
FloCan—A Revised Checklist for the Flora of the Canary Islands



1
Department of Biogeography, University of Bayreuth, Universitätsstr. 30, 95440 Bayreuth, Germany
2
Geographical Institute Bayreuth (GIB), 95440 Bayreuth, Germany
3
Bayreuth Center of Ecology and Environmental Science (BayCEER), 95440 Bayreuth, Germany
4
Botanical Garden, Friedrich-Alexander-University Erlangen-Nürnberg, Loschgestraße 1, 91054 Erlangen, Germany
*
Author to whom correspondence should be addressed.
Diversity 2021, 13(10), 480; https://0-doi-org.brum.beds.ac.uk/10.3390/d13100480
Submission received: 25 August 2021
/
Revised: 21 September 2021
/
Accepted: 23 September 2021
/
Published: 29 September 2021
(This article belongs to the Special Issue 2021 Feature Papers by Diversity’s Editorial Board Members)
Abstract
:The flora of the Canary Islands has been subject to botanical studies for more than 200 years. Several biodiversity databases are available for the archipelago. However, there are various drivers of change in real biodiversity and the knowledge about it constantly needs to be kept track of. Island floras are both: exposed to species loss and to species introductions, either through natural processes or by anthropogenic drivers. Additionally, the evolution of endemic plant species plays a substantial role. Endemic species are sensitive to population decline due to small population sizes and possible low competitiveness against incoming species. Additionally, there is continuous progress in systematics and taxonomy. Species names or their taxonomic attribution can be modified. Here, we check published plant lists for the Canary Islands and literature, and compile currently accepted taxa into an updated checklist. For this FloCan checklist, several sources were compiled, checked for completeness and quality, and their taxonomy was updated. We illustrate how far plant names are considered in regional or global databases. This work represents the current state of knowledge on Canary Island plant diversity, including introduced and recently described taxa. We provide a comprehensive and updated basis for biogeographical and macroecological studies. Particularly, the number of non-native species is being extended substantially. The adaptation to standard international nomenclature supports integration into large-scale studies.
1. Introduction
The Flora of the Canary Islands archipelago has attracted botanists for centuries. Alexander von Humboldt, for example, spent a week on Tenerife in the year 1799. During this short stay, he described the elevational distribution of plant species at the slope of Mount Teide. The iconic figure displaying the altitudinal distribution key species was published after his return to Europe in 1826 [1]. In the same year as Humboldt’s ascent to Mont Teide, in 1799, the first natural history book on the Canary Islands comprising species lists, and written by Viera y Clavijo, was released [2].
Knowing about Humboldt’s experiences, Charles Darwin was keen to see the vegetation of Tenerife in January 1832 when the Beagle arrived offshore. However, nobody was allowed to leave the Beagle at the harbour of Santa Cruz de Tenerife due to quarantine restrictions because of the Cholera epidemic in London at that time. Darwin depicts the situation in his report published in 1839 [3] “Oh misery, misery—we were just preparing to drop our anchor within 1/2 a mile of Santa Cruz when a boat came alongside bringing with it our death-warrant. The consul declared we must perform a rigorous quarantine of twelve days. Those who have never experienced it can scarcely conceive what a gloom it cast on every one: Matters were soon decided by the Captain ordering all sail to be set & make a course for the Cape Verd Islands. We have left perhaps one of the most interesting places in the world, just at the moment when we were near enough for every object to create, without satisfying, our utmost curiosity”. Captain Fitzroy, manoeuvring the Beagle, wrote about this situation “this was a great disappointment to Mr Darwin, who had cherished a hope of visiting the peak. To see it, to anchor and be on the point of landing, yet be obliged to turn away without the slightest prospect of beholding Tenerife again, was indeed to him a real calamity”. Having a strong background in botany, Darwin might have had an eye-opening experience if this incidence had not prevented him from exploring the flora of the Canary Islands.
The middle of the 19th century saw a strong stimulation of botanical assessments with the outstanding “Histoire naturelle des Iles Canaries” written by Philip Barker Webb and Sabin Berthelot (and Afred Moquin-Tandon) [4]. This book became the most important landmark for a complete flora of the archipelago. A few decades later, Hermann Christ [5,6] published another list of plant species in the archipelago. During the 20th century, an extensive series of floristic studies and species lists were edited with the pioneering work of Charles-Josef Pitard [7]. Increasingly new contributions by botanists such as Bornmüller, Buch, Sventenius, Broussonet, Masferrer, Burchard, Hansen, Kunkel, Sunding, Bramwell, Santos Guerra, and Schönfelder, to name but a few, increased the knowledge about the Canarian flora [8,9,10,11,12,13]. Additionally, nowadays the archipelago continues to attract the attention of international naturalists and scientists. This long legacy of botanical research evokes the impression that the plant species of the islands are well known, and it may be one of the best investigated regions of the planet. However, even though the Canary Islands were colonised and settled by Europeans centuries ago and have become an attractive destination for tourists, no complete survey of the entire islands could ever be conducted. The steep terrain of remote mountain slopes, inaccessible gorges (barrancos), and rugged cliffs at their coastline are restricting human investigations and hamper accessibility [14,15]. Substantial parts of the archipelago cannot be reached and are not even accessible to climbers due to the loose parent material of young volcanic rock.
In recent years, several new plant species have been described in the archipelago [16,17,18,19,20,21]. However, the publication of a new species does not translate directly into being incorporated and accepted in standard international databases. This process takes time. Additionally, expert knowledge exists about species that exist in nature but has not yet been addressed in official and accepted scientific publications. Several species are known and mentioned by experts but not officially described yet. This even applies to woody species of considerable size and with clear morphological distinction (Figure 1).
Furthermore, plant species recorded in the past have been reported to have disappeared (e.g., Hypecoum procumbens L. or Grammitis quaerenda Bolle), to have extinguished populations, or to have become extinct [22]. Some endemic plant species are currently at the brink of extinction (e.g., Lotus eremiticus A. Santos). This is a common syndrome on islands, where species populations can be small and viable population sizes can easily be undershot. Comparable examples exist in the Galapagos islands for previously important key species from the endemic genus Scalesia [23] or on Mauritius, exhibiting the prominent example of Sideroxylon grandiflorum [24]. Others are likely to have disappeared in the past and might only be reflected as genera in rare studies of pollen records [25]. Many islands have an extensive legacy of biodiversity loss. However, there is little quantitative evidence for this due to the limited preservation of plant remains.
Because of the continuous progress in plant systematics and taxonomy, there is a need to update and unify nomenclature, particularly for those genera or families under debate and for those experiencing considerable upheaval. Understandably, it is mostly those groups that have undergone rapid radiation and diversification in the archipelago, respectively, where the identification of species and their relatedness is work in progress [26,27,28].
During recent decades, several floras and species lists of the Canary Islands have been published and updated as online databases reflecting the state of knowledge and its rapid development. Here, we can just mention prominent examples of this vast literature [22,29,30,31,32,33,34,35]. It is difficult to decide on one work as a standard. Some are mainly rooted in regional and local knowledge; others are better related to international taxonomic standards; some are more recently published; others follow a more rigorous understanding of systematics; some are continuously updated online databases; others are printed books. When looking closer at some taxa, there is disagreement in many details, including the acceptance of species and differences in the perceptions of their systematic and taxonomic status. However, there is a general need to unify and update taxonomic and spatial information on species [36] to reflect real biodiversity at its best and to enable, e.g., inter-island comparison.
Regional flora and checklists of the Canary Islands are increasingly used in macro-ecological and biogeographical studies [37,38,39], which, however, may reflect just a subsection of the real species diversity of the archipelago [40,41]. The resulting findings may be questionable if a substantial part of the existing species is being ignored [42]. Furthermore, outdated taxonomy might inhibit or weaken studies at larger scales.
Openly available public webpages on endemic plants of the Canary Islands, for instance in Wikipedia, differ strongly in content between languages. The Spanish site lists 122 endemic plant species [43]; the English version linked to the same page informs about only 68 endemic plant species [44]! This illustrates that there is not a clear common ground on this topic. A reason for these differences might be the definition of endemism. It is by its nature scale-dependent [45,46] and may be subjectively defined if the area it is related to is not clearly limited [47]. Additionally, there is human bias to be considered meaning that people might be used to certain species names or taxa that have a high value in nature conservation, and that persist even if there is scientific evidence that such names can no longer be accepted and must be updated.
The advancement of knowledge and confusion due to new findings for the Canary Island flora is understandable at best with the example of the dragon tree (Dracaena draco (L.) L.). The dragon tree is maybe the most iconic plant species in the archipelago. Alexander von Humboldt was already fascinated by the impressive life form of several specimen in the valley of Orotava, Tenerife, back in his days. Then, 200 years later in 1997, a small population of a subspecies of Dracaena draco was found in the Moroccan High Atlas Mountains (subsp. ajgal Benabid & Cuzin). In consequence, Dracaena draco was no longer a Macaronesian endemic species (to the Canary Islands and Cabo Verde) but an endemic subspecies (D. draco subsp. draco) for these islands. One year later, in 1998, another Dracaena species Dracaena tamaranae A. Marrero, R.S. Almeida & M. Gonzalez-Martin was found on the island of Gran Canaria, again adding an endemic Dracaena species to the archipelago [48]. Such surprises and discoveries would rather be expected for less famous plants or for cases where the deterministic traits are not so obvious but are astonishing for the most well-known plant taxa of the islands.
Another prominent example is the Canary Island laurel tree, a key species of the laurel forest, which is best preserved in this archipelago. Laurus novocanariensis Rivas Mart., Lousã, Fern. Prieto, E. Días, J.C. Costa & C. Aguiar was named Laurus azorica (Seub.) Franco before and even further back in time was named Laurus canariensis Webb & Berthel. non Willd. To add to the confusion, Laurus canariensis Willd. is a published but invalid synonym for another Lauracean species of the island: Apollonias barbujana (Cav.) Bornm. In summary, Laurus novocanariensis, as it is currently named, was an archipelago endemic species first, became a Macaronesian endemic, and back again an archipelago endemic taxon. The name has changed due to an increasing understanding of phylogenetic relatedness. However, it may happen that in the future this taxon could be understood as a synonym to Laurus nobilis L. from the Mediterranean which would mean that one more classified endemic species would be lost from the list of plant species just in the human understanding of biological taxonomy—without any consequences for nature [49].
Changes in the number of species that are described for islands are mostly due to newly introduced and even invasive species. These species are being introduced by past and present trade and travelling activities that connect the islands to other parts of the world [50]. This applies strongly to the Canary Islands that have been used as a testing ground for the acclimatisation of exotic species from the New World that were intended to be introduced to the Mediterranean as early as in the 16th century [51]. The subtropical and oceanic climate supports the establishment of many species across tropical and subtropical biogeographic realms. The differentiation of climatic conditions within the archipelago and even within topographically diverse islands with pronounced elevational zones and differences in precipitation and moisture regimes between leeward and windward sides adds to the spectrum of available habitats for the establishment of non-native biota [52]. Many plants have been introduced for ornamental purposes. Those species may remain confined to gardens or parks, but they may also start reproducing and spreading after a certain time lag and establish a legacy of offspring. However, the same mechanism applies for newly introduced species as for unknown endemics; they must first be detected before they can be registered in any database or list. Too often knowledge about cultivated ornamental plants that were established outside their natural range has been ignored until such species turned “wild” and created problems and damage.
Another process that is modifying the diversity of islands is extinction. Many of these extinction events may have occurred unnoticed since invasive species, unknown herbivores such as rabbits or goats, or other predators and pathogens have been introduced. However, the pressure of introduced herbivores on island species that have not evolved defence mechanisms is still pertinent [53]. Many endemic plant species have dramatically declined in distribution and abundance with the consequence of becoming hyper-endemic [46], surviving as remnant populations at the brink of extinction. In the case of Lotus eremiticus A. Santos only one “population” of few specimens remains in nature, most probably built up by a single clone [54]. One single (local) disturbance event could erase such a species globally. Considering the vulnerability of such hyper-endemics, it becomes evident that the human impact has clearly reduced intraspecific diversity. Generally, it is not a given fact that island taxa exhibit low genetic diversity as concluded from their small populations [55,56].
The genus Aeonium is iconic to the archipelago and is also considered as an example of adaptive radiation. Of the 42 species of the genus, 36 are endemic to the Canary Islands. Surprisingly, the phylogenetically oldest species do not originate from the continent (Morocco, Eastern Africa, Yemen), but the continental species instead represent the young branches of the phylogenetic tree [57]. The earth history of “Paleo-Macaronesia” is one explanation for this pattern [58].
Many more volcanic islands more existed in this tectonically complex area at the edge of the oceanic and the continental crust during the last 80 Mio. years. These former islands have disappeared from the ocean surface due to erosion. However, they can be detected with the help of bathymetry as guyots at the sea level of the Last Glacial Maximum 200 (LGM), which was their basis for erosion [59]”. Furthermore, the spatial fluctuation of island area during the Pleistocene had an influence on plant species richness [60].
Increasingly, modern molecular methods allow for new insights into the distinction of species and to their attribution to higher phylogenetic units, influencing nomenclature, systematics, and biogeography [61,62,63,64]. Realistically, this process is a continuous one that will not end soon because of the immense diversity of plant populations and the historical focus on mere morphological traits for classification. Hidden relations between taxa need to be uncovered, and cryptic species that resemble other published species need to be identified. Consequently, data sources for regional and trans-regional assessments need to be updated continuously and adapted to international standards in nomenclature to avoid artefacts just through deviating terminologies.
The Canary Islands are an outstanding example of an essential field for ecological research, the preservation of endemic taxa, and for biogeographical studies. Here, we offer an overview of the current knowledge and perception of the flora of the Canary Islands, knowing that near future developments in phylogeny and discoveries of new species might modify this picture. Nevertheless, we identify that the customary reference to one specific list of plant species has an influence on the scientific statements made. Based on the comparison of established approaches, databases, and lists of the Canary Islands, we identified common general taxonomic agreement but also inconsistencies. Based on this overview, we present an updated and revised flora for the archipelago that is open for additions and corrections.
2. Materials and Methods
We first reviewed and compiled published species lists for the Canary Islands [8,9,10,11,12,13,22,29,30,31,32,33,34,35]. The resulting plant species list was then complemented with individual publications on specific taxonomic groups for specific regions or islands. In addition, documented cultivated ornamental plants and crops were included. This study is focused on the 7 major islands of the Canary Islands archipelago: El Hierro, La Palma, La Gomera, Tenerife, Gran Canaria, Fuerteventura, and Lanzarote. With very few exceptions, all taxa are attributed to their occurrence on these islands.
Small islets in the close vicinity of Lanzarote and Fuerteventura were excluded as they have been covered in a recent study that provided a checklist for these islets [65]. However, there were no additional species listed in comparison with our checklist.
Criteria for accepting a taxon in this new FloCan plant checklist for the Canary Islands were the reliability of records and whether the respective species or subspecies name is accepted or considered a synonym following international standards of nomenclature and systematics. In the case of questionable records, additional proof was explicitly searched, and if a record for a given species on an individual island or for the entire archipelago was not found, this species was deleted from the list. The same applied if a plant population was considered an independent subspecies (or species), but this taxonomic categorization did not align with international standards. However, such synonyms or erroneously reported species are also maintained in an appendix to allow checking for the existence of these taxa in the future. For highly debated taxa, we also consider current scientific literature on plant phylogenetics (e.g., [66] for the genus Micromeria).
The resulting list was then compared to international standard taxonomic checklists prioritising those species that are listed in Plants of the World online (POWO) [67]. As a result, published species names in one list or dataset can translate into subspecies and vice versa.
Additionally, we consulted World Flora Online (WFO) [68] for all taxa, which is the replacement of The Plant List (TPL) [69] that has not been updated since 2013. Therefore, TPL was not considered explicitly in this checklist to avoid redundance and outdated viewpoints. We further compared all taxa with Catalogue of Life [70], which generally accepts more taxa as valid names than World Flora Online. We do not suggest that one of these databases is superior to another one. However, for standardisation we decided to follow the suggested plant names of one checklist, generally Plants of the World Online, indicating nevertheless whether other databases share the same name for a respective plant species, or suggest a synonym as the accepted name.
Additionally, we checked names in GBIF [71], which is not a taxonomic database, but is frequently used and to be considered a standard to obtain species’ distribution data. Furthermore, we screened the TRY [72,73] database for plant functional traits. These international databases differ in terms of scientific scrutiny and their general philosophy. For example, TRY does not provide author citation, which is a substantial part of botanical names.
In this study, databases are not evaluated. Rather it shall be illustrated which taxa are considered under the name given in this checklist (or under a synonym) in these pre-valent databases. We follow, with only very few exceptions, in the naming for plant species, genus, family, and subspecies the standards of POWO. Other infraspecific units (e.g., varieties) are only considered in a few cases. There is no clear agreement across checklists to which degree infraspecific taxa should be considered below the level of subspecies.
Deviating nomenclature or missing representation in other plant lists is protocolled. This option in our checklist illustrates which taxa are generally accepted and where no consistent opinion across published plant species lists can be seen. There are cases where the same species’ name was published by several authors. Confusion could result from missing author citations if one of these apparently equitable names is defined as a synonym of another accepted species.
For list comparison we used a semi-automatised approach and compared our species list with global databases via the “taxize” package [74] applied in R [75]. Every species for which several or no results were given were rechecked manually. However, this filter was complemented by individual reviews and revisions for all taxa, to overcome, for example, deviating spelling of names. Botanical publications that are based on taxa should follow the International Code of Nomenclature for algae, fungi, and plants (ICN) as a global compilation of published plant names in scholarly publications serves the International Plant Names Index (IPNI) [76]. However, IPNI is not aiming to provide the latest state of knowledge for the progress in taxonomy and its reflection in botanical nomenclature.
Island biota and endemic species, but also recently published data on taxa with revised nomenclature, are likely to be underrepresented in global databases. This needs to be considered to avoid data bias in trans-regional biodiversity studies. In our checklist, the current state of representation of Canary Island plants in these databases is protocolled, even though such data repositories will further develop towards higher completeness and representation. This comparison serves as orientation on the reliability of research that uses such open data sources without scrutinising and comparing every single species.
In addition to published floras and plant lists, we reviewed the current botanical literature, focusing on studies about recently documented established non-native species and taxonomic studies for selected species groups [77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108]. These studies are sources for additional species records and changes in plant names, which are not yet included in global or regional databases.
To indicate taxa that are highly debated compared to others, we checked the number of published synonyms in POWO. Infraspecific taxa such as subspecies and varieties are also listed if they are officially published. We separate our analyses for different taxonomic levels. In some genera (e.g., Aeonium, Micromeria) a series of hybrids are described. Hybrids are listed in a separate table in order not to ignore this aspect of biodiversity, but also to show that such organisms, that might appear with a certain regularity, have not evolved (yet) to accepted species.
Finally, we suggest an updated plant list including information on the spatial distribution of taxa across the Canary Islands and their status. We classify native species as probably native and surely native. Non-native species were categorised as probably non-native in cases where this is not certain, surely non-native and invasive non-native (i.e., intruding into and substantially modifying natural ecosystems). The term “non-native” is being used as a synonym of the term “alien”. Our study aims to improve the picture of the currently existing taxa on the island, but also to acknowledge that numerous taxa are unanimous or under debate.
3. Results
This new FloCan checklist informs about the current state of knowledge of the flora of the Canary Islands considering regional floras and international taxonomic databases as well as specific publications (Supplement Materials Part S1–S6). It aims at providing a transparent overview of the acceptance of species and infraspecific names, suggesting a revised checklist. The total number of accepted taxa in this checklist sums up to 2812 (1781 native, 1031 non-native), comprising 2416 species (1452 native, 964 non-native) and 396 infraspecific taxa (329 native, 67 non-native) (Supplement Materials Part S1, Main Table for species and Infraspecific taxa)”.
There are substantial differences between islands in the number and proportion of native and non-native taxa, species, and infraspecific taxa (A species list can be found in Supplement Materials Part S2). The proportion of native taxa is high for the arid islands with less topographic diversity in the eastern part of the archipelago (Fuerteventura and Lanzarote) (Figure 2). Islands with a large human population such as Tenerife or Gran Canaria posess a large number of non-native species. Generally, there are not many non-native infraspecific taxa, which reflects that such biota are an indicator of ongoing speciation.
Within the native taxa (Figure 3), we find a consistent proportion of endemic taxa between approx. 20 and 40 per cent. The proportion of single-island endemic species (SIE) is only higher in comparison with multi-island (archipelago) endemics (MIE) for the sum of all taxa and species across all islands. On individual islands, the proportion of MIE is always larger than that of SIE. The arid islands with less pronounced relief again show comparable patterns, dominated by native species that are non-endemic. Surprisingly, the proportion (and absolute number) of SIE is very small also for El Hierro, whereas the other mountainous islands exhibit a consistently high proportion of SIE. Infraspecific taxa exhibit higher proportions of SIE and MIE compared to species or taxa level consistently for all individual islands but show the opposite pattern for the entire archipelago.
As this FloCan checklist provides an updated list of plant taxa, including their representation in other existing floras and databases, we want to illustrate to what degree other lists are deviating from the here suggested taxonomy (Figure 4). Uncertainty is generally high for infraspecific taxa. The reflection of accepted taxa is good in GBIF. However, many accepted taxa, mainly endemic and infraspecific taxa are not well represented in the TRY database, which can cause bias in ecological studies.
Generally, the compiled list of taxa is well reflected in two currently applied reference lists for the flora of the Canary Islands [22,34] (Figure 5). However, a substantial number of taxa appear in these lists as not-accepted synonyms, and also in international reference databases (Figure 4).
In addition to the main checklist, we add a list of currently not considered taxa that were published for the Canary Islands before, illustrating the reason for exclusion such as being a synonym of another accepted taxon or an obviously erroneous record or misunderstanding (Supplement Materials Part S5). As it does not make sense to list all synonyms ever published, which can sum up to more than 100 for a single species, we report those synonyms in detail in Supplement Materials Part S6 that are used in the above-mentioned datasets and floras referred to in this checklist to guide readers towards accepted names in this checklist.
Published hybrids are listed in addition, in a separate list (Supplement Materials Part S3), as this field is highly likely to be incomplete, and less clearly regulated. Particularly in specific genera (e.g., Micromeria) several hybrids are described. As hybrid speciation can be an important process, hybrids contribute to the biodiversity of the Canary Islands.
Furthermore, we add a list of not (yet) naturalized plants planted in gardens and parks (Supplement Materials Part S4), focusing on recorded perennial, long-lived plants because those species might naturalise and establish in the future. This list contains non-native exotic species that have not been found and reported yet to produce natural offspring. The species of this list are not included in the here presented analyses as we could not compile an exhaustive list of these species given the information available.
In Table 1, we provide a comparison for the taxa accepted in the FloCan Checklist (Supplement Materials Part S1, Main Table) with other modern plant lists such as Atlantis Biota or Muert et al. Major differences relate to the consideration of recently recorded non-native species, but also to taxonomic revisions.
Differences in the number of families are related to deviating reflection of changes in plant taxonomy and systematics. The splitting up of the family Scrophulariaceae is one of several examples of fundamental changes in plant families. Additionally, the increasing consideration and recording of exotic species contributes to an increase in plant families, respectively.
Progress in the exploration of species and in systematics reveals that a flora (plant list) is a moving target due to processes such as the increasing identification of endemism (Table 2) through records of new species or molecular identification of their taxonomic separation. Another process contributing to the ongoing changes in regional biodiversity is the establishment of non-native species (Table 3). Both processes are considered in the updated FloCan checklist.
4. Discussion
This new checklist reflects the current state of knowledge on the Flora of the Canary Islands, one of the most important evolutionary arenas in the holarctic realm [109]. This knowledge is under constant change and development. Therefore, it seems impossible to provide one final product, even in the case of the Canary Islands archipelago, which has been subject to botanical studies since the beginning of the 19th century. New species are still being found that were not known to science before, while small endemic populations of species are at the brink of extinction. Additionally, new non-native species are introduced as crops, ornamental plants or accidentally, and then may become established in the natural or semi-natural environment close to settlements or along roadsides. Some of these species become invasive, intruding into natural communities, and modifying ecosystems and their functioning, partly resulting in a deterioration of ecosystem services. Such ongoing and even accelerated changes resulting from the mostly undersaturated floras of islands combined with increasing connectivity due to transport and traffic make island biota a moving target.
For only very few taxa, no occurrence records could be attributed to specific islands, although these taxa occur or have occurred in the archipelago. Such species are listed in some sources as occurring in the archipelago, but without a precise location on specific islands or without confirmation during the last decades. One endemic plant species (Solanum nava Webb. & Berthel.) is likely completely extinct now, although this was already thought to be the case in the 1970s. One species has most likely become regionally extinct (Grammitis quaerenda Bolle), and three others have not been recorded for years (Glinus lotoides L., Hypecoum procumbens L., Picris hieracioides L.). Nevertheless, such taxa were not excluded as there is a chance of rediscovery. Other species with no clear local records in this list are part of complex groups that require specialist knowledge (e.g., Taraxacum campylodes G.E. Haglund). Currently missing clear local records also apply to some ornamental plants such as Acanthus spinosus L., Amphilophium crucigerum (L.) L.G. Lohmann, or Syagrus weddelliana (H.Wendl.) Becc. and to some tree species that have been planted in forestry (e.g., Pinus sylvestris L.).
In Supplement Materials Part S4 we provide an additional list of plant species recorded in gardens and parks. This list aims to create awareness about possible additional invasion processes even if the specimens do not yet show natural regeneration and dispersal. We encourage, however, the monitoring of these species, as some of these have been recorded to become invasive in other places of the world (e.g., Artemisia absinthium L.). Some herbaceous species on this list, such as Sanguisorba minor L. could naturalise rapidly without necessarily becoming invasive. As several of these garden and park species are trees and shrubs, they might invade and modify natural ecosystems. Nevertheless, such a list can only be incomplete as there is a constant import of ornamental plants. For the reasons of non-proven natural regeneration and incompleteness, this list was separated from the general checklist. Consequently, the main FloCan checklist does not comprise these ornamental garden plants. Here, we exclude them from the analyses, even if they can become non-native members of the natural vegetation quite rapidly.
Progress in taxonomy and systematics results in modified attribution of organisms to species, subspecies and even genera and families. This may be uncomfortable for practitioners that are used to specific terms and may even affect legal regulations with fixed terminology and nomenclature, but it is an intrinsic and essential condition of botanical sciences. Because of this continuously ongoing struggle to improve the understanding of nature, there is no complete agreement on all facets of taxonomy within the scientific community. Even if there are clear rules and regulations for accepting a scientific name, it may take time until such insights are generally accepted and translated into floristic inventories. However, deviating viewpoints will always exist.
Global databases are “work in progress” constantly being subject to change. Moreover, they can be incomplete, particularly for island biota. This applies mainly to the TRY database [72], which includes only very few infraspecific taxa, does not inform about species names’ authors (which can result in errors due to synonyms), and misses many endemic species. However, the frequent use of such sources in large-scale studies implies the necessity to clarify to which degree island biota are covered and which restrictions need to be considered.
A global invader can serve as an example for a possible confusion of plant names. Pennisetum setaceum (Forssk.) Chiov. appears in GBIF [71] as Pennisetum setaceum (Forssk.) Chiov. as well as Cenchrus setaceus (Forssk.) Morrone, with deviating occurrence records for the Canary Islands. A total of 8970 records are provided for C. setaceus for all Canary Islands but only 4473 records are given for P. setaceum, including no reference for the island La Gomera (status 28 June 2021). This illustrates that a critical screening of available data is recommended because this is just one species, and a fully automated data mining can hardly recognise such errors that can even occur for widespread species when the nomenclature has changed or synonyms have been published. This species is listed in our checklist as Cenchrus setaceus (Forssk.) Morrone. This is also the name of the species in the Atlantis Biota data base of the Cabildo Insular of the Canary Islands [22]. However, it appears under a synonym in Muer et al. [34]. Our reference database for international standards, Plants of the World Online [67], also accepts this name, as well as Catalogue of Life [70], whereas it is seen as ambiguous in World Flora Online [68]. Comparable thorough screening across databases was done for all taxa in our checklist.
If endemic plants on oceanic islands are seen as a reflection of ongoing evolutionary processes, infraspecific variation cannot be ignored. Subspecies or varieties may not be very precisely defined and can be seen as beyond the biological species concept. However, speciation has many facets, including hybrid speciation and apomixis. Hence, biogeographical assessments which are aiming to characterise the moving target of evolution cannot ignore such infraspecific units. Again, it was Charles Darwin who was very aware of this fact. In August 1857 he wrote to J.D. Hooker: “I am got extremely interested in tabulating according to mere size of genera, the species having any varieties marked by greek letters or otherwise: the result as far as I have yet gone seems to me one of the most important arguments I have yet met with, that varieties are only small species—or species only strongly marked varieties. The subject is in many ways so very important for me; I wish much you would think of any well-worked Floras with from 1000–2000 species, with the varieties marked. It is good to have hair-splitters & lumpers” [110]. Hopefully, Charles Darwin would have been happy with the plant list provided herewith.
Today, big data algorithms are applied in biogeographic research relying on the correctness of species names, records and occurrence. (e.g., [38,39,40,41,42,60,111,112]). Such analyses are dependent on the expert knowledge fed into databases and their maintenance and quality control. However, there are many sources of error in databases because real-time control cannot be implemented and autotomized, or AI (artificial intelligence) algorithms are not (yet) able to replace expert knowledge, including specific challenges for certain taxa that exhibit, for instance, apomictic processes for reproduction. Additionally, the progress in phylogenetic research is not equal across taxonomic groups, with some being more thoroughly scrutinised because there is a larger scientific community working with these. In contrast, others are more neglected with the consequence of a possibly outdated nomenclature.
The Canary Islands are of outstanding importance to biodiversity covered by the European Union and should be considered strongly in the implementation of the EU Biodiversity Strategy for 2030 [113]. Ongoing land use changes, pressures related to tourism, climate change and additionally, the negative impact of introduced alien herbivores [114] are calling for a reinforced commitment in nature conservation [115]. The designation of protected areas is a common tool in conservation, requiring a sound knowledge of the uniqueness of biota, including infraspecific taxa. Databases and checklists are an important basis for such strategies [116]. The Canary Island protected area network is a work in progress. Besides the management of protected areas, the entire archipelago should be seen as a cradle of nature [109]. To achieve the objective to preserve its biodiversity of the future, laws and regulations need to be efficiently implemented, but in addition, enhanced support in human resources and financial endowment is required.
This FloCan checklist is the result of an in-depth survey on regional literature and web-based platforms, including own experience, data recording in the field and screening of international standard databases. It reflects the current understanding of taxa and the recent information about species records. Each flora, however, is a work in progress or can even be seen as a “moving target”. Additional non-native species can be expected to establish, and even endemic species not yet known to science can be discovered in remote places. Additionally, cryptic taxa can be hidden under the disguise of morphological similarity and will be detectable with molecular methods. Here, we update the state of knowledge in the present moment. Nevertheless, our transparent approach to illustrate converging, but also deviating perceptions and points of view in common plant lists enables more critical and realistic biogeographical assessments.
5. Conclusions
The Canary Island archipelago is a hotspot of plant endemism and a safe site for remnant populations of plant taxa that have become extinct on neighbouring continents during Pleistocene and even Holocene climatic fluctuations. The oceanic climate, combined with pronounced topography, offers suitable habitats for a wide range of species and plant functional groups. However, the total species richness is, like on all islands, relatively low due to dispersal filters. Non-native species have become abundant since humans contributed to their transport, establishment, and provision of disturbed and anthropogenic habitats. Processes that are contributing to phytodiversity, such as invasion, extinction, or evolution, are progressing with varying momentum and different speeds. Furthermore, they do not proceed equally on all islands or in all ecosystems. The global importance of the Canary Islands requires a continuous survey and monitoring of biodiversity. The FloCan checklist aims to reflect the state of knowledge in July 2021 and is very likely to be amended and adapted in the future. Progress in phylogenetics may modify the status of well-known taxa. Still, new species are being discovered, and more and more non-native species are likely to become establish and detected. Additionally, ornamental plants may start regenerating after a lag period or develop possible invasive population dynamics under climate change. Therefore, this study explicitly includes many non-native plants that were missing in previous lists. Being adapted to the current international taxonomy standards, this list can be used for trans-regional or even global biogeographical studies.
Supplementary Materials
The following material is available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/d13100480/s1, Part S1: FloCan Main List (Accepted plant species and infraspecific taxa); Part S2: FloCan Plant Species List; Part S3: Published hybrids (not included in biogeographical analyses); Part S4: Recorded non-naturalized garden plants and plants in public places such as parks (not included in biogeographical analyses); Part S5: List of previously reported plants that were excluded and explanation for exclusion; Part S6: Synonyms used in currently published plant lists (but not accepted in FloCan).
Author Contributions
Conceptualization, C.B.; methodology, C.B. and A.W.; validation, C.B. and W.W.; formal analysis, A.W.; investigation, C.B. and A.W.; resources, C.B. and W.W.; data curation, C.B., A.W. and W.W.; writing—original draft preparation, C.B.; writing—review and editing, A.W. and W.W.; visualization, A.W.; supervision, C.B.; project administration, C.B.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data provided by this study are documented in the Supplementary Materials.
Acknowledgments
This work was greatly supported by Kira Meyer in data compilation, and by Reinhold Stahlmann for all technical aspects of data handling. We are grateful to Felix Medina, from the Cabildo Insular on the Island of La Palma, Spain, for his support in field research. Thanks to Alessandro Chiarucci, Bologna, Italy, Ole Vetaas, Bergen, Norway, Severin Irl, Frankfurt, Germany, Carsten Hobohm, Flensburg, Germany, and Richard Field, Nottingham, UK, for their company in the field and stimulating discussions that generated the need for improved botanical data availability.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Humboldt, A.V.; Bonpland, A. Viage á Las Regiones Equinocciales del Nuevo Continente; Casa de Rosa: Paris, France, 1826. [Google Scholar]
- Viera y Clavijo, D.J. Diccionario de Historia Natural de las Islas Canarias, o Índice Alfabético Descriptivo de sus Tres Reinos, Animal, Vegetal y Mineral; Excma. Mancomunidad de Cabildos de Las Palmas, Plan Cultural: Las Palmas de Gran Canaria, Spain, 1799. [Google Scholar]
- Darwin, C. Voyage of the Beagle. In The Narrative of the Voyages of H.M. Ships Adventure and Beagle; Henry Colburn: London, UK, 1839; Volume 3. [Google Scholar]
- Webb, P.B.; Berthelot, S. Histoire naturelle des Iles Canaries. In Phytogeographia Canariensis; Béthune: Paris, France, 1844; 1430p. [Google Scholar]
- Christ, H. Vegetation und Flora der Canarischen Inseln. Engler’s Bot. Jahrbücher 1885, 6, 458–526. [Google Scholar]
- Christ, H. Specilegium canariense. Engler’s Bot. Jahrbücher 1887, 9, 86–172. [Google Scholar]
- Pitard, J.; Proust, L. Les Iles Canaries. In Flore de l’Archipel; Béthune: Paris, France, 1908; 503p. [Google Scholar]
- Eriksson, O.; Hansen, A.; Sunding, P. Flora of Macaronesia: Checklist of Vascular Plants; Sommerfeltia: Umea, Sweden, 1974. [Google Scholar]
- Kunkel, G. Flora y Vegetación del Archipiélago Canario. Tratado Florístico. 2ª Parte. Dicotiledóneas; Ed. Edirca: Las Palmas de Gran Canaria, Spain, 1991; 312p. [Google Scholar]
- Kunkel, G. Die Kanarischen Inseln und ihre Pflanzenwelt; Gustav Fischer Verlag: Stuttgart, Jena, Germany; New York, NY, USA, 1993. [Google Scholar]
- Bramwell, D. Flora de las Islas Canarias; Rueda: Madid, Spain, 1997. [Google Scholar]
- Schönfelder, P.; Schönfelder, I. Die Kosmos-Kanarenflora; Kosmos: Stuttgart, Germany, 2012; 319p. [Google Scholar]
- Santos Guerra, A. Vegetacion y Flora de La Palma; Editorial Interinsular Canaria: Santa Cruz de Tenerife, Spain, 1983; 348p. [Google Scholar]
- Fernandez-Palacios, J.M.; Martin Esquivel, J.L. Naturaleza de las Islas Canarias; Editorial Interinsular Canaria: Santa Cruz de Tenerife, Spain, 2002. [Google Scholar]
- Pott, R.; Hüppe, J.; Wildpret de la Torre, W. Die Kanarischen Inseln—Natur- und Kulturlandschaften; Ulmer: Stuttgart, Germany, 2003. [Google Scholar]
- Montelongo, V.; Bramwell, D.; Fernández-Palacios, O. Parolinia glabriuscula (Brassicaceae), una nueva especie para Gran Canaria (Islas Canarias, España). Bot. Macaronésica 2003, 24, 67–72. [Google Scholar]
- Gil Gonzalez, J.; Gil Gonzales, M.L.; Morales Mateos, J.B.; Mesa Coello, R. Vicia vulcanorum (Fabaceae) a new species from the island of Lanzarote (Canary Islands). Collect. Bot. 2012, 31, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Bañares Baudet, Á.; Acevedo Rodríguez, A.; Rebolé Beaumont, Á. Monanthes subrosulata, a new species of M. sect. Sedoidea (Crassulaceae) from La Palma, Canary Islands, Spain. Willdenowia 2013, 43, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Crawford, D.J.; Mort, M.E.; Archibald, J.K. Tolpis santosii (Asteraceae, Chichorieae), a new species from La Palma, Canary Islands. Vieraea 2013, 41, 169–175. [Google Scholar]
- Gil Gonzalez, J.; Morales Mateos, J.B.; Gil Gonzales, M.L.; Mesa Coello, R. Vicia voggenreiteriana (Fabaceae) a new species from the island of La Gomera (Canary Islands). Viaeraea 2013, 41, 189–201. [Google Scholar]
- Friesen, N.; Herden, T.; Schönfelder, P. Allium canariense (Amaryllidaceae), a species endemic of the Canary Islands. Phytotaxa 2015, 221, 1–20. [Google Scholar] [CrossRef]
- Atlantis Biota Database. Available online: https://www.biodiversidadcanarias.es/biota/ (accessed on 5 July 2021).
- Mauchamp, A.; Atkinson, R. Rapid, Recent and Irreversible Habitat Loss: Scalesia Forest in the Galapagos Islands; Galapagos Report 2009–2010; GNPD; GCREG; CDF; GC: Galápagos, Ecuador, 2010; pp. 108–112. [Google Scholar]
- Wittmer, M.C. The dodo and the tambalacoque tree: An obligate mutualism reconsidered. Oikos 1991, 61, 133–137. [Google Scholar] [CrossRef] [Green Version]
- de Nascimento, L.; Willis, K.J.; Fernandez-Palacios, J.M.; Criado, C.; Whittaker, R.J. The long-term ecology of the lost forests of La Laguna, Tenerife (Canary Islands). J. Biogeogr. 2009, 36, 499–514. [Google Scholar] [CrossRef]
- Bañares Baudet, Á.; Marrero Gómez, M.V.; Scholz, S. Taxonomic and nomenclatural notes on Crassulaceae of the Canary Islands, Spain. Willdenowia 2008, 38, 475–489. [Google Scholar] [CrossRef] [Green Version]
- Peroni, A.; Peroni, G. Considerazioni tassonomiche sul genere Cystopteris Bernh. (Athyriaceae: Pteridophyta) di El Hierro (Isole Canarie). Ann. Mus. Cov. Rovereto Sez. Arch. St. Sc. Nat. 2006, 21, 207–210. [Google Scholar]
- Vitales, D.; Garnatje, T.; Pellicer, J.; Vallès, J.; Santos-Guerra, A.; Sanmartin, I. The explosive radiation of Cheirolophus (Asteraceae, Cardueae) in Macaronesia. BMC Evol. Biol. 2014, 14, 118. [Google Scholar] [CrossRef] [Green Version]
- Hansen, A.; Sunding, P. Flora of Macaronesia: Checklist of Vascular Plants, 4th ed.; Sommerfeltia: Umea, Sweden, 1993; Volume 17, pp. 1–295. [Google Scholar]
- Hohenester, A.; Welss, W. Exkursionsflora für die Kanarischen Inseln. Mit Ausblicken auf ganz Makaronesien; Eugen Ulmer: Stuttgart, Germany, 1993. [Google Scholar]
- Bramwell, D.; Bramwell, Z. Flores Silvestres de las Islas Canarias, 4th ed.; Rueda Ed.: Madrid, Spain, 2001; 437p. [Google Scholar]
- Izquierdo, I.; Martín, J.L.; Zurita, N.; Arechavaleta, M. (Eds.) Lista de especies silvestres de Canarias (hongos, plantas y animales terrestres). In Consejería de Medio Ambiente y Ordenación Territorial; Gobierno de Canarias: Las Palmas de Gran Canaria, Spain, 2004; 500p. [Google Scholar]
- Arechavaleta, M.; Rodriguez, S.; Zurita, N.; Garcia, A. Lista de Especies Silvestres de Canarias. Hongos, Plantas y Animales Terrestres; Gobierno de Canarias: Las Palmas de Gran Canaria, Spain, 2010; 579p. [Google Scholar]
- Muer, T.; Sauerbier, H.; Cabrera Calixto, F. Die Farn- und Blütenpflanzen der Kanarischen Inseln; Markgraf Publishers: Weikersheim, Germany, 2016. [Google Scholar]
- Flora de Canarias (Gil). Available online: http://www.floradecanarias.com/ (accessed on 5 July 2021).
- Freiberg, M.; Winter, M.; Gentile, A.; Zizka, A.; Muellner-Riehl, A.N.; Weigelt, A.; Wirth, C. LCVP, The Leipzig catalogue of vascular plants, a new taxonomic reference list for all known vascular plants. Sci. Data 2020, 7, 416. [Google Scholar] [CrossRef]
- Kier, G.; Kreft, H.; Lee, T.G.; Jetz, W.; Ibisch, P.L.; Nowicki, C.; Mutke, J.; Barthlott, W. A global assessment of endemism and species richness across island and mainland regions. Proc. Natl. Acad. Sci. USA 2009, 106, 9322–9327. [Google Scholar] [CrossRef] [Green Version]
- Steinbauer, M.; Beierkuhnlein, C. Characteristic pattern of species diversity on the Canary Islands. Erdkunde 2010, 64, 57–71. [Google Scholar] [CrossRef]
- Lenzner, B.; Weigelt, P.; Kreft, H.; Beierkuhnlein, C.; Steinbauer, M. The general dynamic model of island biogeography revisited at the level of major flowering plant families. J. Biogeogr. 2017, 44, 1029–1040. [Google Scholar] [CrossRef]
- Fernández-Palacios, J.M.; Otto, R.; Borregard, M.K.; Kreft, H.; Price, J.P.; Steinbauer, M.J.; Weigelt, P.; Whittaker, R.J. Evolutionary winners are ecological losers among oceanic island plants. J. Biogeogr. 2021, 48, 2186–2198. [Google Scholar] [CrossRef]
- Cutts, V.; Hanz, D.M.; Barjas-Barbosa, M.P.; Algar, A.C.; Steinbauer, M.J.; Irl, S.D.H.; Kreft, H.; Weigelt, P.; Fernandez-Palacios, J.M.; Field, R. Scientific floras can be reliable sources for some trait data in a system with poor coverage in global trait databases. J. Veg. Sci. 2021, 32, e12996. [Google Scholar] [CrossRef]
- Hortal, J.; Lobo, J.M.; Jiménez-Valverde, A. Limitations of Biodiversity Databases: Case Study onSeed-Plant Diversity in Tenerife, Canary Islands. Conserv. Biol. 2007, 21, 853–863. [Google Scholar] [CrossRef] [PubMed]
- Wikipedia Spanish on Categoría: Flora endémica de Canarias. Available online: https://es.wikipedia.org/wiki/Categor%C3%ADa:Flora_end%C3%A9mica_de_Canarias (accessed on 28 June 2021).
- Wikipedia English on Endemic Plants of the Canary Islands. Available online: https://en.wikipedia.org/wiki/Category:Endemic_flora_of_the_Canary_Islands (accessed on 28 June 2021).
- Hobohm, C.; Janišová, M.; Steinbauer, M.; Landi, S.; Field, R.; Vanderplank, S.; Beierkuhnlein, C.; Grytnes, J.A.; Vetaas, O.R.; Fidelis, A.; et al. Global endemics-area relationships of vascular plants. Perspect. Ecol. Conserv. 2019, 17, 41–49. [Google Scholar] [CrossRef]
- Hohbohm, C.; Moro-Richter, M.; Beierkuhnlein, C. Distribution and Habitat Affinity of Endemic and Threatened Species—Global Assessment. In Perspectives for Biodiversity and Ecosystems; Hobohm, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2021. [Google Scholar]
- Thompson, J.D. Plant Evolution in the Mediterranean: Insights for Conservation; Oxford University Press: Oxford, UK, 2020. [Google Scholar]
- Marreiro, A.; Almeida, R.S.; Gonzalez-Martin, M. A new species of wild dragon tree, Dracaena (Dracaenaceae) from Gran Canaria and its taxonomic and biogeographic implications. Bot. J. Linn. Soc. 1998, 128, 291–314. [Google Scholar]
- Rodríguez-Sánchez, F.; Guzmán, B.; Valido, A.; Vargas, P.; Arroyo, J. Late Neogene history of the laurel tree (Laurus, L.; Lauraceae) based on phylogeographical analyses of Mediterranean and Macaronesian populations. J. Biogeogr. 2009, 36, 1270–1281. [Google Scholar] [CrossRef] [Green Version]
- Pyšek, P.; Jarosik, V.; Hulme, P.E.; Hejda, M.; Schaffner, U.; Vila, M.A. Global assessment of invasive plant impacts on resident species, communities and ecosystems: The interaction of impact measures, invading species traits and environment. Glob. Chang. Biol. 2011, 18, 1725–1737. [Google Scholar] [CrossRef]
- Ríos-Mesa, D.; Pereira-Lorenzo, S.; González-Díaz, A.; Hernádez-González, J.; González-Diaz, E.; Saúco, V.G. The status of Chestnut cultivation and utilization in the Canary Islands. Adv. Hortic. Sci. 2011, 25, 90–98. [Google Scholar]
- Irl, S.; Schweiger, A.; Steinbauer, M.; Ah-Peng, C.; Arévalo, J.R.; Beierkuhnlein, C.; Chiarucci, A.; Daehler, C.C.; Fernandez-Palacios, J.M.; Flores, O.; et al. Human impact, climate and dispersal strategies determine plant invasion on islands. J. Biogeogr. 2021. early view. [Google Scholar] [CrossRef]
- Irl, S.; Steinbauer, M.; Messinger, J.; Blume-Werry, G.; Palomares-Martínez, A.; Beierkuhnlein, C.; Jentsch, A. Burned and devoured—Introduced herbivores, fire and the endemic flora of the high elevation ecosystem on La Palma, Canary Islands. Arct. Antarct. Alp. Res. 2014, 46, 859–869. Available online: http://www.gobiernodecanarias.org/boc/2006/237/011.html (accessed on 28 October 2020). [CrossRef] [Green Version]
- Frankham, R. Do island populations have less genetic variation than mainland populations? Heredity 1997, 78, 311–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Mazuecos, M.; Vargas, P. Genetically depauperate in the continent but rich in oceanic islands: Cistus monspeliensis (Cistaceae) in the Canary Islands. PLoS ONE 2011, 6, e17172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Verdugo, C.; Calleja, J.A.; Vargas, P.; Silva, L.; Moreira, O.; Pulido, F. Polyploidy and microsatellite variation in the relict tree Prunus lusitanica L.: How effective are refugia in preserving genotypic diversity of clonal taxa? Mol. Ecol. 2013, 22, 1546–1557. [Google Scholar] [CrossRef] [PubMed]
- Mort, M.E.; Soltis, D.E.; Soltis, P.S.; Francisco-Ortega, J.; Santos-Guerra, A. Phylogenetics and evolution of the Macaronesian clade of Crassulaceae inferred from nuclear and chloroplast sequence data. Syst. Bot. 2002, 27, 271–288. [Google Scholar]
- Fernández-Palacios, J.M.; de Nascimento, L.; Otto, R.; Delgado, J.D.; García-del-Rey, E.; Arévalo, J.R.; Whittaker, R.J. A reconstruction of Palaeo-Macaronesia, with particular reference to the long-term biogeography of the Atlantic island laurel forests. J. Biogeogr. 2011, 38, 226–246. [Google Scholar] [CrossRef]
- Van den Boogard, P. The origin of the Canary Island Seamount Province—New ages of old seamounts. Sci. Rep. 2017, 3, 2107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weigelt, P.; Steinbauer, M.; Cabral, J.S.; Kreft, H. Late Quaternary climate change shapes island biodiversity. Nature 2016, 532, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Aparicio, A.; Martín-Hernanz, S.; Parejo-Farnés, C.; Arroyo, J.; Lavergne, S.; Yeşilyurt, E.B.; Yeşilyurt, M.-L.; Rubio, E.; Albaladejo, R.G. Phylogenetic reconstruction of the genus Helianthemum (Cistaceae) using plastid and nuclear DNA-sequences: Systematic and evolutionary inferences. Taxon 2017, 66, 868–885. [Google Scholar] [CrossRef] [Green Version]
- Sánchez Meseguer, A.; Aldasoro, J.J.; Sanmartín, I. Bayesian inference of phylogeny, morphology and range evolution reveals a complex evolutionary history in St. John’s wort (Hypericum). Mol. Phylogenetics Evol. 2013, 67, 379–403. [Google Scholar] [CrossRef] [PubMed]
- Graham, R.E.; Reyes-Betancourt, J.A.; Chapman, M.A. Inter-island differentiation and contrasting patterns of diversity in the iconic Canary Island sub-alpine endemic Echium wildpretii (Boraginaceae). Syst. Biodivers. 2021, 19, 507–521. [Google Scholar] [CrossRef]
- Schmitt, T. Molekulare Biogeographie; Haupt Verlag: Bern, Germany, 2020; 504p. [Google Scholar]
- Fernández-Palacios, J.M.; Negrín, Z.; Fernández Lugo, S.; Arévalo, J.R.; de Nascimento, L. Terrestrial Biota Checklist of the Chinijo Archipelago and Lobos (Canary Islands). Rev. Sci. Insul. 2018, 1, 51–86. [Google Scholar] [CrossRef]
- Puppo, P.; Curto, M.; Gusmão-Guedes, J.; Cochofel, J.; Pérez de Paz, P.L.; Bräuchler, C.; Meimberg, H. Molecular phylogenetics of Micromeria (Lamiaceae) in the Canary Islands, diversification and inter-island colonization patterns inferred from nuclear genes. Mol. Phylogenetics Evol. 2015, 89, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Plants of the World Online (Kew). Available online: http://www.plantsoftheworldonline.org/ (accessed on 5 June 2021).
- World Flora Online. Available online: http://www.worldfloraonline.org/ (accessed on 5 June 2021).
- The Plant List. Available online: http://www.theplantlist.org/ (accessed on 5 June 2021).
- Roskov, Y.; Abucay, L.; Orrell, T.; Nicolson, D.; Bailly, N.; Kirk, P.M.; Bourgoin, T.; DeWalt, R.E.; Decock, W.; De Wever, A.; et al. (Eds.) Species 2000 & ITIS Catalogue of Life, 2018 Annual Checklist. Species 2000; Naturalis: Leiden, The Netherlands, 2018; Available online: www.catalogueoflife.org/annual-checklist/2018 (accessed on 5 June 2021).
- GBIF. Available online: https://www.gbif.org/ (accessed on 5 June 2021).
- TRY. Available online: https://www.try-db.org/TryWeb/Home.php (accessed on 5 June 2021).
- Kattge, J.; Bönisch, G.; Díaz, S.; Lavorel, S.; Prentice, I.C.; Leadley, P.; Tautenhahn, S.; Werner, G.D.A.; Aakala, T.; Abedi, M.; et al. TRY plant trait database—Enhanced coverage and open access. Glob. Chang. Biol. 2020, 26, 119–188. [Google Scholar] [CrossRef] [Green Version]
- Chamberlain, S.; Szocs, E. Taxize—Taxonomic search and retrieval in R. F1000Research 2013, 2, 191. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- International Plant Names Index IPNI. Available online: https://www.ipni.org/ (accessed on 5 June 2021).
- Nezadal, W.; Lindacher, R.; Welss, W. Lokalendemiten und Phytodiversität der westkanadischen Inseln La Palma und La Gomera. Feddes Repert. 1999, 110, 19–30. [Google Scholar] [CrossRef]
- Barone, R. Notas corológicas sobre dos especies de Atriplex, L. (Chenopodiaceae) en Tenerife, Islas Canarias. Botánica Macaronésica 2003, 24, 161–164. [Google Scholar]
- Leibbach, R.; Peter, R. Bemerkungen zu den Orchideen von La Palma (Kanarische Inseln). J. Eur. Orch. 2007, 39, 55–70. [Google Scholar]
- Otto, R.; Scholz, H.; Scholz, S. Supplements to the flora of the Canary Islands, Spain: Poaceae. Willdenowia 2008, 38, 491–496. [Google Scholar] [CrossRef] [Green Version]
- Prina, A.O.; Martinez-Laborde, J.B. A taxonomic revision of Crambe section Dendrocrambe (Brassicaceae). Bot. J. Linn. Soc. 2008, 156, 291–304. [Google Scholar] [CrossRef] [Green Version]
- Verloove, F.; Reyes-Betancort, J. Additions to the flora of Tenerife (Canary Islands, Spain). Collect. Bot. 2011, 30, 63–78. [Google Scholar] [CrossRef] [Green Version]
- Sánchez de Lorenzo Cáceres, J. Sobre la presencia de Balanites aegyptiaca (L.) Del. (Zygophyllaceae) en Tenerife, Islas Canarias. Bouteloua 2012, 10, 82–84. [Google Scholar]
- Reisigl, H. Gefährdete endemische Blütenpflanzen der Trockeninsel Fuerteventura: Herkunft, Ökologie, Gesellschaft. Bauhinia 2013, 24, 39–52. [Google Scholar]
- Verloove, F. New xenophytes from Gran Canaria (Canary Islands, Spain), with emphasis on naturalized and (potentially) invasive species. Collect. Bot. 2013, 32, 59–82. [Google Scholar] [CrossRef] [Green Version]
- Santos Guerra, A.; Reyes Betancort, J.A.; Padrón Mederos, M.A.; Mesa Coello, R. Plantas poco o nada conocidas de la flora vascular silvestre de las Islas Canarias. Bot. Complut. 2013, 37, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Verloove, F.; Guiggi, A. Some new xenophytes from Fuerteventura (Canary Islands, Spain). Bouteloua 2013, 13, 13,–42. [Google Scholar]
- Santos Guerra, A.; Reyes-Betancort, J. Nuevas adiciones y citas de interés para la flora autóctona de las islas Canarias. Vieraea 2014, 42, 249–258. [Google Scholar]
- Otto, R.; Verloove, F. New xenophytes from La Palma (Canary Islands, Spain), with emphasis on naturalized and (potentially) invasive species. Collect. Bot. 2016, 35, e001. [Google Scholar]
- Marrero Rodríguez, Á. Eucaliptos en Gran Canaria, identificación y corología. Hacia una reseña histórica. Botánica Macaronésica 2016, 29, 91–137. [Google Scholar]
- Salas-Pascual, M.; Quintana Vega, G. Salvinia molesta D. S. Mitch. (Salviniaceae), nueva cita para Canarias y España. Botánica Macaronésica 2016, 29, 73–81. [Google Scholar]
- Verloove, F.; Alves, P. New vascular plant records for the western part of the Iberian Peninsula (Portugal and Spain). Folia Bot. Extrem. 2016, 10, 5–23. [Google Scholar]
- Galan de Mera, A.; Linares-Perea, E.; Vicente-Orellana, J.A. Taraxacum (Asteraceae) in the Azores, Madeira and the Canary Island. Ann. Bot. Fenn. 2017, 54, 273–285. [Google Scholar] [CrossRef]
- Verloove, F. New xenophytes from the Canary Islands (Gran Canaria and Tenerife; Spain). Acta Bot. Croat. 2017, 76, 120–131. [Google Scholar] [CrossRef] [Green Version]
- Verloove, F.; Ojeda-Land, E.; Smith, G.F.; Guiggi, A.; Reyes-Betancort, J.A.; Samarín, C.; González Hernández, A.; Barone, R. New records of naturalised and invasive cacti (Cactaceae) from Gran Canaria and Tenerife, Canary Islands, Spain. Bradleya 2017, 35, 58–79. [Google Scholar] [CrossRef]
- Otto, R.; Verloove, F. New xenophytes from La Palma (Canary Islands, Spain), with emphasis on naturalized and (potentially) invasive species—Part 2. Collect. Bot. 2018, 37, e005. [Google Scholar]
- Expósito, A.B.; Siverio, A.; Bermejo, L.A.; Sobrino-Vesperinas, E. Checklist of alien plant species in a natural protected area: Anaga Rural Park (Tenerife, Canary Islands) effect of human infrastructures on their abundance. Plant Ecol. Evol. 2018, 151, 142–152. [Google Scholar] [CrossRef]
- Verloove, F.; Salas-Pascual, M.; Marrero Rodríguez, Á. New records of alien plants for the flora of Gran Canaria (Canary Islands, Spain). Flora Mediterr. 2018, 28, 119–135. [Google Scholar]
- Portero Alvarez, A.M.; Martin-Carbajal Gonzalez, J.; Reyes-Betancort, J.; Mesa Coello, R.A. Lotus gomerythus (Fabaceae-Loteae) Spec. Nova. Botanica Macaronesica 2019, 30, 89–98. [Google Scholar]
- Marrero, A. Vicia tenoi (Fabaceae) una nueca especie del macizo de Teno, Tenerife (Islas Canarias). Bot. Macaronesica 2019, 30, 153–156. [Google Scholar]
- Arango Toro, O. Aeoniun lioi (Crassulaceae): Una nueva especie de Tenerife, Islas Canarias. Bot. Macaronesica 2019, 30, 7–22. [Google Scholar]
- Verloove, F.; Thiede, J.; Marrero Rodríguez, Á.; Salas-Pascual, M.; Reyes-Betancort, J.; Ojeda-Land, E.; Smith, G. A synopsis of feral Agave and Furcraea (Agavaceae, Asparagaceae s. lat.) in the Canary Islands (Spain). Plant Ecol. Evol. 2019, 152, 470–498. [Google Scholar] [CrossRef]
- Marrero Rodríguez, Á. Adiciones corológicas a la flora vascular de Gran Canaria, especies xenófitas, ocasionales o potenciales invasoras. Botánica Macaronésica 2019, 30, 121–142. [Google Scholar]
- Otto, R.; Verloove, F. New xenophytes from La Palma (Canary Islands, Spain), with emphasis on naturalized and (potentially) invasive species—Part 3. Collect. Bot. 2020, 39, e001. [Google Scholar]
- Verloove, F.; Déniz Suárez, E.; Salas Pascual, M. New records of non-native vascular plants in Gran Canaria (Spain, Canary Islands). Flora Mediterr. 2020, 30, 121–136. [Google Scholar]
- White, O.W.; Reyes-Betancort, J.A.; Chapman, M.A.; Carine, M.A. Geographical isolation, habitat shifts and hybridisation in the diversification of the Macaronesian endemic genus Argyranthemum (Asteraceae). New Phytol. 2020, 228, 1953–1971. [Google Scholar] [CrossRef] [PubMed]
- Verloove, F. New records in vascular plants alien to Tenerife (Spain, Canary Islands). Biodivers. Data J. 2021, 9, e62878. [Google Scholar] [CrossRef] [PubMed]
- White, O.W.; Reyes-Betancort, J.A.; Chapman, M.A.; Carine, M.A. Recircumscription of the Canary Island endemics Argyranthemum broussonetii and A. callichrysum (Asteraceae: Anthemideae) based on evolutionary relationships and morphology. Willdenowia 2021, 51, 129–139. [Google Scholar] [CrossRef]
- Nürk, N.M.; Linder, H.P.; Onstein, R.E.; Larcombe, M.J.; Hughes, C.E.; Fernández, L.P.; Schlüter, P.M.; Valente, L.; Beierkuhnlein, C.; Cutts, V.; et al. Diversification in evolutionary arenas—Assessment and synthesis. Ecol. Evol. 2020, 10, 6163–6182. [Google Scholar] [CrossRef] [PubMed]
- Darwin, C. Letter no. 2130. To, J.D. Hooker, 1 August 1857. Darwin Correspondence Project. Available online: https://www.darwinproject.ac.uk/ (accessed on 5 July 2021).
- Irl, S.; Schweiger, A.; Medina, F.M.; Fernández-Palacios, J.M.; Harter, D.; Jentsch, A.; Provenzale, A.; Steinbauer, M.J.; Beierkuhnlein, C. An island view of endemic rarity—Environmental drivers and consequences for nature conservation. Divers. Distrib. 2017, 23, 1132–1142. [Google Scholar] [CrossRef] [Green Version]
- García-Verdugo, C.; Baldwin, B.G.; Fay, M.F.; Caujapé-Castells, J. Life history traits and patterns of diversification in oceanic archipelagos: A meta-analysis. Bot. J. Linn. Soc. 2014, 174, 334–348. [Google Scholar] [CrossRef] [Green Version]
- EU Biodiversity Strategy for 2030. Available online: https://www.eea.europa.eu/policy-documents/eu-biodiversity-strategy-for-2030-1 (accessed on 17 July 2021).
- Gangoso, L.; Donázar, J.A.; Scholz, S.; Palacios, C.J.; Hiraldo, F. Contradiction in conservation of island ecosystems: Plants, introduced herbivores and avian scavengers in the Canary Islands. Biodivers. Conserv. 2006, 15, 2231–2248. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Palacios, J.M.; Martin Esquivel, J.L. Naturaleza de las Islas Canarias—Ecologia y Conservation; Publicaciones Turquesa: Santa Cruz de Tenerife, Spain, 2001; 474p. [Google Scholar]
- Reyes-Betancort, J.A.; Santos-Guerra, A.; Guma, I.R.; Humphries, C.J.; Carine, M.A. Diversity, rarity and the evolution and conservation of the Canary Islands endemic flora. An. Jardín Botánico Madr. 2008, 65, 25–45. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
On the island of La Palma (Canary Islands) just one species of the genus Carlina L. with narrow leaves is listed in most floras. This species (C. falcata Svent.) (a) is relatively abundant and widespread on the island. However, in remote parts of northern cliffs a subspecies of Carlina canariensis (L.f.) Cav. was recorded recently (b), which has not been scientifically described up to now. A regular publication of this taxon following the rules of ICN (International Code of Nomenclature for algae, fungi, and plants) is in preparation. This illustrates that even for well-known genera of woody species, new species’ descriptions are still to be expected.
Figure 1.
On the island of La Palma (Canary Islands) just one species of the genus Carlina L. with narrow leaves is listed in most floras. This species (C. falcata Svent.) (a) is relatively abundant and widespread on the island. However, in remote parts of northern cliffs a subspecies of Carlina canariensis (L.f.) Cav. was recorded recently (b), which has not been scientifically described up to now. A regular publication of this taxon following the rules of ICN (International Code of Nomenclature for algae, fungi, and plants) is in preparation. This illustrates that even for well-known genera of woody species, new species’ descriptions are still to be expected.

Figure 2.
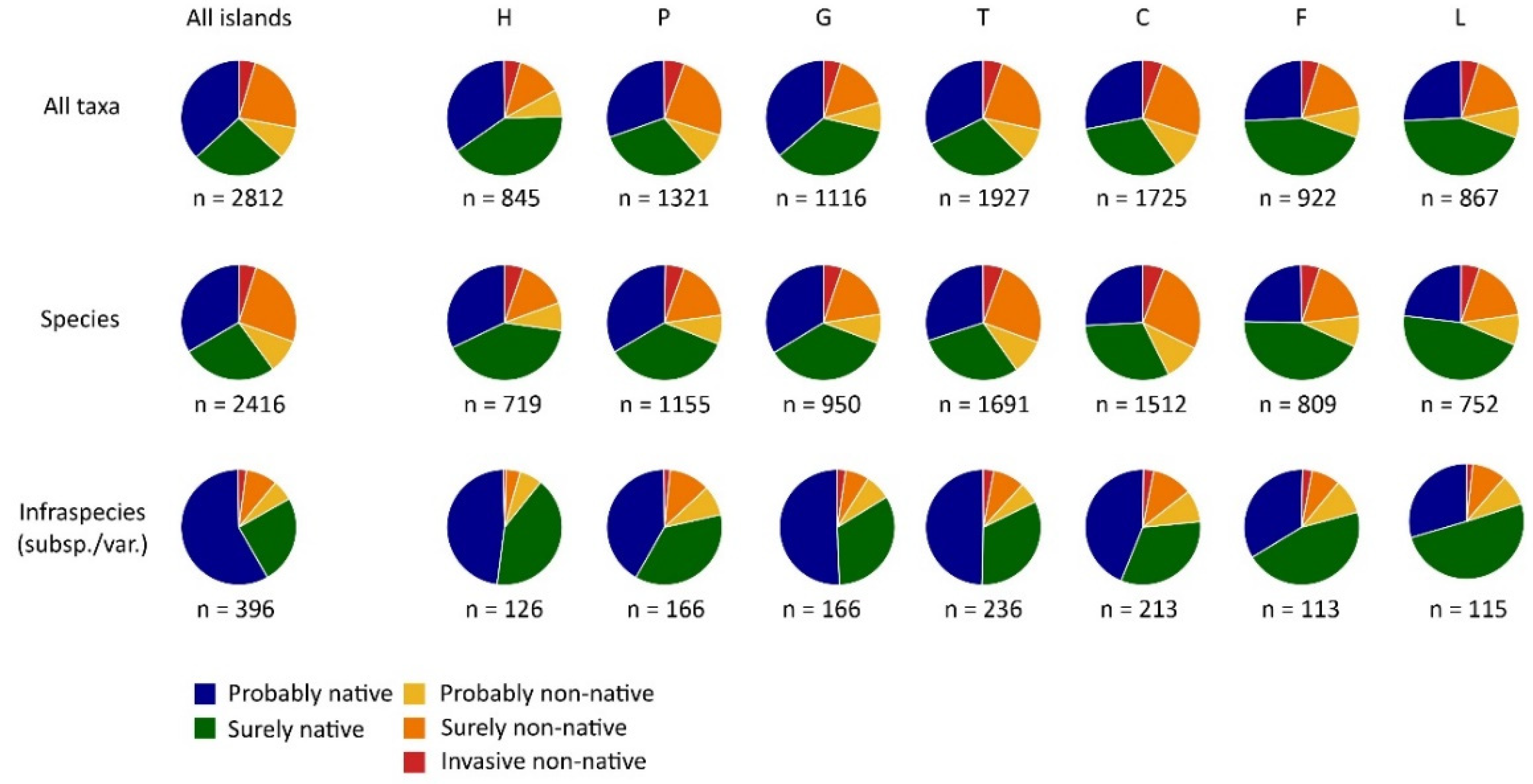
Categories of native and non-native taxa in the Canary Islands and for the individual islands El Hierro (H), La Palma (P), La Gomera (G), Tenerife (T), Gran Canaria (C), Fuerteventura (F) and Lanzarote (L) based on the here presented checklist. Proportions are given at the level of all taxa, and separately for species and infra specific taxa (sub-species and varieties). Absolute numbers of taxa are given below the pie charts. Generally, the proportion of accepted native infraspecific units (subspecies, varieties) is higher compared with the accepted species. Highest numbers of taxa are listed for the large islands with pronounced topography and diverse climatic conditions.
Figure 2.
Categories of native and non-native taxa in the Canary Islands and for the individual islands El Hierro (H), La Palma (P), La Gomera (G), Tenerife (T), Gran Canaria (C), Fuerteventura (F) and Lanzarote (L) based on the here presented checklist. Proportions are given at the level of all taxa, and separately for species and infra specific taxa (sub-species and varieties). Absolute numbers of taxa are given below the pie charts. Generally, the proportion of accepted native infraspecific units (subspecies, varieties) is higher compared with the accepted species. Highest numbers of taxa are listed for the large islands with pronounced topography and diverse climatic conditions.

Figure 3.
Categories for native taxa subdivided into native non-endemic species (blue), multi-island endemics (green) and single island endemics (yellow). Proportions are given at the level of all taxa, and separately for species and infraspecific taxa (sub-species and varieties). Absolute numbers of taxa are given below the pie charts (n). The relations are illustrated for the entire Canary Islands archipelago (All islands) and for the individual islands El Hierro (H), La Palma (P), La Gomera (G), Tenerife (T), Gran Canaria (C), Fuerteventura (F) and Lanzarote (L). Surprisingly, approximately one quarter of all species in the entire archipelago are SIE. The highest proportions of endemic taxa relate to the islands with pronounced topography and diverse climatic conditions (H, P, G, T, C).
Figure 3.
Categories for native taxa subdivided into native non-endemic species (blue), multi-island endemics (green) and single island endemics (yellow). Proportions are given at the level of all taxa, and separately for species and infraspecific taxa (sub-species and varieties). Absolute numbers of taxa are given below the pie charts (n). The relations are illustrated for the entire Canary Islands archipelago (All islands) and for the individual islands El Hierro (H), La Palma (P), La Gomera (G), Tenerife (T), Gran Canaria (C), Fuerteventura (F) and Lanzarote (L). Surprisingly, approximately one quarter of all species in the entire archipelago are SIE. The highest proportions of endemic taxa relate to the islands with pronounced topography and diverse climatic conditions (H, P, G, T, C).

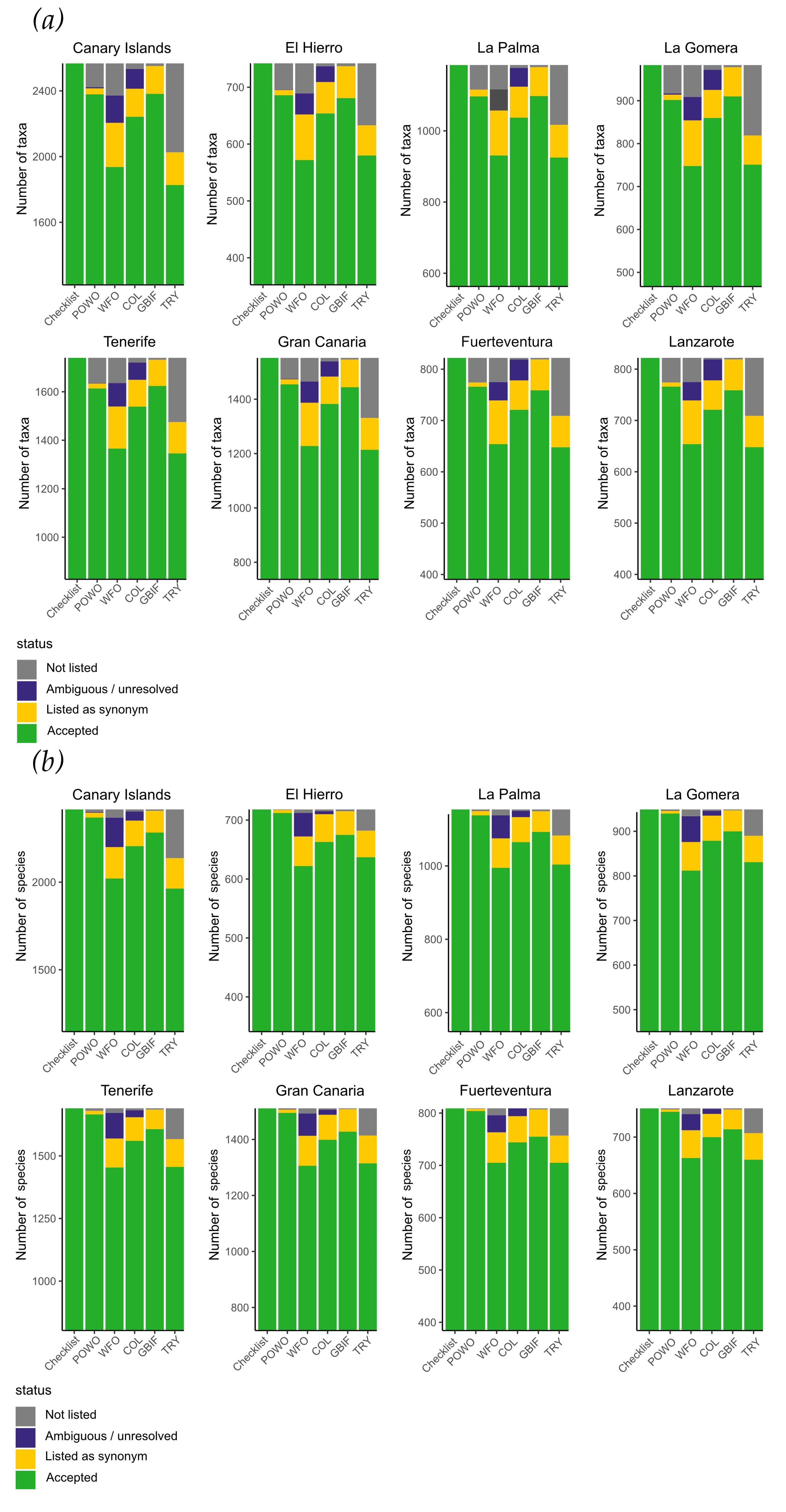
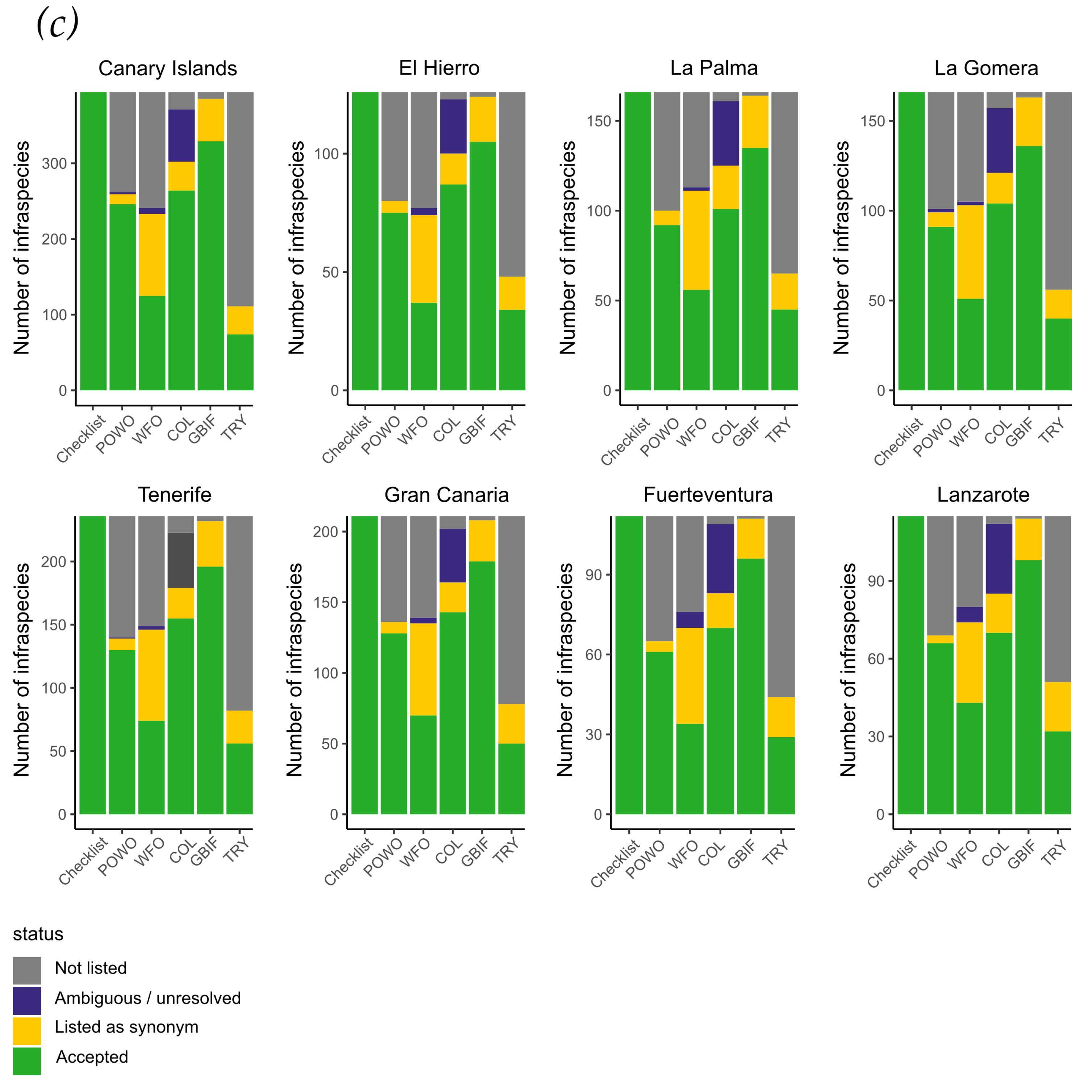
Figure 4.
Representation of the accepted taxa suggested in this checklist for the Canary Islands in global databases. (a) for all taxa, (b) for species, and (c) for infraspecific taxa (subspecies and varieties). Note that scales for (a,b) do not begin with 0 taxa/species but start at 50% of each individual y-axis to work out differences. Axis (c) for infraspecific taxa start in 0. Numbers of accepted taxa diverge between POWO and WFO, reflecting that POWO was taken as a reference. COL numbers are close to those of POWO, which supports the decision to select this database for reference. The patterns for individual islands resemble the pattern of the entire archipelago.
Figure 4.
Representation of the accepted taxa suggested in this checklist for the Canary Islands in global databases. (a) for all taxa, (b) for species, and (c) for infraspecific taxa (subspecies and varieties). Note that scales for (a,b) do not begin with 0 taxa/species but start at 50% of each individual y-axis to work out differences. Axis (c) for infraspecific taxa start in 0. Numbers of accepted taxa diverge between POWO and WFO, reflecting that POWO was taken as a reference. COL numbers are close to those of POWO, which supports the decision to select this database for reference. The patterns for individual islands resemble the pattern of the entire archipelago.


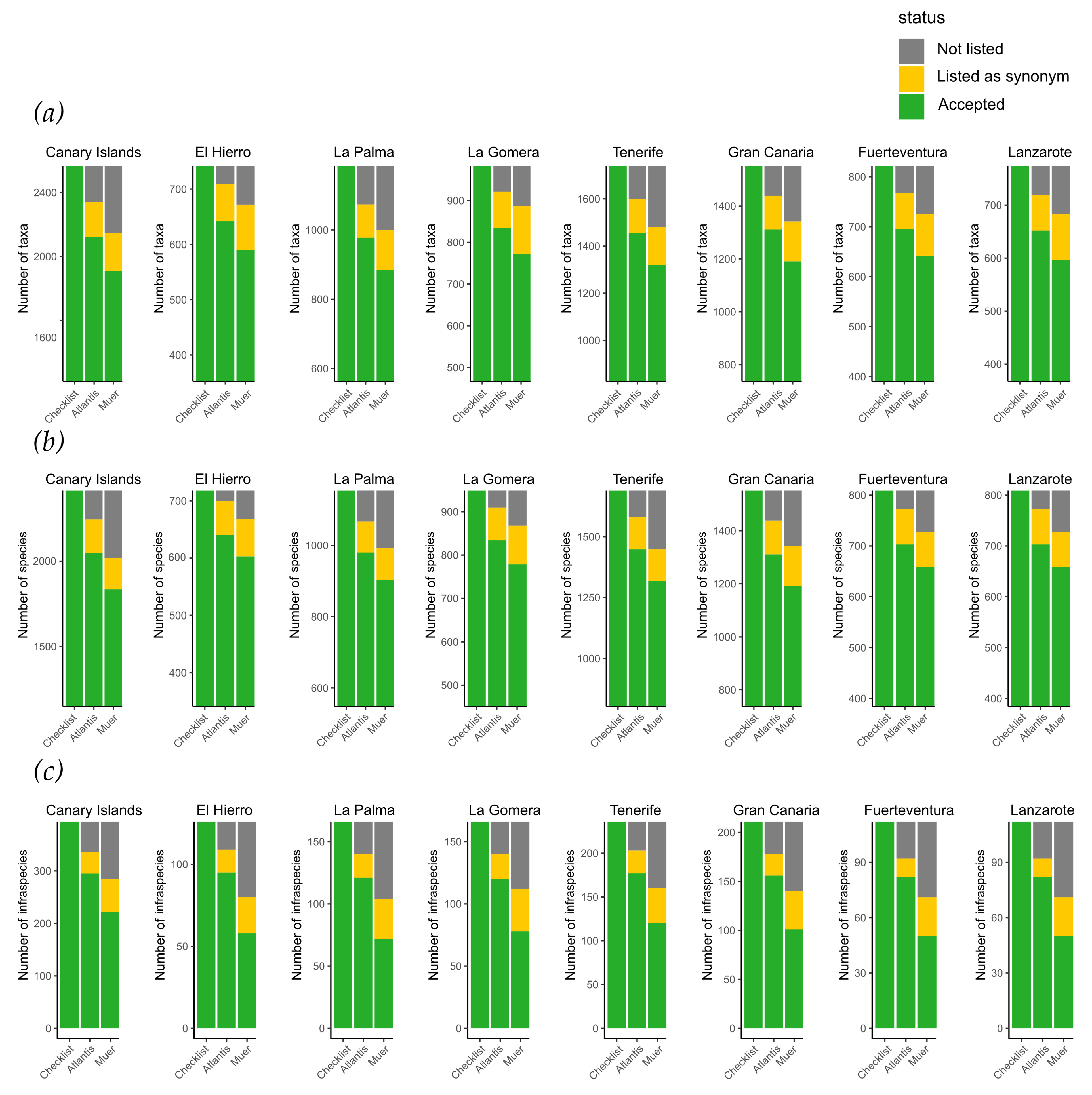
Figure 5.
Comparing numbers of the here presented FloCan Checklist (without hybrid taxa, synonyms, and non-naturalized ornamental garden and park plants) with the Atlantis database [22] and with the taxa of Muer et al. [34] separately for (a) taxa, (b) species and (c) infraspecies (subspecies, varieties). Categories given are “not listed” in the respective database, “listed as synonym” and “accepted”. This comparison illustrates how many of the accepted taxa in FloCan are also reflected in other established references.
Figure 5.
Comparing numbers of the here presented FloCan Checklist (without hybrid taxa, synonyms, and non-naturalized ornamental garden and park plants) with the Atlantis database [22] and with the taxa of Muer et al. [34] separately for (a) taxa, (b) species and (c) infraspecies (subspecies, varieties). Categories given are “not listed” in the respective database, “listed as synonym” and “accepted”. This comparison illustrates how many of the accepted taxa in FloCan are also reflected in other established references.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Numbers of accepted plant taxa (families, genera, species, subspecies, varieties) in the FloCan checklist for the Canary Island archipelago in comparison with other current plant lists (Atlantis, Muer) and with reference to the accepted taxa in international taxonomic data bases Plants of the World Online (POWO), Catalogue of Life (COL), World Flora Online (WFO), GBIF and TRY.
Table 1.
Numbers of accepted plant taxa (families, genera, species, subspecies, varieties) in the FloCan checklist for the Canary Island archipelago in comparison with other current plant lists (Atlantis, Muer) and with reference to the accepted taxa in international taxonomic data bases Plants of the World Online (POWO), Catalogue of Life (COL), World Flora Online (WFO), GBIF and TRY.
| Taxonomic Level | FloCan | Atlantis Biota | Muer et al. | POWO | COL | WFO | GBIF | TRY |
|---|---|---|---|---|---|---|---|---|
| Year | 2021 | 2021 | 2016 | 2021 | 2021 | 2021 | 2021 | 2021 |
| Families | 171 | 162 | 155 | 171 | 166 | 164 | 170 | 166 |
| Genera | 863 | 747 | 699 | 854 | 815 | 775 | 832 | 788 |
| Species | 2416 | 2050 | 1834 | 2370 | 2207 | 2022 | 2284 | 1965 |
| Infraspecifics | 396 | 295 | 222 | 247 | 264 | 125 | 329 | 74 |
Table 2.
Numbers of endemic species and endemic infraspecific taxa (subspecies, varieties) in current plant lists of the Canary Island archipelago.
Table 2.
Numbers of endemic species and endemic infraspecific taxa (subspecies, varieties) in current plant lists of the Canary Island archipelago.
| Taxonomic Level | FloCan | Atlantis Biota | Muer et al. |
|---|---|---|---|
| Year | 2021 | 2021 | 2016 |
| Species | 608 | 541 | 499 |
| Infraspecifics | 197 | 147 | 152 |
Table 3.
Numbers of non-native species and non-native infraspecific taxa (subspecies, varieties) in current plant lists of the Canary Island archipelago.
Table 3.
Numbers of non-native species and non-native infraspecific taxa (subspecies, varieties) in current plant lists of the Canary Island archipelago.
| Taxonomic Level | FloCan | Atlantis Biota | Muer et al. |
|---|---|---|---|
| Year | 2021 | 2021 | 2016 |
| Species | 964 | 781 | 662 |
| Infraspecifics | 67 | 44 | 16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Beierkuhnlein, C.; Walentowitz, A.; Welss, W. FloCan—A Revised Checklist for the Flora of the Canary Islands. Diversity 2021, 13, 480. https://0-doi-org.brum.beds.ac.uk/10.3390/d13100480
AMA Style
Beierkuhnlein C, Walentowitz A, Welss W. FloCan—A Revised Checklist for the Flora of the Canary Islands. Diversity. 2021; 13(10):480. https://0-doi-org.brum.beds.ac.uk/10.3390/d13100480
Chicago/Turabian StyleBeierkuhnlein, Carl, Anna Walentowitz, and Walter Welss. 2021. "FloCan—A Revised Checklist for the Flora of the Canary Islands" Diversity 13, no. 10: 480. https://0-doi-org.brum.beds.ac.uk/10.3390/d13100480
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.