Endoparasitic Mites (Rhinonyssidae) on Urban Pigeons and Doves: Updating Morphological and Epidemiological Information


1
Department of Functional and Evolutionary Ecology, Experimental Station of Arid Zones (EEZA-CSIC), 04120 Almería, Spain
2
Departament of Human Anatomy, State Pediatric Medical University, Litovskaya st. 2, 194100 St. Petersburg, Russia
3
Department of Microbiology and Parasitology, Faculty of Pharmacy, University of Sevilla, Profesor García González 2, 41012 Sevilla, Spain
*
Author to whom correspondence should be addressed.
Diversity 2021, 13(1), 11; https://0-doi-org.brum.beds.ac.uk/10.3390/d13010011
Submission received: 27 November 2020
/
Revised: 21 December 2020
/
Accepted: 28 December 2020
/
Published: 31 December 2020
(This article belongs to the Special Issue Bird Parasites)
Abstract
:Rhynonyssidae is a family of endoparasitic hematophagous mites, which are still largely unknown even though they could act as vector or reservoir of different pathogens like dermanyssids. Sampling requirements have prevented deeper analysis. Rhinonyssids have been explored in a few host specimens per species, leading to undetailed morphological descriptions and inaccurate epidemiology. We explore the relationships established between these parasites in two Columbiformes urban birds (domestic pigeon (Columba livia domestica) and Eurasian collared dove (Streptopelia decaocto)), assesing 250 individuals of each type in Seville (Spain). As expected, Mesonyssus melloi (Castro, 1948) and Mesonyssus columbae (Crossley, 1950) were found in domestic pigeons, and Mesonyssus streptopeliae (Fain, 1962) in Eurasian collared doves. However, M. columbae was found for the first time in Eurasian collared doves. This relationship could be common in nature, but sampling methodology or host switching could also account for this result. An additional unknown specimen was found in a Eurasian collared dove, which could be a new species or an aberrant individual. We also provide an epidemiological survey of the three mite species, with M. melloi being the most common one followed by M. streptopeliae and M. columbae. High variation between previous epidemiological measurements and ours highlights the importance of developing deeper studies to uncover the factors regulating mite prevalence and intensities of infection.
1. Introduction
Rhinonyssidae Trouessart, 1895 (Acari: Parasitiformes: Mesostigmata) is a family of hematophagous mites that inhabits the nasal cavities of various orders of birds [1]. Currently, the family Rhinonyssidae includes more than 600 species arranged in 11 genera: Larinyssus Strandtmann, 1948; Locustellonyssus Bregetova, 1965; Mesonyssus Fain, 1960; Ptilonyssoides Vitzthum, 1935; Ptilonyssus Berlese et Trouessart, 1889; Rallinyssus Strandtmann, 1948; Rhinoecius Cooreman, 1946; Rhinonyssus Trouessart, 1894; Sternostoma Berlese et Trouessart, 1889; Tinaminyssus Strandtmann et Wharton, 1958; Vitznyssus Castro, 1948. It is currently considered the most diverse family of nasal mites [2].
During their feeding activity, rhinonyssids can damage the nasal cavities of birds, leading to the development of Rhinonyssidosis avium disease [3]. Some species, like Sternostoma tracheacolum Lawrence, 1948, can reach not only the nasal passages of birds but also their tracheae, lungs and air sacs [4]. As a result, they are highly pathogenic in some host species like canaries and Gouldian finches [4,5]. Furthermore, even when not experimentally demonstrated, they could act as a potential reservoir or vector of some pathogens, as has been shown for the red poultry mite Dermanyssus gallinae, De Geer, 1778, transmitting Salmonella enterica [6] and avian influenza virus [7]. In fact, a recent metagenomics study has shown the presence of pathogenic bacteria like Clostridioides difficile in Tinaminyssus (=Mesonyssus) melloi [8], suggesting that deeper studies are necessary to explore the real potential of these mites as vectors or reservoirs.
Unfortunately, some aspects of the family Rhinonyssidae have been neglected in research, and our knowledge about them is scarce. Most studies are focused on identifying the participants in the host-parasite relationships through the study of just a few of specimens of each host species, which leads to inaccurate epidemiology [9,10,11,12,13]. This is partially due to the tiny size of the individuals, the expertise needed for their morphological identification and the requirements for sampling. Correct sampling of nasal mites requires dead hosts and expertise to open bird nasal cavities [14]. Nonetheless, some generalities about this group are known. They are endoparasites that frequently inhabit avian hosts and lack an off-host stage. They are mainly transmitted directly through oral cavities when hosts exchange food during nestling feeding or courtship behaviour, but indirect transmission can also take place via water, perches and other contaminated surfaces [15]. Furthermore, their degree of host specificity is variable, ranging from specialists that parasitize species to others that can infect avian hosts across different families [16].
The development of deeper studies on rhinonyssids is hampered by the lack of a reliable identification of closely related species of the family Rhinonyssidae. This is a consequence of the absence of species-level keys and the scattered literature on morphometrical information [17]. The genus Mesonyssus Fain, 1960, that infests columbiform birds is directly affected by this fact. Since its species show a very similar morphology, they are usually arranged into three species groups where specific identification is difficult: melloi, columbae and zenaidurae [18]. Currently, the genus Mesonyssus includes 24 species that parasitize Columbiformes and six species that are associated with Pelecaniformes [2]. Columbiformes that coexist close to human populations (e.g., Columba livia domestica and Streptopelia decaocto) are considered of special interest for public health [19], so their symbiont relationships deserve to be explored.
In this study we take advantage of a population control program developed in the city of Seville, to examine the rhinonyssids present in two sympatric species of Columbiformes that were captured and culled in an urban area: Columba livia domestica (domestic pigeon) and Streptopelia decaocto (Eurasian collared dove). Our aim is to offer new insight into the relationships established between these organisms in order to expand on and improve the scarce knowledge available of this host-parasite system. We report an updated morphological description, illustrations, and an epidemiological assessment of Mesonyssus melloi, Mesonyssus columbae and Mesonyssus streptopeliae, based on a remarkable number of hosts, to develop a comprehensive survey of the parasitic relationships in urban Columbiformes. We also explore the coexistence of mite species and possible host switching events. Such information may be useful to understand key aspects of the biology of these species and to develop future studies on them.
2. Materials and Methods
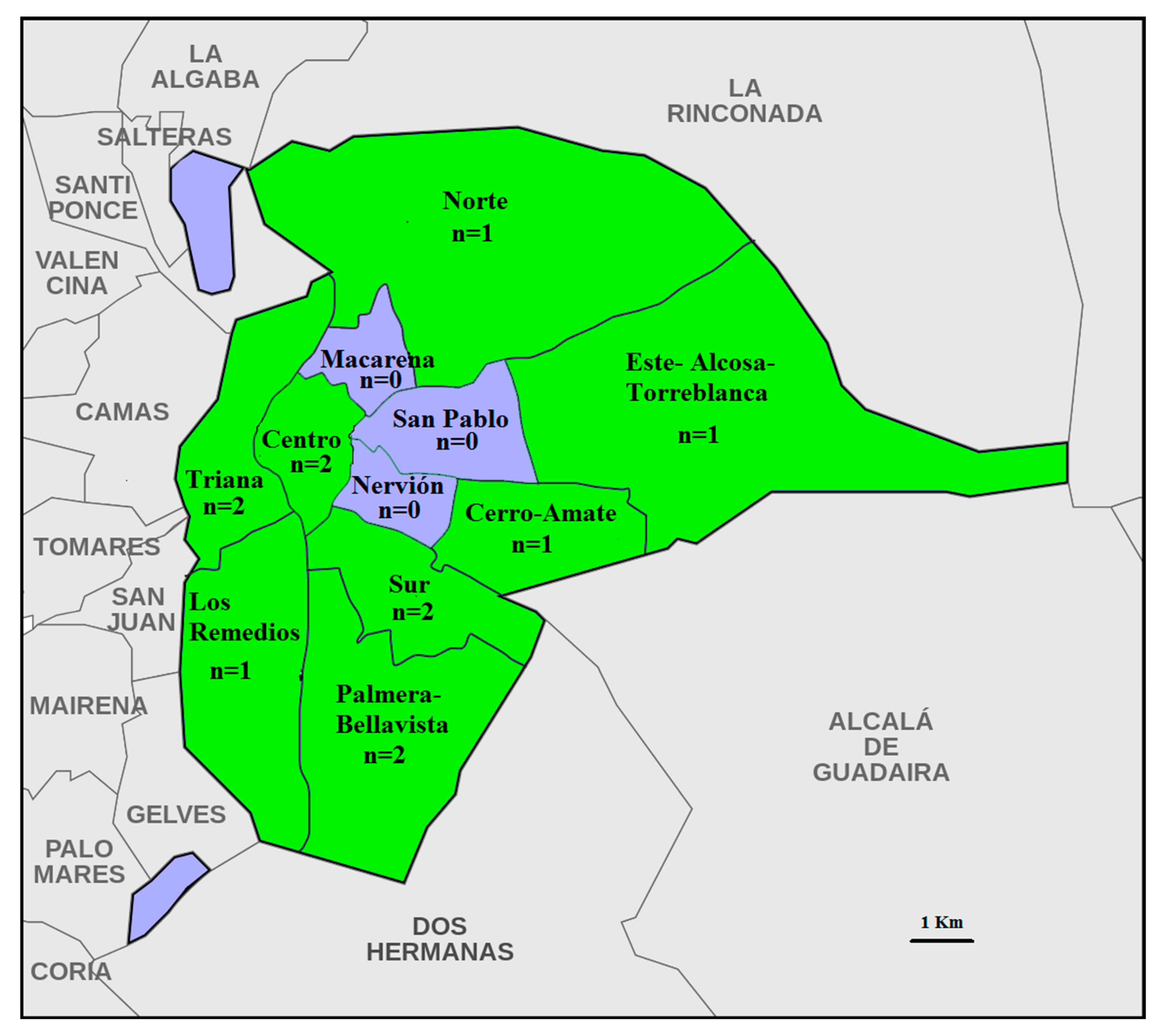
In this paper, 500 urban columbiform hosts (250 Columba livia domestica and 250 Streptopelia decaocto) were examined from the municipality of Seville, Andalusia, Spain. The individuals were attracted using water and food and captured using trap cages in different urban areas of Seville throughout 2018 and 2019. From the eleven districts of Seville, eight were sampled in one or two locations (total locations = 12, Figure 1). The “Centro Zoosanitario de Sevilla”, which is in charge of the bird population control program developed in the city of Seville, captured the individuals and provided them to us for parasite examination. Once the birds were captured and euthanized by institute staff, samples of each specimen were placed into individual plastic bags and refrigerated at 4 °C until examination. The procedure for bird trapping, handling and euthanizing used here was included in the animal control program of the Seville City Council and approved by the “Consejo Municipal de Protección y Bienestar Animal (COMBA)”.
To collect the mites, the nasal cavities were opened under an Olympus binocular stereo microscope with a scalpel and small scissors. First, the ossa mandibulae was removed together with the hyoid brush apparatus (apparatus hyobranchialis), then the scalp and feathers were removed with tweezers. Following this, a transverse section was made in the papillae pharyngeales region. A medial section was made, starting from the rima infundibuli through the choana and ending in the ruga palatina mediana. The nostrils in the area of the operculum were examined, and then the os maxillare was opened at the level of its fusion with the palate. Then, the maxillary bone was removed and the nasal cavity with its three conchae was opened: the largest of all rostral concha nasalis rostralis; the middle, concha nasalis media; and the caudal, concha, nasalis caudalis. Examination of all nasal cavities was carried out under a dissecting microscope with tweezers and a dissecting needle [14]. To avoid contamination, tweezers and needles were sterilized between host specimens by flaming over a Bunsen burner.
The mites obtained were washed in saline solution of 0.9% and fixed in 70% ethanol. They were then clarified with lactic acid and mounted on slides using Hoyer’s medium. Photographs were taken using an Olympus CX 21 microscope with an attached camera system. Illustrations were made, and measurements were taken using a contrast phase Zeiss microscope with camera lucida.
The identification of the mites was based on morphological characteristics in accordance with previous studies [14,18,20,21,22,23,24,25,26,27,28]. Mites were sexed based on the position of the genital pore, the presence of spermatodactyl in the chelicerae and a pair of long setae in the ventral zone, close to gnathosoma. For the epidemiological analysis, we assessed the prevalence and its confidence interval (Clopper–Pearson interval), and the mean intensities of each mite species in each host species. Fisher’s exact test was used to compare the prevalence between the coexisting mite species, M. melloi and M. columbae, and bootstrap t-test (after 9999 bootstrap replicates) was used to compare the mean intensities (see [29]). Slope calculations and the Kendall correlation test were respectively employed to check for male to female mite ratios and correlations for each parasite species.
3. Results
A total of 1491 adult nasal mites (range 0–114) belonging to three species were collected in the analysis of 500 Columbiformes. Mesonyssus melloi and M. columbae in C. livia domestica and M. columbae and M. streptopeliae in S. decaocto (Figure 2). One additional individual with no morphological coincidence with any species of rhinonyssids was found. Since the non-adult stages of these mites are hard to identify correctly, they were not explored in the current study.
3.1. Morphological Analysis
All mites isolated from urban Columbiformes were included in the genus Mesonyssus, which is specific to this order of birds and also associated with Pelecaniformes [14]. For each species identified, a complete updated morphometric study was carried out (measurements not shown). The morphological features of the species re-descriptions were as follows:
- Genus Mesonyssus Fain, 1960 [35]. Mesonyssus [14,21,23,35,36,37,38]. Tinaminyssus [24,25,39,40,41]. Type species: Neonyssus treronis [42].Diagnosis: Idiosoma oval, 450–630 μm long. Dorsal side of idiosoma with podosomal and opisthosomal shields. Pygidial shield absent. Stigmata with oblong peritrems located dorsolaterally. Mesosomal shields absent. Poststigmal sclerites present or absent. Gnathosoma located terminally. Deutosternal denticles absent. Tritosternum absent. Sternal shield absent. Genital shield present. Anal shield surrounding anus, located ventrally. Cribrum (Aspero) present. Convexities and spines on coxae present or absent.
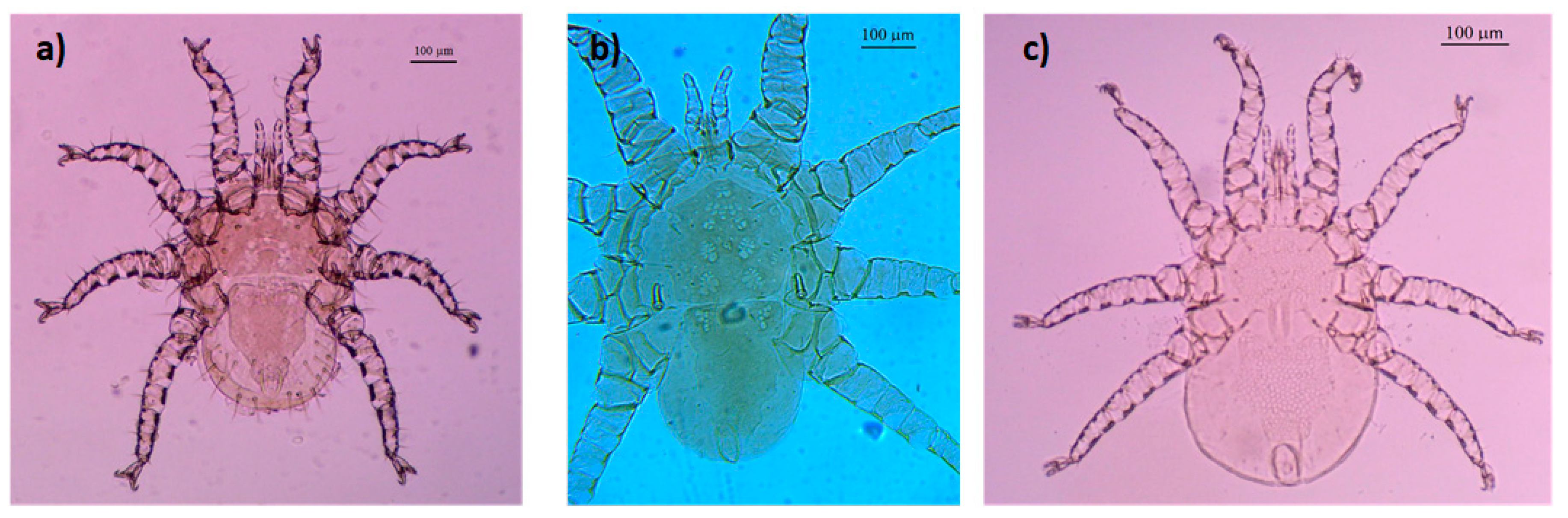
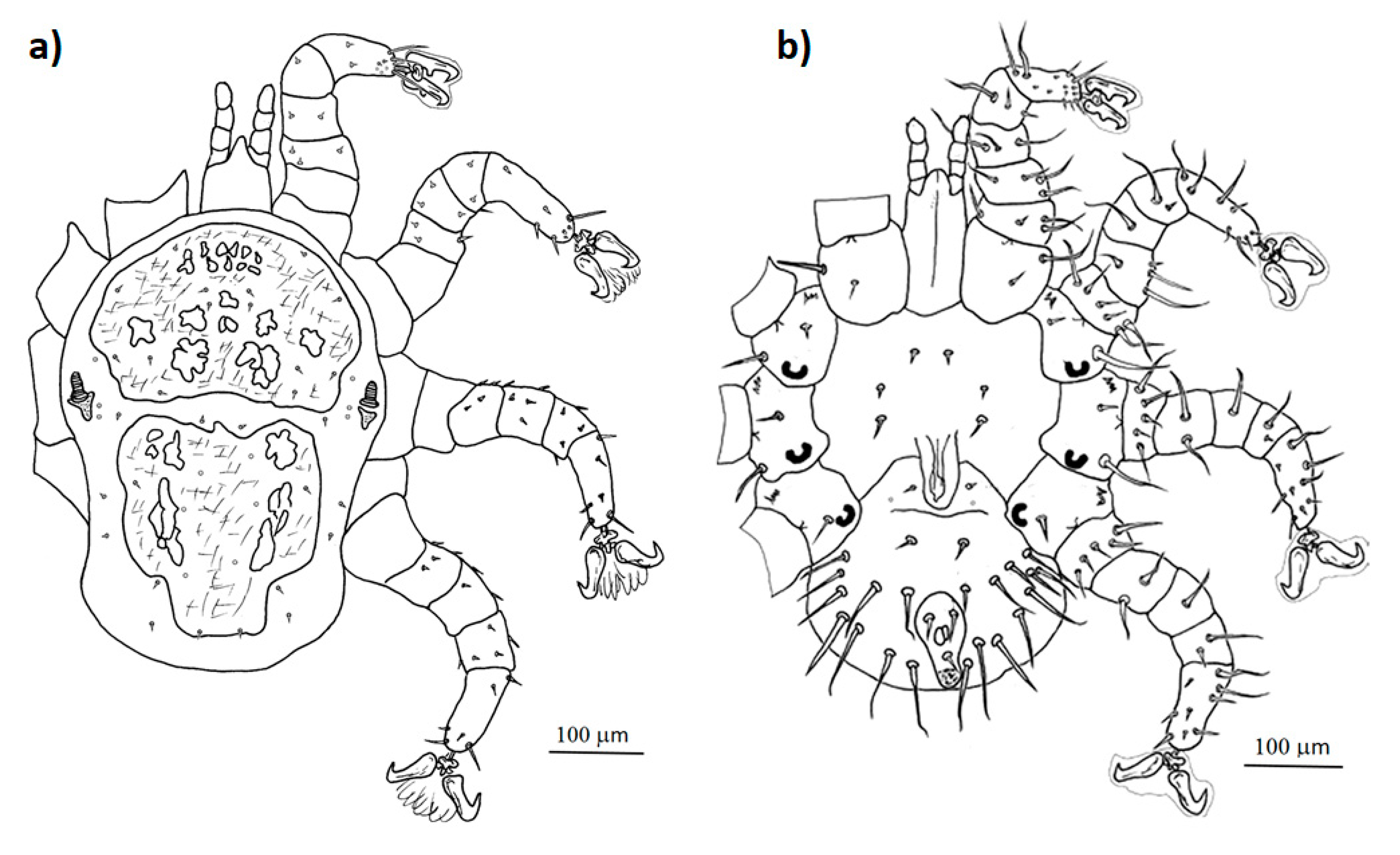
- Mesonyssus melloi (Castro, 1948) [43] (Figure 2a). Neonyssus melloi [22,43,44]. Mesonyssus melloi [14,21,27,45]. Tinaminyssus melloi [24,25,26,40]. Mesonyssus melloi melloi [23]. Neonyssus hirsutus [46].Dorsum (Figure 3a): Podosomal shield wide, strongly sclerotized, with anterior margin convex, almost semi-circular. Podosomal shield shows seven pairs of setae. Stigmata with short peritrems, located dorsolaterally, at level of coxae III. Post-stigmatic sclerites of triangular shape. On podosoma cuticle three pairs of mesolateral setae can be observed. Opisthosomal cuticle presents six pairs of fine setae. Opisthosomal shield wide, large, occupies almost entire opisthosoma. Anterior margin of this shield sinuous, and includes three pairs of pores and two pairs of setae.Ventrum: (Figure 3b) Sternal shield absent. three pairs of sternal setae present; Genital shield of medium size, thin and narrow. A pair of genital setae situated lateral to genital shield. Ventral opisthosoma with a pair of short setae Jv1 and 10 pairs of long setae. Anal shield pear-shaped, broadened anteriorly, bearing a pair of preanal setae, located anterior to anus, and one posterior postanal seta Cribrum (Aspero) present. Coxae I–III with two setae of different length. Posterior parts of coxae II–IV convexitas coxae. Anterior part of coxae II–IV with spina coxae.Gnathosoma: Oblong. Gnathosomal formula: 2-4-2. Three pairs of hypostomal setae and one pair of subcapitular setae present.
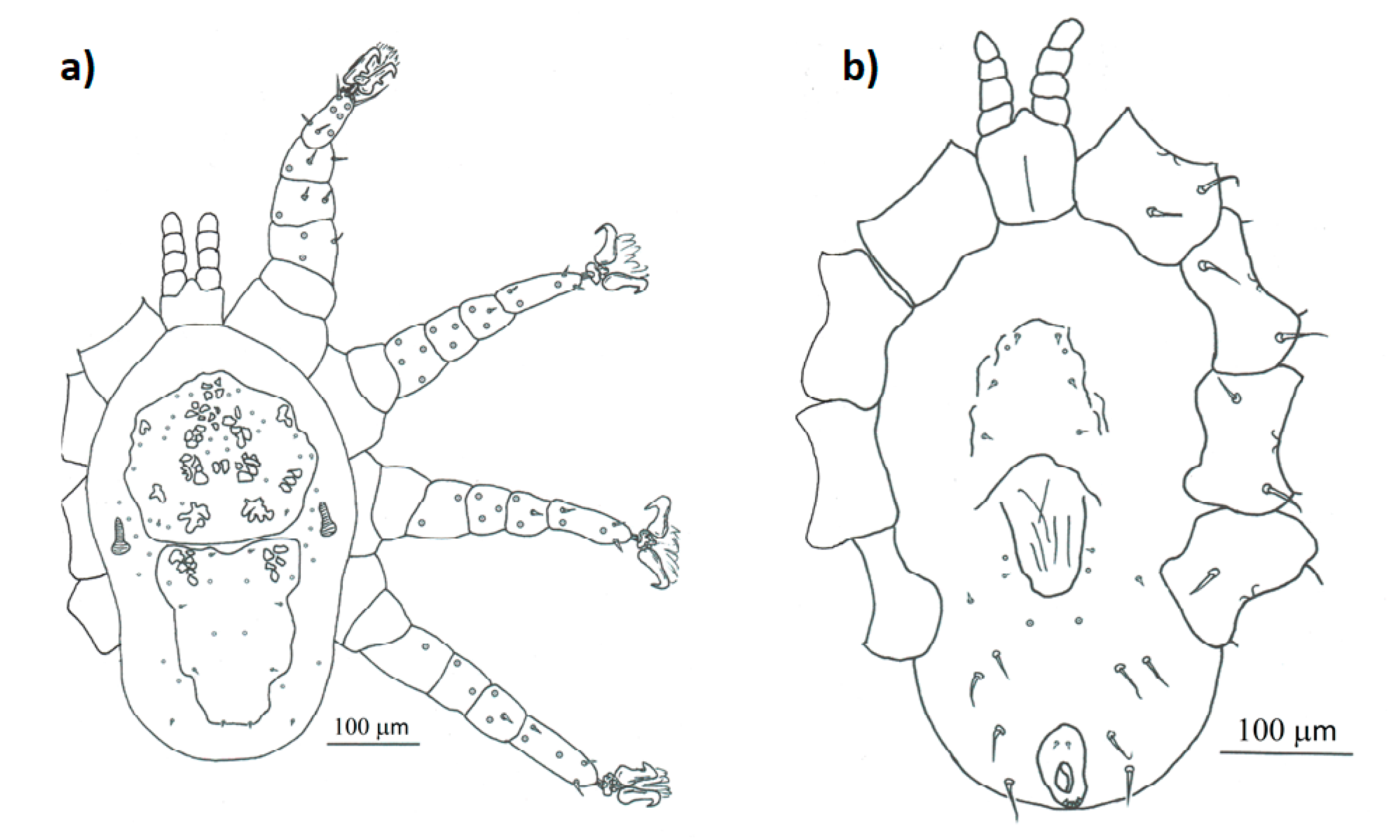
- Mesonyssus columbae (Crossley, 1950) [47] (Figure 2b). Neonyssus columbae [22,47]. Mesonyssus columbae [14,21,23,45]. Tinaminyssus columbae [24,25,40].Dorsum (Figure 4a): Podosomal and opisthosomal shields strongly sclerotized, which present on their surface 26 setae. Stigmata with peritrems, located at level of coxae III. The opisthosomal shield shows four pairs of setae and three pairs of pores. Dorsal podosomal cuticle includes three pairs of mesolateral setae. Dorsal opisthosomal cuticle presents four pairs of setae.Ventrum (Figure 4b): Sternal shield absent. Three pairs of sternal setae present. Genital shield wide, one pair of genital setae and one pair of pores situated lateral to this shield. Ventral opisthosomal idiosoma with six pairs of setae. Anal shield narrow oval and includes one pair of preanal setae. Aspero present.Gnathosoma: Wide. Gnathosomal formula (2-4-0). Subcapitular setae absent.
- Mesonyssus streptopeliae Fain, 1962 [18] (Figure 2c). Mesonyssus melloi streptopeliae [18]. Mesonyssus streptopeliae [20,21,27,28].Dorsum (Figure 5a): Podosomal and opisthosomal shields strongly sclerotized. Podosomal shield with round form with seven pairs of setae. Stigmata with peritrems, located at level of coxae III. Post-stigmatic sclerites of triangular shape. The opisthosomal shield wave-shaped with four pairs of setae. Dorsal podosomal cuticle includes three pairs of mesolateral setae. Dorsal opisthosomal cuticle presents four long pairs and three short pairs of setae.Ventrum (Figure 5b): There is a subtle rudiment of sternal shield. Three pairs of short sternal setae present. Genital shield wide. Ventral opisthosomal idiosoma with six pairs of setae and four pores. Anal shield round and includes one pair of preanal setae. Aspero absent.Gnathosoma: Thin and oblong. Gnathosomal formula (2-4-2). Palps long, with short and thin setae.
3.2. Epidemiological Analysis
C. livia domestica had a higher prevalence of Mesonyssus melloi than of M. columbae (Table 1; Fisher test, p < 0.001). Specifically, 85 out of 250 C. livia domestica harboured Mesonyssus melloi while 14 of them harboured M. columbae (Table 1). Mixed infection of the former species of mites occurs in 8.8% of the infested C. livia domestica. Regarding S. decaocto, 26 out of 250 individuals harboured M. streptopeliae (Table 1). Furthermore, one S. decaocto infested with M. streptopeliae also harboured three M. columbae mites.
The highest mean infection intensity was obtained for M. melloi, followed by M. columbae and then by M. streptopeliae (Table 1). The mean infection intensity of M. melloi on C. livia domestica was significantly higher than that of M. columbae (t = 2.06, Df = 54.96, bootstrapped p-value = 0.037).
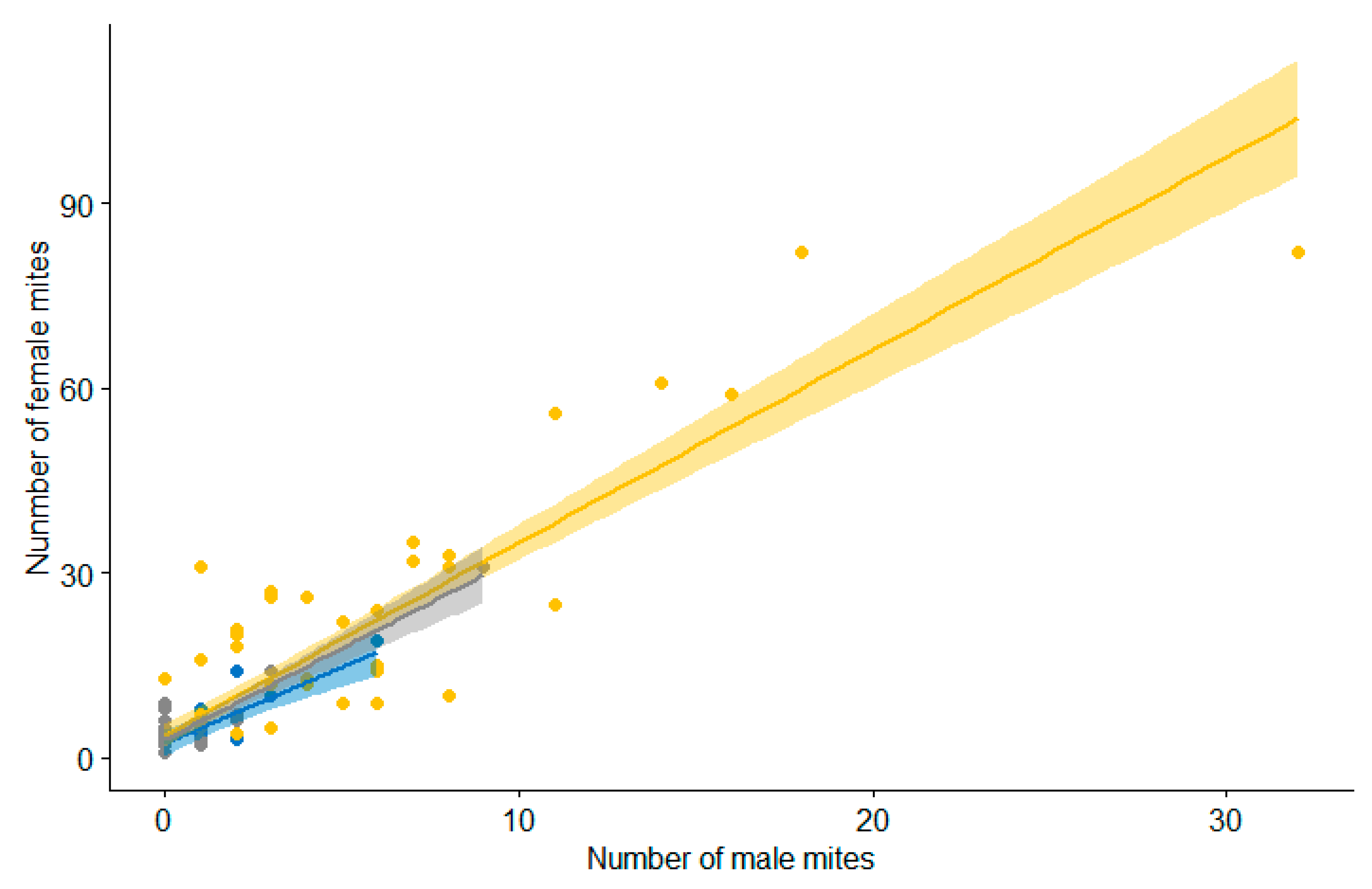
The number of female mites was positively correlated with the number of males in all three species of mites (M. melloi: τ = 0.73, p-value < 0.01; M. columbae: τ = 0.77, p-value < 0.01; M. streptopeliae: τ = 0.44; p-value < 0.01), with similar slopes (number of females per males of M. melloi: m = 3.12, M. columbae: m = 2.47 and M. streptopeliae: m = 2.97; Figure 6).
4. Discussion
This study reveals the species of rhinonyssids harboured by the sympatric host species C. livia domestica and S. decaocto, with an updated morphological description and an accurate epidemiological survey. The morphological information published until now on rhinonyssids is abundant but not very detailed, and the illustrations are not as exhaustive as the ones we provide. Furthermore, epidemiological studies are frequently based on the analyses of a few hosts, which makes the epidemiological survey inaccurate due to the low prevalence of the species studied [9,10,11,12,13] (but see [48]). We offer a detailed survey of the nasal mite species found in 250 C. livia domestica and 250 S. decaocto. Furthermore, we have detected M. columbae infecting S. decaocto for the first time.
The taxonomic assignment of the mite species isolated is still controversial. Since their original description in the 1950s, these species have been included in different genera, such as Neonyssus [42], Mesonyssus [23,35,36,37,38] and Tinaminyssus [24,39,40]. Lately they have been placed in the genus Tinaminyssus by some authors [25,41] and in Mesonyssus by others [2,14,21]. As suggested by the latter, in this paper the species are arranged in the genus Mesonyssus because the genus Tinaminyssus mainly includes species associated with Tinamiformes.
The morphological features of the mite species sampled match those previously described by other authors for Messonyssus melloi [21,22,25,26,27], M. columbae [14,21,22,23,24,25] and M. streptopeliae [18,20,21,27,28]. Nevertheless, the published descriptions and illustrations of these species tended to be simple and only focused on few characteristics. We offer a complete re-description together with highly detailed illustrations of the species mentioned, including all the features that other authors noted (Figure 3, Figure 4 and Figure 5). This information will help develop future investigations based on M. melloi, M. columbae and M. streptopeliae, making their identification easier and clearer.
It is noteworthy to state that a specimen of Mesonyssus was collected from S. decaocto showing the gnathosomal formula (2 + 2 + 2), completely different from the gnathosomal formula of the detected species (2 + 4 + 2) for M. melloi and M. streptopeliae, and (2 + 4 + 0) for M. columbae. Only two species worldwide follow the former formula: Mesonyssus juxtamelloi Pence et Canaris, 1976, and Mesonyssus carapachibeyus Dusbábek, 1969. However, these species and the mentioned specimen have different characteristics. M. juxtamelloi shows 32 setae on the dorsal idiosoma and 12 setae on the ventral idiosoma, and M. carapachibeyus presents three dorsal shields while the specimen collected has 21 dorsal setae, 10 ventral setae and two dorsal shields. Therefore, this individual could be a new species or an aberrant specimen. Deeper studies are needed to detect more individuals with similar characteristics.
To understand the relationship between parasites and hosts, it is crucial to obtain basic epidemiological information such as the prevalence, the intensity of infection, and the identity of the species involved. Until now, the most explored epidemiological aspect in the family Rhinonyssidae is the identification of the interacting species [48,49]. Nevertheless, in most of the species, mite information related with prevalence and intensity is almost unknown. We explored these epidemiological parameters focusing on the relationship of the nasal mite parasitic fauna of two avian species: C. livia domestica and S. decaocto, exploring 250 individuals of each one.
Traditionally, different studies on rhinonyssids have been performed throughout the world exploring prevalence; based on a scarce number of host specimens, these show low values. However, these studies include a high number of host species but a low number of specimens of each host species. For instance, Pence in 1973 [16] examined 1927 birds of 193 species and determined a total prevalence of 16.2%, Spicer in 1987 [50] studied 502 bird hosts of 103 species and found a prevalence of 17% and Knee et al. in 2008 [25] carried out a study of 450 birds of 154 species and showed a prevalence of 15%. However, each specific host-parasite relationship could have different values of prevalence, which could lead to inaccurate estimations if only a few individuals are examined. Thus, exploring specific relationships within a high number of hosts for each of these relationships is key.
We explored 250 C. livia domestica and 250 S. decaocto, finding Mesonyssus melloi and M. columbae in C. livia domestica and M. streptopeliae, and M. columbae in S. decaocoto. While most of these relationships have already been described, this is the first time that M. columbae has been detected in S. decaocoto. The sterilization of instruments prior to carrying out a new host analysis and the meticulousness during mounting, together with the fact that this species appears least frequently, leads us to think that it is not due to contamination. Thus, three possible explanations could account for this result: i) this relationship could be common in nature but the scarce epidemiological surveys together with its low prevalence have precluded its detection; ii) this is an incipient event of host switching, which has been found to possibly have important evolutionary consequences in other groups of avian mites [51]; iii) this encounter is a consequence of the sampling method, which could lead to the infection of doves with mites from pigeons. Some authors have found that indirect transmission can take place via water, perches and other contaminated surfaces [15], so the sampling method of employing traps with water and food to catch individuals could lead to this event. Nevertheless, it is important to note that these two host species usually share feeding habitats, which could lead to this event in natural conditions. Studies that explore a greater number of individuals of each host species would help uncover these kinds of relationships where prevalence could be too small.
Mesonyssus melloi was significantly more prevalent and produced higher infection intensities in C. livia domestica than M. columbae. Furthermore, as previously described [14,21], they are able to coexist, infecting the same host (Table 1). Streptopelia decaocoto are almost always infected by a single mite species, M. streptopeliae (with the exception previously described). Unfortunately, the comparison of these data with other publications is difficult. First, because most studies concern morphological description where the number of host specimens considered is low, this leads to inaccurate estimates. To our knowledge, this is the first paper that explores these estimates for M. streptopeliae. Regarding M. columbae, only three publications explore this kind of data: Crossley (1952), who reported a prevalence of 20% in 60 pigeons [52]; Wilson (1964), who found a prevalence of 7% after examining 15 pigeons [23]; Rojas et al. (2020), who found a prevalence of 1.9% after examining 262 pigeons in St. Petersburg (Russia) [48]. Meanwhile, we found a prevalence of 6%. In the case of M. melloi, there is more information available: Crossley (1952) found a prevalence of 25% after examining 60 pigeons in Texas (USA) [52]; Wilson (1964) found a prevalence of 7% after examining 15 pigeons in Indiana (USA) [23]; Rózsa (1990) found a prevalence of 23% or 42%, depending on the host population, in Hungary [53]; Foronda et al. (2004) found a prevalence of 10% in 50 pigeons in Tenerife (Spain) [54]; and Rojas et al. (2020) found a prevalence of 1.1% after examining 262 pigeons in St. Petersburg (Russia) [48]. Meanwhile, we found a prevalence of 34%, one of the highest ever reported. It is noteworthy to mention, even with the scarce information available, the high variation in the prevalence reported by different authors in the same host-parasite system. Similar concerns about the abundance and intensity of infection arise from information about intensities of infections, but in this case the research is much too scarce. To our knowledge, there are two single studies focused on M. melloi that report intensity of infection: Foronda et al. (2004), who reported a mean of 218.3 ± 117.3 mites per infected host [54], and Rózsa (1990), who reported a mean of 7.9 or 10.3 mites per host [53], depending on the host population. We report an infection intensity of 14.4 mites per infected host. It is clear that infection intensities are highly variable. It is important to highlight the study performed by Rózsa (1990) that compared the prevalence and intensities between two pigeon populations located in an urban and a farmland area, finding higher values of both parameters in the urban area [53]. Prevalence and intensities are highly variable not only among host-parasite relationships but also among populations. Thus, there must be important factors behind the difference in prevalence and intensities of infection that deserve to be explored. Deeper studies at a larger spatial scale analysing a higher number of hosts must be performed in order to reveal the factors accounting for these results.
The female-biased sex ratio is known by acarologists focused on Rhinonyssidae, and because of that, morphological keys are usually based on female characteristics. Biased sex ratios could be selected from different mating systems or population structures [55,56]. However, there are no studies about ratio or reproduction mode in the bibliography, even though these are key to predict and understand the mating system of these organisms. We found a relatively constant sex ratio within three specific parasite-host relationships, which could be due to similar pressures on the sex allocation. Nevertheless, specific studies on different species focused on the mating system must be developed to understand this ratio.
5. Conclusions
Knowledge of the relationships established between birds and their rhinonyssids parasites is scarce. The challenge of identification and studies based on a few host specimens could impede the development of further studies and lead to inaccurate results, respectively. We detailed morphological descriptions of three closely related species in order to facilitate their study. Furthermore, we estimated basic epidemiological parameters based on a high number of individuals in order to obtain accurate estimates. Deep differences between previously reported results and the ones shown here suggest that various factors could be behind these differences. Future studies will be essential to assess the mechanisms ruling the variation in prevalence and intensity.
Author Contributions
Conceptualization, M.d.R. and J.V.; methodology, I.D. and M.d.R.; software, J.V.; validation, M.d.R. and J.V.; formal analysis, J.V.; investigation, M.d.R.; resources, M.d.R. and I.D.; data curation, M.d.R. and J.V.; writing—original draft preparation, M.d.R. and J.V.; writing—review and editing, M.d.R., I.D. and J.V.; visualization, M.d.R., I.D. and J.V.; supervision, M.d.R.; project administration, M.d.R.; funding acquisition, M.d.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by “V Plan Propio de Investigación of the University of Seville, Spain” and by “Ministero de Ciencia, Innovación y Universidades (MCIU/AEI/FEDER, UE)” project number PGC2018-097426-B-C22.
Institutional Review Board Statement
The study was conducted according to the guide-lines of the Declaration of Helsinki, and approved by the Institutional Review Board of animal control program of the Seville City Council and approved by the “Consejo Municipal de Protec-ción y Bienestar Animal (COMBA)”.
Data Availability Statement
Not applicable.
Acknowledgments
We thank the “Centro Municipal Zoosanitario of Sevilla” and especially Francisco Peña Fernández and Rafael Cuadrado Nieto for providing the samples.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vitzthum, H.G. Milben aus der Nasenhöhle von Vögeln. J. Ornithol. 1935, 83, 563–587. [Google Scholar] [CrossRef]
- Dimov, I.D. Morphology of Nasal Mites of the Family Rhinonyssidae—Scanning Electron Microscopy Saint Petersburg 2020; LLC Zhigulin: Saint Petersburg, Russia, 2020; ISBN 9785604468708. [Google Scholar]
- Dimov, I.D. Rhinonyssidosis avium. Vetpharma 2011, 3, 88–90. [Google Scholar]
- Stephan, S.A.R.; Kaschula, V.R.; Canham, A.S. Respiratory acariasis of canaries. J. S. Afr. Vet. Assoc. 1950, 21, 103–107. [Google Scholar]
- Tidemann, S.C.; McOrist, S.; Woinarski, J.C.Z.; Freeland, W.J. Parasitism of wild Gouldian finches (Erythrura gouldiae) by the air-sac mite Sternostoma tracheacolum. J. Wildl. Dis. 1992, 28, 80–84. [Google Scholar] [CrossRef]
- Moro, C.V.; Chauve, C.; Zenner, L. Experimental infection of Salmonella Enteritidis by the poultry red mite, Dermanyssus gallinae. Vet. Parasitol. 2007, 146, 329–336. [Google Scholar] [CrossRef]
- Sommer, D.; Heffels-Redmann, U.; Köhler, K.; Lierz, M.; Kaleta, E.F. Role of the poultry red mite (Dermanyssus gallinae) in the transmission of avian influenza A virus. Tierarztliche Praxis Ausgabe G Grosstiere/Nutztiere 2016, 44, 26–33. [Google Scholar] [CrossRef]
- Osuna-Mascaró, C.; Doña, J.; Johnson, K.P.; Esteban, R.; De Rojas, M. Complete mitochondrial genomes and bacterial metagenomic data from two species of parasitic avian nasal-mites (Rhinonyssidae: Mesostigmata). Front. Ecol. Evol. 2020, 8, 142. [Google Scholar] [CrossRef]
- Feider, Z.; Mironescu, I. Rhinonyssides parasites des oiseaux des genres Podiceps et Tringa. Anaele Stiint. Univ. Al. I. Cuza Iasi Sect. II A 1972, 18, 317–330. [Google Scholar]
- Guevara Benítez, D.C.; Úbeda Ontiveros, J.M. Ácaros del género Ptilonyssus Berlese y Trouessart, 1889 (mesostigmata: Rhynonyssidae) parásitos de fosas nasales de paseriformes españolas. I. Ptilonyssus hirsti (Castro y Pereira, 1947) Pereira y Castro, 1949. Rev. Iber. Parasitol. 1978, 38, 591–614. [Google Scholar]
- Kadosaka, T.; Kaneko, K.; Asanuma, K. A new species and new records of avian nasal mites (Acarina: Rhinonyssidae) from Japan. Jpn. J. Sanit. Zool. 1987, 38, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Spicer, G.S. Nasal mites from birds of a Guatemalan cloud forest (Acarina: Rhinonyssidae). J. Parasitol. 1984, 70, 794–802. [Google Scholar] [CrossRef]
- Da Silva, D.S.; Scheer, S.; Muller, G. Nasal mites (Mesostigmata, Rhinonyssidae) in Sternidae (Aves: Charadriiformes) on the southern coast of Brazil. Rev. Bras. Parasitol. Veterinária 2018, 27, 109–111. [Google Scholar] [CrossRef] [Green Version]
- Dimov, I.D. Kleshchi-rinonissidy ptic Severo-Zapada Ross; LLC Zhigulin: Saint Petersburg, Russia, 2018; ISBN 9785600021778. [Google Scholar]
- Bell, P.J. The life history and transmission biology of Sternostoma tracheacolum Lawrence (Acari: Rhinonyssidae) associated with the Gouldian finch Erythrura gouldiae. Exp. Appl. Acarol. 1996, 20, 323–341. [Google Scholar] [CrossRef]
- Pence, D.B. The nasal mites of birds from Louisiana. IX. Synopsis. J. Parasitol. 1973, 881–892. [Google Scholar] [CrossRef]
- Baulieu, F.; Knee, W.; Nowell, V.; Schwarzfeld, M.; Lindo, Z.; Behan-Pelletier, V.M.; Lumley, L.; Young, M.R.; Smith, I.; Proctor, H.C. Acari of Canada. Zookeys 2019, 77. [Google Scholar] [CrossRef]
- Fain, A. Les Rhinonyssides parasites des Pigeons. Rev. Zool. Bot. Afr. 1962, 65, 305–324. [Google Scholar]
- Bonnefoy, X.; Kampen, H.; Sweeney, K. Public Health Significance of Urban Pests; World Health Organization: Geneva, Switzerland, 2008; ISBN 9289071885. [Google Scholar]
- Fain, A. Diagnoses d’acariens parasites nouveaux. Rev. Zool. Bot. Afr 1965, 72, 152–160. [Google Scholar]
- Butenko, O.M. Rhinonyssid Mites of Non-Passerine Birds of the USSR; Academy of Sciences of USSR: Moscow, Russia, 1984. [Google Scholar]
- Fain, A. Les acariens des familles Epidermoptidae et Rhinonyssidae parasites des fosses nasales d’oiseaux au Ruanda-Urundi et au Congo belge. Proc. Ann. Musée R. Congo Belge 1957, 60, 1–176. [Google Scholar]
- Wilson, N. New records and descriptions of Rhinonyssidae mostly from New Guinea. Pac. Insects 1964, 6, 357–388. [Google Scholar]
- Pence, D.B. Keys, Species and Host List, and Bibliography for Nasal Mites of North. American Birds (Acarina-Rhinonyssinae, Turbinoptinae, Speleognathinae, and Cytoditidae); Texas Tech Press: Lubbock, TX, USA, 1975. [Google Scholar]
- Knee, W.; Proctor, H.; Galloway, T. Survey of nasal mites (Rhinonyssidae, Ereynetidae, and Turbinoptidae) associated with birds in Alberta and Manitoba, Canada. Can. Entomol. 2008, 140, 364–379. [Google Scholar] [CrossRef]
- Domrow, R. Bird nasal mites in New Zealand. Tuatara 1972, 19, 99–100. [Google Scholar]
- Shumilo, R.; Lunkashu, M. Rhinonyssid Mites from Southeastern URSS; Editorial Academy of Sciences of MSSR; Academy of Sciences of USSR: Moscow, Russia, 1970. [Google Scholar]
- Fain, A.; Herin, A.; Puylaert, F. Notes sur les Acariens nasicoles d’Oiseaux du Cameroun. Rev. Zool. Afr. 1977, 91, 385–396. [Google Scholar]
- Rózsa, L.; Reiczigel, J.; Majoros, G. Quantifying parasites in samples of hosts. J. Parasitol. 2000, 86, 228–232. [Google Scholar] [CrossRef]
- R Core Team. R: A language and environment for statistical computing. R Found. Stat. Comput. Vienna Austria. 2020. Available online: http://www.R-project.org/ (accessed on 28 December 2020).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 3319242776. [Google Scholar]
- Kassambara, A. ggpubr: “ggplot2” Based Publication Ready Plots. Available online: https://cran.r-project.org/package=ggpubr (accessed on 28 December 2020).
- Devleesschauwer, B.; Torgerson, P.; Charlier, J.; Levecke, B.; Praet, N.; Roelandt, S.; Smit, S.; Dorny, P.; Berkvens, D.; Speybroeck, N. Prevalence: Tools for prevalence assessment studies. R Packag. Version. 2015. Available online: http://cran.r-project.org/package=prevalence (accessed on 28 December 2020).
- Kohl, M. MKinfer: Inferential Statistics. R Packag. Version 0.5. 2020. Available online: http://cran.r-project.org/package=prevalence (accessed on 28 December 2020).
- Alec, F. Sur le statut de quelques espèces de Rhinonyssidae décrites par Hirst. Rev. Zool. Bot. Afr. 1960, 59, 310–314. [Google Scholar]
- Domrow, R. Some mite paraites of Australian birds. Proc. Linn. Soc. N. S. W. 1966, 90, 190–217. [Google Scholar]
- Fain, A.; Aitken, T.H.G. Les acariens parasites nasicoles des oiseaux de Trinidad (Indes Occidentales): I. Rhinonyssidae mesostigmates. Bull. Ann. Soc. R. D’Entomologie Belg. 1967, 43, 1–44. [Google Scholar]
- Sixl, W. Nasale Milben einheimischer Vögel. Mitteilungen Naturwissenschaftlichen Vereines Steiermark 1969, 99, 217–218. [Google Scholar]
- Strandtmann, R.W.; Wharton, G.W. A Manual of Mesostigmatid Mites Parasitic on Vertebrates; Institute of Acarology, University Maryland: College Park, MD, USA, 1958. [Google Scholar]
- Domrow, R. The nasal mites of Queensland birds (Acari: Dermanyssidae, Ereynetidae, and Epidermoptidae). Proc. Linnean Soc. N. S. W. 1969, 93, 297–426. [Google Scholar]
- Úbeda, J.M.; De Rojas, M.; Mora, M.D.; Guevara, D.C. Nasal mites (Mesostigmata, Rhinonyssidae) parasitic on columbiform birds from the Iberian Peninsula. Comparison of ribosomal ITS 1 region for species differentiation. Boletín la Real Soc. Española Hist. Nat. Sección Biológica 2018, 98, 65–75. [Google Scholar]
- Fain, A. Les acariens de la famille Rhinonyssidae Vitzthum 1935 parasites des fosses nasales des oiseaux au Ruanda-Urundi. Rev. Zool. Bot. Afri. 1956, 53, 131–157. [Google Scholar]
- Castro, M.P. de Reestruturação genérica da familia Rhinonyssidae Vitzthum, 1935 (Acari: Mesostigmata: Gamasides) e descrição de algumas espécies novas. Arq. Inst. Biol. (Sao Paulo) 1948, 18, 253–284. [Google Scholar]
- Pereira, C.; Castro, M.P. Revisão da subfamília Ptilonyssinae Castro, 1948 (Acari: Mesostigmata: Rhinonyssidae) com a descrição de algumas espécies novas. Arq. Inst. Biol (Sao. Paulo) 1949, 19, 218–235. [Google Scholar]
- Fain, A.; Sixl, W.; Moritsch, C. The nasal mites of the family Rhinonyssidae with description of a new species (Acarina). Mitteilungen der Abteilung Zool. Bot. Landesmuseum Joanneum Graz 1974, 3, 1–9. [Google Scholar]
- Feider, Z. Neonyssus hirsutus n.sp. (Rhinonyssidae, Parasitiformes), un nou parazit in cavitatea nazala la porumbelul domestic. Stidii. Cerc. Sti. Iasi. 1962, 13, 57–68. [Google Scholar]
- Crossley, D.A. A new species of nasal mite, Neonyssus (Neonyssus) columbae, from the pigeon (Acarina, Mesostigmata, Rhinonyssidae). Proc. Entomol. Soc. Wash 1950, 52, 309–312. [Google Scholar]
- De Rojas, M.; Doña, J.; Dimov, I. A comprehensive survey of Rhinonyssid mites (Mesostigmata: Rhinonyssidae) in Northwest Russia: New mite-host associations and prevalence data. Biodivers. Data J. 2020, 8, e49535. [Google Scholar] [CrossRef]
- Beron, P. Acarorum Catalogus VI. Order Mesostigmata. Gamasina: Dermanyssoidea (Rhinonyssidae, Spinturnicidae). In National Museum of Natural History, Sofia Bulgarian Academy of Sciences; Pensoft Publishers: Sofia, Bulgaria, 2020. [Google Scholar] [CrossRef]
- Spicer, G.S. Prevalence and host-parasite list of some nasal mites from birds (Acarina: Rhinonyssidae, Speleognathidae). J. Parasitol. 1987, 73, 259–264. [Google Scholar] [CrossRef]
- Doña, J.; Sweet, A.D.; Johnson, K.P.; Serrano, D.; Mironov, S.; Jovani, R. Cophylogenetic analyses reveal extensive host-shift speciation in a highly specialized and host-specific symbiont system. Mol. Phylogenet. Evol. 2017, 115, 190–196. [Google Scholar] [CrossRef]
- Crossley, D.A. Two new nasal mites from columbiform birds. J. Parasitol. 1952, 38, 385–390. [Google Scholar] [CrossRef]
- Rózsa, L. The ectoparasite fauna of feral pigeon populations in Hungary. Parasitol. Hungarica 1990, 23, 115–119. [Google Scholar]
- Foronda, P.; Valladares, B.; Rivera-Medina, J.A.; Figueruelo, E.; Abreu, N.; Casanova, J.C. Parasites of Columba livia (Aves: Columbiformes) in Tenerife (Canary Islands) and their role in the conservation biology of the laurel pigeons. Parasite 2004, 11, 311–316. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, W.D. Extraordinary sex ratios. Science 1967, 156, 477–488. [Google Scholar] [CrossRef]
- Herre, E.A. Optimality, plasticity and selective regime in fig wasp sex ratios. Nature 1987, 329, 627–629. [Google Scholar] [CrossRef]
Figure 1.
Sampled districts of Seville. Districts not sampled are blue, while sampled districts are green; “n” shows the number of locations sampled per district.
Figure 1.
Sampled districts of Seville. Districts not sampled are blue, while sampled districts are green; “n” shows the number of locations sampled per district.

Figure 2.
Female mite species from Columba livia domestica and Streptopelia decaocto: (a) Mesonyssus melloi; (b) Mesonyssus columbae; (c) Mesonyssus streptopeliae.
Figure 2.
Female mite species from Columba livia domestica and Streptopelia decaocto: (a) Mesonyssus melloi; (b) Mesonyssus columbae; (c) Mesonyssus streptopeliae.

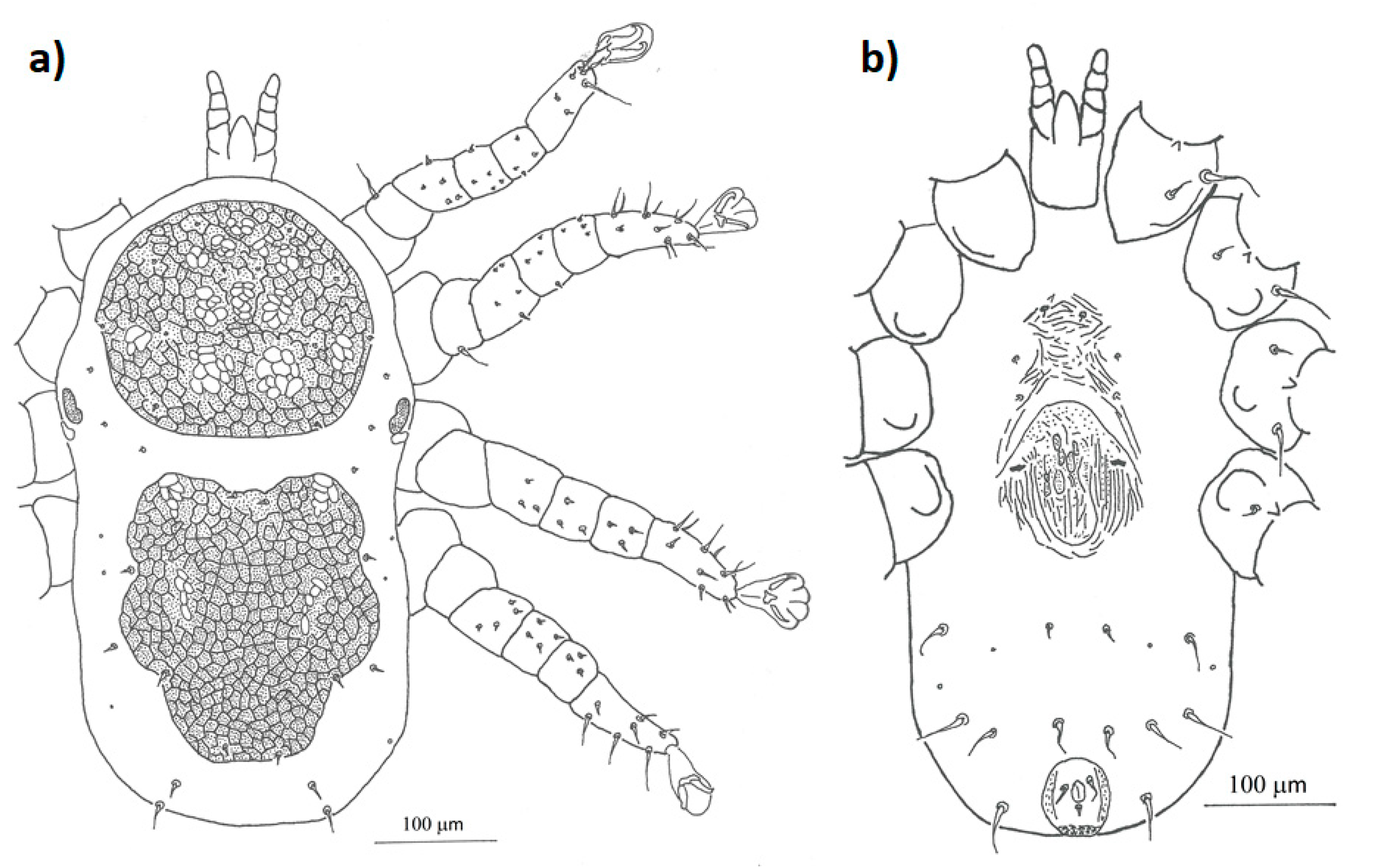
Figure 3.
Detailed drawing of the morphological characteristics of female Mesonyssus melloi: (a) dorsal view; (b) ventral view.
Figure 3.
Detailed drawing of the morphological characteristics of female Mesonyssus melloi: (a) dorsal view; (b) ventral view.

Figure 4.
Detailed drawing of the morphological characteristics of female Mesonyssus columbae: (a) dorsal view; (b) ventral view.
Figure 4.
Detailed drawing of the morphological characteristics of female Mesonyssus columbae: (a) dorsal view; (b) ventral view.

Figure 5.
Detailed drawing of the morphological characteristics of female Mesonyssus streptopeliae: (a) dorsal view; (b) ventral view.
Figure 5.
Detailed drawing of the morphological characteristics of female Mesonyssus streptopeliae: (a) dorsal view; (b) ventral view.

Figure 6.
Number of females per male of each parasitic mite species. The solid line represents the linear regression line, the shadows represent the 95% confidence interval and the dots represent each individual datum for each mite species, depending on the colour: yellow represents M. melloi, blue represents M. columbae and grey represents M. streptopeliae.
Figure 6.
Number of females per male of each parasitic mite species. The solid line represents the linear regression line, the shadows represent the 95% confidence interval and the dots represent each individual datum for each mite species, depending on the colour: yellow represents M. melloi, blue represents M. columbae and grey represents M. streptopeliae.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Epidemiological information of each parasitic mite species in each host species. The table shows the prevalence with 95% confidence intervals (in brackets) and the mean intensities with their standard error and the range of mites collected per host. Total number of each mite species collected per host species (n), total number of male and female mites of each species and total number of infested hosts with each mite species is also shown.
Table 1.
Epidemiological information of each parasitic mite species in each host species. The table shows the prevalence with 95% confidence intervals (in brackets) and the mean intensities with their standard error and the range of mites collected per host. Total number of each mite species collected per host species (n), total number of male and female mites of each species and total number of infested hosts with each mite species is also shown.
| Host Species | Parasite Species (n) | Number of Males | Number of Females | Number of Infested Hosts | Prevalence (IC95%) | Mean intensity±SE (Range) |
| Columba livia domestica | M. melloi (n = 1223) | 219 | 1004 | 85 | 0.34 (0.28–0.40) | 14.4 ± 2.39 (0–114) |
| M. columbae (n = 110) | 22 | 88 | 14 | 0.06 (0.03–0.09) | 7.86 ± 2.08 (0–25) | |
| Streptopelia decaocto | M. streptopeliae (n = 158) | 20 | 138 | 26 | 0.10 (0.07–0.15) | 6.08 ± 1.52 (0–40) |
| M. columbae (n = 3) | - | 3 | 1 | 0.004 (0.0001–0.22) | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Veiga, J.; Dimov, I.; de Rojas, M. Endoparasitic Mites (Rhinonyssidae) on Urban Pigeons and Doves: Updating Morphological and Epidemiological Information. Diversity 2021, 13, 11. https://0-doi-org.brum.beds.ac.uk/10.3390/d13010011
AMA Style
Veiga J, Dimov I, de Rojas M. Endoparasitic Mites (Rhinonyssidae) on Urban Pigeons and Doves: Updating Morphological and Epidemiological Information. Diversity. 2021; 13(1):11. https://0-doi-org.brum.beds.ac.uk/10.3390/d13010011
Chicago/Turabian StyleVeiga, Jesús, Ivan Dimov, and Manuel de Rojas. 2021. "Endoparasitic Mites (Rhinonyssidae) on Urban Pigeons and Doves: Updating Morphological and Epidemiological Information" Diversity 13, no. 1: 11. https://0-doi-org.brum.beds.ac.uk/10.3390/d13010011
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.