Species-Specific Functional Morphology of Four US Atlantic Coast Dune Grasses: Biogeographic Implications for Dune Shape and Coastal Protection

 , , , , , ,
, , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- How do the four dominant dune grass species compare in their distribution and abundance with latitude and location on the foredune profile?
- How do the four species of dune grasses differ in their functional morphology and sand accretion in the field?
2. Materials and Methods
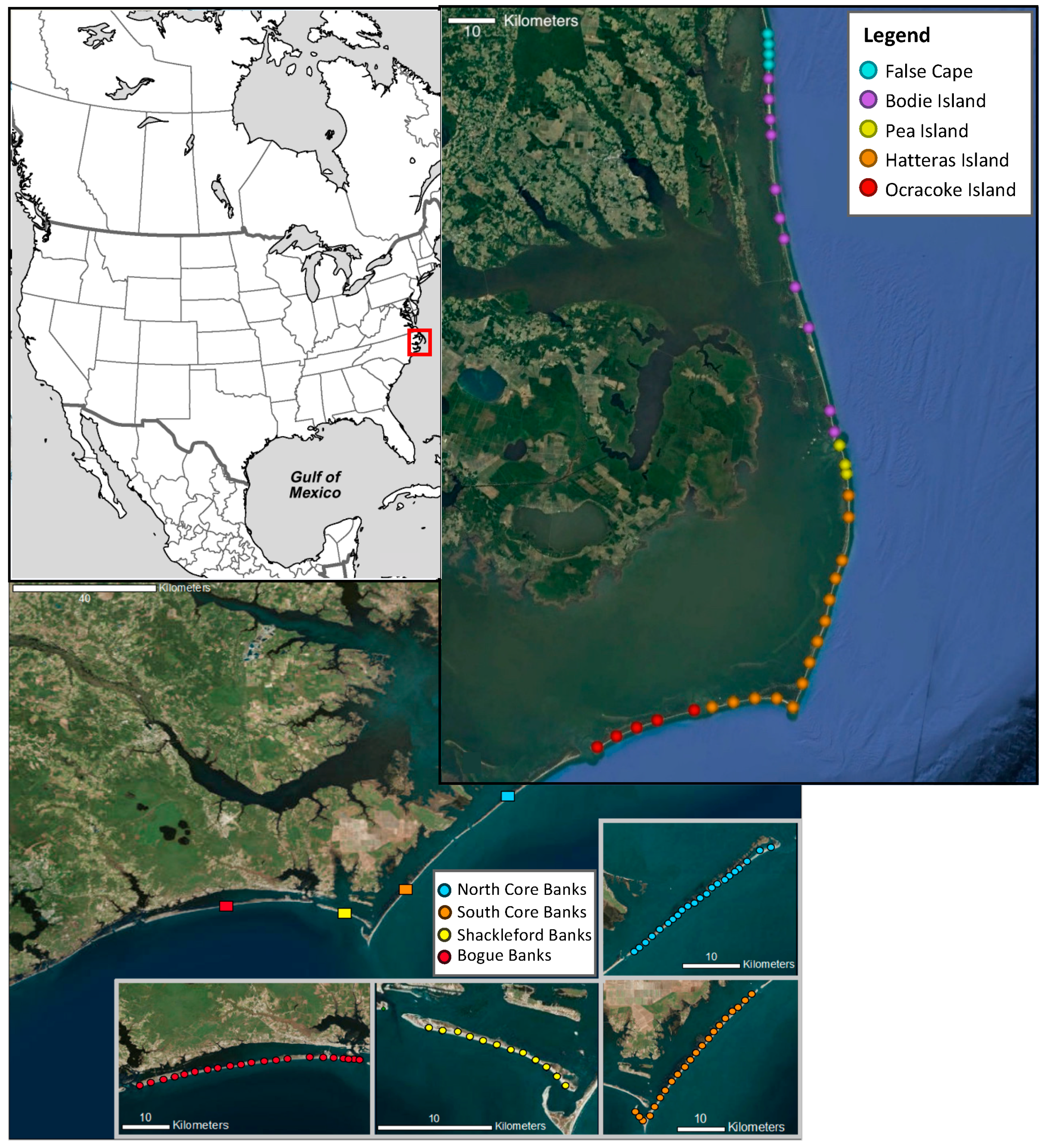
2.1. Field Setting
2.2. Foredune Plant and Grass Distribution and Abundance Along the US Outer Banks Islands
2.3. Dune Grass Growth Form, Morphology, and Changes in Sand Elevation
2.4. Data and Statistical Analyses
3. Results
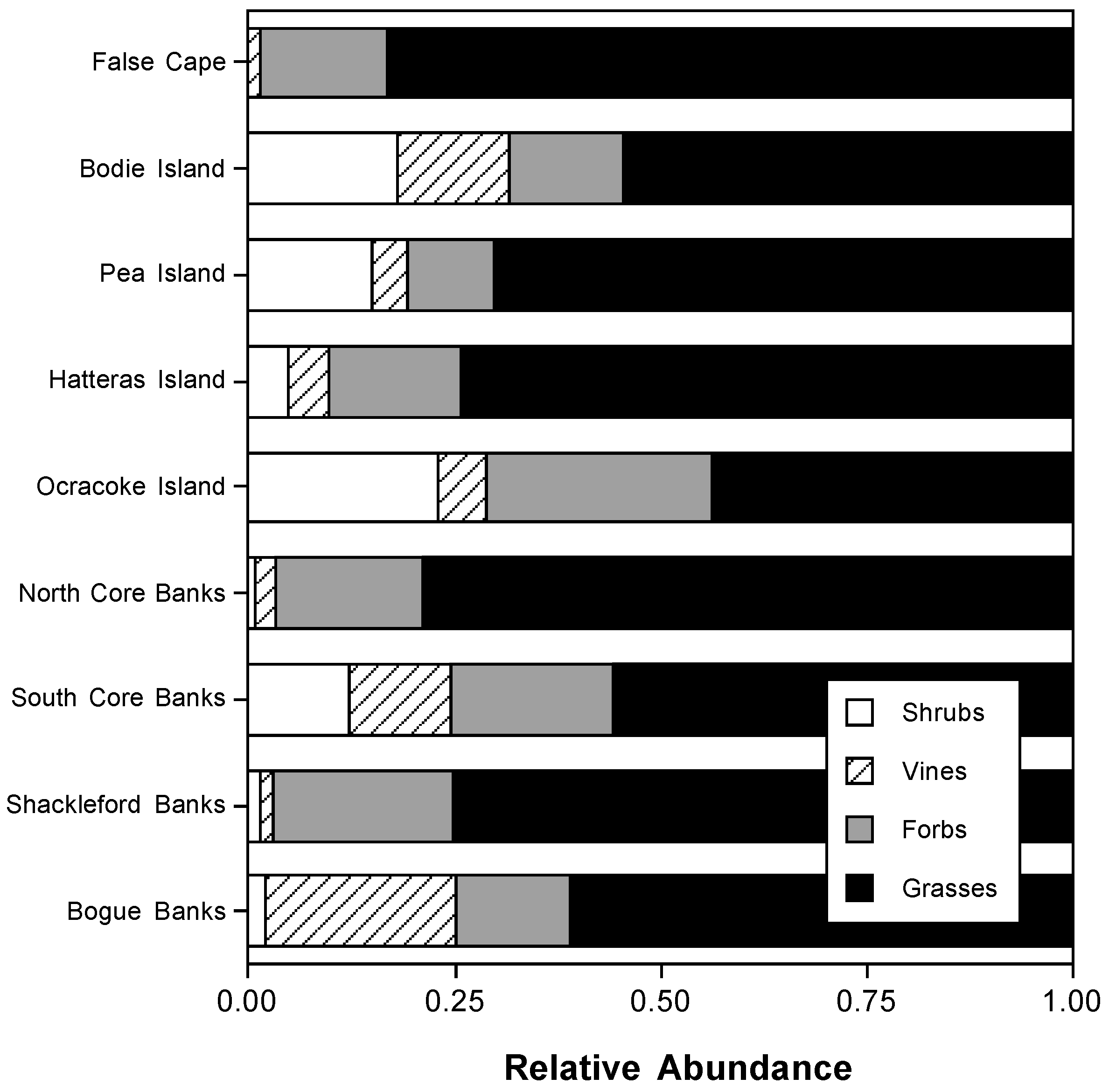
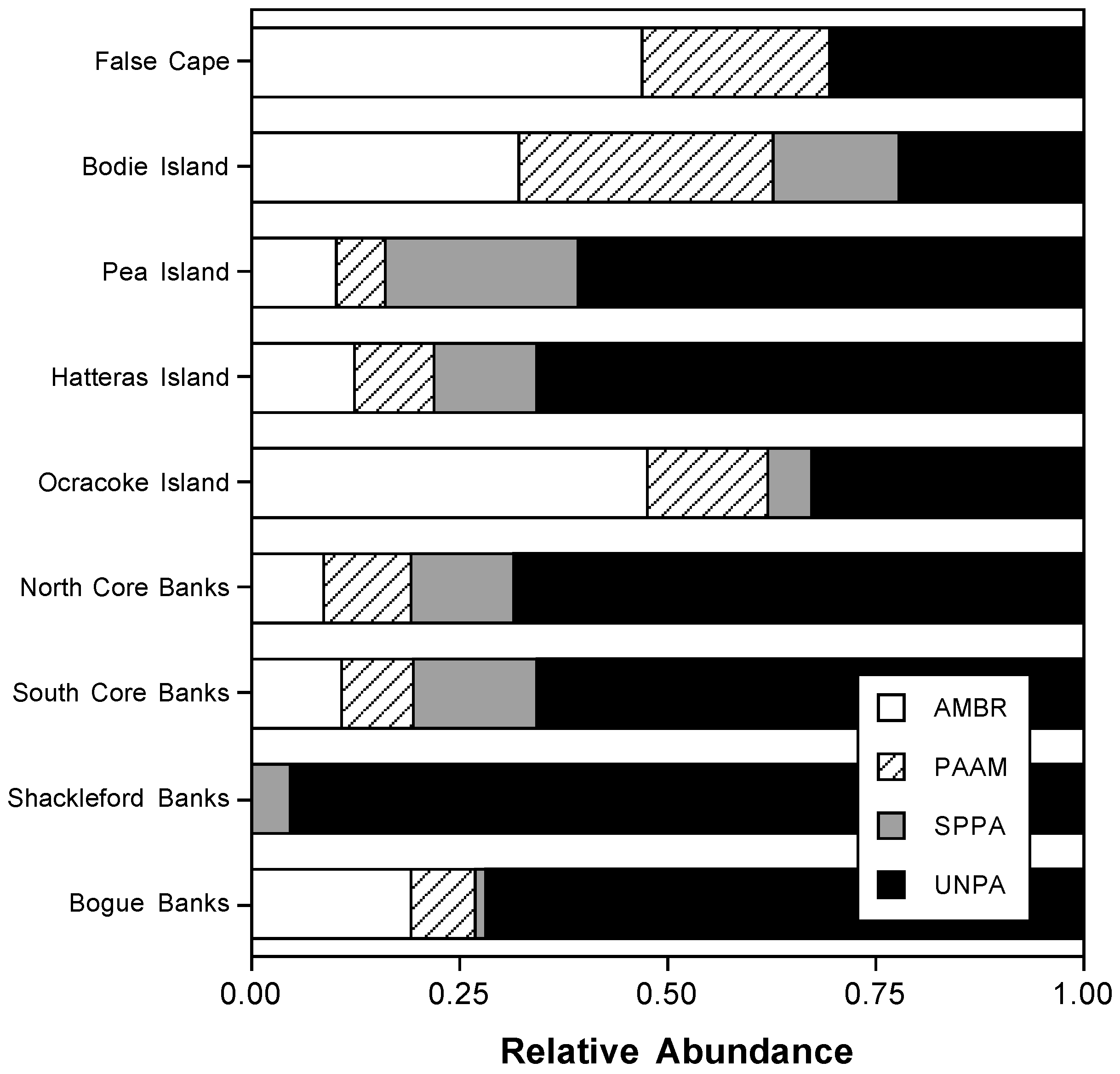
3.1. Foredune Plant and Grass Distribution and Abundance Along the Outer Banks Foredunes
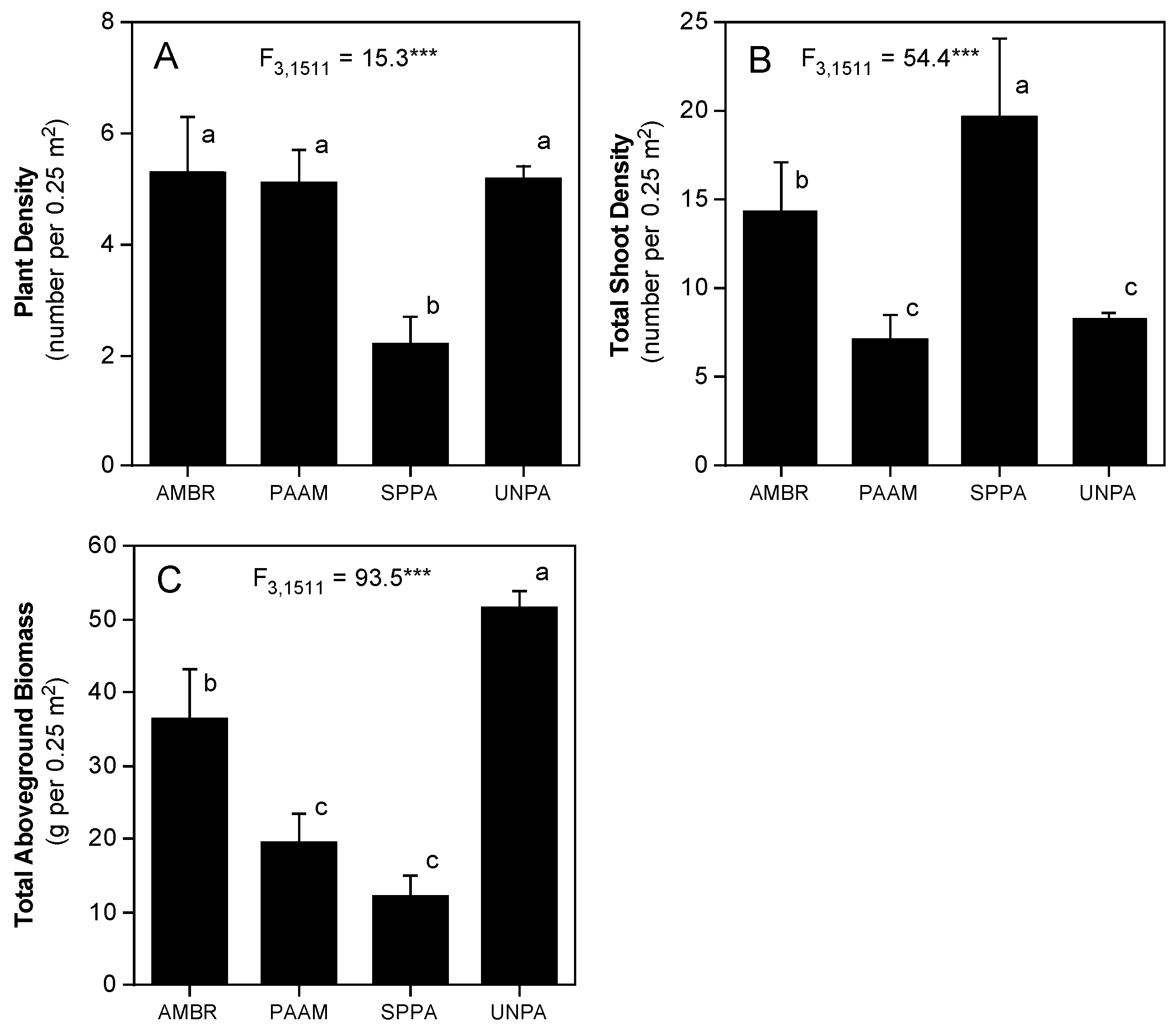
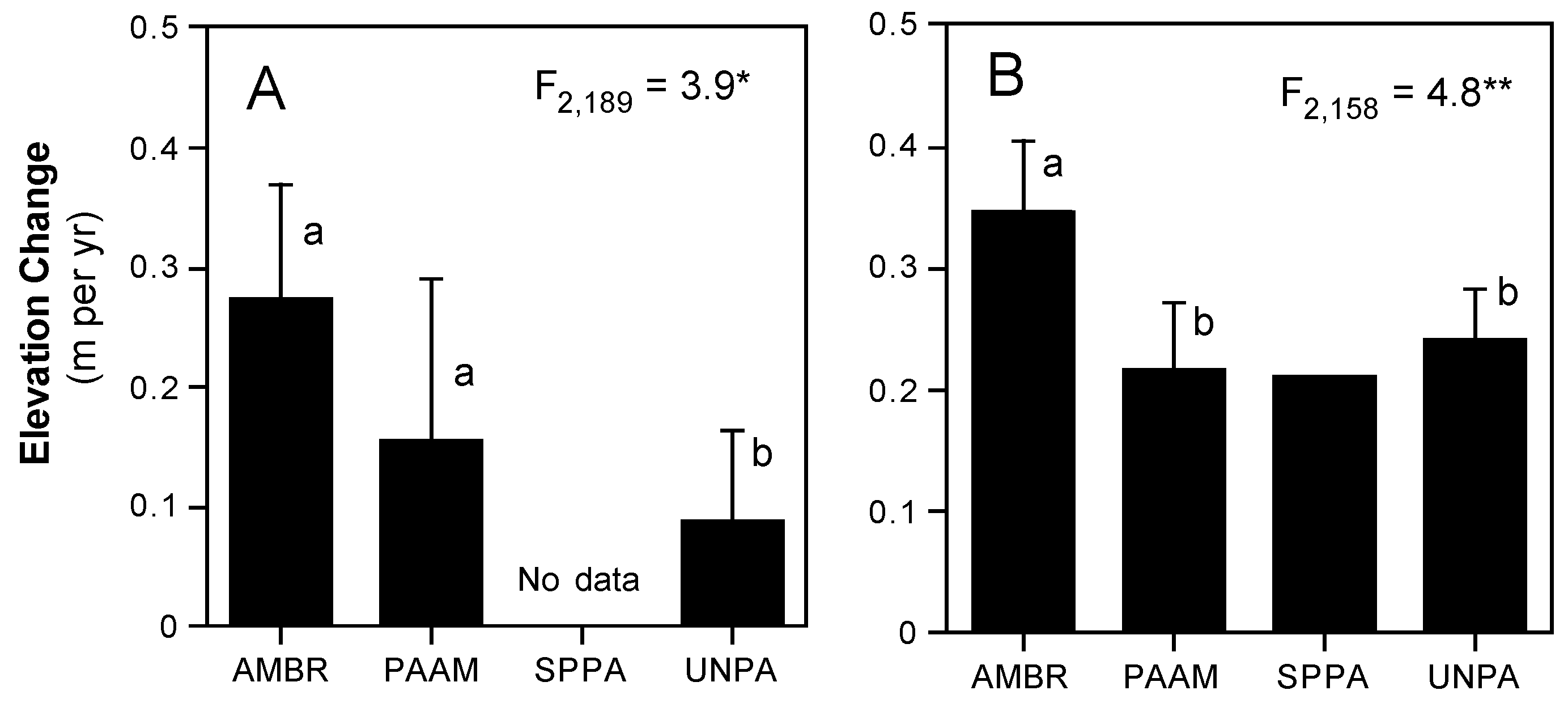
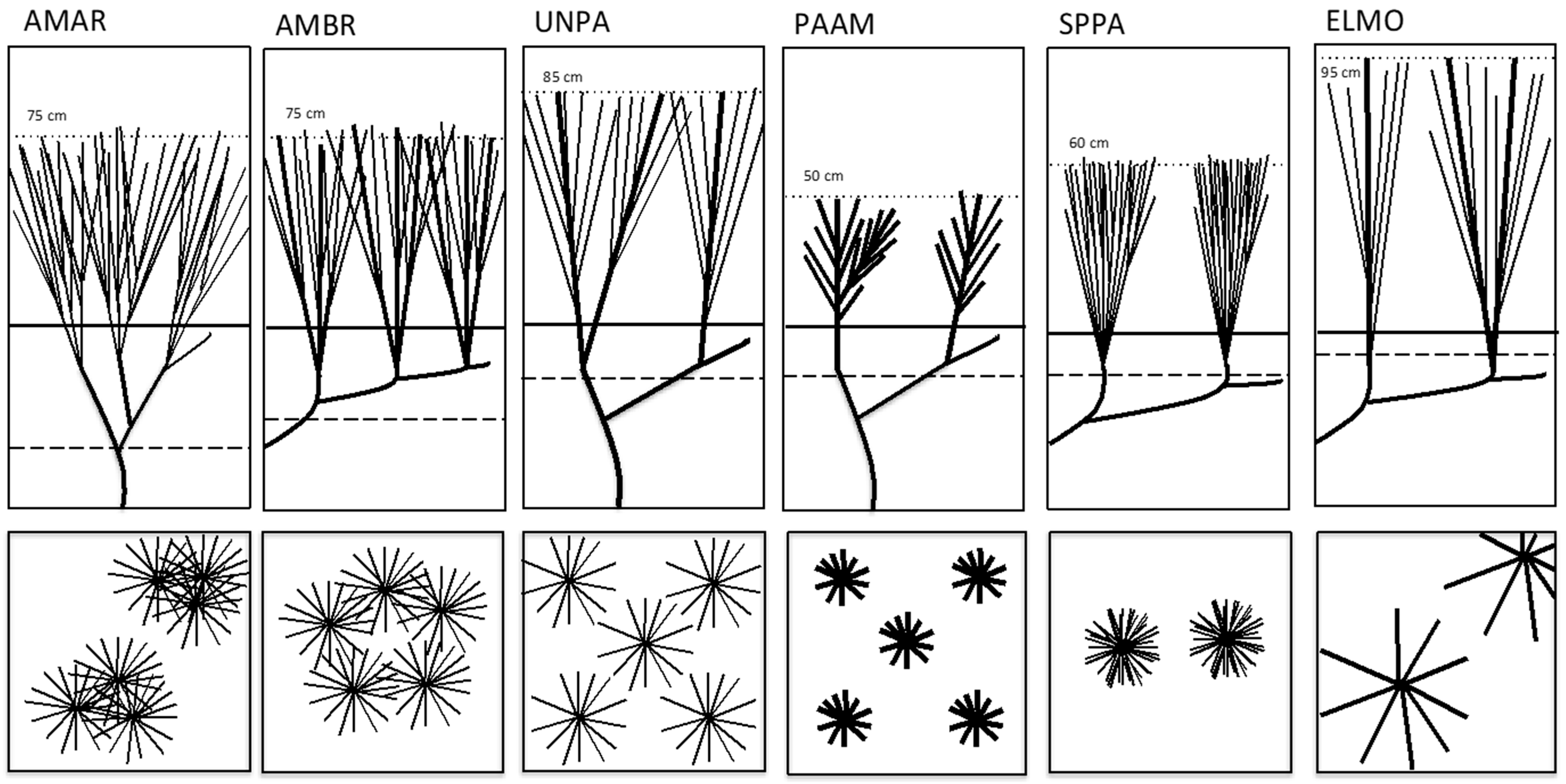
3.2. Dune Grass Growth Form and Morphology and Changes in Sand Elevation
4. Discussion
4.1. Species–Specific Functional Morphology and Associated Changes in Sand Elevation
4.2. Dune Grass Distributions and the Consequences for Foredune Geomorphology and Coastal Protection Along the Outer Banks
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Barbier, E.B.; Koch, E.W.; Silliman, B.; Hacker, S.D.; Wolanski, E.; Primavera, J.; Granek, E.; Polasky, S.; Aswani, S.; Cramer, L.A.; et al. Coastal ecosystem-based management with non-linear ecological functions and values. Science 2008, 319, 321–323. [Google Scholar] [CrossRef]
- Barbier, E.; Hacker, S.D.; Kennedy, C.; Koch, E.; Stier, A.D.; Silliman, B. The value of estuarine and coastal ecosystem services. Ecol. Monogr. 2011, 81, 169–193. [Google Scholar] [CrossRef]
- Olson, J.S. Lake Michigan dune development 2. Plants as agents and tools in geomorphology. J. Geol. 1958, 66, 345–351. [Google Scholar] [CrossRef]
- Woodhouse, W.W.; Seneca, E.D.; Broome, S.W. Effects of species on dune grass growth. Int. J. Biometeorol. 1977, 21, 256–266. [Google Scholar] [CrossRef]
- Hesp, P. A review of the biological and geomorphological processes in the initiation and development of incipient foredunes. Proc. R. Soc. Edin. 1989, 96, 191–202. [Google Scholar] [CrossRef]
- Duarte, C.M.; Losada, I.J.; Hendriks, I.E.; Mazarrasa, I.; Marba, N. The role of coastal plant communities for climate change mitigation and adaptation. Nat. Clim. Chang. 2013, 3. [Google Scholar] [CrossRef]
- Seabloom, E.W.; Ruggiero, P.; Hacker, S.D.; Mull, J.; Zarnetske, P.L. Invasive grasses, climate change, and exposure to storm-wave overtopping in coastal dune ecosystems. Glob. Chang. Biol. 2013, 19, 824–832. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, J.L.; Jones, C.G.; Byers, J.E.; Arkema, K.K.; Berkenbusch, K.; Committo, J.A.; Duarte, C.M.; Hacker, S.D.; Hogarth, P.J.; Lambrinos, J.G.; et al. Physical ecosystem engineers and the functioning of estuaries and coasts. In Functioning of Estuaries and Coastal Ecosystems; Heip, C.H.R., Philippart, C.J.M., Middelburg, J.J., Eds.; Elsevier: London, UK, 2011; Volume 7, pp. 53–81. [Google Scholar]
- McLeod, E.; Chmura, G.L.; Bouillon, S.; Salm, R.; Bjork, M.; Duarte, C.M.; Lovelock, C.E.; Schlesinger, W.H.; Silliman, B.R. A blueprint for blue carbon: Toward an improved understanding of the role of vegetated coastal habitats in sequestrating CO2. Front. Ecol. Environ. 2011, 9, 552–560. [Google Scholar] [CrossRef]
- Woodhouse, W.W. Dune Building and Stabilization with Vegetation; U.S. Army Corps of Engineers, Coastal Research Center: Fort Belvoir, VA, USA, 1978.
- Hacker, S.D.; Zarnetske, P.; Seabloom, E.; Ruggiero, P.; Mull, J.; Gerrity, S.; Jones, C. Subtle differences in two non-native congeneric beach grasses significantly affect their colonization, spread, and impact. Oikos 2012, 121, 138–148. [Google Scholar] [CrossRef]
- Zarnetske, P.L.; Hacker, S.D.; Seabloom, E.W.; Ruggiero, P.; Killian, J.R.; Maddux, T.B.; Cox, D. Biophysical feedback mediates effects of invasive grasses on coastal dune shape. Ecology 2012, 93, 1439–1450. [Google Scholar] [CrossRef] [PubMed]
- Zarnetske, P.L.; Ruggiero, P.; Hacker, S.D.; Seabloom, E.W. Coastal foredune evolution: The relative influence of vegetation and sand supply in the US Pacific Northwest. J. Roy. Soc. Interface 2015, 12. [Google Scholar] [CrossRef]
- Wolner, C.V.; Moore, L.J.; Young, D.R.; Brantley, S.T.; Bissett, S.N.; McBride, R.A. Ecomorphodynamic feedbacks and barrier island response to disturbance: Insights from the Virginia Barrier Islands, Mid-Atlantic Bight, USA. Geomorphology 2013, 199, 115–128. [Google Scholar] [CrossRef]
- Brantley, S.T.; Bissett, S.N.; Young, D.R.; Wolner, C.W.; Moore, L.J. Barrier island morphology and sediment characteristics affect the recovery of dune building grasses following storm-induced overwash. PLoS ONE 2014, 9, e104747. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.E. Effects of disturbance on vegetation by sand accretion and erosion across coastal dune habitats on a barrier island. AoB Plants 2015, 7, plv003. [Google Scholar] [CrossRef] [PubMed]
- Charbonneau, B.R.; Wnek, J.P.; Langley, J.A.; Lee, G.; Balsamo, R.A. Above vs. belowground plant biomass along a barrier island: Implications for dune stabilization. J. Environ. Manag. 2016, 182, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, E.B.; Moore, L.J.; Durán Vinent, O. Lateral vegetation growth rates exert control on coastal foredune “hummockiness” and coalescing time. Earth Surf. Dyn. 2017, 5, 417–427. [Google Scholar] [CrossRef]
- Biel, R.G.; Hacker, S.D.; Ruggiero, P. Elucidating coastal foredune ecomorphodynamics in the US Pacific Northwest via Bayesian networks. J. Geophys. Res. Earth 2019, in press. [Google Scholar]
- Psuty, N.P. A dune/beach interaction model and dune management. Thalasses 1986, 1, 11–15. [Google Scholar]
- Kuriyama, Y.; Mochizuki, N.; Nakashima, T. Influence of vegetation on Aeolian sand transport rate from a backshore to a foredune at Hasaki, Japan. Sedimentology 2005, 52, 1123–1132. [Google Scholar] [CrossRef]
- Durán, O.; Moore, L.J. Vegetation controls on the maximum size of coastal dunes. Proc. Natl. Acad. Sci. USA 2013. [Google Scholar] [CrossRef]
- Durán Vinent, O.; Moore, L.J. Bistability of barrier islands induced by biophysical interactions. Nat. Clim. Chang. 2015, 5, 158–162. [Google Scholar] [CrossRef]
- Charbonneau, B.R.; Casper, B.B. Wind tunnel tests inform Ammophila planting spacing for dune management. Shore Beach 2018, 86, 37–46. [Google Scholar]
- Buckley, R. The effect of sparse vegetation on the transport of dune sand by wind. Nature 1987, 325, 426–428. [Google Scholar] [CrossRef]
- Hesp, P. Foredunes and blowouts: Initiation, geomorphology and dynamics. Geomorphology 2002, 48, 245–268. [Google Scholar] [CrossRef]
- Van Dijk, P.M.; Arens, S.M.; van Boxel, J.H. Aeolian processes across transverse dunes. II. Modelling in the sediment transport and profile development. Earth Surf. Proc. Land. 1999, 24, 319–333. [Google Scholar] [CrossRef]
- Woodhouse, W.W.; Hanes, R.E. Dune Stabilization with Vegetation on the Outer Banks of North Carolina; U.S. Army Corps of Engineers, Coastal Research Center: Fort Belvoir, VA, USA, 1967.
- Seneca, E.D.; Woodhouse, W.W., Jr.; Broome, S.W. Dune Stabilization with Panicum amarum along the North Carolina Coast; U.S. Army Corps of Engineers, Coastal Research Center: Fort Belvoir, VA, USA, 1976.
- Goldstein, E.B.; Mullins, E.V.; Moore, L.J.; Biel, R.G.; Brown, J.K.; Hacker, S.D.; Jay, K.R.; Mostow, R.S.; Ruggiero, P.; Zinnert, J.C. Literature-based latitudinal distribution and possible range shifts of two US east coast dune grass species (Uniola paniculata and Ammophila breviligulata). PeerJ 2018, 6, e4932. [Google Scholar] [CrossRef]
- Bryant, M.A.; Hesser, T.J.; Jensen, R.E. Evaluation Statistics Computed for the Wave Information Studies (WIS); U.S. Army Engineer Research and Development Center Technical Note CHL I-91; U.S. Army Engineer Research and Development Center: Vicksburg, MS, USA, 2016.
- Miller, T.L.; Morton, R.A.; Sallenger, A.H. The National Assessment of Shoreline Change: A GIS Compilation of Vector Shorelines and Associated Shoreline Change Data for the U.S. Southeast Atlantic Coast; U.S. Geological Survey Open File Report No. 2005-1326; U.S. Geological Survey: Reston, VA, USA, 2006.
- Lazarus, E.D.; Murray, A.B. An integrated hypothesis for regional patterns of shoreline change along the Northern North Carolina Outer Banks, USA. Mar. Geol. 2011, 281, 85–90. [Google Scholar] [CrossRef]
- Armstrong, S.B.; Lazarus, E.D. Masked shoreline erosion at large spatial scales as a collective effect of beach nourishment. Earth’s Future 2019, 7, 74–84. [Google Scholar] [CrossRef]
- Kaczkowski, H.L.; Kana, T.W.; Traynum, S.B.; Visser, R. Beach-fill equilibration and dune growth at two large-scale nourishment sites. Ocean Dyn. 2018, 68, 1191–1206. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; Available online: http://ww.R-project.org/ (accessed on 15 January 2019).
- Mullins, E.; Moore, L.J.; Goldstein, E.B.; Jass, T.; Bruno, J.; Durán Vinent, O. Investigating dune-building feedback at the plant level: Insights from a multispecies field experiment. Earth Surf. Proc. Land. 2019. [Google Scholar] [CrossRef]
- Godfrey, P.J.; Godfrey, M.M. Barrier Island Ecology of Cape Lookout National Seashore and Vicinity, North Carolina; National Park Service Scientific Monograph Series; National Park Service: Cape Hatteras National Seashore, NC, USA, 1976.
- Stallins, J.A. Stability domains in barrier island dune systems. Ecol. Complex. 2005, 2, 410–430. [Google Scholar] [CrossRef]
- Seneca, E.D.; Cooper, A.W. Germination and seedling response to temperature, daylength, and salinity by Ammophila breviligulata from Michigan and North Carolina. Bot. Gaz. 1971, 132, 203–215. [Google Scholar] [CrossRef]
- Emery, S.M.; Rudgers, J.A. Biotic and abiotic predictors of ecosystem engineering traits of the dune building grass, Ammophila breviligulata. Ecosphere 2014, 5, 87. [Google Scholar] [CrossRef]
- Wagner, R.H. The ecology of Uniola paniculata L. in the dune-strand habitat of North Carolina. Ecol. Monogr. 1964, 34, 79–96. [Google Scholar] [CrossRef]
- Lonard, R.I.; Judd, F.W.; Stalter, R. Biological flora of coastal dunes and wetlands: Uniola paniculata L. J. Coast. Res. 2011, 27, 984–993. [Google Scholar] [CrossRef]
- Lonard, R.I.; Judd, F.W.; Stalter, R. The biological flora of coastal dunes and wetlands: Spartina patens (W. Aiton) GH Muhlenberg. J. Coast. Res. 2010, 26, 935–946. [Google Scholar] [CrossRef]
- Lonard, R.I.; Judd, F.W. The biological flora of coastal dunes and wetlands: Panicum amarum S. Elliott and Panicum amarum S. Elliott var. amarulum (AS Hitchcock and MA Chase) P. Palmer. J. Coast. Res. 2011, 27, 233–242. [Google Scholar]
- Harris, A.L.; Zinnert, J.C.; Young, D.R. Differential response of barrier island dune grasses to species interactions and burial. Plant Ecol. 2017, 218, 609–619. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.K.; Zinnert, J.C.; Young, D.R. Emergent interactions influence functional traits and success of dune building ecosystem engineers. J. Plant Ecol. 2018, 11, 524–532. [Google Scholar] [CrossRef]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hacker, S.D.; Jay, K.R.; Cohn, N.; Goldstein, E.B.; Hovenga, P.A.; Itzkin, M.; Moore, L.J.; Mostow, R.S.; Mullins, E.V.; Ruggiero, P. Species-Specific Functional Morphology of Four US Atlantic Coast Dune Grasses: Biogeographic Implications for Dune Shape and Coastal Protection. Diversity 2019, 11, 82. https://0-doi-org.brum.beds.ac.uk/10.3390/d11050082
Hacker SD, Jay KR, Cohn N, Goldstein EB, Hovenga PA, Itzkin M, Moore LJ, Mostow RS, Mullins EV, Ruggiero P. Species-Specific Functional Morphology of Four US Atlantic Coast Dune Grasses: Biogeographic Implications for Dune Shape and Coastal Protection. Diversity. 2019; 11(5):82. https://0-doi-org.brum.beds.ac.uk/10.3390/d11050082
Chicago/Turabian StyleHacker, Sally D., Katya R. Jay, Nicholas Cohn, Evan B. Goldstein, Paige A. Hovenga, Michael Itzkin, Laura J. Moore, Rebecca S. Mostow, Elsemarie V. Mullins, and Peter Ruggiero. 2019. "Species-Specific Functional Morphology of Four US Atlantic Coast Dune Grasses: Biogeographic Implications for Dune Shape and Coastal Protection" Diversity 11, no. 5: 82. https://0-doi-org.brum.beds.ac.uk/10.3390/d11050082