Bacterial Diversity and Community Structure in Typical Plant Rhizosphere



Food Security and Safety Niche, Faculty of Natural and Agricultural Sciences, North-West University, Private Mail Bag X2046, Mmabatho 2735, South Africa
*
Author to whom correspondence should be addressed.
Diversity 2019, 11(10), 179; https://0-doi-org.brum.beds.ac.uk/10.3390/d11100179
Submission received: 11 September 2019
/
Revised: 24 September 2019
/
Accepted: 24 September 2019
/
Published: 30 September 2019
Abstract
:Bacteria play a vital role in the quality of soil, health, and the production of plants. This has led to several studies in understanding the diversity and structure in the plant rhizosphere. Over the years, there have been overwhelming advances in molecular biology which have led to the development of omics techniques which utilize RNA, DNA, or proteins as biomolecules; these have been gainfully used in plant–microbe interactions. The bacterial community found in the rhizosphere is known for its colonization around the roots due to availability of nutrients, and composition, and it affects the plant growth directly or indirectly. Metabolic fingerprinting enables a snapshot of the metabolic composition at a given time. We review metabolites with ample information on their benefit to plants and which are found in rhizobacteria such as Pseudomonas spp. and Bacillus spp. Exploring plant-growth-promoting rhizobacteria using omics techniques can be a true success story for agricultural sustainability.
1. Introduction
Soils are home to a variety of bacteria, be they neutral, negative (pathogenic), allelopathic, or beneficial (symbiotic) to plants, and the interactions of these bacteria happen within the soil matrix (Table 1). This gives elucidation as to why soils are the most diverse habitats in the planet to the extent that even in harsh soil conditions bacteria still thrive, as reported for the coldest, driest desert on Earth [1]. The plant rhizosphere consists of diverse microorganisms which modulate the physiology and morphology and in the process, improve plant growth through the promotion of hormones and also serve as protectants against plant pathogens [2]. It is a critical zone of soil encompassing the plant root, thereby making it a hot spot of high abundance and diversity of microorganisms. The plant rhizosphere also attracts bacteria from the soil environment, such as the plant-growth-promoting rhizobacteria [3].
The soil has a great effect on plants and bacteria and vice versa. This is a result of the influence of plants on the soil through rhizodeposits, water, plant litter, gas, and nutrient exchanges [4]. Where rhizodeposits are present, bacteria abound and so such environments are richer in microflora and available nutrients than the external environment, which could be termed as the bulk soil [5]. The interface between root and soil is a strategic entryway for plants to absorb water and mineral nutrients from the soil environment and release rhizodeposits into the soil. These rhizodeposits which occur in different forms facilitate below-ground interactions between plants and microorganisms, and consequently affect the biodiversity of the region. Naturally, root–microbial interactions are very complex since such interactions involve myriads of microorganisms [6].
Plants, being the major sources of organic carbon in the soil, can be drivers of microbial growth and activity; hence, plants can affect the structure of the bacterial community in the soil [7]. Additionally, plant root–soil and microbial interaction has a key influence on plant community dynamics and nutrient cycling. A transformation in the plant community structure affects litter composition, which changes nutrient turnover rates and soil characteristics. The change in soil characteristics might further cause a change in plant community structure and composition [8].
Under unfavorable conditions, plant-growth-promoting rhizobacteria alleviate the effects in plants of such parameters as germination rate, drought tolerance, and plant yield components. The use of microorganisms in agriculture for crop protection against plant pathogens and pests because of the metabolites they produce, and also biological control against diseases, may present an alternative for plant disease prevention [9,10].
Since microorganisms found in the rhizosphere are important for plant health and biogeochemical cycles, engineering the rhizosphere may finally put an end to the use of agrochemicals by substituting their functions with beneficial microbes. Therefore, understanding the community structure and diversity of active microorganisms in the rhizosphere is key to enhancing plant growth and increasing agricultural productivity [11].
2. Rhizosphere Bacterial Community
The rhizosphere is home to enormous numbers of diverse bacterial species, some of which are culturable and some are yet unculturable. Culturable bacteria form an important part of the rhizosphere, the majority of which are Gram negative [21,22]. Among such bacteria are the rhizobacteria, which are characterized by aggressive colonization and subsequent establishment on plant roots. Many studies have characterized rhizobacteria in the rhizosphere of different plant species. Rhizobacterial genera viz., Acetobacter, Acinetobacter, Alcaligenes, Arthrobacter, Azoarcus, Azospirillum, Azotobacter, Bacillus, Beijerinckia, Burkholderia, Derxia, Enterobacter, Gluconacetobacter, Herbaspirillum, Klebsiella, Ochrobactrum, Pantoae, Pseudomonas, Rhodococcus, Serratia, Stenotrophomonas, and Zoogloea have been subjected to extensive research for decades [5,23] and have been implicated for beneficial effects on plant yield components. Of special interest are those that are common and peculiar to a locality [22] and those that have plant-growth-promoting traits such as Pseudomonas fluorescens NWU65, Vibrio fluvialis NWU37, Ewingella americana NWU59, P. putida NWU12 [22], the Actinobacteria [24,25], and many proteobacteria [26,27,28].
The bacterial population in the soil can grow very rapidly and make use of a wide range of different substances as nutrient sources. Over the last decade, the role of plant growth promoting rhizobacteria (PGPR) cannot be overemphasized due to their positive effect on plants. About 2–5% of bacteria found in the rhizosphere have plant-growth-promoting traits. Thus, they are potential tools for sustainable agriculture in the future [29]. The PGPR can promote growth either in direct or indirect ways by means of various mechanisms, which include providing a nitrogen source for plants through nitrogen fixation, applying biological control measures in combating soil-borne pathogens, producing plant growth substances (phytohormones) that improve plant development, and producing metabolites such as siderophores, antibiotics, cyanides, and ammonia [30].
It is conventional to observe cultivar effects on the bacterial composition of the rhizosphere because of the different compounds released by the root, collectively termed root exudate (Figure 1). Root exudates and other root deposits confer physical and chemical changes to the soil rhizosphere when compared to the bulk soil. The root exudates released are quickly assimilated by the root-associated microbes and modified before being discharged into the rhizosphere soil by microorganisms [8]. The absorption of root deposits by rhizosphere microbes enhances soil quality as seen in a study by Beauregard et al. [31], where root polysaccharides caused the biofilm matrix production of Bacillus subtilis, a microorganism beneficial to plants. These microbes do not only perceive secreted signals from the plant root, but also release different signaling molecules to control their host plant by improving biotic and abiotic stress tolerance or resistance, plant growth, and root development [8].
Bacteria in the rhizosphere respond differently to root exudates which stimulate great bacterial biomass and activity. A typical cultivar effect was reported among the cowpea cultivars—Rabuor (Kenya) local, IT95K-286-4, and IT94D-437-1 [9]. The effect of cultivars on pod characteristics was highly significant for certain microbial inoculants and helps in the yield of cowpea plants.
3. Factors Controlling the Distribution and Abundance of Bacterial Community in the Rhizosphere
Bacterial functional diversity is the ability of bacterial communities to use a wide spectrum of different compounds such as cellulose, sugars, and lignin that occur in plant tissues and whose proportional abundance changes during decomposition [32]. Other investigations indicate that the type and amount of available organic substrates in the rhizosphere strongly influence the abundance of bacterial groups and their functional diversity [33,34]. Minerals such as nitrogen and iron can also affect the number of bacteria than can be found in the rhizosphere and change the composition of the rhizobacterial community. It may be possible that potential nitrogen fixing and phosphate solubilizing bacteria add to plant nutrients by improving nitrogen and phosphorus intake by the plants, which could lead to an increase in plant yield [35].
Rhizosphere-inhabiting bacteria are most likely to emanate from surrounding bulk soil and often times from the seeds, then thrive under conditions that prevail in the plant roots. Plant type, plant age, soil pH, plant growth stages, organic compounds, and nutrient availability are some of the factors that influence the abundance and distribution of rhizobacteria in the soil. In other words, plant species, plant cultivars, plant developmental stages, and soil characteristics have thus been identified as major factors that contribute to the determination of composition of the rhizosphere community [36].
4. How Plant Growth Stages Affect Bacterial Community Dynamics
The bacterial communities in the rhizosphere region benefit from the nutrients that are secreted by younger roots and are under constant influence from their environment thus bringing changes to the community. Rhizobacteria are not static; they are subjected to changing environmental conditions of temperature, water content, and nutrient availability. It has been discovered that in young plant roots, bacterial communities are controlled by r-strategists, which are species having growth rates and abilities to make use of simple substrates [37,38]. As the roots of these young plants mature, there are specific elements of root exudates that can have selective control in the rhizosphere by spurning some species and strengthening the competitive activity and population of other species. As a result, there is a slight movement in dominance to bacterial communities with relatively slow growth rates and those that have the ability to degrade more complex substrates (k-strategists). As the root tips of the plant matures, soil microorganisms will colonize the roots and the population densities will increase rapidly a few centimeters from the root tips, and that is where insoluble, soluble, and volatile root exudates are used by the rhizobacteria for growth and metabolism [39]. The different root zones in a plant can support distinct bacterial communities showing the differences in root exudation qualitatively and quantitatively [40]. Soil type also plays a crucial role in the determination of the unique dominant bacteria that colonize the rhizosphere [41]. Therefore, the rhizosphere bacterial population belonging to the same plant species may differ both spatially and temporally.
5. Plant–Root Interactions in the Rhizosphere
The key roles of plant roots are to anchor the plant, absorb water and essential nutrients as well as store the nutrients and accumulate and secrete a diverse array of compounds including several primary and secondary metabolites, proteins, and peptides [42]. There is therefore a relationship between the plant roots and the underground parts of the soil. The interface between plant root and soil is influenced by different interactions triggered by soil microorganisms and plants roots.
Plants release nutrients and other organic substances in the rhizosphere region which attract different kinds of bacteria [24]. There is a signal in the rhizosphere when a plant releases exudate from its root. These signals can recruit nitrogen-fixing and growth-promoting bacteria, for instance rhizobia. Plant species belonging to the family Fabaceae mainly benefit in this association. In addition, plant-produced flavonoids are also involved in the establishment of these associations [43]. Bacteria colonize the rhizoplane and the rhizosphere, which is an important interaction between the plant roots and the surrounding soil. There is a regular flow of organic substrates from the plant which the bacteria take advantage of and as a result promote the growth of the plant through provision of soluble inorganic nutrients and production of growth-promoting substances [44]. Hence, the level of interaction with the plant root is shaped by the nature of rhizodeposits and soil properties. In other words, rhizodeposition varies depending on the growth stages and species of the plant, in addition to the conditions of the environment [45].
The soil physical structure supports the above ground part of the plant by the root system. The plant also requires certain essential elements from the soil such as N, P, K, S, Mg, and Ca (macronutrients) as well as B, Cu, Fe, Mn, Mo, Ni, and Zn (micronutrients). The presence of toxic elements in the soil can also limit plant growth.
6. Influence of Plant Exudates on Rhizosphere Microbial Dynamics
In a study by Mendes et al. [46] on soybean plants, certain bacterial groups with specific nutritional functions were more present in the rhizosphere when compared to the bulk soil. The bacterial groups included those that were involved in iron uptake and metabolism, membrane transport, nitrogen, phosphorus, and potassium metabolism. These nutritional traits might be of benefit to the plants and this implies that soybean plants may attract specific microbial groups from the bulk soil into the rhizosphere on the basis of the functional qualities that enhance their productivity. “Secretion system type IV” which falls under the membrane transport is involved in the mutualistic associations between bacteria and other organisms. This membrane transport is present in Acidobacteria, Proteobacteria, Actinobacteria and Bacteroidetes as reported by Mendes et al [46]. Thus, it is expected that those plants that need the secretion system for their optimal performance will have more bacterial communities associated with the above listed phyla (Table 2). Microorganisms implicated in the metabolism of potassium are Rhizobium, Pseudomonas, Bacillus, and Flavobacterium. These bacteria are able to make potassium more available for plant use [47], while the rhizobia are involved in nitrogen metabolism including nitrogen fixation.
The exudate from plants is also a determining factor of species richness and abundance in the rhizosphere. Carbon is one of the photosynthates fixed by plants which is partly transported into the root zone and excreted from root tissues. Various organic acids such as aliphatic and aromatic acids, amides, carbohydrates (glucose and xylose), fructose, lactic, malic, oxalic, pyruvic, succinic, and amino acids secreted from the root play roles in chemotaxis and are also referred to as PGPR bioactive factors [5]. Some examples of the amino acids are alanine, asparagine, glutamic acid, glutamine, isoleucine, leucine, serine, and valine. These acids, as well as other compounds, are released into the rhizosphere and thus serve as nutrients for rhizobacteria. The composition of plant exudates is cultivar dependent and is influenced by plant exposure to stress and the plant growth stage.
7. Metabolites
The rhizosphere possesses a detailed dynamic ecological relationship between plant and microbes. A wide range of chemicals are usually secreted, and can be grouped as signaling compounds, growth regulators, and nutrient solubilizers. The determination of chemicals involved in this signaling can help in sustainable agriculture, most especially as biocontrol agents [66,67].
Some of the metabolites secreted by plant roots are (Table 3) glucoberin, 4-methyl-2-pentanone, alkaloids, amino acids, benzaldehyde, biochanin A, calystegine, cyanidin, dehydrotomatine flavonoids, formononetin, furfural, glycoalkaloids, glycosides, kaempferol, lectins, maeckiain, medicarpin, okundoperoxide, p-coumaric acid, peptides, phylloquinone, polyphenols, quercetin glycosides, rhamnoside, salicylic acid, terpenoids, tryptophan, ϒ-butyrolatone, ϒ-terpinene, α-tomatine, and α-tocopherol [68]. Besides plant metabolites, bacteria also secrete bacterial metabolites. Some of the bacterial metabolites are 2,4-diacetylphloroglucinol, 2-amino phenol, 2-amino phenoxazone, antibiotics, auxofuran, bacillomycin D, beta-1,3-glucanase, butyl 2-pyrolidone-5-carboxylate, chitinase, exoprotease, fengycins, hydrogen cyanide (HCN), indole-acetic-acid (IAA), lipopeptides, phenazine-1-carboxamide, phenazines, phosphate, phytotoxins, prodigiosin protease, pyochelin, pyoluteorin, pyoverdine, pyrrolnitrin, salicylic acid, siderophores, surfactins, and viscosinamide [69].
Accumulations of secondary metabolites often occur in plants when they undergo various stresses, elicitors, or signal molecules. Synthesis of secondary metabolites increases when certain factors are involved. These factors are physical, chemical, and microbial factors [70].
8. Metabolic Fingerprinting
Biochemical fingerprinting can be of great benefit for scientific and commercial purposes in relation to plant breeding, response of plants to biotic and abiotic stresses, and characterization of genetic mutants. The study of metabolic fingerprinting in the plant rhizosphere is important, because metabolites are low-molecular-weight compounds synthesized by plants for essential functions, especially growth and development, and specific functions such as defense mechanisms. Metabolites can be analyzed using various approaches which can be classed into five categories [79]: (i) Metabolite target analysis—this approach is target-driven where one or a few single compounds are analyzed, (ii) metabolic profiling—an assortment of metabolites already defined according to a class of compounds or based on their correlation with a particular pathway. Examples of such class of compounds are carbohydrates, fatty acids, organic phosphates, or amino acids, (iii) metabolic fingerprinting—fast analysis of samples through pattern recognition using high throughput technology, (iv) metabolomics—non-biased identification and quantification of all the metabolites in a biological system, and (v) metabonomics—the measurement of dynamic changes across the metabolome of living systems with respect to time in response to physiological stimuli or genetic alteration [79]. Multiple methods are required to analyze different subsets of metabolites due to differences in volatility, polarity, solubility, and chromatographic behavior [80].
In the metabolic fingerprinting approach, the goal is not to classify each detected metabolite but to correlate patterns and fingerprint the metabolites that change in response to disease, toxin exposure, and environmental or genetic alteration [79].
9. The Use of Omics Techniques to Analyze the Rhizosphere
Next generation sequencing (NGS) has been used to infer the microbial community structure of the rhizosphere and the core microbial community. This has led to better understanding of the structure, abundance, spatial distribution diversity, and important members of the rhizosphere community [81]. Plants in which this platform has been studied include pea, soybean, corn, wheat, and oat [82]. The NGS tools are metagenomics, metatranscriptomics, metaproteomics, and metabolomics. A review on this has been done by White III et al. [81]. Although each omics study has its advantages and disadvantages, to better capture the structure and diversity in the rhizosphere, it is advisable to carry out multi-omics analysis which is a modern approach in system biology.
10. Conclusions and Future Outlooks
This review showed that different metabolites are found in rhizobacteria. They have a relevant role to play in agriculture as these metabolites are responsible for stimulating plant growth, suppressing diseases such as fungal phytopathogens and plant–parasitic nematodes by production of cyanide, siderophores, ammonia, and other volatile metabolites. Another benefit is that they have a positive effect on the growth of plants under abiotic stress such as salinity or drought. In crop production, these rhizobacteria can be used to improve crop yield by acting as biofertilizers, thereby helping to reduce reliance on chemical fertilizers and pesticides, which are known to pollute the environment. Furthermore, biofertilizers have the advantage of being eco-friendly and cheap.
Research in soil microbiology has largely been focused on the analysis of microbial processes that take place in the soil. We need to understand the microbial populations because they are the basic assemblages that drive these processes. It is undergoing a rapid transition where interdisciplinary approaches are required. It involves the collaboration in many areas of science like biology, physics, bioinformatics, mathematics, statistics, and computer science. Thus, to better understand the rhizosphere and develop new natural products, in-depth research will be needed in future where techniques, state-of-the-art technologies, along with robust data software and analysis are used.
The study of the rhizosphere is a rapidly advancing area of research and there has been tremendous progress in the usage of genetic fingerprinting techniques to study rhizobacterial communities that are found in the rhizosphere. However, quantitative assessment is still a huge challenge, due to the biases which are associated with the isolation of nucleic acids (DNA or RNA) and PCR, and none of these techniques provide full understanding and access of the genetic diversity of the bacterial community. Thus, omics studies are important tools which can be used to identify and characterize the microbial genes and functions that help microorganisms thrive in the plant rhizosphere. Such information will help improve the ability to fight plant diseases, and promote beneficial bacterial functions for agriculture, which will have direct positive effects on global food production, thereby enhancing food security.
Author Contributions
All authors contributed equally.
Funding
National Research Foundation, South Africa: UID119948.
Acknowledgments
T.T.A. would like to thank the North-West University for postgraduate bursary. O.O.B. thanks the National Research Foundation, South Africa for grant (UID119948) that has supported research in our laboratory.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Babalola, O.O.; Kirby, B.M.; Roes-Hill, L.; Cook, A.E.; Cary, S.C.; Burton, S.G.; Cowan, D.A. Phylogenetic analysis of actinobacterial populations associated with Antarctic Dry Valley mineral soils. Environ. Microbiol. 2009, 11, 566–576. [Google Scholar] [CrossRef]
- Philippot, L.; Raaijmakers, J.; Lemanceau, P.; Van Der Putten, W. Going back to the roots: The microbial ecology of the rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, R.; Gao, J.; Wang, X.; Fan, F.; Ma, X.; Yin, H.; Zhang, C.; Feng, K.; Deng, Y. Thirty-one years of rice-rice-green manure rotations shape the rhizosphere microbial community and enrich beneficial bacteria. Soil Biol. Biochem. 2017, 104, 208–217. [Google Scholar] [CrossRef]
- Fanin, N.; Bertrand, I. Aboveground litter quality is a better predictor than belowground microbial communities when estimating carbon mineralization along a land-use gradient. Soil Biol. Biochem. 2016, 94, 48–60. [Google Scholar] [CrossRef]
- Babalola, O.O. Beneficial bacteria of agricultural importance. Biotechnol. Lett. 2010, 32, 1559–1570. [Google Scholar] [CrossRef]
- Preece, C.; Penuelas, J. Rhizodeposition under drought and consequences for soil communities and ecosystem resilience. Plant Soil 2016, 409, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Lange, M.; Eisenhauer, N.; Sierra, C.; Bessler, H.; Engels, C.; Griffiths, R.; Mellado-Vázquez, P.; Malik, A.; Roy, J.; Scheu, S. Plant diversity increases soil microbial activity and soil carbon storage. Nat. Commun. 2015, 6, 6707. [Google Scholar] [CrossRef]
- Zhang, R.; Vivanco, J.; Shen, Q. The unseen rhizosphere root–soil–microbe interactions for crop production. Curr. Opin. Microbiol. 2017, 37, 8–14. [Google Scholar] [CrossRef]
- Babalola, O.O.; Sanni, A.I.; Odhiambo, G.D.; Torto, B. Plant growth-promoting rhizobacteria do not pose any deleterious effect on cowpea and detectable amounts of ethylene are produced. World J. Microbiol. Biotechnol. 2007, 23, 747–752. [Google Scholar] [CrossRef]
- Enebe, M.; Babalola, O. The influence of plant growth-promoting rhizobacteria in plant tolerance to abiotic stress: A survival strategy. Appl. Microbiol. Biotechnol. 2018, 102, 7821–7835. [Google Scholar] [CrossRef]
- Li, H.; Su, J.; Yang, X.; Zhu, Y. Distinct rhizosphere effect on active and total bacterial communities in paddy soils. Sci. Total Environ. 2019, 649, 422–430. [Google Scholar] [CrossRef]
- Caballero-Mellado, J.; Onofre-Lemus, J.; Estrada-De Los Santos, P.; Martínez-Aguilar, L. The tomato rhizosphere, an environment rich in nitrogen-fixing Burkholderia species with capabilities of interest for agriculture and bioremediation. Appl. Environ. Microbiol. 2007, 73, 5308–5319. [Google Scholar] [CrossRef]
- Petatan-Sagahon, I.; Anducho-Reyes, M.; Silva-Rojas, H.; Arana-Cuenca, A.; Tellez-Jurado, A.; Cárdenas-Álvarez, I.; Mercado-Flores, Y. Isolation of bacteria with antifungal activity against the phytopathogenic fungi Stenocarpella maydis and Stenocarpella macrospora. Int. J. Mol. Sci. 2011, 12, 5522–5537. [Google Scholar] [CrossRef]
- Saubidet, M.; Fatta, N.; Barneix, A. The effect of inoculation with Azospirillum brasilense on growth and nitrogen utilization by wheat plants. Plant Soil 2002, 245, 215–222. [Google Scholar] [CrossRef]
- Yanni, Y.; El-Fattah, F. Towards integrated biofertilization management with free living and associative dinitrogen fixers for enhancing rice performance in the Nile delta. Symbiosis 1999, 27, 319–331. [Google Scholar]
- Muratova, A.; Turkovskaya, O.; Antonyuk, L.; Makarov, O.; Pozdnyakova, L.; Ignatov, V. Oil-oxidizing potential of associative rhizobacteria of the genus Azospirillum. Microbiology 2005, 74, 210–215. [Google Scholar] [CrossRef]
- Åström, B.; Gustafsson, A.; Gerhardson, B. Characteristics of a plant deleterious rhizosphere Pseudomonad and its inhibitory metabolite (s). J. Appl. Bacteriol. 1993, 74, 20–28. [Google Scholar] [CrossRef]
- Murphy, J.; Zehnder, G.; Schuster, D.; Sikora, E.; Polston, J.; Kloepper, J. Plant growth-promoting rhizobacterial mediated protection in tomato against Tomato mottle virus. Plant Dis. 2000, 84, 779–784. [Google Scholar] [CrossRef]
- Carvalho, D.; Oliveira, D.; Corrêa, R.; Campos, V.; Guimarães, R.; Coimbra, J. Rhizobacteria able to produce phytotoxic metabolites. Braz. J. Microbiol. 2007, 38, 759–765. [Google Scholar] [CrossRef] [Green Version]
- Manivanh, L.; Pierret, A.; Rattanavong, S.; Kounnavongsa, O.; Buisson, Y.; Elliott, I.; Maeght, J.; Xayyathip, K.; Silisouk, J.; Vongsouvath, M. Burkholderia pseudomallei in a lowland rice paddy: Seasonal changes and influence of soil depth and physico-chemical properties. Sci. Rep. 2017, 7, 3031. [Google Scholar] [CrossRef]
- Babalola, O.; Akindolire, A. Identification of native rhizobacteria peculiar to selected food crops in Mmabatho municipality of South Africa. Biol. Agric. Hortic. 2011, 27, 294–309. [Google Scholar] [CrossRef]
- Hou, M.P.; Babalola, O.O. Evaluation of plant growth promoting potential of four rhizobacterial species for indigenous system. J. Cent. South Univ. 2013, 20, 164–171. [Google Scholar] [CrossRef]
- Olanrewaju, O.; Glick, B.; Babalola, O. Mechanisms of action of plant growth promoting bacteria. World J. Microbiol. Biotechnol. 2017, 33, 197. [Google Scholar] [CrossRef]
- Adegboye, M.; Babalola, O. Phylogenetic characterization of culturable antibiotic producing Streptomyces from rhizospheric soils. Mol. Biol. S 2013, 1, 001. [Google Scholar]
- Adegboye, M.F.; Babalola, O.O.; Ngoma, L.; Okoh, A.I. Analysis of Streptomyces spp. native to Mahikeng soils in South Africa. J. Pure Appl. Microbiol. 2012, 6, 1001–1010. [Google Scholar]
- Babalola, O.O.; Glick, B.R. The use of microbial inoculants in African agriculture: Current practice and future prospects. J. Food Agric. Environ. 2012, 10, 540–549. [Google Scholar]
- Babalola, O.O. Molecular techniques: An overview of methods for the detection of bacteria. Afr. J. Biotechnol. 2003, 2, 710–713. [Google Scholar] [Green Version]
- Singh, B.K.; Munro, S.; Potts, J.M.; Millard, P. Influence of grass species and soil type on rhizosphere microbial community structure in grassland soils. Appl. Soil Ecol. 2007, 36, 147–155. [Google Scholar]
- Goswami, D.; Thakker, J.; Dhandhukia, P. Portraying mechanics of plant growth promoting rhizobacteria (PGPR): A review. Cogent Food Agric. 2016, 2, 1127500. [Google Scholar] [CrossRef]
- Bumunang, E.W.; Babalola, O.O.; Barros, E. Bacterial community profiling in the rhizosphere of field grown GM and non-GM maize. J. Pure Appl. Microbiol 2013, 7, 1–21. [Google Scholar]
- Beauregard, P.; Chai, Y.; Vlamakis, H.; Losick, R.; Kolter, R. Bacillus subtilis biofilm induction by plant polysaccharides. Proc. Natl. Acad. Sci. USA 2013, 110, E1621–E1630. [Google Scholar] [CrossRef]
- Lahav, I.; Steinberger, Y. Soil bacterial functional diversity in a potato field. Eur. J. Soil Biol. 2001, 37, 59–67. [Google Scholar] [CrossRef]
- Grayston, S.; Griffith, G.; Mawdsley, J.; Campbell, C.; Bardgett, R.D. Accounting for variability in soil microbial communities of temperate upland grassland ecosystems. Soil Biol. Biochem. 2001, 33, 533–551. [Google Scholar] [CrossRef]
- Lejon, D.P.; Sebastia, J.; Lamy, I.; Chaussod, R.; Ranjard, L. Relationships between soil organic status and microbial community density and genetic structure in two agricultural soils submitted to various types of organic management. Microb. Ecol. 2007, 53, 650–663. [Google Scholar] [CrossRef]
- Yadav, B.; Akhtar, M.; Panwar, J. Rhizospheric plant-microbe interactions: Key factors to soil fertility and plant nutrition. In Plant Microbes Symbiosis: Applied Facets; Springer: New Delhi, India, 2015; pp. 127–145. [Google Scholar]
- Schlemper, T.; Leite, M.; Lucheta, A.; Shimels, M.; Bouwmeester, H.; van Veen, J.; Kuramae, E. Rhizobacterial community structure differences among sorghum cultivars in different growth stages and soils. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef] [Green Version]
- Brimecombe, M.J.; Leij, F.A.; Lynch, J.M. Nematode community structure as a sensitive indicator of microbial perturbations induced by a genetically modified Pseudomonas fluorescens strain. Biol. Fertil. Soils 2001, 34, 270–275. [Google Scholar] [CrossRef]
- Zhou, D.; Huang, X.; Chaparro, J.; Badri, D.; Manter, D.; Vivanco, J.; Guo, J. Root and bacterial secretions regulate the interaction between plants and PGPR leading to distinct plant growth promotion effects. Plant Soil 2016, 401, 259–272. [Google Scholar] [CrossRef]
- Marschner, P.; Joergensen, R.; Piepho, H.-P.; Buerkert, A. Legume rotation effects on early growth and rhizosphere microbiology of sorghum in West African soils. Plant Soil 2004, 264, 325–334. [Google Scholar] [CrossRef] [Green Version]
- Reinhold-Hurek, B.; Bünger, W.; Burbano, C.; Sabale, M.; Hurek, T. Roots shaping their microbiome: Global hotspots for microbial activity. Annu. Rev. Phytopathol. 2015, 53, 403–424. [Google Scholar] [CrossRef]
- Marschner, P.; Yang, C.-H.; Lieberei, R.; Crowley, D. Soil and plant specific effects on bacterial community composition in the rhizosphere. Soil Biol. Biochem. 2001, 33, 1437–1445. [Google Scholar] [CrossRef]
- Weisskopf, L.; Abou-Mansour, E.; Fromin, N.; Tomasi, N.; Santelia, D.; Edelkott, I.; Neumann, G.; Aragno, M.; Tabacchi, R.; Martinoia, E. White lupin has developed a complex strategy to limit microbial degradation of secreted citrate required for phosphate acquisition. Plant Cell Environ. 2006, 29, 919–927. [Google Scholar] [CrossRef] [Green Version]
- Rasmann, S.; Turlings, T. Root signals that mediate mutualistic interactions in the rhizosphere. Curr. Opin. Plant Biol. 2016, 32, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Kai, M.; Effmert, U.; Piechulla, B. Bacterial-plant-interactions: Approaches to unravel the biological function of bacterial volatiles in the rhizosphere. Front. Microbiol. 2016, 7, 108. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; Van Themaat, E.; Schulze-Lefert, P. Structure and functions of the bacterial microbiota of plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef]
- Mendes, L.W.; Kuramae, E.E.; Navarrete, A.A.; Van Veen, J.A.; Tsai, S.M. Taxonomical and functional microbial community selection in soybean rhizosphere. ISME J. 2014, 8, 1577–1587. [Google Scholar] [CrossRef]
- Raghavendra, M.; Nayaka, S.; Nuthan, B. Role of rhizosphere microflora in potassium solubilization. In Potassium Solubilizing Microorganisms for Sustainable Agriculture; Springer: New Delhi, India, 2016; pp. 43–59. [Google Scholar]
- Bloemberg, G.; Lugtenberg, B. Molecular basis of plant growth promotion and biocontrol by Rhizobacteria. Curr. Opin. Plant Biol. 2001, 4, 343–350. [Google Scholar] [CrossRef]
- Dimkpa, C.; Svatoš, A.; Dabrowska, P.; Schmidt, A.; Boland, W.; Kothe, E. Involvement of siderophores in the reduction of metal-induced inhibition of auxin synthesis in Streptomyces spp. Chemosphere 2008, 74, 19–25. [Google Scholar] [CrossRef]
- Koumoutsi, A.; Chen, X.; Henne, A.; Liesegang, H.; Hitzeroth, G.; Franke, P.; Vater, J.; Borriss, R. Structural and functional characterization of gene clusters directing nonribosomal synthesis of bioactive cyclic lipopeptides in Bacillus amyloliquefaciens strain FZB42. J. Bacteriol. 2004, 186, 1084–1096. [Google Scholar] [CrossRef]
- Höfte, M.; Bakker, P. Competition for iron and induced systemic resistance by siderophores of plant growth promoting rhizobacteria. In Microbial Siderophores; Springer: Berlin/Heidelberg, Germany, 2007; pp. 121–133. [Google Scholar]
- Chen, X.; Koumoutsi, A.; Scholz, R.; Eisenreich, A.; Schneider, K.; Heinemeyer, I.; Morgenstern, B.; Voss, B.; Hess, W.; Reva, O. Comparative analysis of the complete genome sequence of the plant growth–promoting bacterium Bacillus amyloliquefaciens FZB42. Nat. Biotechnol. 2007, 25, 1007. [Google Scholar] [CrossRef]
- Kalbe, C.; Marten, P.; Berg, G. Strains of the genus Serratia as beneficial rhizobacteria of oilseed rape with antifungal properties. Microbiol. Res. 1996, 151, 433–439. [Google Scholar] [CrossRef]
- Raaijmakers, J.; Vlami, M.; De Souza, J. Antibiotic production by bacterial biocontrol agents. Antonie Van Leeuwenhoek 2002, 81, 537. [Google Scholar] [CrossRef]
- Arora, N.; Kang, S.; Maheshwari, D. Isolation of siderophore-producing strains of Rhizobium meliloti and their biocontrol potential against Macrophomina phaseolina that causes charcoal rot of groundnut. Curr. Sci. Bangalore 2001, 81, 673–677. [Google Scholar]
- Chin-A-Woeng, T.; van den Broek, D.; Lugtenberg, B.; Bloemberg, G. The Pseudomonas chlororaphis PCL1391 sigma regulator psrA represses the production of the antifungal metabolite phenazine-1-carboxamide. Mol. Plant Microbe Interact. 2005, 18, 244–253. [Google Scholar] [CrossRef]
- Boruah, H.; Kumar, B. Plant disease suppression and growth promotion by a Fluorescent Pseudomonas strain. Folia Microbiol. 2002, 47, 137–143. [Google Scholar] [CrossRef]
- Nagarajkumar, M.; Bhaskaran, R.; Velazhahan, R. Involvement of secondary metabolites and extracellular lytic enzymes produced by Pseudomonas fluorescens in inhibition of Rhizoctonia solani, the rice sheath blight pathogen. Microbiol. Res. 2004, 159, 73–81. [Google Scholar] [CrossRef]
- Loper, J.; Gross, H. Genomic analysis of antifungal metabolite production by Pseudomonas fluorescens Pf-5. In New Perspectives and Approaches in Plant Growth-Promoting Rhizobacteria Research; Springer: Dordrecht, The Netherlands, 2007; pp. 265–278. [Google Scholar]
- Park, M.; Kim, Y.; Lee, S.; Kim, I. Identification of an ISR-related metabolite produced by rhizobacterium Klebsiella oxytoca C1036 active against soft-rot disease pathogen in tobacco. Pest Manag. Sci. Former. Pestic. Sci. 2009, 65, 1114–1117. [Google Scholar] [CrossRef]
- Riedlinger, J.; Schrey, S.; Tarkka, M.; Hampp, R.; Kapur, M.; Fiedler, H. Auxofuran, a novel metabolite that stimulates the growth of fly agaric, is produced by the mycorrhiza helper bacterium Streptomyces strain AcH 505. Appl. Environ. Microbiol. 2006, 72, 3550–3557. [Google Scholar] [CrossRef]
- Gealy, D.; Gurusiddaiah, S.; Ogg, A. Isolation and characterization of metabolites from Pseudomonas syringae-strain 3366 and their phytotoxicity against certain weed and crop species. Weed Sci. 1996, 44, 383–392. [Google Scholar] [CrossRef]
- Gealy, D.; Gurusiddaiah, S.; Ogg, A.; Kennedy, A. Metabolites from Pseudomonas fluorescens strain D7 inhibit downy brome (Bromus tectorum) seedling growth. Weed Technol. 1996, 10, 282–287. [Google Scholar] [CrossRef]
- Jousset, A.; Lara, E.; Wall, L.; Valverde, C. Secondary metabolites help biocontrol strain Pseudomonas fluorescens CHA0 to escape protozoan grazing. Appl. Environ. Microbiol. 2006, 72, 7083–7090. [Google Scholar] [CrossRef]
- Kumar, R.; Ayyadurai, N.; Pandiaraja, P.; Reddy, A.; Venkateswarlu, Y.; Prakash, O.; Sakthivel, N. Characterization of antifungal metabolite produced by a new strain Pseudomonas aeruginosa PUPa3 that exhibits broad-spectrum antifungal activity and biofertilizing traits. J. Appl. Microbiol. 2005, 98, 145–154. [Google Scholar] [CrossRef]
- Verma, A.; Kumar, S.; Kumar, G.; Saini, J.; Agrawal, R.; Satlewal, A.; Ansari, M. Rhizosphere metabolite profiling: An opportunity to understand plant-microbe interactions for crop improvement. In Crop Improvement through Microbial Biotechnology; Elsevier: Amsterdam, The Netherlands, 2018; pp. 343–361. [Google Scholar]
- Beneduzi, A.; Ambrosini, A.; Passaglia, L. Plant growth-promoting rhizobacteria (PGPR): Their potential as antagonists and biocontrol agents. Genet. Mol. Biol. 2012, 35, 1044–1051. [Google Scholar] [CrossRef]
- Bais, H.; Park, S.; Weir, T.; Callaway, R.; Vivanco, J. How plants communicate using the underground information superhighway. Trends Plant Sci. 2004, 9, 26–32. [Google Scholar] [CrossRef]
- Santoyo, G.; Orozco-Mosqueda, M.; Govindappa, M. Mechanisms of biocontrol and plant growth-promoting activity in soil bacterial species of Bacillus and Pseudomonas: A review. Biocontrol Sci. Technol. 2012, 22, 855–872. [Google Scholar] [CrossRef]
- Thakur, M.; Bhattacharya, S.; Khosla, P.; Puri, S. Improving production of plant secondary metabolites through biotic and abiotic elicitation. J. Appl. Res. Med. Aromat. Plants 2019, 12, 1–12. [Google Scholar] [CrossRef]
- Cirlini, M.; Dall’Asta, C.; Silvanini, A.; Beghè, D.; Fabbri, A.; Galaverna, G.; Ganino, T. Volatile fingerprinting of chestnut flours from traditional Emilia Romagna (Italy) cultivars. Food Chem. 2012, 134, 662–668. [Google Scholar] [CrossRef]
- Choi, H.; Choi, Y.; Verberne, M.; Lefeber, A.; Erkelens, C.; Verpoorte, R. Metabolic fingerprinting of wild type and transgenic tobacco plants by 1H NMR and multivariate analysis technique. Phytochemistry 2004, 65, 857–864. [Google Scholar] [CrossRef]
- Sachdev, D.; Chaudhari, H.; Kasture, V.; Dhavale, D.; Chopade, B. Isolation and characterization of indole acetic acid (IAA) producing Klebsiella pneumoniae strains from rhizosphere of wheat (Triticum aestivum) and their effect on plant growth. Indian J. Exp. Biol. 2009, 47, 993–1000. [Google Scholar]
- Ndakidemi, P.; Dakora, F. Legume seed flavonoids and nitrogenous metabolites as signals and protectants in early seedling development. Funct. Plant Biol. 2003, 30, 729–745. [Google Scholar] [CrossRef]
- Armero, J.; Requejo, R.; Jorrín, J.; López-Valbuena, R.; Tena, M. Release of phytoalexins and related isoflavonoids from intact chickpea seedlings elicited with reduced glutathione at root level. Plant Physiol. Biochem. 2001, 39, 785–795. [Google Scholar] [CrossRef]
- Friedman, M. Potato glycoalkaloids and metabolites: Roles in the plant and in the diet. J. Agric. Food Chem. 2006, 54, 8655–8681. [Google Scholar] [CrossRef]
- Efange, S.; Brun, R.; Wittlin, S.; Connolly, J.; Hoye, T.; McAkam, T.; Makolo, F.; Mbah, J.; Nelson, D.; Nyongbela, K. Okundoperoxide, a bicyclic cyclofarnesylsesquiterpene endoperoxide from Scleria striatinux with antiplasmodial activity. J. Nat. Prod. 2009, 72, 280–283. [Google Scholar] [CrossRef]
- Kozukue, N.; Han, J.; Lee, K.; Friedman, M. Dehydrotomatine and α-tomatine content in tomato fruits and vegetative plant tissues. J. Agric. Food Chem. 2004, 52, 2079–2083. [Google Scholar] [CrossRef]
- Garcia-Perez, I.; Vallejo, M.; Garcia, A.; Legido-Quigley, C.; Barbas, C. Metabolic fingerprinting with capillary electrophoresis. J. Chromatogr. A 2008, 1204, 130–139. [Google Scholar] [CrossRef]
- Jorge, T.; Rodrigues, J.; Caldana, C.; Schmidt, R.; van Dongen, J.; Thomas-Oates, J.; António, C. Mass spectrometry-based plant metabolomics: Metabolite responses to abiotic stress. Mass Spectrom. Rev. 2016, 35, 620–649. [Google Scholar] [CrossRef]
- White III, R.; Rivas-Ubach, A.; Borkum, M.; Köberl, M.; Bilbao, A.; Colby, S.; Hoyt, D.; Bingol, K.; Kim, Y.; Wendler, J. The state of rhizospheric science in the era of multi-omics: A practical guide to omics technologies. Rhizosphere 2017, 3, 212–221. [Google Scholar] [CrossRef]
- Newman, M.; Lorenz, N.; Hoilett, N.; Lee, N.; Dick, R.; Liles, M.; Ramsier, C.; Kloepper, J. Changes in rhizosphere bacterial gene expression following glyphosate treatment. Sci. Total Environ. 2016, 553, 32–41. [Google Scholar] [CrossRef] [Green Version]
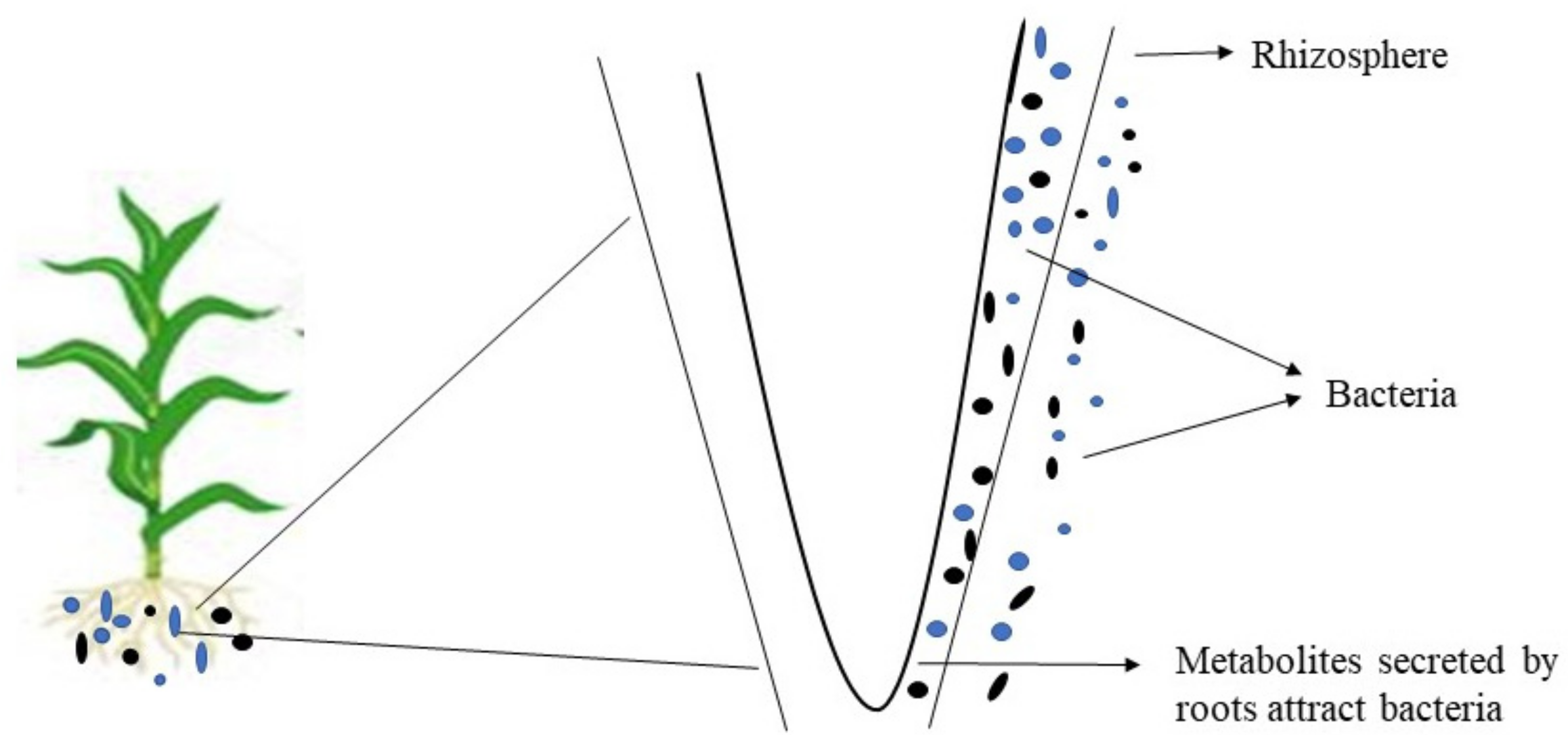
Figure 1.
Young roots shape the bacterial population in the rhizosphere community by secreting more organic materials when compared to the older roots.
Figure 1.
Young roots shape the bacterial population in the rhizosphere community by secreting more organic materials when compared to the older roots.


{kind=link}
Table 1.
Interactions occurring between the rhizosphere and bacteria can be positive, negative, or neutral.
Table 1.
Interactions occurring between the rhizosphere and bacteria can be positive, negative, or neutral.
| Plant Rhizosphere | Rhizobacteria | Effect | References |
|---|---|---|---|
| Positive effect | |||
| Tomato (Lycopersicon esculentum var. esculentum) | Burkholderia cepacia; B. unamae; B. tropica; B. xenovorans | N2 fixation, plant growth promotion, and bioremediation | [12] |
| Corn (Zea mays) | Bacillus subtilis; Pseudomonas fluorescens; Pantoea agglomerans | Antifungal activity | [13] |
| Wheat (Triticum aestivum) | Azospirillum brasilense | Promotes the uptake of NO3−, K+, and H2PO4− | [14] |
| Rice (Oryza sativa) | Azotobacter vinelandii; Azotobacter chroococcum | Plant growth promotion | [15] |
| Wheat | Azospirillum lipoferum | Promotes the development of the root system of wheat even under the contamination of crude oil | [16] |
| Negative effect | |||
| Wheat | Pseudomonas fluorescens angstrom313 | Plant growth reduction | [17] |
| Tomato | Bacillus subtilis | Causes sour skin, a bacterial soft-rotting disease of onion | [18] |
| Castor oil plant (Ricinus communis) | Bacillus cereus | Causes rootlet rot | [19] |
| Rice paddies | Burkholderia pseudomallei | Causative agents of melioidosis | [20] |
Table 2.
Interactions of metabolites released by the rhizobacteria and their importance to plants.
| Rhizobacteria | Metabolites | Function of the Metabolites | References |
|---|---|---|---|
| Pseudomonas spp. | Phenazines, pyrrolnitrin, pyoluteorin, viscosinamide | Viscosinamide found to prevent the infection of sugar beet by Pythium ultimum | [48] |
| Streptomyces spp. | Siderophores | Alleviate metal-contamination stress on plants | [49] |
| Bacillus amyloliquefaciens strain FZB 42 | lipopeptides, surfactins, bacillomycin D, fengycins | Antifungal activity | [50] |
| P. aeruginosa | Pyoverdine, pyochelin, salicylic acid | Induces resistance to plant diseases caused by Botrytis cinerea on bean and tomato, Colletotrichum lindemuthianum | [51] |
| Bacillus amyloliquefaciens | Bacillomycin D, fengycins, surfactins | Enhance plant growth and suppress plant pathogenic organisms | [52] |
| Serratia spp. | Siderophores, pyrrolnitrin, prodigiosin | Antifungal activity against different phytopathognic fungi Verticillium dahlia, Rhizoctonia solani, Sclerotinia sclerotiorum | [53] |
| P. fluorescens | Pyoluteorin, pyrrolnitrin, phenazine-1-carboxylic acid, 2,4-diacetylphloroglucinol | Suppress soil borne plant pathogens, including fungi and nematodes | [54] |
| Rhizobium meliloti | Siderophores | Antifungal activity against Macrophomina phaseolina | [55] |
| P. chlorophis PCL1391 | Phenazine-1-carboxamide | Antifungal | [56] |
| P. fluorescens | Siderophore pyoverdin | Antibacterial and antifungal activity. Enhanced plant growth | [57] |
| P. fluorescens PfMDUS | Chitinase, beta-1,3-glucanase, siderophores, salicylic acid, hydrogen cyanide | Inhibit mycelial growth of R. solani | [58] |
| P. fluorescens Pf-5 | Antibiotics, siderophores | Antibiotics toxic to soil-borne fungi and oomycetes that infect plant roots/siderophores involved in iron acquisition | [59] |
| Klebsiella oxytoca C1036 | Butyl 2-pyrolidone-5-carboxylate | Active against soft-rot disease pathogen in tobacco | [60] |
| Streptomyces AcH 505 | Auxofuran | Stimulates the growth of fly agaric, suppresses growth of ectomycorrhizal fungi | [61] |
| P. syringae strain 366 | Phenazine-1-carboxylic acid, 2-amino phenoxazone, 2-amino phenol | Inhibit downy brome root growth | [62] |
| P. fluorescens D7 | Uncharacterized phytotoxins | Inhibit downy brome root growth | [63] |
| P. fluorescens CHAO | Antibiotics, hydrogen cyanide, and an exoprotease | Protists’ growth inhibition, encystation, paralysis, and cell lysis | [64] |
| P. aeruginosa PUPa3 | Phenazine-1-carboxamide | Broad-spectrum antifungal activity and biofertilizing traits | [65] |
Table 3.
Plant roots secrete a range of metabolites into the rhizosphere.
| Plant | Metabolites | Function of the Metabolites | References |
|---|---|---|---|
| Sweet chestnut (Castanea sativa Mill.) | ϒ-butyrolactone, ϒ-terpinene, furfural, benzaldehyde, 4-methyl-2-pentanone | Peculiar aroma, the most typical organoleptic characteristic of chestnut-based products | [71] |
| Tobacco (Nicotiana tabacum) | Salicylic acid, alkaloids, flavonoids | Plays a role in plant growth and development, photosynthesis, ion uptake, and transport. Resistant to pathogens by inducing production of pathogenesis-related proteins. UV filtration and symbiotic nitrogen fixation | [72] |
| Wheat (Triticum spp.) | Tryptophan | For the biosynthesis of indole-3-acetic acid by associative bacteria | [73] |
| Legume seeds | Flavonoids, alkaloids, terpenoids, peptides, amino acids | Serve as eco-sensing signals for suitable rhizobacteria and arbuscular mycorrhizal fungal towards symbiotic mutualisms. Defense molecules against pathogens and insect pests. | [74] |
| Chickpea (Cicer arietinum) | Biochanin A, medicarpin, formononetin, maeckiain | Not stated | [75] |
| Potato (Solanum tuberosum) | Glycoalkaloids, calystegine, lectins | They protect plants against phytopathogens | [76] |
| Spice (Scleria stiatinux) | Okundoperoxide | It contains antiviral and antifungal properties, also used as herbal tea for fevers | [77] |
| Tomato (Solanum lycopersicum) | α-tomatine, dehydrotomatine | Defend plants against attack by microorganisms and herbivores due to their insecticidal activity and have allelopathic effects on many weeds. | [78] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Alawiye, T.T.; Babalola, O.O. Bacterial Diversity and Community Structure in Typical Plant Rhizosphere. Diversity 2019, 11, 179. https://0-doi-org.brum.beds.ac.uk/10.3390/d11100179
AMA Style
Alawiye TT, Babalola OO. Bacterial Diversity and Community Structure in Typical Plant Rhizosphere. Diversity. 2019; 11(10):179. https://0-doi-org.brum.beds.ac.uk/10.3390/d11100179
Chicago/Turabian StyleAlawiye, Temitayo Tosin, and Olubukola Oluranti Babalola. 2019. "Bacterial Diversity and Community Structure in Typical Plant Rhizosphere" Diversity 11, no. 10: 179. https://0-doi-org.brum.beds.ac.uk/10.3390/d11100179
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.