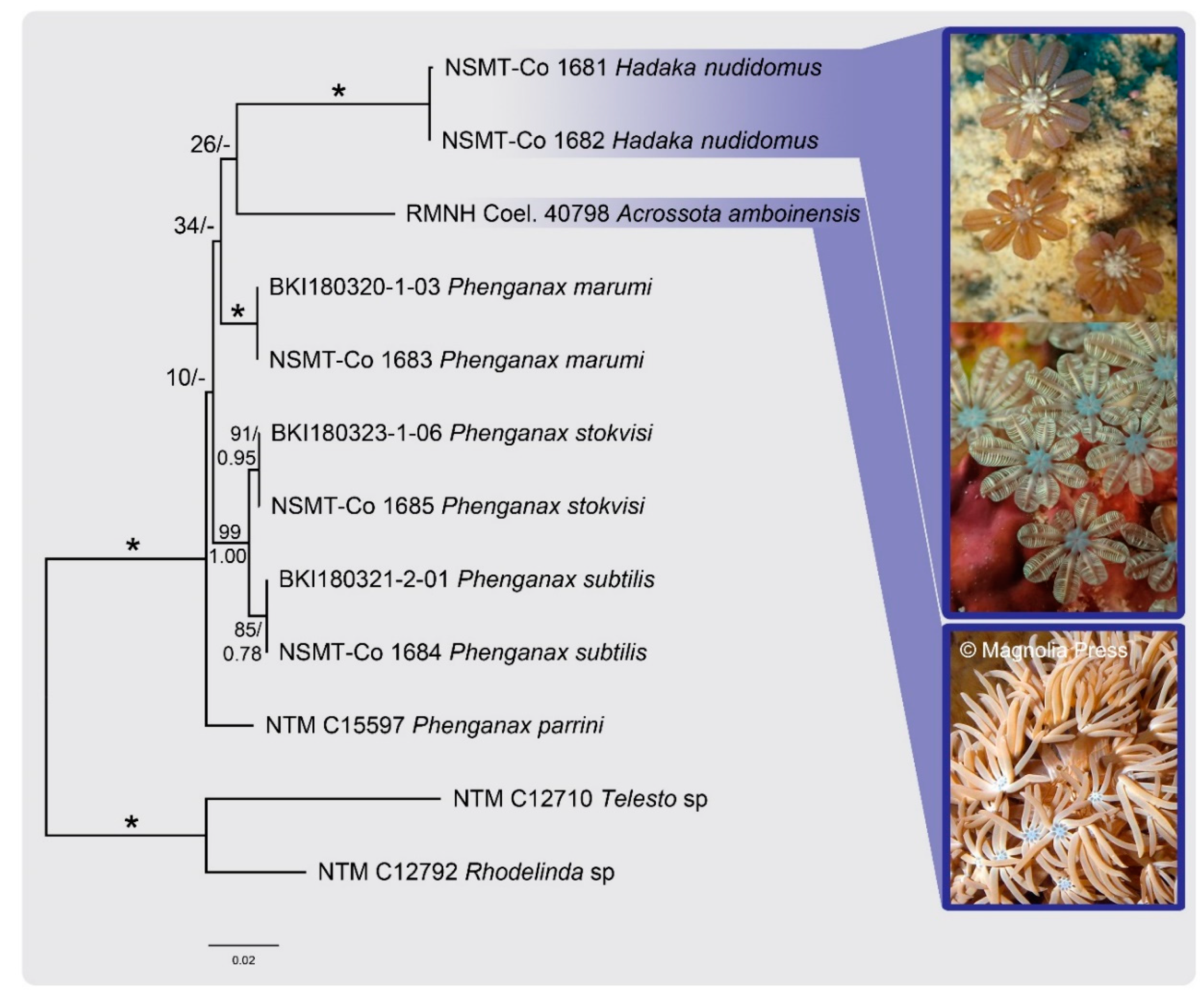
4. Molecular Results
5. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Buddemeier, R.W.; Kleypas, J.A.; Aronson, R.B. Coral Reefs and Global Climate Change: Potential Contributions of Climate Change to Stresses on Coral Reef Ecosystems, Report; Pew Centre on Global Climate Change: Arlington, VA, USA, 2004; 56p, Available online: http://www.reefresilience.org/pdf/Coral_Reefs_and_Global_Climate_Change.pdf (accessed on 2 April 2019).
- Chen, P.Y.; Chen, C.C.; Chu, L.; McCarl, B. Evaluating the economic damage of climate change. Glob. Environ. Chang. 2015, 30, 12–20. [Google Scholar] [CrossRef]
- Hughes, T.P.; Baird, A.H.; Bellwood, D.R.; Card, M.; Connolly, S.R.; Folke, C.; Grosberg, R.; Hoegh-Guldberg, O.; Jackson, J.B.C.; Kleypas, J.; et al. Climate change, human impacts, and the resilience of coral reefs. Science 2003, 301, 929–933. [Google Scholar] [CrossRef] [PubMed]
- Pandolfi, J.M.; Bradbury, R.H.; Sala, E.; Hughes, T.P.; Bjorndal, K.A.; Cooke, R.G.; McArdle, D.; McClenachan, L.; Newman, M.J.H.; Paredes, G.; et al. Global trajectories of the long-term decline of coral reef ecosystems. Science 2003, 301, 955–958. [Google Scholar] [CrossRef] [PubMed]
- Meyer, C.P.; Geller, J.B.; Paulay, G. Fine scale endemism on coral reefs: Archipelagic differentiation in turbinid gastropods. Evolution 2005, 59, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Kerry, J.T.; Baird, A.H.; Connolly, S.R.; Dietzel, A.; Eakin, C.A.; Heron, S.F.; Hoey, A.S.; Hoogenboom, M.O.; Liu, G.; et al. Global warming transforms coral reef assemblages. Nature 2018, 556, 492–496. [Google Scholar] [CrossRef] [PubMed]
- Mittermeier, R.A.; Myers, N.; Thomsen, J.B.; da Fonseca, G.A.B.; Olivieri, S. Biodiversity hotspots and major tropical wilderness areas: Approaches to setting conservation priorities. Conserv. Biol. 1998, 12, 516–520. [Google Scholar] [CrossRef]
- Mittermeier, R.A.; Myers, N.; Mittermeier, C.G. Hotspots—Earth’s Biologically Richest and Most Endangered Terrestrial Ecoregions; Cemex: Mexico City, Mexico, 1999; 431p. [Google Scholar]
- Allen, G.R. Conservation hotspots of biodiversity and endemism for Indo-Pacific coral reef fishes. Aquat. Conserv. 2008, 18, 541–556. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Meyer, C.P. Searching for heat in a marine biodiversity hotspot. J. Biogeogr. 2009, 36, 569–576. [Google Scholar] [CrossRef]
- Appeltans, W.; Ahyong, S.T.; Anderson, G.; Angel, M.V.; Artois, T.; Bailly, N.; Bamber, R.; Barber, A.; Bartsch, I.; Berta, A.; et al. The magnitude of global marine species diversity. Curr. Biol. 2012, 22, 2189–2202. [Google Scholar] [CrossRef]
- Troudet, J.; Grandcolas, P.; Blin, A.; Vignes-Lebbe, R.; Legendre, F. Taxonomic bias in biodiversity data and societal preferences. Sci. Rep. 2017, 7, 9132. [Google Scholar] [CrossRef]
- Mace, G.M.; Balmford, A.; Boitani, L.; Cowlishaw, G.; Dobson, A.P.; Faith, D.P. It’s time to work together and stop duplicating conservation efforts. Nature 2000, 405, 393. [Google Scholar] [CrossRef] [PubMed]
- Hooper, J.N.A.; Kennedy, J.A.; Quinn, R.J. Biodiversity ‘hotspots’, patterns of richness and endemism, and taxonomic affinities of tropical Australian sponges (Porifera). Biodivers. Conserv. 2002, 11, 851–885. [Google Scholar] [CrossRef]
- Kerr, J.T. Species richness, endemism, and choice of areas for conservation. Conserv. Biol. 1997, 11, 1094–1100. [Google Scholar] [CrossRef]
- Puglise, K.A.; Hinderstein, L.M.; Marr, J.C.A.; Dowgiallo, M.J.; Martinez, F.A. Mesophotic Coral Ecosystems Research Strategy: International Workshop to Prioritize Research and Management Needs for Mesophotic Coral Ecosystems, Jupiter, Florida, 12–15 July 2008; NOAA Technical Memorandum NOS NCCOS 98 and OAR OER 2; NOAA National Centers for Coastal Research, NOAA Undersea Research Program: Silver Spring, MD, USA, 2009; 24p.
- Kahng, S.E.; Garcia-Sais, J.R.; Spalding, H.L.; Brokovich, E.; Wagner, D.; Weil, E.; Hinderstein, L.; Toonen, R.J. Community ecology of mesophotic coral reef ecosystems. Coral Reefs 2010, 29, 255–275. [Google Scholar] [CrossRef]
- Sinniger, F.; Harii, S. Studies on Mesophotic Coral Ecosystems in Japan. In Coral Reef Studies of Japan. Coral Reefs of the World; Iguchi, A., Hongo, C., Eds.; Springer: Singapore, 2018; Volume 13, pp. 149–162. [Google Scholar]
- Laverick, J.H.; Piango, S.; Andradi-Brown, D.A.; Exton, D.A.; Bongaerts, P.; Bridge, T.C.; Lesser, M.P.; Pyle, R.L.; Slattery, M.; Wagner, D.; et al. To what extent do mesophotic coral ecosystems and shallow reefs share species of conservation interest? A systematic review. Environ. Evid. 2018, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- Pyle, R.L. Ocean Pulse: A Critical Diagnosis; Tanacredi, J.T., Loret, J., Eds.; Springer: Boston, MA, USA, 1998; pp. 71–88. [Google Scholar]
- Rocha, L.A.; Pinheiro, H.T.; Shepherd, B.; Papastamatiou, Y.P.; Luiz, O.J.; Pyle, R.L.; Bongaerts, P. Mesophotic coral ecosystems are threatened and ecologically distinct from shallow water reefs. Science 2018, 361, 281–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosaki, R.K.; Pyle, R.L.; Leonard, J.C.; Hauk, B.B.; Whitton, R.K.; Wagner, D. 100% endemism in mesophotic reef fish assemblages at Kure Atoll, Hawaiian Islands. Mar. Biodivers. 2017, 47, 783–784. [Google Scholar] [CrossRef]
- Roberts, C.M.; McClean, C.J.; Veron, J.E.N.; Hawkins, J.P.; Allen, J.R.; McAllister, D.E.; Mittermeier, C.G.; Schueler, F.W.; Spalding, M.; Wells, F.; et al. Marine biodiversity hotspots and conservation priorities for tropical reefs. Science 2002, 295, 1280–1285. [Google Scholar] [CrossRef]
- Kizaki, K. Geology and tectonics of the Ryukyu Islands. Tectonophysics 1986, 125, 193–207. [Google Scholar] [CrossRef]
- Reimer, J.D.; Biondi, P.; Lau, Y.W.; Masucci, G.; Nguyen, X.H.; Santos, M.E.A.; Wee, H.B. Marine biodiversity research in the Ryukyu Islands, Japan: Current status and trends. PeerJ 2019, 7, e6532. [Google Scholar] [CrossRef]
- Andres, M.; Park, J.H.; Wimbush, M.; Zhu, X.H.; Chang, K.I.; Ichikawa, H. Study of the Kuroshio/Ryukyu current system based on satellite-altimeter and in situ measurements. J. Oceanogr. 2008, 64, 937–950. [Google Scholar] [CrossRef]
- Veron, J.E.N. Conservation of biodiversity: A critical time for the hermatypic corals of Japan. Coral Reefs 1992, 11, 13–21. [Google Scholar] [CrossRef]
- Rodriguez-Lanetty, M.; Hoegh-Guldberg, O. Symbiont diversity within the widespread scleractinian coral Plesiastrea versipora, across the northwestern Pacific. Mar. Biol. 2003, 143, 501–509. [Google Scholar] [CrossRef]
- Fabricius, K.; Alderslade, P. Soft Corals and Sea Fans: A Comprehensive Guide to the Tropical Shallow-Water Genera of the Central-West Pacific, the Indian Ocean and the Red Sea; Australian Institute of Marine Science: Townsville, Australia, 2001. [Google Scholar]
- Daly, M.; Brugler, M.R.; Cartwright, P.; Collins, A.G.; Dawson, M.N.; Fautin, D.G.; France, S.C.; McFadden, C.S.; Opresko, D.M.; Rodriguez, E.; et al. The phylum Cnidaria: A review of phylogenetic patterns and diversity 300 years after Linnaeus. Zootaxa 2007, 1668, 127–182. [Google Scholar]
- McFadden, C.S.; van Ofwegen, L.P. Stoloniferous octocorals (Anthozoa, Octocorallia) from South Africa, with descriptions of a new family of Alcyonacea, a new genus of Clavulariidae, and a new species of Cornularia (Cornulariidae). Invertebr. Syst. 2012, 26, 331–356. [Google Scholar] [CrossRef]
- Lau, Y.W.; Stokvis, F.R.; Van Ofwegen, L.P.; Reimer, J.D. Stolonifera from shallow waters in the north-western Pacific: A description of a new genus and two new species within the Arulidae (Anthozoa, Octocorallia). ZooKeys 2018, 790, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Lau, Y.W.; Reimer, J.D. A first phylogenetic study on stoloniferous octocorals off the coast of Kota Kinabalu, Sabah, Malaysia, and the description of three new genera and six new species. ZooKeys 2019, 872, 127–158. [Google Scholar] [CrossRef]
- Lau, Y.W.; Stokvis, F.R.; Imahara, Y.; Reimer, J.D. The stoloniferous octocoral, Hanabira yukibana, gen. nov., sp. nov., of the southern Ryukyus has morphological and symbiont variation. Contrib. Zool. 2019, 54–77. [Google Scholar] [CrossRef]
- Van de Water, J.A.J.M.; Allemand, D.; Ferrier-Pagès, C. Host-microbe interactions in octocoral holobionts-recent advances and perspectives. Microbiome 2018, 6, 64. [Google Scholar] [CrossRef]
- Van Oppen, M.J.H.; Mieog, J.C.; Sánchez, C.A.; Fabricius, K.E. Diversity of alga symbionts (zooxanthellae) in octocorals: The roles of geography and host relationships. Mol. Ecol. 2005, 14, 2403–2417. [Google Scholar] [CrossRef]
- Goulet, T.L.; Simmons, C.; Goulet, D. Worldwide biogeography of Symbiodinium in tropical octocorals. Mar. Ecol. Prog. Ser. 2008, 355, 45–58. [Google Scholar] [CrossRef]
- LaJeunesse, T.C.; Bhagooli, R.; Hidaka, M.; de Vantier, L.; Done, T.; Schmidt, G.W.; Fitt, W.K.; Hoegh-Guldberg, O. Closely related Symbiodinium spp. differ in relative dominance in coral reef host communities across environmental, latitudinal and biogeographic gradients. Mar. Ecol. Prog. Ser. 2004, 284, 147–161. [Google Scholar] [CrossRef]
- Fabricius, K.E.; De’ath, G. Photosynthetic symbionts and energy supply determine octocoral biodiversity in coral reefs. Ecology 2008, 89, 3163–3173. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, M.; Wilson, A.; Stones-havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Ranwez, V.; Harispe, S.; Delsuc, F.; Douzery, E.J.P. MACSE: Multiple Alignment of Coding SEquences accounting for frameshifts and stop codons. PLoS ONE 2011, 6, e22594. [Google Scholar] [CrossRef]
- Gaurav, V.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analyses and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Aberer, A.J.; Kobert, K.; Stamatakis, A. ExaBayes: Massively parallel Bayesian tree inference for the whole-genome era. Mol. Biol. Evol. 2014, 31, 2553–2556. [Google Scholar] [CrossRef] [PubMed]
- Foster, P.G. Modeling compositional heterogeneity. Syst. Biol. 2014, 53, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Rambout, A. FigTree. 2014. Available online: tree.bio.ed.ac.uk/software/figtree/ (accessed on 17 April 2019).
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- McFadden, C.S.; Benayahu, Y.; Pante, E.; Thoma, J.N.; Nevarez, P.A.; France, S.C. Limitations of mitochondrial gene barcoding in Octocorallia. Mol. Ecol. Resour. 2011, 11, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Alderslade, P.; McFadden, C.S. Pinnule-less polyps: A new genus and new species of Indo-Pacific Clavulariidae and validation of the soft coral genus Acrossota and the family Acrossotidae (Coelenterata: Octocorallia). Zootaxa 2007, 1400, 27–44. [Google Scholar] [CrossRef]
- LaJeunesse, T.C.; Parkinson, J.E.; Gabrielson, P.W.; Jeong, H.J.; Reimer, J.D.; Voolstra, C.R.; Santos, S.R. Systematic revision of Symbiodiniaceae highlights the antiquity and diversity of coral endosymbionts. Curr. Biol. 2018, 28. [Google Scholar] [CrossRef] [PubMed]
- Benayahu, Y.; McFadden, C.S.; Shoham, E.; Van Ofwegen, L.P. Search for mesophotic octocorals (Cnidaria, Anthozoa) and their phylogeny: II. A new zooxanthellate species from Eilat, northern Red Sea. ZooKeys 2017, 676. [Google Scholar] [CrossRef]
- LaJeunesse, T.C.; Thornhill, D.J. Improved resolution of reef-coral endosymbiont (Symbiodinium) species diversity, ecology, and evolution through psbA non-coding region genotyping. PLoS ONE 2011, 6, e29013. [Google Scholar] [CrossRef]
- Alderslade, P.; McFadden, C.S. A new sclerite-free genus and species of Clavulariidae (Coelenterata: Octocorallia). Zootaxa 2011, 3104, 64–68. [Google Scholar] [CrossRef] [Green Version]
- Benayahu, Y.; McFadden, C.S.; Shoham, E. Search for mesophotic octocorals (Cnidaria, Anthozoa) and their phylogeny: I. A new sclerite-free genus from Eilat, northern Red Sea. ZooKeys 2017, 680. [Google Scholar] [CrossRef]
- Baldwin, C.C.; Pitassy, D.E.; Robertson, D.R. A new deep-reef scorpionfish (Teleostei, Scorpaenidae, Scorpaenodes) from the southern Caribbean with comments on depth distributions and relationships of western Atlantic members of the genus. ZooKeys 2016, 606, 141–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loya, Y.; Eyal, G.; Treibitz, T.; Lesser, M.P.; Appeldoorn, R. Theme section on mesophotic coral ecosystems: Advances in knowledge and future perspectives. Coral Reefs 2016, 35. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Species | Catalogue Number | Locality/GPS (DMS) | Symbiodiniaceae Genus | GenBank Accession Numbers | ||||
|---|---|---|---|---|---|---|---|---|---|
| 28S rDNA | COI | mtMutS | ND6 | ITS | |||||
| Clavulariidae | Hadaka nudidomus gen. nov. et sp. nov. | NSMT-Co 1681 (holotype) | NE Uchibanare, Iriomote Isl./24.370413 N, 123.736428 E | Cladocopium | MN488601 | MN488603 | MN488605 | n.a. | MN488607 |
| Hadaka nudidomus gen. nov. et sp. nov. | NSMT-Co 1682 (paratype) | Entrance Hedo Dome, Cape Hedo, Okinawa Isl./26.856412 N, 128.245093 E | Cladocopium | MN488602 | MN488604 | n.a. | MN488606 | MN488608 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lau, Y.W.; Reimer, J.D. Zooxanthellate, Sclerite-Free, and Pseudopinnuled Octocoral Hadaka nudidomus gen. nov. et sp. nov. (Anthozoa, Octocorallia) from Mesophotic Reefs of the Southern Ryukyus Islands. Diversity 2019, 11, 176. https://0-doi-org.brum.beds.ac.uk/10.3390/d11100176
Lau YW, Reimer JD. Zooxanthellate, Sclerite-Free, and Pseudopinnuled Octocoral Hadaka nudidomus gen. nov. et sp. nov. (Anthozoa, Octocorallia) from Mesophotic Reefs of the Southern Ryukyus Islands. Diversity. 2019; 11(10):176. https://0-doi-org.brum.beds.ac.uk/10.3390/d11100176
Chicago/Turabian StyleLau, Yee Wah, and James Davis Reimer. 2019. "Zooxanthellate, Sclerite-Free, and Pseudopinnuled Octocoral Hadaka nudidomus gen. nov. et sp. nov. (Anthozoa, Octocorallia) from Mesophotic Reefs of the Southern Ryukyus Islands" Diversity 11, no. 10: 176. https://0-doi-org.brum.beds.ac.uk/10.3390/d11100176