Inventory and Historical Changes in the Marine Flora of Tomioka Peninsula (Amakusa Island), Japan

1
National Scientific Center of Marine Biology, Far Eastern Branch, Russian Academy of Science, A.V. Zhirmunsky Institute of Marine Biology, Palchevskogo 17, Vladivostok 690041, Russia
2
Amakusa Marine Biological Laboratory, AMBL-Kyushu University, Reihoku-Amakusa, Kumamoto 863-2507, Japan
3
State Key Laboratory of Marine Resource Utilization in South China Sea, College of Marine Science, Hainan University, 58th Renmin Avenue, Haikou 570228, China
*
Author to whom correspondence should be addressed.
Diversity 2019, 11(9), 158; https://0-doi-org.brum.beds.ac.uk/10.3390/d11090158
Submission received: 4 July 2019
/
Revised: 30 August 2019
/
Accepted: 3 September 2019
/
Published: 6 September 2019
(This article belongs to the Special Issue Biodiversity of Macroalgae)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Intensive algal sampling was conducted from 2012 to 2017 in the Tomioka Peninsula, Amakusa-Shimoshima Island (the East China Sea, Japan), yielding a total of 293 benthic macroalgal taxa, of which 63% were red algae, 16% were brown algae, and 19% were green algae. The majority (69%) of species were previously recorded only for the tropics and/or subtropics, whereas 31% of species were recorded for temperate latitudes. Among all species of algae found from 2012 to 2017 in the Tomioka Peninsula, 163 species (56%) were newly identified species for Amakusa-Shimoshima Island, including six taxa, which were recorded in Japan for the first time. Comparison of the current data from the Amakusa-Shimoshima Island with those of nearby tropical regions suggested that the recent marine flora of the Amakusa-Shimoshima Island was more closely affiliated with the flora of a warm-temperate region. Moreover, we found that the benthic flora of the Tomioka Peninsula was significantly changed between the 1950s (Segawa & Yoshida 1961) and 2012–2017. For example, the species diversity was increased by two times, mainly at the expense of red and green algae, and the biogeographic status of the benthic flora was changed from the flora of a cold-temperate region to the flora of a warm-temperate region, which could be attributed to an introduction of red algae from the tropics of South East Asia and widespread opportunistic green algae, as well as the disappearance of cold-water brown algae. Collectively, our data suggested that these changes were driven by the global warming effect on the ocean.
1. Introduction
Amakusa-Shimoshima Island (32°25’ N, 130°7’ E) is located at the north-eastern periphery of the East China Sea, 22 kilometers off the west coast of Kyushu Island (Figure 1A). The island has an area of ~400 km2. This region has a humid subtropical climate with the highest summer temperature of 32 °C in August and the lowest temperature of below 5 °C in winter months. Precipitation is significant throughout the year, and its annual rainfall reaches ~2800 mm, with the heaviest precipitation in June and July. A relatively cold period lasts from approximately December to March. The water temperature usually fluctuates between 13 °C and 30 °C during a year. Generally speaking, the salinity of this area is above 30 practical salinity units (PSU), and it can drop to 14 PSU during the rainy season [1,2].
The benthic flora of the island has not been well documented. Only one algal sampling to study species diversity and composition around the Tomioka Peninsula (32°31’19.1’’ N, 130°1’56’’ E) and near Amakusa Marine Biological Laboratory, Kyushu University (32°31’9’’ N, 130°2’34.7’’ E) was conducted in 1959–1960 by Segawa & Yoshida [3]. In the subsequent years, several non-taxonomic investigations concerning the marine algae of the Tomioka Peninsula and other areas of the Amakusa-Shimoshima Island have been published [4,5,6,7].
From 2012 to 2017, we assessed the diversity of seaweeds, taxonomic composition, and the structure of algal communities around the Tomioka Peninsula at the same sites, where Segawa and Yoshida [3] conducted their field work. In the present study, we aimed to fulfill an inventory of the recent marine flora around the Tomioka Peninsula, document possible changes in the marine flora in this area during the past 50 years, and reveal the possible causes underlying these changes.
2. Materials and Methods
2.1. Research Area
Investigations on marine flora were conducted in the north-western corner of the Amakusa-Shimoshima Island around the Tomioka Peninsula (Figure 1A). The rocky-cobble shores of the northern, west and south-west coastline are exposed to wave action of the East China Sea, which are not urbanized, and the east coast faces the Tomioka Bay with a sandy shore. The neck of the peninsula is occupied by hotels, residential houses, and a power plant [8]. In Tomioka Bay, there is a small fishing port and some passenger berths, through which the Amakusa Island and Hiroshima City are connected. Moreover, Tomioka Bay is also the terminal, where loose construction materials are discharged. In addition, some farms for the aquaculture of molluscs and marine plantations, such as green alga Monostroma nitidum, are also located in the bay. The inner part of the bay is completely protected from storms, whereas the coast opposite a bay outlet is exposed to wave action.
2.2. Study Sites
A total of six sampling locations were selected as follows. (1) Akaiwa (Figure 1A, sampling points 4, 5, 6) was characterized by stony substrates in the intertidal zone and rocky substrates in the upper subtidal zone (Figure 1B, sampling point 4). (2) Magarizaki (Figure 1A,C, sampling point 7) was characterized by large boulders from the intertidal to upper subtidal zones. (3) The intertidal and upper subtidal zones of Shikizaki Bay (Figure 1A,D, sampling point 3) were characterized by rocky substrates with many intertidal pools and some large subtidal pools with sandy bottoms and separate rocks protruding among the sand (Figure 1D, sampling point 3). (4) Shiraiwazaki Bay (Figure 1A, sampling points 1, 2) was characterized by rocky and stony substrates with pebbles and boulders (Figure 1E, sampling point 2) as well as an artificial underwater embankment and breakwater structures along the coast (sampling point 1). In open areas of the coast, the intertidal zone was characterized by pools of various depths, and the upper subtidal zone was characterized by rocky substrates (rocky outcrops among sand) with separate colonies of hermatypic corals [5,8]. In artificially created areas in the lower intertidal and upper subtidal zones, the bottom was represented by silted sand and individual large stones. (5) At Tomioka Harbor (Figure 1A, sampling points 8–12), algae were collected from three sites with artificial substrates (concrete and concrete-stony walls of piers, concrete slabs, metal and plastic, mooring-lines, and so on) and at sites outside the port with natural substrates (rocky, stony, and sandy bottoms covered with silt). The intertidal and upper subtidal zones of the harbor were characterized by a silt–sand mixture with rocky outcrops (Figure 1F, sampling points 9, 11). (6) Tsujishima Islet located to the north-east of Tomioka Harbor (Figure 1A, sampling points 13, 14) was characterized by rocky substrates in the intertidal zone and upper subtidal zone (Figure 1G, sampling point 14).
2.3. Sampling Timing and Strategy
Macroalgal sampling was conducted in November–December 2012, April and August 2013, January 2014, October–November 2015 and November 2017. At each sampling point, algae were collected at a water depth of 5 m from four different tidal zones as follows: the upper intertidal zone (+ the splash zone), middle and lower intertidal zones, as well as the upper subtidal zone (division into tidal zones followed) [9] (Table S1).
2.4. Collection, Identification and Conservation of Marine Plants
Marine plants at every sampling point were collected from 0–5 m depth via foot or snorkeling during low tides and SCUBA diving during high tides. In order to study the species composition of the benthic flora and the taxonomic composition of algal communities, the detailed algal sampling and the processing of the obtained materials were as follows (Table S1):
- (1)
- In each tidal zone, algal turf communities (with thalli less than 5 cm in height), crustose coralline algae (CCA) and large upright-growing algae (with thalli more than 5 cm in height) in an area of ~100 m2 were visually identified. The selected communities were photographed many times at a right angle. In the communities of algal turf and CCA, samples were collected from three randomly selected square plots, each plot measuring ~100 cm2. In communities of upright-growing algae, samples were also collected from three squares, each measuring from 0.5 m2 to 1 m2. Algae were also separately collected from areas outside the selected square plots.
- (2)
- Algae collected from different communities were stored in separate plastic bags and then placed in the refrigerator prior to further processing. Freshly collected material was identified using monographic publications, floristic studies, and systematic articles described by Titlyanova et al. [10]. The systematics and nomenclature followed Guiry & Guiry [11]. The previously known and newly recorded species for Amakusa-Shimoshima Island were verified using the article by Segawa & Yoshida [3] and for Japan by AlgaeBase [11]. Hierarchical classification of the Phylum Rhodophyta (Rh) was carried out as Saunders & Hommersand [12]. The classification system of the Phyla Chlorophyta (Ch) and Ochrophyta followed Tsuda [13].
- (3)
- The collections of both macrophytes and their epiphytes were preserved as dried herbarial specimens and deposited in the herbarium at the A.V. Zhirmunsky Institute of Marine Biology, National Scientific Center of Marine Biology, Far Eastern Branch, Russian Academy of Science, Vladivostok 690041, Russian Federation.
During our work at the Amakusa-Shimoshima Island, a total of more than 600 samples from algal turf communities and more than 300 samples from communities of large upright-growing algae were processed (Table S1). Abundance was visually determined, and the mean substrate surface area occupied by marine plants (relative percentage of coverage) was estimated using slides of algal turf communities. The following codes were used: (+) rare sighting—overall representatives of a certain species were found only one–two times with less than 5% relative coverage of substrata; (++) common—representatives of the species were found in the most samples with 5% to 20% relative coverage; (+++)abundant—algae were found in communities with 30% to 100% relative coverage. Dominance in the communities was also visually determined and defined as monodominant if one algal species occupied more than 50% of the surface area; bidominant if two species occupied more than 50% of the surface area; and polydominant if more than two species predominated.
2.5. Terms of the Comparative Analysis, when Studying Historical Changes in the Flora
Under the comparison of two collections of marine plants sampled by different collectors, the question arises about correctness of the comparison, because of possible differences in aims of studies, quality and quantity, season, and sites of samplings. In previous studies on decadal changes in the benthic flora, we postulated four items within a framework promoting success in the comparison of earlier and later collections: (1) collections of algae should be sampled in the same season of year or in all seasons; (2) algal sampling should be carried out at sites with similar geographic and zonal features; (3) assessment of species diversity of the marine flora should be the aim of algal sampling; (4) collections should be compared using the same sampling methods [14]. Through the comparison of the early collection by Segawa and Yoshida (the 1950s, early collection, EC) [3] with our new collection (2012–2017, late collection, LC), these items were kept.
3. Results
3.1. Species and Life Form Diversity
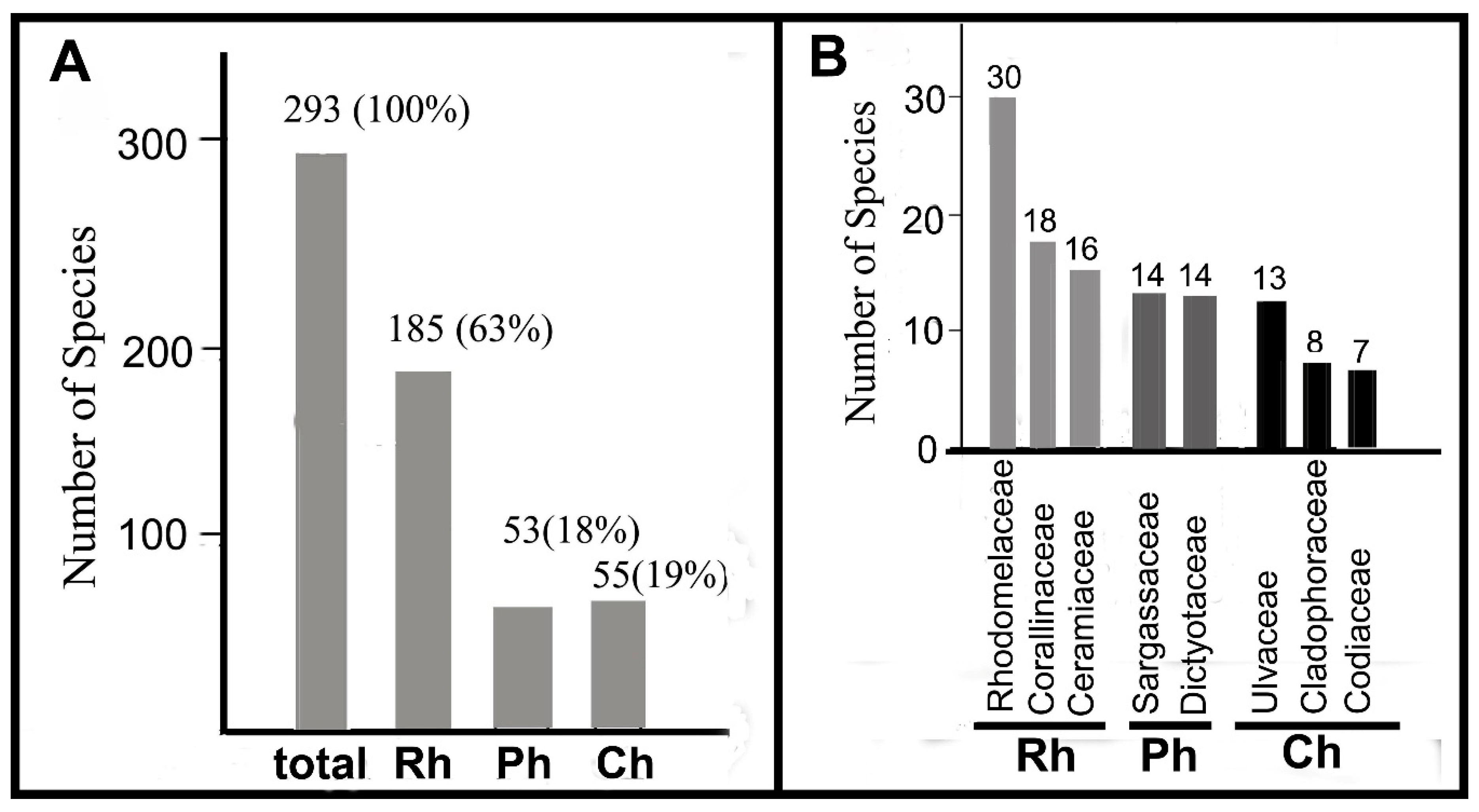
Table S2 and Figure 2, Figure 3, Figure 4, Figure 5, Figure 6 and Figure 7 show the results of the present study, including 293 species of benthic macroalgae. The Phylum Rhodophyta consisted of four classes, 17 orders, 38 families, 87 genera, and 185 species (63% of all species). Ochrophyta was composed of one class Phaeophyceae (Ph), nine orders, 12 families, 25 genera, and 53 species (18%); and Chlorophyta consisted of one class (Ulvophyceae), five orders, 16 families, 25 genera and 55 species (19%). Among Rhodophyta, the largest taxa belonged to the Rhodomelaceae, Corallinaceae, and Ceramiaceae; among brown algae Sargassaceae, Dictyotaceae; and among Chlorophyta it belonged to Ulvaceae, Cladophoraceae, and Codiaceae (Figure 2). In terms of algal species richness, the sampling localities could be ranked as follows: Shiraiwazaki Bay > Tomioka Harbor > Shikizaki Bay > Tsujishima Islet > Akaiwa > Magarizaki (106) (Tables S1 and S2).
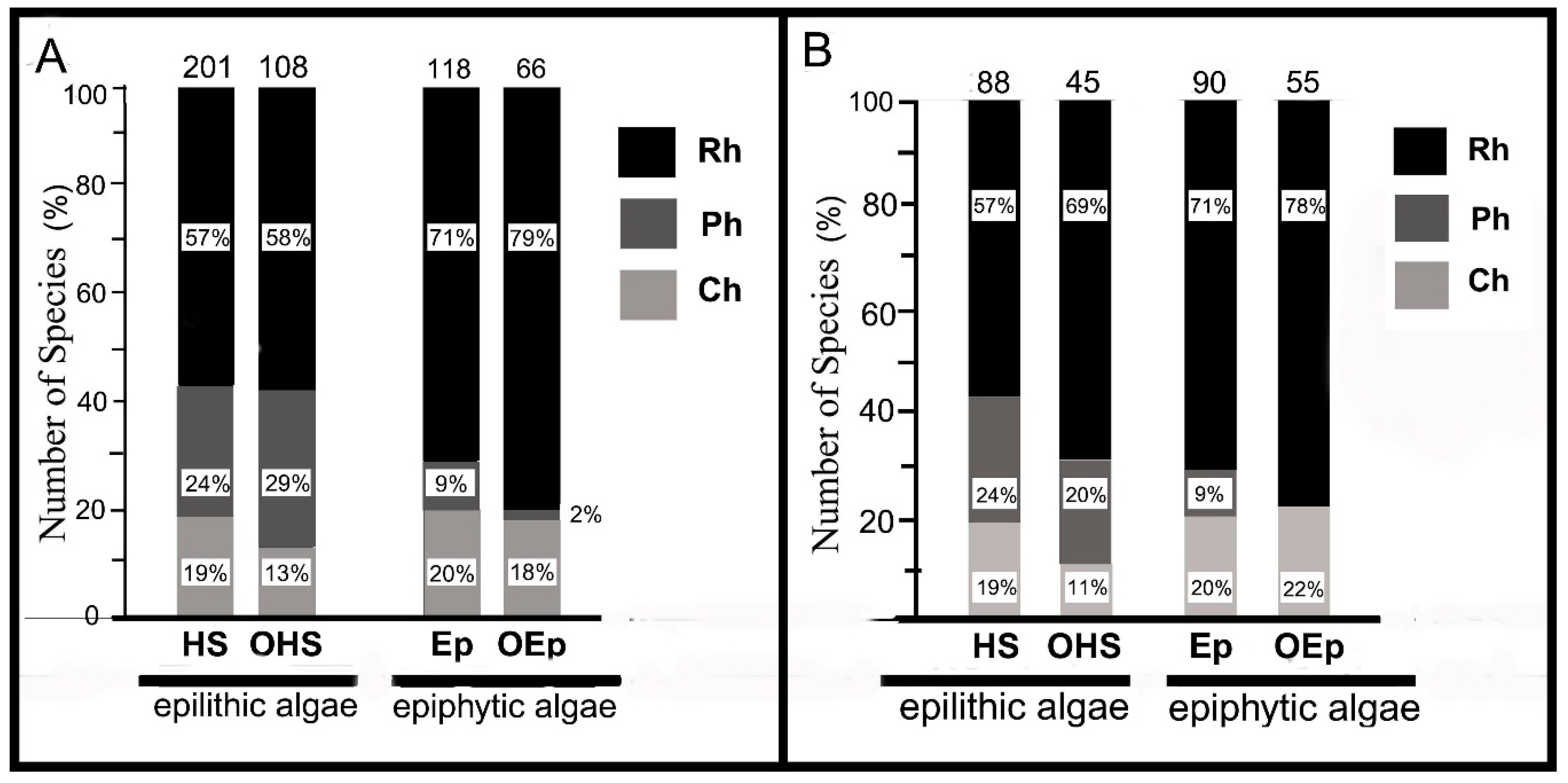
The investigated area was dominated by algae growing on solid natural substrates (epilithic algae) (201 species), and many of epilithic algae (20%) also grew epiphytically. A total of 108 species of algae grew only on hard substrates, among which red and brown algae predominated (Figure 3A).
A total of 118 epiphytes were identified, and half of them also grew on solid natural and/or artificial substrates. Moreover, 66 species (or 23% of all collected species) only epiphytically grew. Red and green algae predominated among these epiphytes (Figure 3A).
A total of 69 species (24%) grew on various artificial substrates, including concrete blocks, plastic ropes, metal and wooden constructions of the port. In addition, 31 species grew on shells of live mollusks (epizoic algae), and three species (Pseudocladophora conchopheria, Champia japonica, and Polysiphonia senticulosa) were observed only on shells. Two collected species, Gracilaria parvispora and Agarophyton vermiculophyllum, being only inhabitants of soft substrates (epipelithic algae) were attached to small pebbles. Three species, Acrochaete geniculata, Ulvella viridis (Ch), and Acrochaetium colaconemoides (Rh), were found growing endophytically. The green alga Percursaria percursa were found only floating on the water surface (Table S2).
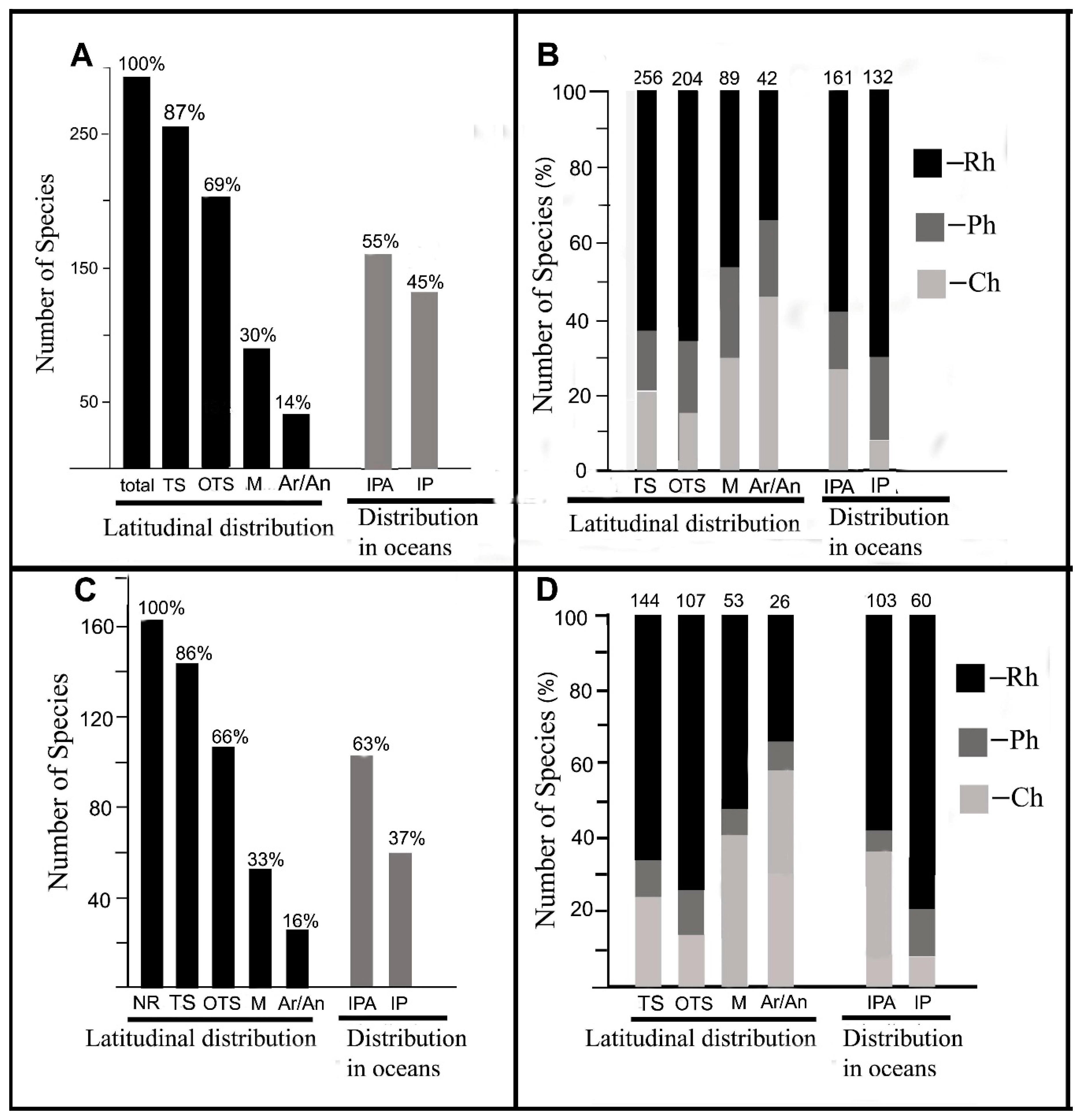
In the collection of 2012–2017, the dominated group of species (256 species, or 87%) was found earlier in the tropics and subtropics (Guiry & Guiry 2018), and the rest 13% species were recorded for the subtropics. A total of 204 species (69%) were recorded only for the tropics and/or subtropics, among which red algae accounted for 67%, brown algae accounted for 19%, and green algae accounted for 14%. Algae recorded for the tropics and/or subtropics to temperate zones included 89 species (30% of the collection). In this group, red algae accounted for 53%, brown algae accounted for 17%, and green algae accounted for 30%. Cosmopolitan species inhabiting from the tropics to the Arctic/Antarctic zones consisted of 42 species (14% of the collection), where green algae predominated (Table S2, Figure 4A,B).
Algae widely distributed in the tropical and subtropical zones of Atlantic, Indian, and Pacific Oceans accounted for more than half of our collection, of which red, brown, and green algae accounted for 58%, 24%, and 27%, respectively. Algae inhabiting only the Indo-Pacific contained 132 species, of which green algae predominated (Table S2, Figure 4A,B).
3.2. Newly Recorded Species for Amakusa-Shimoshima Island
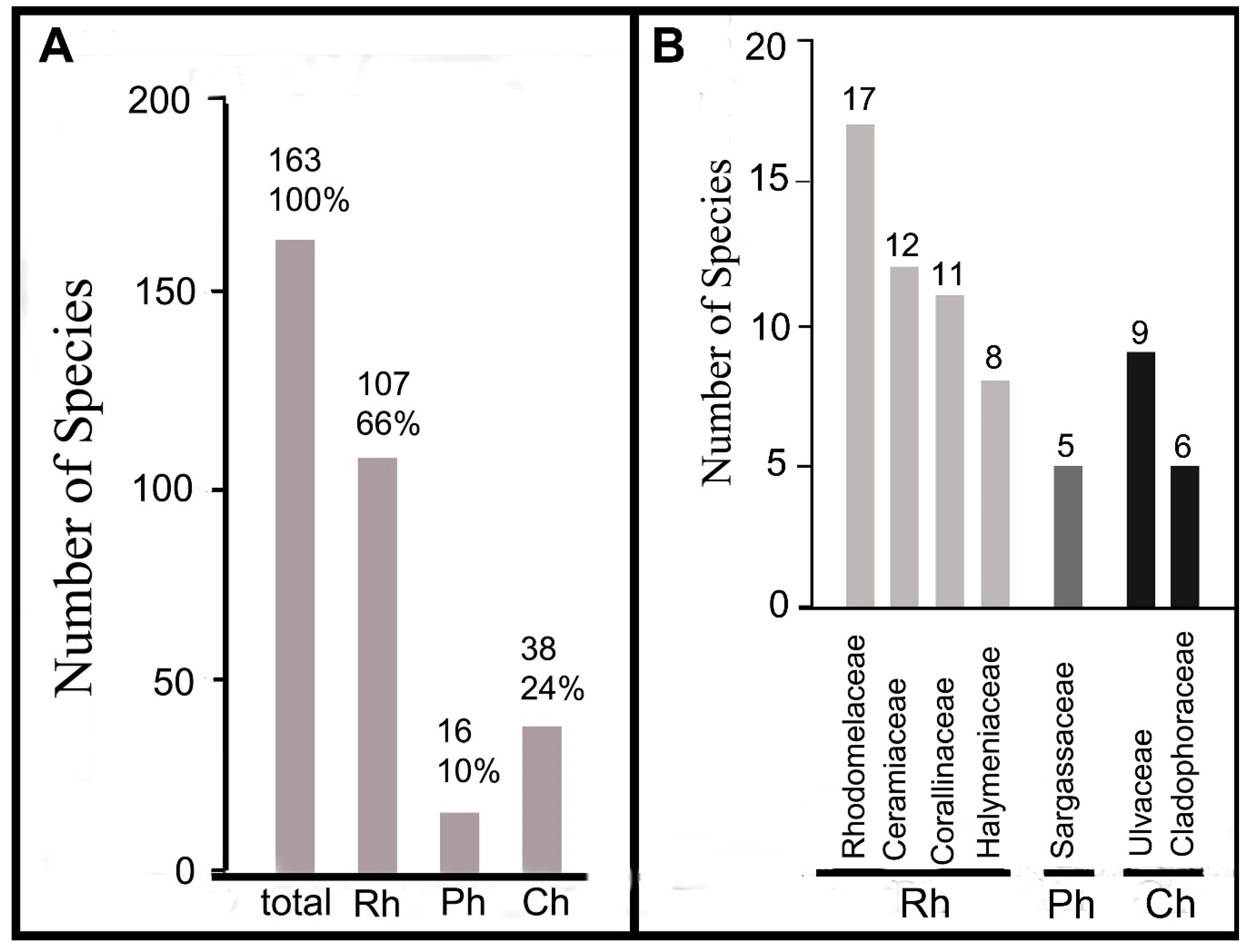
Among all species of algae found in 2012–2017 around the Tomioka Peninsula, 163 taxa (56% of all found algae) were new records for Amakusa. The largest number of new records was Rhodophytes (66% of all found species). Most taxa belonged to Rhodomelaceae, Ceramiaceae, Corallinaceae, and Halymeniaceae, and brown algae accounted for 10% of new records. The greatest number of taxa belonged to Sargassaceae. Green algae accounted for 24% of new records, and the greatest number of taxa belonged to Ulvaceae and Cladophoraceae (Table S2, Figure 5A,B).
In the new records (NR) group, epiphytic algae predominated (90 species, or 55% of NR), and many species of this group also occupied hard substrata, artificial substrata, and shells of mollusks. Moreover, 55 algal species (34% NR) grew only epiphytically. Among the “pure” epiphytes, red algae predominated. In addition, 45 species (27% NR) grew only on natural hard substrata, and red algae predominated among these algae and epiphytes (Figure 3B).
In the group of newly recorded species (144 species or 86% of NR), as well as in all collections of dominated species inhabiting tropical and subtropical seas, the remaining 14% of the NR were not recorded for tropical waters. A group of algae inhabiting only tropical and subtropical waters consisted of 107 species or 66% of NR, of which red, brown, and green algae accounted for 75%, 11%, and 14%, respectively. A group of NR algae recorded from the tropics and/or subtropics to temperate zones consisted of 53 species or 33% of the NR, of which the relative proportion of red, brown, and green algae was 53%, 7%, and 40%, respectively. Cosmopolitan species of the NR collection widely spread from the tropical to Arctic/Antarctic zones accounted for 26 species or 16% of NR, of which green algae predominated (Table S2, Figure 4C,D).
A group of NR species distributed in the tropical and subtropical zones of three oceans (Atlantic, Indian, and Pacific Oceans) included 103 taxa (63% of NR), including 58% red algae, 15% brown algae, and 27% green algae. Algae inhabiting the Indo-Pacific included 60 species (37% of NR), and the relative proportion of red, brown, and green algae in this group was 80%, 12%, and 8%, respectively (Table S2, Figure 4C,D).
3.3. Newly Recorded Species for Japan
Six species of our collection were new records to Japan. The endophytic alga Acrochaete geniculata (N.L. Gardner) O’Kelly (Ch) was found between outer cells of Hypnea sp. growing in the lower intertidal and upper subtidal zones in Shikizaki Bay and Ceramium borneense from Tsujishima Islet. The species has been previously found in Kurile Islands and Sakhalin Island as Pseudodictyon geniculatum Gardner [15] and in Hainan Island [16] (Figure 6A).
Antithamnionella longicellulata Perestenko (Rh) growing as an epiphyte on Ceramium cimbricum was found in the low intertidal zone of Tomioka Bay. This species was also recorded from Peter the Great Bay, the Sea of Japan and Busse Lagoon, and South Sakhalin [9] (Figure 6B).
Melanothamnus pseudovillum (Hollenberg) Díaz-Tapia & Maggs (Rh) was found growing as an epiphye on Colpomenia sinuosa in low intertidal zone in Tomioka Bay. This alga is distributed in Florida, Atlantic Ocean [17], Johnston Island, Central Pacific [11] and also found in Hainan Island, China [18] (Figure 6C).
Chaetomorpha javanica Kützing (Ch) was found on high-level rocks in Shikizaki Bay. The species is distributed in the tropics and subtropics of the Atlantic, Indian, and Pacific Oceans (Figure 6D).
Ulva chaetomorphoides (Börgesen) Hayden, Blomster, Maggs, P.C. Silva, M.J. Stanhope & J.R. Waaland (Ch) was found on hard substratum in the lower intertidal zone in Shiraiwazaki Bay. This species was distributed in the tropics and subtropics of the Atlantic, Indian, and Pacific Oceans (Figure 6E).
Ulva ralfsii (Harvey) Le Jolis (Ch) was found on rocks in the middle intertidal zone of Tsujishima Islet. The alga was distributed from the tropical to temperate latitudes of the Atlantic, Indian, and Pacific Oceans (Figure 6F).
3.4. Distribution of Algal Species and their Communities in Tidal Zones
The distribution of algae in tidal zones was uneven, and the largest number of species (218) was found in the lower intertidal zone, followed by 173 species in the upper subtidal zone and only 61 species in the upper and middle intertidal zones. Moreover, half of the species were found in the intertidal pools. Nine species of the red algae, such as Chondracanthus intermedius, Chroodactylon ornatum, Erythrotrichia carnea, Gayliella mazoyerae, Herposiphonia secunda, H. secunda f. tenella, Hydrolithon farinosum, Sahlingia subintegra, Stylonema alsidii, the brown alga Sphacelaria rigidula, and three species of green algae, such as Ulva clathrata, U. compressa, Rhizoclonium riparium, were found in all tidal zones (Table S2).
In the upper intertidal zone, rocky substrates were dominated by monodominant communities of brown crust algae, such as Neoralfsia expansa (Figure S1A), Ralfsia verrucosa and Petrospongium rugosum, and the red alga Pyropia [Porphyra] suborbiculata. The middle intertidal zone was occupied by monodominant (rarely bidominant) communities of the brown alga Ishige foliacea, the green alga Monostroma nitidum (Figure S1B) and the red alga Gloiopeltis furcata. Moreover, N. expansa, P. rugosum, Feldmannia mitchelliae (Ph), Corallina pilulifera, C. intermedius, Centroceras clavulatum (Rh) and Ulva intestinalis (Ch) were often found among the communities.
In the low intertidal zone, monodominant communities of the brown algae Ishige okamurae or Sargassum thunbergii (Figure S1C) and a mosaic polydominant community composed of C. undulatus, I. okamurae, Colpomenia sinuosa, Ulva lactuca, and Cladophora laetevirens predominated on rocky substrata (Figure S1D). Among sandy substrata, Agarophyton vermiculophyllum and Gracilaria parvispora were found on separate pebbles.
In the upper subtidal zone, the abundant species included Amphiroa beauvoisii, C. clavulatum, Gelidium amansii, and G. elegans (Rh); Dictyopteris prolifera, Dictyota coriacea, C. sinuosa, Sargassum hemiphyllum, S. horneri, S. piluliferum (Oc); Caulerpa webbiana f. tomentella and C. subserrata (Ch). Polydominant communities were formed with mosaic inclusions of algae, such as Amphiroa beauvoisii, Gelidium amansii, and G. elegans (Rh); C. sinuosa, Dictyota coriacea (Oc); C. webbiana f. tomentella and C. subserrata (Ch) in the upper subtidal zone (Figure S1E). Sargassaceae were common in these communities. Dictyopteris prolifera formed a monodominant community with a negligible inclusion of other epilithic algae (Figure S1F). In this zone, the largest number of epiphytic algae included red algae, such as Acrochaetium microscopicum, Colaconema hypneae, C. clavulatum, Ceramium borneense, C. cimbricum, C. tenerrimum, Crouania attenuate, and Gayliella fimbriata.
Only at Shiraiwazaki Bay, the upper subtidal zone at a depth of 3–4 m was occupied by hermatypic corals and algae-macrophytes, including Sargassum yamadae, S. piluliferum, Zonaria diesingiana, Dictyopteris prolifera, Dictyota coriacea, D. friabilis (Ph), Plocamium telfairiae, Chondracanthus tenellus, Gelidium amansii, and G. elegans (Rh).
4. Discussion
4.1. Recent Seaweed Flora
According to the scheme of Briggs [19] on marine biogeographic regions, Amakusa Island is situated on the northern border of warm-temperate region of the Northern Hemisphere, as well as Kyushu and Shikoku Islands.
The marine flora of cold-temperate, warm-temperate, and tropical regions in both hemispheres differs in species diversity and composition of large taxonomic algal groups (Rhodophyta, Phaeophyceaceae, and Chlorophyta) [20,21]. Unfortunately, there are no generalized data on the diversity of the recent benthic algal flora along the coasts of East Asia. However, based on past studies covering this region [9,10,16,22,23,24,25,26,27], it can be summarized that an average of about 200 common species of benthic macroalgae are found in cold-temperate region, and more than 500 species are found in warm-temperate and tropical seas of East Asia.
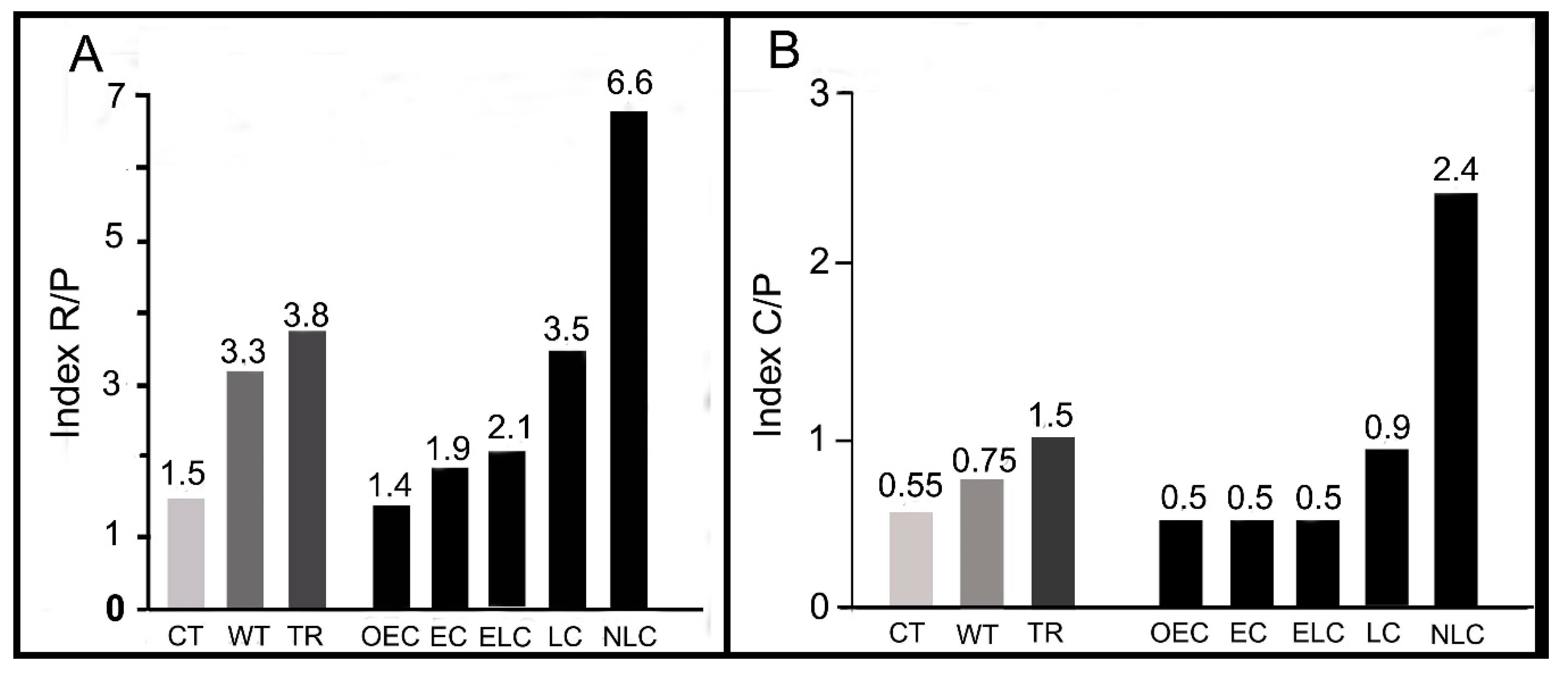
The indices R/P (number of Rhodophyta species divided by the number of Phaeophycean species) and C/P (number of Chlorophyta species divided by the number of Phaeophycean species) in a given flora are very important characteristics of the marine flora in the biogeographic regions [20,21].
According to Santelices et al. [20], these indices for the marine flora of cold waters (~45°N) are 1.0–2.0 and 0.3–0.8, respectively, these indices for warm waters (~30° N) are 2.5–4.0 and 0.5–1.0, respectively, and the values for tropical waters (~10° N) are 3.0–4.5 and 1.0–2.0, respectively (Figure 7). The marine floras of cold waters, both in the northern and southern hemispheres, are dominated by red and brown algae, and the floras of warm waters are dominated by red and green algae. In the marine floras of cold waters, red, brown, and green algae account for 40–50%, 20–25%, and 10–20%, respectively [24,26]. In the marine floras of warm waters (subtropical and tropical latitudes), red, brown, and green algae account for 50–70%, 10–15%, and 20–30%, respectively [10,14,22,23,27]. The main characteristic of the benthic marine floras in the East Asia warm-temperate region is the presence of the warm-water genus Sargassum (S. fusiforme, S. horneri, S. thunbergii, etc.) in the intertidal and subtidal zones. Other dominant species in the subtidal zones of cold- and warm-temperate regions in the northern part of the East China Sea and the Sea of Japan include brown algae, such as Undaria pinnatifida, Eisenia bicyclis, and Ecklonia cava, and also common red algae, such as Nemalion vermiculare and Corallina pilulifera [26,28,29,30].
Our sampling in 2012–2017 along the Tomioka Peninsula coast of Amakusa-Shimoshima Island documented 293 species of benthic macroalgae. Ratios of species numbers of algal taxonomic groups were R/P = 3.5 and C/P = 0.9 (Figure 7). The taxonomic compositions of large taxonomic groups were as follows: Rh 63%, Ph 18%, and Ch 19% (Figure 2A). These basic characteristics of the benthic floras of Tomioka Peninsula in 2012–2017 indicated the close association between the floras of warm temperate regions [20].
However, in terms of parameters—such as a high level of species diversity, the predominance in the floras by the number of species and biomass of red algae from the families Rhodomelaceae and Ceramiaceae, brown algae from Sargassaceae, and Dictyotaceae and green algae from families Ulvaceae and Cladophoraceae, the formation on hard substrates of bidominant and polydominant algal turf communities, a high degree of epiphyticism, and the species distribution and their communities in tidal zones—the floras of the Tomioka Peninsula (Amakusa-Shimoshima Island) demonstrated similarity with those collected from some localities in South Asia tropical biogeographical regions, such as southern Vietnam, Nha Trang Bay, 20° S of the Tomioka Peninsula; Hainan Island, China (Sanya Bay, 14° S); and Japan, Yonaguni Island, 8° S [14,31,32].
4.2. Historical Changes in the Marine Flora
Special interest represents the comparison of the recent flora of the Tomioka Peninsula with earlier flora (the 1950s) documented by Segawa and Yoshida [3] and analysis of changes occurring during the past 50 years. We have previously studied decadal changes in the benthic flora of three localities in the tropic biogeographic region of the South-East Asia, including Southern Vietnam, Nha Trang Bay; China, Hainan Island, Sanya Bay; and Japan, Yonaguni Island [14,31,32]. In all these localities, we have found serious changes and even catastrophic changes in some areas which could be attributed as follows. In Sanya Bay and in Nha Trang Bay, these changes are caused by irrational exploitation of coral reefs and water eutrophication. In coastal waters of Yonaguni Island, these changes are caused by the periodic prolonged influence of high temperature, leading to the damage and mortality of hermatypic corals, followed by the appearance of free substrates (dead and damaged coral colonies) and their occupation by sessile organisms.
Algal sampling conducted in the 1950s in the Tomioka Peninsula (early collection, EC) has documented 202 species [3]. In fact, according to Guiry & Guiry [11] and Segawa and Yoshida [3], they have found 194 species, including 57% red algae, 28% brown algae, and 15% green algae, and the ratios of taxonomic algal groups in the flora are R:P = 2.0 and C:P = 0.5. These taxonomic indices of the benthic flora in the Tomioka Peninsula coast recorded at the end of the 1950s are significantly different from our current collections in the same places during 2012–2017 (late collection, LC) and correspond to the values for these indices for the marine flora of cold-temperate region of the Northern Hemisphere (Figure 7) [19,20,24].
In 2012–2017, we found only 130 same species of algae for EC and LC, where red algae accounted for 59%, brown algae accounted for 28% and green algae accounted for 13%. For example, the taxonomic composition was close (but not identical) to that of Segawa and Yoshida’s collection [3]. The indices R/P and C/P in this group of species were closer to the values for these indices for the marine flora of warm-temperate region, but quite dissimilar to those for cold-temperate region (Figure 7).
At the same time, we did not find 62 species documented by Segawa and Yoshida. In this group, the ratios of species numbers of R/P and C/P were close to the marine flora of a cold-temperate region. In the group of “new records to Amakusa”, taxonomic composition (Rh 66%, Ph 10%, Ch 24%) and indices of ratios of individual algal groups (Figure 7) corresponded to benthic flora of tropical biogeographic regions.
Therefore, the comparison of our data (samplings of 2012–2017) with data of Segawa and Yoshida (samplings in the 1950s) [3] indicated significant changes in the benthic flora during the past 50 years as follows. The species diversity was increased by two times at the expense of red and green algae. The biogeographic status of the benthic flora was changed from the flora of a cold-temperate region to the flora of a warm-temperate region, which could be attributed to the introduction of red algae from tropical region of South-East Asia and widespread green opportunistic algae and also the disappearance of some cold-water brown algae, such as Ecklonia radicosa (Kjellman) Okamura, Desmarestia dudresnayi subsp. tabacoides (Okamura) A.F. Peters, E.C. Yang, F.C. Küpper & Prud’Homme van Reine [=Desmarestia tabacoides Okamura], Carpomitra costata (Stackhouse) Batters [=Carpomitra cabrerae (Clemente) Kützing], and Sporochnus radiciformis (R. Brown ex Turner) C. Agardh) [=Sporochnus scoparius Harvey].
Some decadal changes in the benthic marine flora in the southwest part of Kyushu Island have been documented earlier. For example, in Tosa Bay, the temperate Sargassum species declined and has been replaced by tropical species of Sargassum [33]. Moreover, in the southern part of temperate Japan, the resources of cold-water brown and red algae from genera, such as Sargassum, Eisenia, Ecklonia, Saccharina (Ph), and Gelidium (Rh), have decreased during the past 20 years, while CCA and frondose macroalgae occupy rocks or boulders [34,35]. Along the coast of the Kochi Prefecture in the Pacific region, temperate species, such as Sargassum micracanthum and S. okamurae that dominated during the 1970s, have decreased, and the subtropical species S. duplicatum has become a major component of the flora since the 1990s [36]. The appearance of subtropical Sargassum has also been reported in the area of Nagasaki, in the western Kyushu region [37]. In the Miyazaki Prefecture, the Sargassum flora consisting mainly of temperate species in the 1970s has contained many subtropical species since 2000 [37]. In above-mentioned studies, authors have mainly explained these decadal changes by two determining factors: severe exploitation of brown algae and warming of surrounding waters during some decades. Authors have confirmed these conclusions by data on the dynamics of exploitation (algal harvesting from field and their cultivation) and monitoring of water temperature. Our collections and those made by Segawa and Yoshida were sampled at sites which were not subjected to irrational exploitation, but affected by global warming [34].
The majority of authors of earlier studies have postulated that the most probable effect of direct influence of ocean warming on the coastal marine flora is a poleward shift in the distributional boundaries of species with an associated replacement of cold-water species by warm-water species and also the greatly decreased abundance of many temperature-sensitive algae in these regions [38,39,40,41,42]. Probably, such an observed shift in the benthic flora of the Tomioka Peninsula Amakusa-Shimoshima Island was also the response of benthic algal settlements and coastal ecosystems as a whole to ocean warming.
Supplementary Materials
The following data are available online at https://0-www-mdpi-com.brum.beds.ac.uk/1424-2818/11/9/158/s1, Figure S1: Algal communities in different tidal zones of the Tomioka Peninsula (Amakusa-Shimoshima Island), Table S1: Sampling location, sampling points, number and time of samplings and number of samples and Species. Table S2: List of the recent marine flora of Tomioka Peninsula (Amakusa-Shimoshima Island) found in 2012–2017.
Author Contributions
E.A.T. and X.L. conceived and designed the experiments; E.A.T., T.V.T., and M.T. performed the experiments, E.A.T., and X.L. performed the data analysis, and drafted the manuscript; X.L. supervised the project. All the authors reviewed and approved the final manuscript.
Funding
This work was financially supported by the National Natural Science Foundation of China (41476134), the Open Project of State Key Laboratory of Marine Resource Utilization in South China Sea (DX2017003), the Foundation of Hainan University (KYQD(ZR)1805) and the grant ‘China–Russia special funds 2018’ from the Chinese Academy of Sciences.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Climate-Japan. Available online: https://www.climatestotravel.com/climate/japan (accessed on 29 August 2019).
- Japan Sea temperatures. Available online: https://www.seatemperature.org/asia/japan/ (accessed on 29 August 2019).
- Segawa, S.; Yoshida, T. Decapoda crustacean. In Fauna and Flora of the Sea around the Amakusa Marine Biological Laboratory, Part II; Tomioka, R., Ed.; Kumamoto Prefecture: Amakusa, Japan, 1961; p. 24. [Google Scholar]
- Nandakumar, K.; Tanaka, M.; Kikuchi, T. Interspecific competition among fouling organisms in Tomioka Bay, Japan. Mar. Ecol. 1993, 94, 43–50. [Google Scholar] [CrossRef]
- Tioho, H.; Tokeshi, M.; Nojima, S. Experimental analysis of recruitment in a scleractinian coral at high latitude. Mar. Ecol. 2001, 213, 79–86. [Google Scholar] [CrossRef]
- Nishihara, G.N.; Noro, T.; Terada, R. Effect of temperature and light on the photosynthetic performance of two edible seaweeds: Meristotheca coacta Okamura and Meristotheca papulosa J. Agardh (Solieriaceae, Rhodophyta). Aquac. Sci. 2012, 60, 377–388. [Google Scholar]
- Terada, R.; Tanaka, T.; Uchimura, M. Morphology and distribution of Caulerpa lentillifera J. Agardh (Chrolophyceae) in Japanese waters, including the first record from Southern Kyushu and Northern Ryukyu Islands. J. Jpn. Bot. 2012, 60, 377–388. [Google Scholar]
- Nakano, Y. Global environmental change and coral bleaching. Coral Reefs Jpn. 2004, 42–48. [Google Scholar]
- Perestenko, L.P. Red Algae of the Far-Eastern Seas of Russia; Olga Publishing House: St Petersburg, Russia, 1994; p. 330. [Google Scholar]
- Titlyanova, T.V.; Titlyanov, E.A.; Kalita, T.L. Marine algal flora of Hainan Island: A comprehensive synthesis. Coast. Ecosyst. 2014, 1, 28–53. [Google Scholar]
- AlgaeBase. World-Wide Electronic Publication. Available online: http://www.algaebase.org (accessed on 29 August 2019).
- Saunders, G.W.; Hommersand, M.H. Assessing red algal supraordinal diversity and taxonomy in the context of contemporary systematic data. Am. J. Bot. 2004, 91, 1494–1507. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, R.T. Checklist and Bibliography of the Marine Benthic Algae from the Mariana Islands (Guam and CNMI); University of Guam: Guam, USA, 2003; Volume 107, pp. 1–37. [Google Scholar]
- Titlyanov, E.A.; Titlyanova, T.V.; Belous, O.S. Checklist of the marine flora of Nha Trang Bay (Vietnam, South China Sea) and decadal changes in the species diversity composition between 1953 and 2010. Bot. Mar. 2015, 58, 367–377. [Google Scholar] [CrossRef]
- Vinogradova, K.L. Green algae. In Algae of the Far-Eastern Seas of USSR; Perestenko, L.P., Ed.; Nauka: St. Petersburg, Russia, 1975; p. 147. [Google Scholar]
- Titlyanova, T.V.; Titlyanov, E.A.; Xia, B.; Bartsch, I. New records of benthic marine green algae (Chlorophyta) for the island of Hainan (China). Nova Hedwigia 2012, 94, 441–470. [Google Scholar] [CrossRef]
- Littler, D.M.; Littler, M.M. Caribbean Reef Plants; OffShore Graphics: Washington, DC, USA, 2000; p. 542. [Google Scholar]
- Titlyanova, T.V.; Titlyanov, E.A.; Li, X.B. The significance of new records of benthic red algae (Rhodophyta) for Hainan Island (and China) between 1990 and 2016. Diversity 2017, 9, 24. [Google Scholar] [CrossRef]
- Briggs, J.C. Marine Zoogeography; McGraw-Hill: New York, NY, USA, 1974; p. 475. [Google Scholar]
- Santelices, B.; Bolton, J.J.; Meneses, I. Chapter Six Marine Algal Communities. In Marine Macroecology; Whitman, J.D., Roy, K., Eds.; Chicago University Press: Chicago, IL, USA, 2009; pp. 153–192. [Google Scholar]
- Lüning, K. Seaweeds. Their Environment, Biogeography and Ecophysiology; John Wiley & Sons: Hoboken, NJ, USA, 1990; p. 527. [Google Scholar]
- Pham, H.H. Marine Algae of South Vietnam; Trung Tam Hoc Lieu Saigon: Saigon, Vietnam, 1969; p. 558. [Google Scholar]
- Lewis, J.E.; Norris, J.N. A history and annotated account of the benthic marine algae of Taiwan. Smithson. Contrib. Mar. Sci. 1987, 29, 1–38. [Google Scholar] [CrossRef]
- Perestenko, L.P. Algae of the Peter Great Bay (the Sea of Japan); Nauka: St. Petersburg, Russia, 1980; p. 232. [Google Scholar]
- Nguyen, T.V.; Le, N.H.; Lin, S.M.; Steen, F.; de Clerck, O. Checklist of the marine macroalgae of Vietnam. Bot. Mar. 2013, 56, 207–227. [Google Scholar] [CrossRef]
- Belous, O.S.; Titlyanova, T.V.; Titlyanov, E.A. Marine Plants of the Trinity Bay and Adjacent Water Areas; Dalnauka: Vladivostok, Russia, 2013; p. 264. [Google Scholar]
- Titlyanov, E.A.; Titlyanova, T.V.; Li, X.B.; Kalita, T.L.; Huang, H. Recent (2008–2012) seaweed flora of Hainan Island, South China Sea. Mar. Biol. Res. 2015, 11, 540–550. [Google Scholar] [CrossRef]
- Hayashida, F. Synecological studies of a brown alga, Ecklonia cava Kjellmann, forming aquatic forest. I. Benthic marine algal vegetation at survey area. J. Mar. Sci. Tech-Japan. 1983, 16, 207–212. [Google Scholar]
- Hayashida, F. Synecological studies of a brown alga, Ecklonia cava Kjellmann, forming aquatic forest. II. On growth of Ecklonia cava. J. Mar. Sci. Tech-Jpn. 1984, 18, 275–280. [Google Scholar]
- Maegawa, M.; Yokohama, Y.; Aruga, Y. Critical light conditions for young Ecklonia cava and Eisenia bicyclis with reference to photosynthesis. Hydrobiologia 1987, 447–455. [Google Scholar] [CrossRef]
- Titlyanov, E.A.; Titlyanova, T.V.; Belous, O.S.; Kalita, T.L. Inventory change (1990s–2010s) in the marine flora of Sanya Bay (Hainan Island, China). J. Mar. Biol. Assoc. UK 2015, 95, 461–470. [Google Scholar] [CrossRef]
- Titlyanov, E.A.; Titlyanova, T.V.; Kalita, T.L.; Tokeshi, M. Decadal changes in the algal assemblages of tropical-subtropical Yonaguni Island in the western Pacific. Coast. Ecosyst. 2016, 3, 16–37. [Google Scholar]
- Haraguchi, H.; Sekida, S. Recent changes in the distribution of Sargassum species in Kochi, Japan. Kuroshio Sci. 2008, 2, 41–46. [Google Scholar]
- Nagai, S.; Yoshida, G.; Tarutani, K. Change in species composition and distribution of algae in the coastal waters of western Japan. In Global Warming Impacts: Case Studies on the Economy, Human Health, and on Urban and Natural Environments; Casalegno, S., Ed.; BoD–Books on Demand: Norderstedt, Germany, 2011; pp. 2–31. [Google Scholar]
- Fujita, D. Current status and problems of isoyake in Japan. Bull Fish Res. Agen 2010, 32, 33–42. [Google Scholar]
- Hiraoka, M.; Ura, Y.; Haraguchi, H. Relationship between seaweed beds and seawater temperature in the Tosa Bay. Aquabiology 2005, 27, 485–493. [Google Scholar]
- Aratake, H.; Shimizu, H.; Watanabe, K.; Yoshida, G. Long-term change in Sargassum-bed distribution along the coast of Kushima-city, southern part of Miyazaki Prefecture, Japan. Bull. Miyazaki Prefectural Fish. Exp. Stn. 2007, 11, 1–13. [Google Scholar]
- Lima, F.P.; Ribeiro, P.A.; Queiroz, N.; Hawkins, S.J.; Santos, A.M. Do distributional shifts of northern and southern species of algae match the warming pattern? Glob. Chang. Biol. 2007, 13, 2592–2604. [Google Scholar] [CrossRef]
- Hawkins, S.J.; Moore, P.J.; Burrows, M.T.; Poloczanska, E.; Mieszkowska, N.; Herbert, R.J.H.; Jenkins, S.R.; Thompson, R.C.; Genner, M.J.; Southward, A.J. Complex interactions in a rapidly changing world: Responses of rocky shore communities to recent climate change. Clim. Res. 2008, 37, 123–133. [Google Scholar] [CrossRef]
- Schiel, D.; Steinbeck, J.R.; Foster, M.S. Ten years of induced ocean warming causes comprehensive. Ecology 2004, 85, 1833–1839. [Google Scholar] [CrossRef]
- Araujo, R.M.; Assis, J.; Aquillar, R.; Airoldi, L.; Barbara, I.; Bartsch, I.; Bekkby, T.; Christie, H.; Davoult, D.; Derrien-Courtel, S.; et al. Status, trends and drivers of kelp forests in Europe: An expert assessment. Biodivers. Conserv. 2016, 25, 1319–1348. [Google Scholar] [CrossRef]
- Harley, C.D.G.; Anderson, K.M.; Demes, K.W.; Jorve, J.P.; Kordas, R.L.; Coyle, T.A. Effects of climate change on global seaweed communities. J. Phycol. 2012, 48, 1064–1078. [Google Scholar] [CrossRef]
Figure 1.
Algal sampling sites in the Tomioka Peninsula, Amakusa-Shimoshima Island, in 2012–2017. (A) Sampling location and sampling points (red circles); (B) Akaiwa, sampling point 5; (C) Magarizaki, sampling point 7; (D) Shikizaki Bay, sampling point 3; (E) Shiraiwazaki Bay, sampling point 2; (F) Tomioka Harbor, (a) sampling point 9, (b) sampling point 11; (G) Tsujishima Islet, sampling point 14.
Figure 1.
Algal sampling sites in the Tomioka Peninsula, Amakusa-Shimoshima Island, in 2012–2017. (A) Sampling location and sampling points (red circles); (B) Akaiwa, sampling point 5; (C) Magarizaki, sampling point 7; (D) Shikizaki Bay, sampling point 3; (E) Shiraiwazaki Bay, sampling point 2; (F) Tomioka Harbor, (a) sampling point 9, (b) sampling point 11; (G) Tsujishima Islet, sampling point 14.

Figure 2.
Species diversity and composition of seaweeds′ assemblages along coast of the Tomioka Peninsula in 2012–2017. (A) Absolute and relative species numbers of different taxonomic groups Rhodophyta, Phaeophyceae, Chlorophyta (Rh, Ph, Ch). (B) Species numbers in dominant families.
Figure 2.
Species diversity and composition of seaweeds′ assemblages along coast of the Tomioka Peninsula in 2012–2017. (A) Absolute and relative species numbers of different taxonomic groups Rhodophyta, Phaeophyceae, Chlorophyta (Rh, Ph, Ch). (B) Species numbers in dominant families.

Figure 3.
Relative species number from the Phyla Rh, Ch, and Class Ph in different groups of epilithic and epiphytic benthic algae of the collection of 2012–2017: HS, algae growing on hard natural substrates; OHS, algae growing only on hard natural substrates; Ep, algae growing epiphytically; OEp, algae growing only epiphytically. Numbers above the columns are total number of species in individual groups. (A) Collection of benthic macroalgae sampled in 2012–2017. (B) New findings in the collection of 2012–2017.
Figure 3.
Relative species number from the Phyla Rh, Ch, and Class Ph in different groups of epilithic and epiphytic benthic algae of the collection of 2012–2017: HS, algae growing on hard natural substrates; OHS, algae growing only on hard natural substrates; Ep, algae growing epiphytically; OEp, algae growing only epiphytically. Numbers above the columns are total number of species in individual groups. (A) Collection of benthic macroalgae sampled in 2012–2017. (B) New findings in the collection of 2012–2017.

Figure 4.
Absolute and relative number of algal species in the collection 2012–2017 in groups differing in geographic (latitudinal) distribution. (A) Species number in different algal groups of the collection 2012–2017. (B) Relative number of species from the Phyla Rh, Ch, and the Class Ph in different geographic groups of algae of the collection 2012–2017. (C) Algal species number in geographic groups of new findings, new records (NR) in the collection 2012–2017. (D) Relative algal species number in the Phyla Rh, Ch, and the Class Ph in different geographic algal groups of new findings in 2012–2017. TS, algae inhabiting tropical and subtropical seas (OTS); M, algae inhabiting the tropics and/or subtropics to temperate zones; Ar/An, cosmopolitan algae growing from the tropics to the Arctic and/or Antarctic; IPA, algae inhabiting the Atlantic, Indian and Pacific Oceans; IP, algae inhabiting the Indo-Pacific.
Figure 4.
Absolute and relative number of algal species in the collection 2012–2017 in groups differing in geographic (latitudinal) distribution. (A) Species number in different algal groups of the collection 2012–2017. (B) Relative number of species from the Phyla Rh, Ch, and the Class Ph in different geographic groups of algae of the collection 2012–2017. (C) Algal species number in geographic groups of new findings, new records (NR) in the collection 2012–2017. (D) Relative algal species number in the Phyla Rh, Ch, and the Class Ph in different geographic algal groups of new findings in 2012–2017. TS, algae inhabiting tropical and subtropical seas (OTS); M, algae inhabiting the tropics and/or subtropics to temperate zones; Ar/An, cosmopolitan algae growing from the tropics to the Arctic and/or Antarctic; IPA, algae inhabiting the Atlantic, Indian and Pacific Oceans; IP, algae inhabiting the Indo-Pacific.

Figure 5.
Species diversity and taxonomic composition of new findings of seaweeds along coast of the Tomioka Peninsula in 2012–2017. (A) Absolute and relative species numbers of different taxonomic groups (Rh, Ph, Ch). (B) Species numbers in dominant families.
Figure 5.
Species diversity and taxonomic composition of new findings of seaweeds along coast of the Tomioka Peninsula in 2012–2017. (A) Absolute and relative species numbers of different taxonomic groups (Rh, Ph, Ch). (B) Species numbers in dominant families.

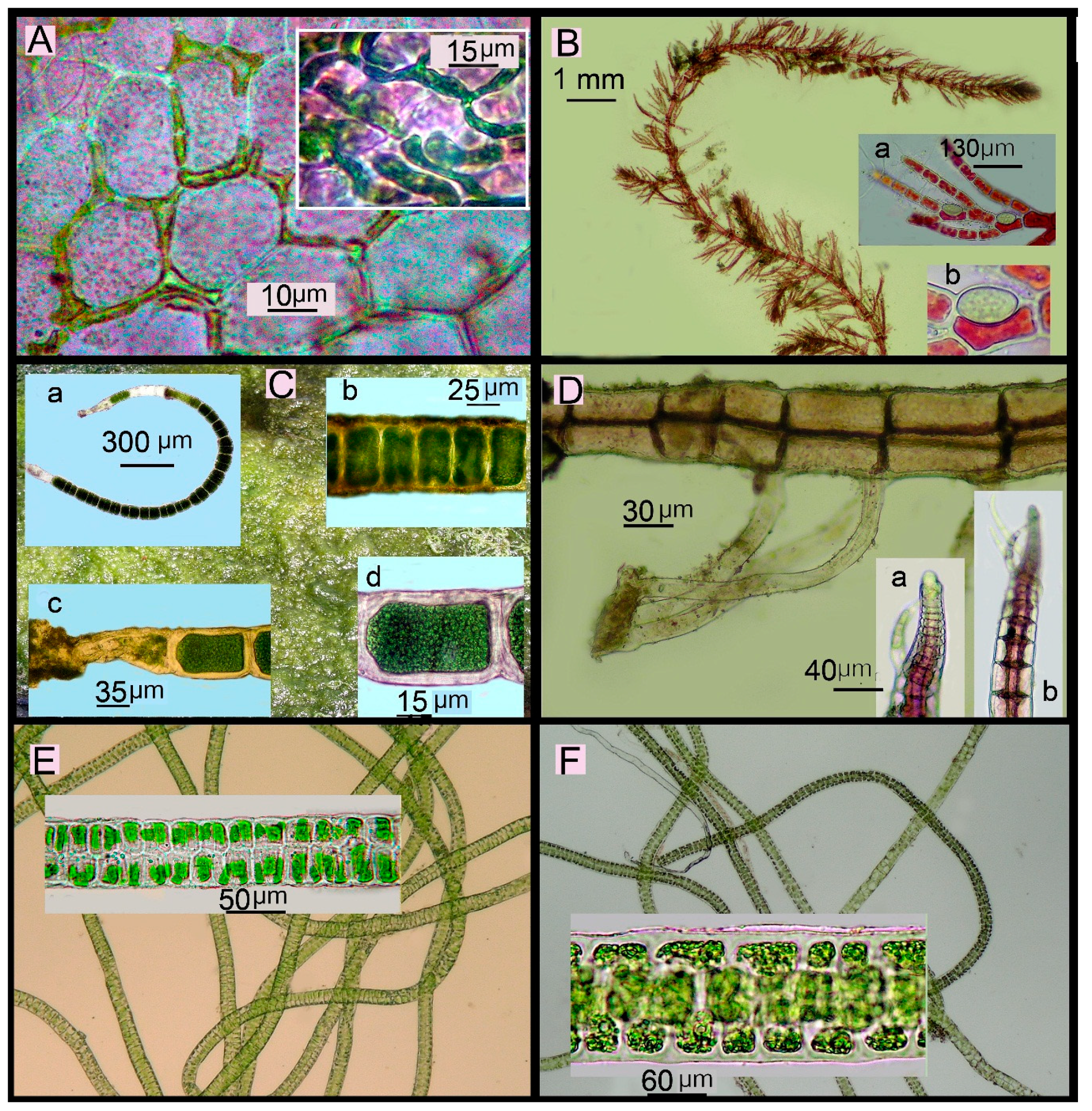
Figure 6.
New records of benthic macroalgae for Japan found in Amakusa-Shimoshima Island. (A) Acrochaete geniculata (N.L. Gardner) O’Kelly, surface view showing cells arranged between outer cells of Hypnea sp. and Ceramium borneense (Inset). (B) Antithamnionella longicellulata Perestenko, habit. Insets: a, b, fragments showing ovoid gland cells. (C) Chaetomorpha javanica Kützing. Insets: a, habit; b, fragment showing dividing cells; d, fragment showing thick striated cell walls; d, basal part. (D) Melanothamnus pseudovillum (Hollenberg) Díaz-Tapia & Maggs, prostrate axis showing unicellular rhizoids proximal on pericentral cells. Insets: a, b, apex of erect axis. (E) Ulva chaetomorphoides (Börgesen) Hayden, Blomster, Maggs, P.C. Silva, M.J. Stanhope & J.R. Waaland, habit. Inset: fragment of filament in surface view. (F) Ulva ralfsii (Harvey) Le Jolis, habit. Inset: fragment of filament in surface view.
Figure 6.
New records of benthic macroalgae for Japan found in Amakusa-Shimoshima Island. (A) Acrochaete geniculata (N.L. Gardner) O’Kelly, surface view showing cells arranged between outer cells of Hypnea sp. and Ceramium borneense (Inset). (B) Antithamnionella longicellulata Perestenko, habit. Insets: a, b, fragments showing ovoid gland cells. (C) Chaetomorpha javanica Kützing. Insets: a, habit; b, fragment showing dividing cells; d, fragment showing thick striated cell walls; d, basal part. (D) Melanothamnus pseudovillum (Hollenberg) Díaz-Tapia & Maggs, prostrate axis showing unicellular rhizoids proximal on pericentral cells. Insets: a, b, apex of erect axis. (E) Ulva chaetomorphoides (Börgesen) Hayden, Blomster, Maggs, P.C. Silva, M.J. Stanhope & J.R. Waaland, habit. Inset: fragment of filament in surface view. (F) Ulva ralfsii (Harvey) Le Jolis, habit. Inset: fragment of filament in surface view.

Figure 7.
The average levels of the R/P indices (number of Rhodophyta species divided by the number of Phaeophycean species) (A) and C/P (number of Chlorophyta species divided by the number of Phaeophycean species) (B) in the marine floras of cold-temperate region (CT), warm-temperate region (WT), tropical region (TR) [20], in the early collection (EC) and late collection (LC), and also in the following groups of new findings in LC (NLC), common species in EC and LC (ELC) and species recorded only in EC (OEC).
Figure 7.
The average levels of the R/P indices (number of Rhodophyta species divided by the number of Phaeophycean species) (A) and C/P (number of Chlorophyta species divided by the number of Phaeophycean species) (B) in the marine floras of cold-temperate region (CT), warm-temperate region (WT), tropical region (TR) [20], in the early collection (EC) and late collection (LC), and also in the following groups of new findings in LC (NLC), common species in EC and LC (ELC) and species recorded only in EC (OEC).

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Titlyanov, E.A.; Titlyanova, T.V.; Tokeshi, M.; Li, X. Inventory and Historical Changes in the Marine Flora of Tomioka Peninsula (Amakusa Island), Japan. Diversity 2019, 11, 158. https://0-doi-org.brum.beds.ac.uk/10.3390/d11090158
AMA Style
Titlyanov EA, Titlyanova TV, Tokeshi M, Li X. Inventory and Historical Changes in the Marine Flora of Tomioka Peninsula (Amakusa Island), Japan. Diversity. 2019; 11(9):158. https://0-doi-org.brum.beds.ac.uk/10.3390/d11090158
Chicago/Turabian StyleTitlyanov, Eduard A., Tamara V. Titlyanova, Mutsunori Tokeshi, and Xiubao Li. 2019. "Inventory and Historical Changes in the Marine Flora of Tomioka Peninsula (Amakusa Island), Japan" Diversity 11, no. 9: 158. https://0-doi-org.brum.beds.ac.uk/10.3390/d11090158
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.