Minimizing Potential Allee Effects in Psittacine Reintroductions: An Example from Puerto Rico

Abstract
:1. Introduction
2. Materials and Methods
2.1. General Release Methodologies
2.1.1. Traditional Release
2.1.2. Precision Release
2.1.3. Soft Release Type A
2.1.4. Soft Release Type B
2.2. Data Analyses and Reporting
3. Results
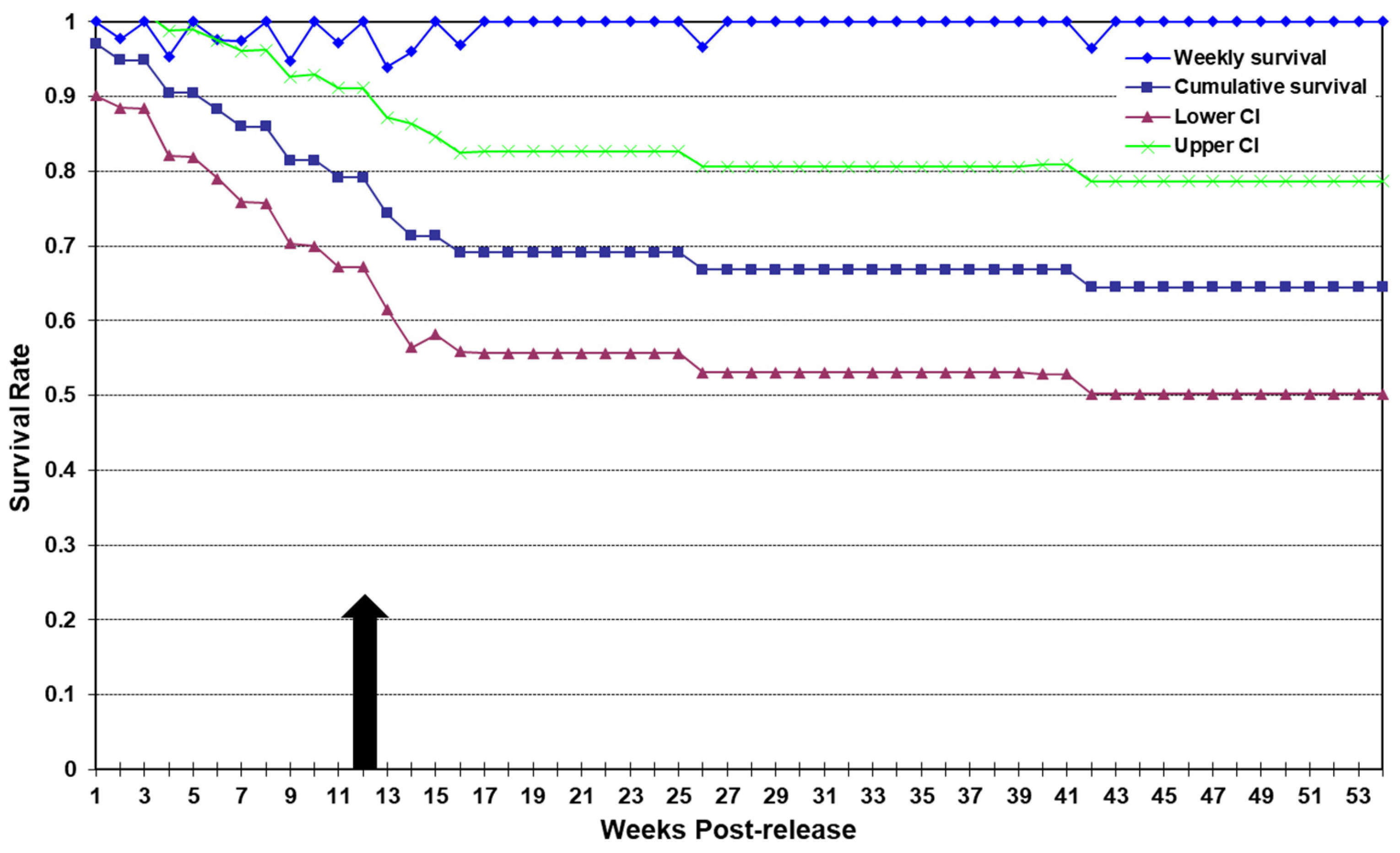
3.1. Traditional Releases
3.1.1. Survival
3.1.2. Site Fidelity
3.1.3. Flock Cohesion
3.1.4. Prompt Reproduction
3.2. Precision Releases
3.2.1. Survival
3.2.2. Site Fidelity
3.2.3. Flock Cohesion
3.2.4. Prompt Reproduction
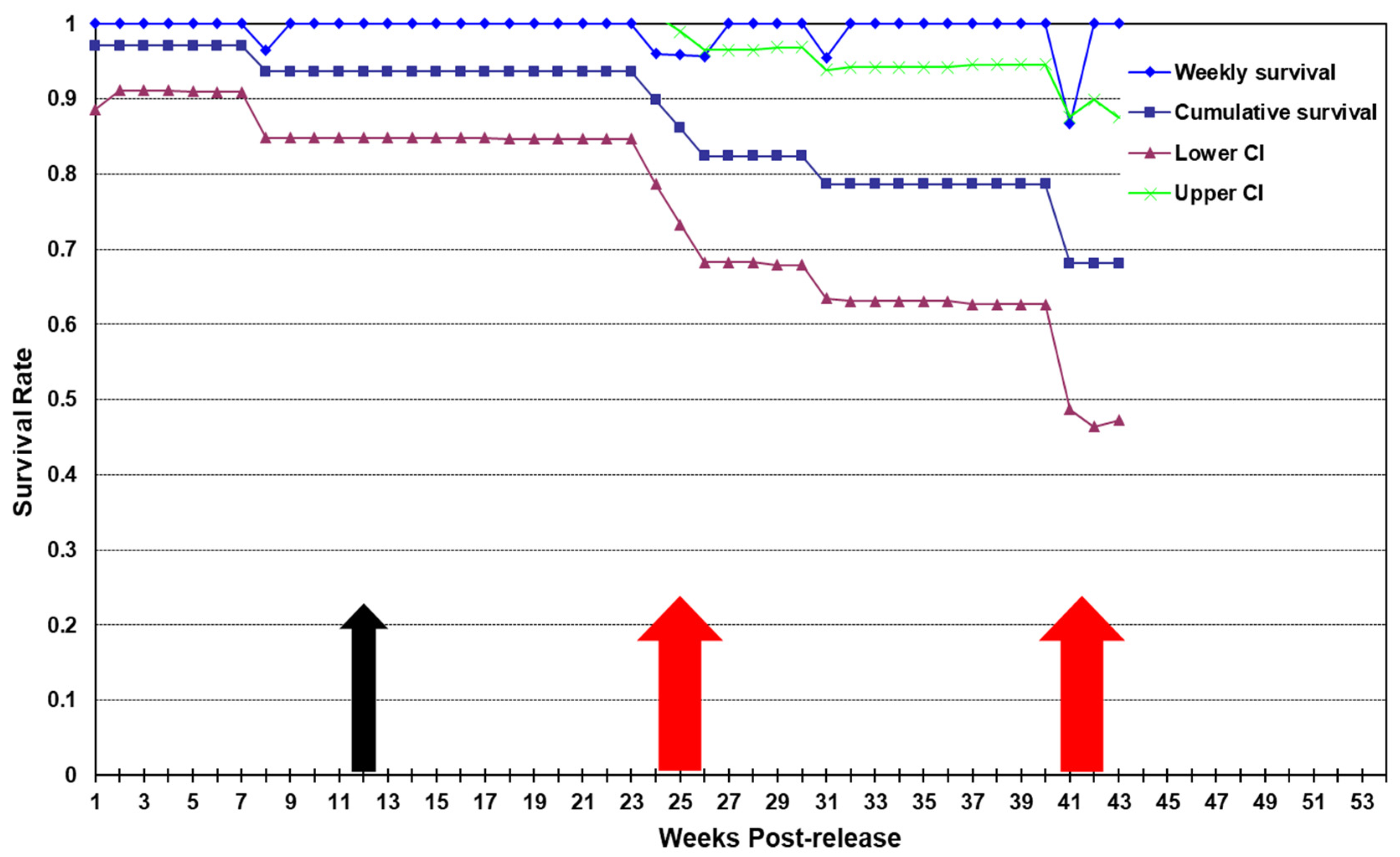
3.3. Soft Release Type A
3.3.1. Survival
3.3.2. Site Fidelity
3.3.3. Flock Cohesion
3.3.4. Prompt Reproduction
3.4. Soft Release Type B
3.4.1. Survival
3.4.2. Site Fidelity
3.4.3. Flock Cohesion
3.4.4. Prompt Reproduction
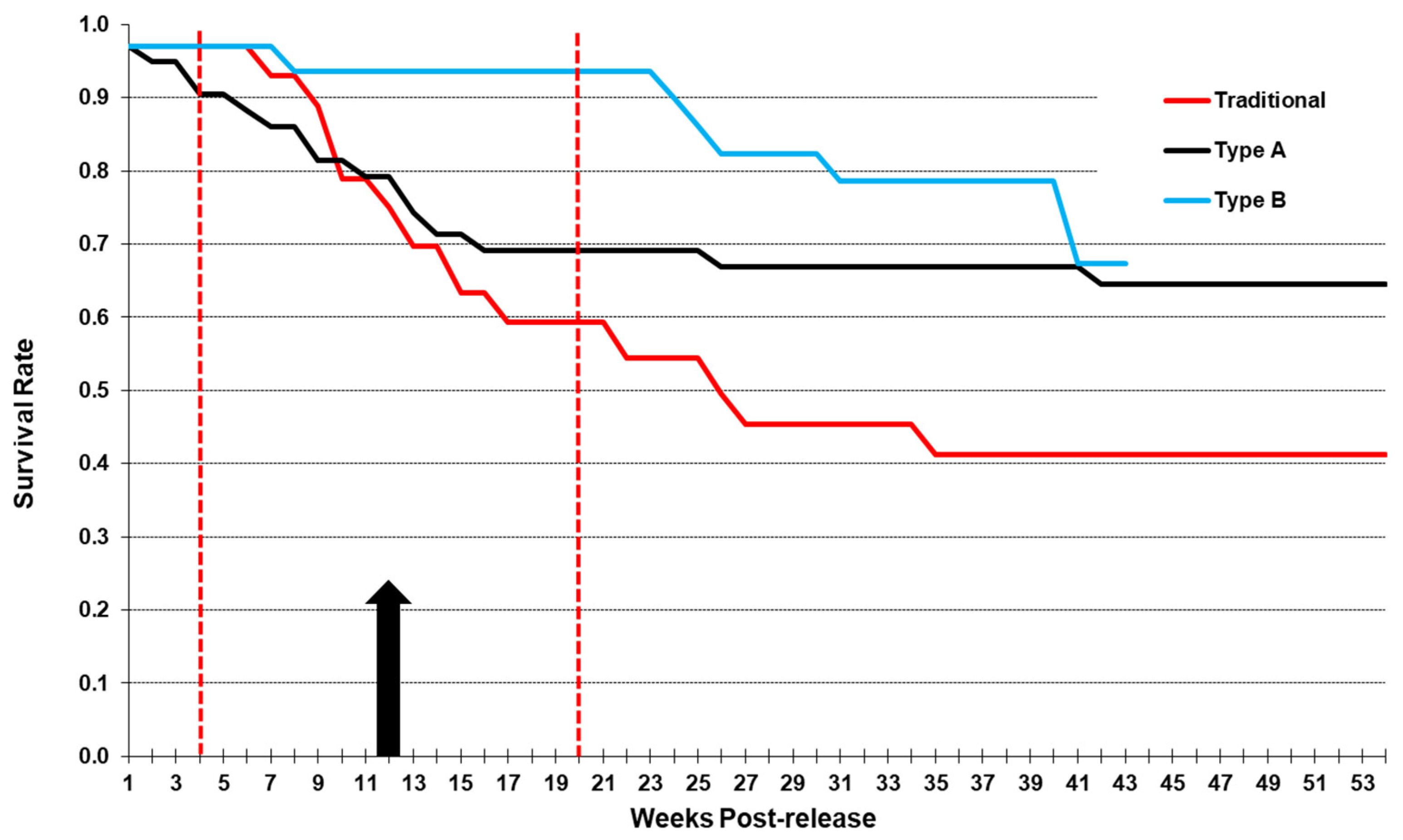
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IUCN Red List of Threatened Species. Version 2020. Available online: http://www.iucnredlist.org/apps/redlist (accessed on 26 October 2020).
- White, T.H., Jr.; Collar, N.J.; Moorhouse, R.J.; Sanz, V.; Stolen, E.D.; Brightsmith, D.J. Psittacine reintroductions: Common denominators of success. Biol. Conserv. 2012, 148, 106–115. [Google Scholar] [CrossRef]
- Snyder, N.F.R.; Koenig, S.E.; Koschmann, J.; Snyder, H.A.; Johnson, T.B. Thick-billed Parrot releases in Arizona. Condor 1994, 96, 845–862. [Google Scholar]
- Brightsmith, D.J.; Hilburn, J.; del Campo, A.; Boyd, J.; Frisius, M.; Frisius, R.; Janik, D.; Guillén, F. The use of hand-raised psittacines for reintroduction: A case study of scarlet macaws (Ara macao) in Peru and Costa Rica. Biol. Conserv. 2005, 121, 465–472. [Google Scholar] [CrossRef]
- Estrada, A. Reintroduction of the scarlet macaw (Ara macao cyanoptera) in the tropical rainforests of Palenque, Mexico: Project design and first-year progress. Trop. Conserv. Sci. 2014, 7, 342–364. [Google Scholar] [CrossRef]
- Wolf, C.M.; Garland, T., Jr.; Griffith, B. Predictors of avian and mammalian translocation success: Reanalysis with phylogenetically independent contrasts. Biol. Conserv. 1998, 86, 243–255. [Google Scholar] [CrossRef]
- Moseby, K.E.; Read, J.L.; Paton, D.C.; Copley, P.; Hill, B.M.; Crisp, H.A. Predation determines the outcome of 10 reintroduction attempts in arid South Australia. Biol. Conserv. 2011, 144, 2863–2872. [Google Scholar] [CrossRef]
- Ewen, J.G.; Armstrong, D.P.; Parker, K.A.; Seddon, P.J. (Eds.) Reintroduction Biology: Integrating Science and Management; John Wiley and Sons: West Sussex, UK, 2012. [Google Scholar]
- Deredec, A.; Courchamp, F. Importance of the Allee effect for reintroductions. Écoscience 2007, 14, 440–451. [Google Scholar] [CrossRef]
- Dennis, B. Allee effects in stochastic populations. Oikos 2002, 96, 389–401. [Google Scholar] [CrossRef] [Green Version]
- Stephens, P.A.; Sutherland, W.J.; Freckleton, R.P. What is the Allee effect? Oikos 1999, 87, 185–190. [Google Scholar] [CrossRef] [Green Version]
- Stephens, P.A.; Sutherland, W.J. Consequences of the Allee effect for behaviour, ecology and conservation. Trends Ecol. Evol. 1999, 14, 401–405. [Google Scholar] [CrossRef]
- Collazo, J.A.; Fackler, P.F.; Pacific, K.; White, T.H., Jr.; Llerandi-Roman, I.; Dinsmore, S.J. Optimal allocation of captive-reared Puerto Rican Parrots: Decisions when divergent dynamics characterize managed populations. J. Wildl. Manag. 2013, 77, 1124–1134. [Google Scholar] [CrossRef]
- Somers, M.J.; Graf, J.A.; Szkman, M.; Slotow, R.; Gusset, M. Dynamics of a small re-introduced population of wild dogs over 25 years: Allee effects and the implications of sociality for endangered species’ recovery. Oecologia 2008, 158, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Courchamp, F.; Clutton-Brock, T.; Grenfell, B. Inverse density dependence and the Allee effect. Trends Ecol. Evol. 1999, 14, 405–410. [Google Scholar] [CrossRef]
- Armstrong, D.P.; Wittmer, H.U. Incorporating Allee effects into reintroduction strategies. Ecol. Res. 2011, 26, 687–695. [Google Scholar] [CrossRef]
- White, T.H., Jr.; Collazo, J.A.; Dinsmore, S.J.; Llerandi-Román, I. Niche restriction and conservatism in a neotropical psittacine: The case of the Puerto Rican parrot. In Habitat Loss: Causes, Effects on Biodiversity and Reduction Strategies; Devore, B., Ed.; Nova Science Publishers: New York, NY, USA, 2014; pp. 1–83. Available online: https://novapublishers.com/wp-content/uploads/2019/05/978-1-63117-231-1_ch1.pdf (accessed on 14 November 2020).
- Snyder, N.F.R.; Wiley, J.W.; Kepler, C.B. The Parrots of Luquillo: Natural History and Conservation of the Puerto Rican Parrot; Western Foundation of Vertebrate Zoology: Los Angeles, CA, USA, 1987. [Google Scholar]
- Collazo, J.A.; White, T.H., Jr.; Vilella, F.J.; Guerrero, S.A. Survival of captive-reared Hispaniolan Parrots released in Parque Nacional del Este, Dominican Republic. Condor 2003, 105, 198–207. [Google Scholar] [CrossRef]
- White, T.H., Jr.; Collazo, J.A.; Vilella, F.J. Survival of captive-reared Puerto Rican Parrots released in the Caribbean National Forest. Condor 2005, 107, 426–434. [Google Scholar] [CrossRef]
- White, T.H., Jr.; Abreu-González, W. Dummy transmitters for pre-release acclimation of captive-reared birds. Reintro. News 2007, 26, 28–30. [Google Scholar]
- USFWS. Recovery Plan for the Puerto Rican Parrot (Amazona vittata); United States Fish and Wildlife Service: Region 4, Atlanta, GA, USA, 2009.
- Llerandi-Román, I.; White, T.H., Jr.; Garcia-Bermúdez, M.A. Successful reintroduction of Puerto Rican parrots in the moist karst region of northcentral Puerto Rico. Unpubl. Abstract. In Proceedings of the 8th Caribbean Biodiversity Congress, Universidad Autónoma de Santo Domingo, Santo Domingo, Dominican Republic, 28 January–1 February 2014. [Google Scholar]
- The 2012 IUCN Guidelines for Reintroductions and Other Conservation Translocations. Available online: http://www.iucnsscrsg.org (accessed on 2 November 2020).
- White, T.H., Jr.; Abreu-González, W.; Toledo-González, M.; Torres-Báez, P. From the field: Artificial nest cavities for Amazona parrots. Wildl. Soc. Bull. 2005, 33, 756–760. [Google Scholar] [CrossRef]
- White, T.H., Jr.; Brown, G.G., Jr.; Collazo, J.A. Artificial cavities and nest site selection by Puerto Rican Parrots: A multiscale assessment. Avian Conserv. Ecol. 2006, 1, 5. [Google Scholar] [CrossRef] [Green Version]
- Oehler, D.A.; Boodoo, D.; Plair, B.; Kuchinski, K.; Campbell, M.; Lutchmendial, G.; Ramsubage, S.; Maruska, E.J.; Malowski, S. Translocation of Blue and Gold Macaw Ara ararauna into its historical range on Trinidad. Bird Conserv. Int. 2001, 11, 29–141. [Google Scholar] [CrossRef] [Green Version]
- Sanz, V.; Grajal, A. Successful reintroduction of captive-raised Yellow-shouldered Amazon Parrots on Margarita Island, Venezuela. Conserv. Biol. 1998, 12, 430–441. [Google Scholar] [CrossRef]
- Harrington, D.P.; Fleming, T.R. A class of rank test procedures for censored survival data. Biometrika 1982, 69, 553–566. [Google Scholar] [CrossRef]
- Caro, T. Antipredator Defenses in Birds and Mammals; Univ. of Chicago Press: Chicago, IL, USA, 2005. [Google Scholar]
- Griffith, B.; Scott, J.M.; Carpenter, J.W.; Reed, C. Translocation as a species conservation tool. Science 1989, 245, 477–480. [Google Scholar] [CrossRef] [PubMed]
- Green, R.E. The influence of numbers released on the outcome of attempts to introduce exotic bird species to New Zealand. J. Animal Ecol. 1997, 66, 25–35. [Google Scholar] [CrossRef]
- Gascoigne, J.C.; Lipsius, R.N. Allee effects driven by predation. J. Appl. Ecol. 2004, 41, 801–810. [Google Scholar] [CrossRef]
- Angulo, E.; Roemer, G.W.; Berek, L.; Gasco, J.; Courchamp, F. Double Allee effects and extinction in the Island fox. Conserv. Biol. 2007, 21, 1082–1091. [Google Scholar] [CrossRef] [Green Version]
- Waas, J.R.; Caulfield, M.; Colgan, P.W.; Boag, P.T. Colony sound facilitates sexual and agonistic activities in royal penguins. Animal Behav. 2000, 60, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Pickering, S.; Creighton, E.; Stephens-Woods, B. Flock size and breeding success in Flamingos. Zoo Biol. 1992, 11, 229–234. [Google Scholar] [CrossRef]
- McCarthy, M.A. The Allee effect, finding mates and theoretical models. Ecol. Model. 1997, 103, 99–102. [Google Scholar] [CrossRef]
- Instituto Chico Mendes de Conservação da Biodiversidade. Executive Summary of the National Action Plan for the Spix’s Macaw Conservation; Instituto Chico Mendes de Conservação da Biodiversidade: Brasilia, Brasil, 2012.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Release Type | Survival 1-Year | Site Fidelity | Flock Cohesion | Prompt Reproduction |
|---|---|---|---|---|
| Traditional | 41% | 30–40% | 20–25% | 0 |
| Precision | 59% | 22–28% | 11–17% | 0 |
| Soft Release A | 64% | 65% | 100% | 18% |
| Soft Release B | 68% 1 | 67% 1 | 95% 1 | 25% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
White, T.H., Jr.; Abreu, W.; Benitez, G.; Jhonson, A.; Lopez, M.; Ramirez, L.; Rodriguez, I.; Toledo, M.; Torres, P.; Velez, J. Minimizing Potential Allee Effects in Psittacine Reintroductions: An Example from Puerto Rico. Diversity 2021, 13, 13. https://0-doi-org.brum.beds.ac.uk/10.3390/d13010013
White TH Jr., Abreu W, Benitez G, Jhonson A, Lopez M, Ramirez L, Rodriguez I, Toledo M, Torres P, Velez J. Minimizing Potential Allee Effects in Psittacine Reintroductions: An Example from Puerto Rico. Diversity. 2021; 13(1):13. https://0-doi-org.brum.beds.ac.uk/10.3390/d13010013
Chicago/Turabian StyleWhite, Thomas H., Jr., Wilfredo Abreu, Gabriel Benitez, Arelis Jhonson, Marisel Lopez, Limary Ramirez, Iris Rodriguez, Miguel Toledo, Pablo Torres, and Jafet Velez. 2021. "Minimizing Potential Allee Effects in Psittacine Reintroductions: An Example from Puerto Rico" Diversity 13, no. 1: 13. https://0-doi-org.brum.beds.ac.uk/10.3390/d13010013