
Unequivocal Differences in Predation Pressure on Large Carabid Beetles between Forestry Treatments



MTA-ELTE-MTM Ecology Research Group, Biological Institute, Eötvös Loránd University, Pázmány Péter Sétány 1/C, H-1117 Budapest, Hungary
*
Author to whom correspondence should be addressed.
Diversity 2021, 13(10), 484; https://0-doi-org.brum.beds.ac.uk/10.3390/d13100484
Submission received: 17 September 2021
/
Revised: 30 September 2021
/
Accepted: 1 October 2021
/
Published: 3 October 2021
(This article belongs to the Special Issue Faunistical and Ecological Studies on Carabid Beetles)
Abstract
:Carabid beetles (Coleoptera: Carabidae) are considered as one of the most cardinal invertebrate predatory groups in many ecosystems, including forests. Previous studies revealed that the predation pressure provided by carabids significantly regulates the ecological network of invertebrates. Nevertheless, there is no direct estimation of the predation risk on carabids, which can be an important proxy for the phenomenon called ecological trap. In our study, we aimed to explore the predation pressure on carabids using 3D-printed decoys installed in two types of forestry treatments, preparation cuts and clear cuts, and control plots in a Hungarian oak–hornbeam forest. We estimated the seasonal, diurnal and treatment-specific aspects of the predation pressure on carabids. Our results reveal a significantly higher predation risk on carabids in both forestry treatments than in the control. Moreover, it was also higher in the nighttime than daytime. Contrarily, no effects of season and microhabitat features were found. Based on these clues we assume that habitats modified by forestry practices may act as an ecological trap for carabids. Our findings contribute to a better understanding of how ecological interactions between species may change in a modified forest environment.

1. Introduction
Predation by various terrestrial animals can be one of the most important drivers of carabid evolution and, together with abiotic factors, determines their activity patterns [1]. Although carabids have various morphological and chemical defense mechanisms to avoid or reduce predation risk [2,3,4], they can also shift their behavior in space and time by being active only in a certain part of the day or using different habitat patches than predators. Another option is to be hidden in the soil, under the leaf litter or understory vegetation. For instance, studies describing the activity patterns of some large carabid species reported relatively long periods of beetle inactivity between movements that can last even for a couple of days [5,6]. In some individuals, regardless of sex, this no-movement behavior dominated during the tracking period [7]. Elek et al. [8] suggested that these periods when beetles are hidden and not moving could be a predator avoidance strategy.
Although carabids are often studied as important natural enemies [9,10,11], they can also be predated due to their middle position in the food chain. In temperate forests of the Northern Hemisphere, carabids represent potential prey for a large spectrum of predators, including bats, hedgehogs, shrews, raccoon dogs, wild boars and frogs [3,12,13,14]. Interactions between carabids and their predators might change due to shifts in the distribution of suitable habitat patches of various sizes within forest stands as a result of forestry practices [15,16,17]. This is especially true for Europe, where the majority of temperate forests have a semi-natural origin [18]. Therefore, it is crucial to explore whether the spatial differences created by forestry practices can affect the predation pressure on carabids. In addition, it can be an important issue to assess whether a certain treatment can act as an ecological trap for carabids [19], a habitat which is suitable for foraging or breeding, despite the fact that mortality can be higher there than in the unmanaged control forest stands.
There is an emerging need to estimate the structural changes in communities towards understanding ecosystem functions. Predation is well known as a key driver of these processes, potentially forming the community itself (i.e., keystone predation), and any change in predation intensity can reflect changes in ecosystem functions [20]. The technique involving artificial decoys as prey provides a powerful tool to assess relative rates of predation or predation pressure across various treatments [21]. Artificial prey is relatively cheap and easy to manipulate, and it is not invasive as it does not involve living specimens [22,23,24]. This can be an issue especially for rare or endangered species with low population densities where collection is nearly impossible [14]. Using 3D-printed decoys seems to be more suitable than involving dried fragile specimens that can be easily damaged [25]. Other frequently used materials, such as clay or plasticine [9,10,26], cannot replicate the narrow parts of beetles’ body, such as legs and antennae [14]. Yet, these materials and techniques can be used for modeling various invertebrates with simple body shapes, such as snails, slugs, caterpillars and earthworms, and is called the sentinel prey method [9,27].
In this paper, we experimentally tested the predation pressure on carabids in the Hungarian managed temperate forest using real-sized 3D-printed models as decoys. As a model organism for 3D decoys, we selected a forest generalist, Carabus coriaceus L., 1758, a large species commonly occurring in Hungarian oak–hornbeam forests that sensitively reacts to forestry practices [8,28]. Two distinct types of forestry treatments, clear cuts and preparations cuts, were used for the field experiment. They considerably differ from surrounded unmanaged forest stands in terms of tree height and the cover of understory vegetation, leaf litter and bare soil [29] as well as in microclimatic conditions [30,31]. Hence, we expected that the predation pressure will vary between forestry treatments and undisturbed control stands due to the different availability of shelters formed by dense ground vegetation or leaf litter. Moreover, the different activity patterns of potential predators may presume that the predation pressure will also have diurnal and seasonal aspects based on predator activity and breeding period. Following these clues, we tested how the predation pressure on large carabids is affected at the different spatio-temporal scales. In particular, we focused on microhabitat characteristic, habitat type and time to determine the most influential factor(s) of the predation pressure in the studied forest stands.
2. Materials and Methods
2.1. Study Area
We conducted our experiment in the vicinity of Pilisszántó village in the Pilis Mountains, the northern part of Hungary (N 47°40′, E 18°54′). The study area (40 ha) is a structurally homogenous, 80-year-old, managed two-layered sessile oak–hornbeam forest. The structure of the stand is a result of the past and recent management under the shelterwood silvicultural system. The upper canopy layer is dominated by sessile oak (Quercus petraea (Matt.) Liebl, 1784) with an average height of 21 m and a mean diameter of 28 cm at breast height. The second most abundant tree species, hornbeam (Carpinus betulus L., 1753), forms a secondary canopy layer with an average height of 11 m and a mean diameter of 12 cm. Other admixing tree species include Quercus cerris L., 1753, Fagus sylvatica L., 1753, Prunus avium L., 1753 and Fraxinus ornus L., 1753. The shrub layer is scarce, mainly consisting of the regeneration of hornbeam and F. ornus, and the understory cover is dominated by mesic forest plants, such as Carex pilosa Scopoli, 1772 and Melica uniflora Retz, 1779 [16,29].
Our study was implemented as a part of the Pilis Forestry System Experiment [32] where four forestry treatments representing two different silvicultural systems were established in 2014. The main aim of this project is to explore the major effect of various treatments on natural forest regeneration and the biodiversity of several taxa including plants, enchytraeid worms, spiders and ground beetles (see Elek et al. [16] for further details). For the field experiment conducted in this study, we used two of the four implemented treatments representing characteristic stages of rotation forestry system: (1) Clear cut (CC) was a circular clear-cutting area of 80 m diameter surrounded by a closed-canopy stand. (2) Preparation cut (P) was created when 30% of the total basal area of the dominant tree layer and the whole secondary tree layer were removed in a spatially uniform way in a circle of 80 m diameter. These two treatments were chosen due to their strong effects on carabids at different levels, from community composition to individual activity [8,16,17].
2.2. Three-Dimensional Printed Decoys and Field Experiment
The 3D model of Carabus coriaceus was prepared in Blender 2.8 [33] based on high-resolution photos of real-sized individuals. One of the advantages of using C. coriaceus is its relatively large body size (33–40 mm), making the species suitable for detailed 3D printing. Unlike the other large species, C. coriaceus is unified in color (black) without iridescence and it is also one of the most abundant species in the area [17]. The beetle model (Supplementary Material Figure S1) was converted into printing data by KISSlicer 1.6, and then life-size three-dimensional decoys were generated by the 3D printer (DeltiQ M, developed by TriLAB Group s.r.o., Brno, Czech Republic). Black polylactic acid (PLA) filament was used as a production material. The printed decoy was the same size as the real specimen (Figure 1a).
The field experiment was conducted in two seasons, spring and autumn, which corresponded with the highest activity peaks of carabids in the area [16,17]. In both treatments (CC and P) and control plots (C), we installed 10 decoys (two lines, each with five beetles) on the ground with an A5 cardboard sheet beneath them. Each plot (both treatments and control) was replicated in three blocks, resulting in nine sampling plots. The distance between decoys was approximately 2.5 m. Decoys were positioned in the center of the green-colored sheet to avoid any unintended attraction due to conspicuous sheet colors (Figure 1b). The exact position was marked by two perpendicular lines drawn on the paper; the decoy was installed at their intersection to be able to record possible predation events. We recorded the cover of bare soil, litter, herbal and shrub layer (in %) in a one-meter-radius circle around each decoy and the number of surrounding trees (see summary Table 1). These environmental variables can potentially affect the distribution of carabids via the availability of shelters [7,8]. In total, 90 C. coriaceus decoys were installed per season. Decoys were checked twice a day, in the morning and evening, for six consecutive days (7–12 September 2020 and 8–13 June 2021). The exposed decoy was considered attacked when it had been turned or moved/relocated from its original position, including its disappearance or damage, such as scratches, bites and missing parts. For turns, we measured the angle; if relocated, the distance was recorded. Although decoy turns suggest that the potential predator was interested in the exposed decoy, the relocation or scratches are clear signs of a predation attempt. After measurements, decoys were repositioned or replaced by new ones when necessary.
2.3. Data Analyses
The predation pressure was considered as a ratio between attack and no-attack events on 10 decoys in a plot per measurement session. This response was coded as two-column matrix [attack; no attack] using the cbind function in R 3.6.1 [34] where all analyses were conducted. We used generalized linear models (the glm function) with a binomial distribution and logit link function. Three different models were built considering various temporal and spatial scales. In the first model, treatment (factor with three levels: control, preparation cuts, clear cuts) was used as a single explanatory variable (spatial scale). In the second model, treatment, cover of leaf litter, bare soil and number of trees in the plot were included (micro-spatial scale). Cover of herbs and shrubs were excluded from the analyses due to a strong negative correlation with leaf litter (Pearson r = −0.48 for herb layer and r = −0.70 for shrubbery; see Supplementary Material Figure S2). The treatment, daytime (factor with two levels: day and night) and season (factor with two levels: spring and autumn) factors were used in the third model (spatio-temporal scale). Then, using the model.sel function from the ‘MuMIn’ package [35], this set of models was tested to select the best model(s) based on information criterion corrected for small sample sizes (Akaike Information Criterion—AICc, [36]). The best model was selected as the most parsimonious explanation of the data when Δ AICc was higher than two (Δ AICc > 2) for other models.
3. Results
In total, we recorded 108 attack events based on 1800 observation events (i.e., the predation rate was 6%). Turn was the most common event in 87 cases, followed by scratches or broken parts in 13 cases and 8 relocations. The majority of turns were between 5 and 10° from the original position, and the maximal turn reached 45°. The mean distance for relocation was 11.8 cm and in one case the decoy was taken. Focusing on possible predators, we observed bird droppings (N = 2) and wild boar hair (N = 1) on turned or scratched decoys. Some of the recorded predation events are shown on Figure 1c–g.
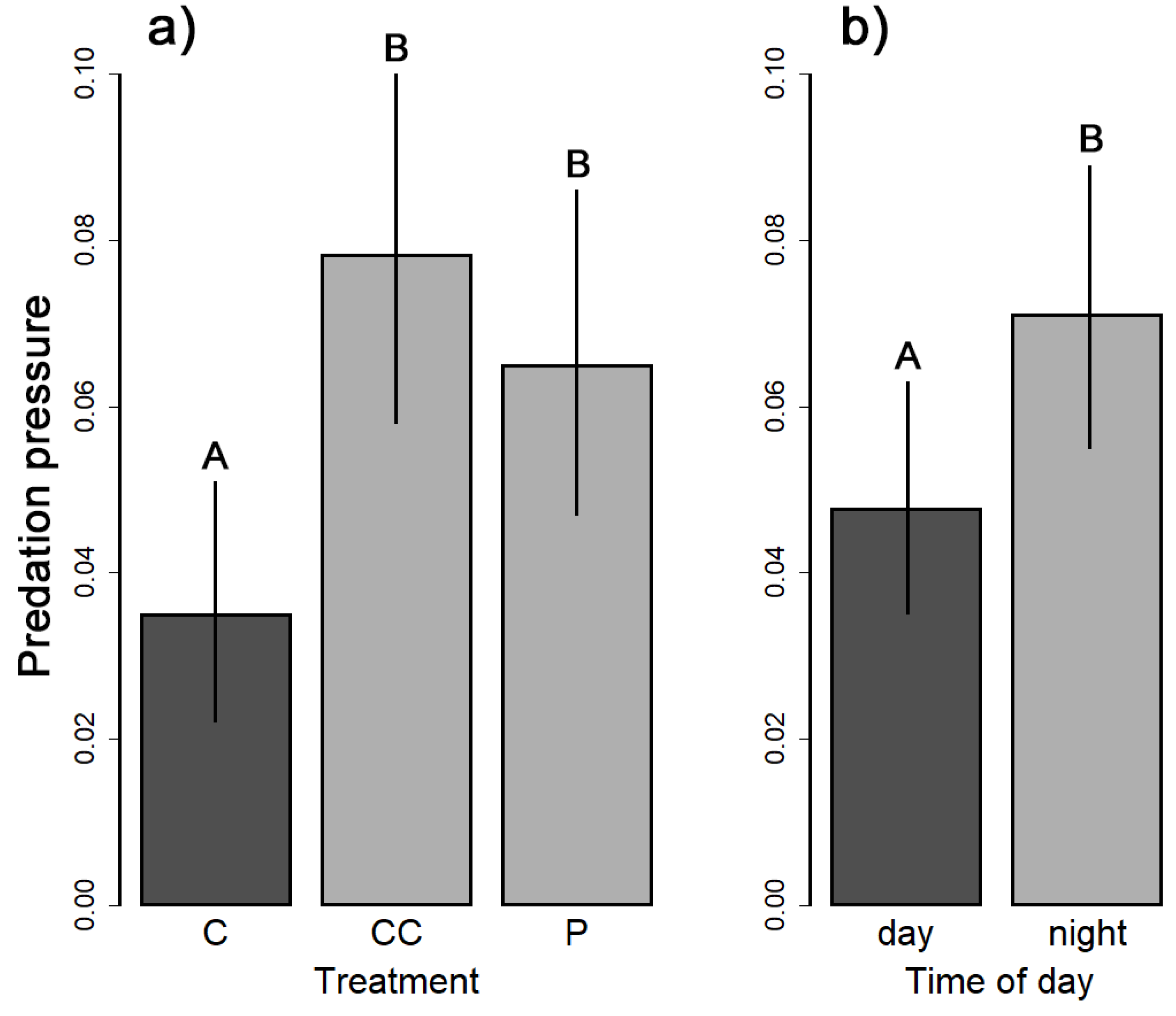
The model selection showed that the “spatio-temporal scale” model was the best one, suggesting spatial and temporal constraints are the most influential explanatory variables on predation pressure (Table 2). Therefore, we revealed that the predation pressure was significantly higher in both treatments than in the control forest (Table 3, Figure 2a). Moreover, the pressure was higher during nights than daytime (Figure 2b). On the contrary, no effect of season or environmental variables was confirmed.
4. Discussion
We revealed that 3D-printed decoys represent a suitable approach for testing predation pressure on large carabids, since predators interacted with them. The predation pressure in our study area was affected by spatial as well as temporal constraints. We recorded more attack events in both treatments than in the control. Moreover, the predation pressure on the exposed carabid decoys was higher at night than during the daytime, and it did not correlate with any microhabitat features, such as leaf litter or bush cover, as they are not important for predators of large carabids. Taking into account our previous findings on habitat utilization by large carabids [28], we suggest that both forestry treatments can act as an ecological trap for carabids, as these plots are used only temporarily and with a higher risk of mortality.
Although marks on decoys made from polylactic acid filament cannot deliver a detailed determination of predator identity in comparison with plasticine models [37], we were able to determine some of the predators and suggest other possible candidates. Considering the types of attack, the turn was the most common event, occurring in most cases. Decoy turn may suggest an interest of the potential predator in the exposed decoy, investigating its edibility. Contrarily, scratches and relocations are clear signs of feeding attempts. Wild boar (Sus scrofa L., 1758) was determined directly since we found its hair on one relocated decoy. Indeed, various carabid species were recorded as a part of the wild boar’s diet [38]. From other mammalian predators, we do not have direct evidence for attacking decoys, but their interest can be presumed as they commonly occur in oak–hornbeam forests of the Pilis Mountains. Eurasian badger (Meles meles L., 1758) is already known for feeding on carabids, including large ones, such as C. coriaceus [39,40]. Other potential predators could be the northern white-breasted hedgehog (Erinaceus roumanicus Martin, 1838) or the greater mouse-eared bat (Myotis myotis Borkhausen, 1797), which collect ground-dwelling insects directly from the ground, and large carabids were previously found in its droppings [13]. Moreover, we also recorded bird droppings on two turned decoys. Since we found these droppings exclusively in the spring season, carabids may provide food not only for adult birds but also for nestlings. In addition, Eurasian nuthatch (Sitta europaea L., 1758) can feed its young by a high proportion of large carabid species, including the genera Carabus, Calosoma or Pterostichus [12].
The predation pressure was higher in treatments than in the control undisturbed forest, thus the risk of beetles’ mortality evidently increased in these treatments. Some large carabids, including C. coriaceus, can penetrate clear cuts and preparation cuts in relatively high numbers, likely for foraging, since there is a high competition for the limited resources in the structurally homogeneous undisturbed forest [28]. This kind of habitat utilization is rather transient, since individuals are able to leave the treatment sites within a couple of days [8,28]. There is a high risk of predation in both treatments modified by forestry practices; therefore, these habitats may act as an ecological trap for carabids. The ecological trap is a habitat that is actively selected by individuals despite higher costs of utilization, such as increased mortality and low breeding success [19]. Modified habitat patches are often mentioned as an example of the ecological trap for various animal groups, including insects [19,41]. Our results do not support the increasing disturbance hypothesis [42], suggesting that predation pressure in modified habitats is lower than in undisturbed ones due to the declining number of predators [43,44]. Treatments included in our study were, however, relatively small (with diameter of 80 m) in comparison with landscape gradients and surrounded by undisturbed semi-natural forest. We can presume that large predators, such as wild boars, could easily move across plots regardless of disturbances.
Forestry treatments differ in terms of microclimatic conditions and the structure of understory vegetation from the control forest [29,30]. Nevertheless, we found no effect of any environmental variable on the predation pressure. Microhabitat features, such as the cover of leaf litter or herbs, of a particular patch are important factors for the distribution of carabids [45,46,47], but not for their predators, as they move at larger scales and likely consider the same patch as structurally homogeneous. Additionally, the predation pressure was higher at night than in the daytime. This seems to be coherent with the fact that C. coriaceus is predominantly nocturnal [6,8] and most mammalian predators are more active during nights, which may result in an overlap in activity of prey and potential predators. One of the options to avoid predators is to switch movement activity to the daytime, when the predation pressure is lower. Indeed, beetles, previously considered as strictly nocturnal, were observed to be active during the daytime as well (see [6,8] for activity patterns of C. coriaceus). However, there is still a risk of diurnal predators, such as birds. Another option is to minimize the encounters with predators by being hidden as much as possible in shelters. Based on radio-tracking studies, large carabids are able to be inactive for a couple of days, hidden under leaf litter or burrowed in soil [5,6,7]. In C. coriaceus, this anti-predation behavior was observed especially in undisturbed forest [8,28], where a high amount of leaf litter seems to be ideal to hide.
It is also worth mentioning some methodological perspectives and limitations of using 3D-printed decoys for testing predation pressure. This approach is suitable for recording the exploratory behavior of various predators, as they frequently interacted with decoys and turned them. Exact predator identification was, however, limited, as the used printing material (polylactic acid filament) is rather tough for recording soft and small bites. Originally, we experimented with a softer and more elastic material; nevertheless, we could not print narrower body parts, such as legs or antennas. The surface of 3D-printed decoys also could not mimic the structural colors of the real carabid cuticle, especially if the model species has metallic iridescent colorization [14], and this may possibly bias attractiveness for predators. Although we installed decoys far away from each other, there is a chance that the same animals might attack them, leading to an overestimation of predation pressure.
5. Conclusions
Nowadays, ecological studies attempt to focus on the functional aspect of the studied ecosystems [17,48]. Although community measures are widely used, new facets appear and serve as a good proxy for ecosystem functioning including animal behavior. The estimation of predation risk can reflect how animals can select their habitat for foraging, breeding or overwintering. However, for making such a conclusion, the knowledge of the spatio-temporal habitat use of beetles is required, suggesting that other methods should be employed alongside decoys, such as pitfall trapping and radio telemetry. Understanding how interactions between prey and predators change in modified habitats is crucial for better predictions of species-specific responses to habitat alteration. We revealed that habitats modified by forestry practices may act as an ecological trap for carabids. We also proved that the estimation of predation on ground-dwelling predators can be a good proxy for identifying key habitats for conservation [49]. These clues mentioned above may also help us to scrutinize the potential knowledge gaps in understanding how animal behavior can be generalized as a functional component of community-level measures.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/d13100484/s1, Figure S1: Three-dimensional model of the Carabus coriaceus decoy used for 3D printing. The original file in .stl format (205 MB) can be provided upon request, Figure S2: The overview of the Spearman’s rank correlation test for exploring the relationship between microhabitat features: cover of bare soil, leaf litter, herbal layer, shrubbery and the presence of tree.
Author Contributions
Conceptualization, J.R. and Z.E.; methodology, J.R. and Z.E.; formal analysis, J.R. and Z.E.; investigation, J.R. and Z.E.; writing—original draft preparation, J.R. and Z.E.; writing—review and editing, J.R. and Z.E.; visualization, J.R. and Z.E.; funding acquisition, Z.E. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Hungarian Research Fund (NKFIH-K 128441) and by the Hungarian Academy of Sciences (MTA KEP, Ecology for Society project).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on reasonable request from the corresponding author.
Acknowledgments
The authors thank Pilisi Parkerdő Ltd. (Péter Csépányi, Viktor Farkas, Gábor Szenthe, László Simon) for the maintenance of the experimental site. We are also grateful to Jan Chloupek and Alena Vláčilová for modeling and printing 3D decoys as well as Péter Ódor for comments on the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lövei, G.L.; Sunderland, K.D. Ecology and behavior of ground beetles (Coleoptera: Carabidae). Annu. Rev. Entomol. 1996, 41, 231–256. [Google Scholar] [CrossRef]
- Brandmayr, P.; Bonacci, T.; Giglio, A.; Talarico, F.F.; Brandmayr, T.Z. The evolution of defence mechanisms in carabid beetles: A review. In Life and Time: The Evolution of Life and Its History; Casellato, S., Burighel, P., Minelli, A., Eds.; Cleup: Padova, Italy, 2009; pp. 25–43. [Google Scholar]
- Sugiura, S. Predators as drivers of insect defenses. Entomol. Sci. 2020, 23, 316–337. [Google Scholar] [CrossRef]
- Giglio, A.; Vommaro, M.L.; Brandmayr, P.; Talarico, F. Pygidial Glands in Carabidae, an Overview of Morphology and Chemical Secretion. Life 2021, 11, 562. [Google Scholar] [CrossRef]
- Niehues, F.J.; Hockmann, P.; Weber, F. Genetics and dynamics of a Carabus auronitens metapopulation in the Westphalian Lowlands (Coleoptera, Carabidae). Ann. Zool. Fenn. 1996, 33, 85–96. [Google Scholar]
- Riecken, U.; Raths, U. Use of radio telemetry for studying dispersal and habitat use of Carabus coriaceus L. Ann. Zool. Fenn. 1996, 33, 109–116. [Google Scholar]
- Růžičková, J.; Veselý, M. Movement activity and habitat use of Carabus ullrichii (Coleoptera: Carabidae): The forest edge as a mating site? Entomol. Sci. 2018, 21, 76–83. [Google Scholar] [CrossRef]
- Elek, Z.; Růžičková, J.; Ódor, P. Individual decisions drive the changes in movement patterns of ground beetles between forestry management types. Biologia 2021, 1–10. [Google Scholar] [CrossRef]
- Ferrante, M.; Cacciato, A.L.; Lövei, G.L. Quantifying predation pressure along an urbanisation gradient in Denmark using artificial caterpillars. Eur. J. Entomol. 2014, 111, 649–654. [Google Scholar] [CrossRef] [Green Version]
- Lövei, G.L.; Ferrante, M. A review of the sentinel prey method as a way of quantifying invertebrate predation under field conditions. Insect Sci. 2017, 24, 528–542. [Google Scholar] [CrossRef]
- Boetzl, F.A.; Konle, A.; Krauss, J. Aphid cards—Useful model for assessing predation rates or bias prone nonsense? J. Appl. Entomol. 2020, 144, 74–80. [Google Scholar] [CrossRef]
- Thiele, H.U. Carabid Beetles in Their Environments; Springer: Berlin/Heidelberg, Germany, 1997; 369p. [Google Scholar] [CrossRef]
- Graclik, A.; Wasielewski, O. Diet composition of Myotis myotis (Chiroptera, Vespertilionidae) in western Poland: Results of fecal analyses. Turk. J. Zool. 2012, 36, 209–213. [Google Scholar] [CrossRef]
- Fukuda, S.; Konuma, J. Using three-dimensional printed models to test for aposematism in a carabid beetle. Biol. J. Linn. Soc. Lond. 2019, 128, 735–741. [Google Scholar] [CrossRef]
- Negro, M.; Vacchiano, G.; Berretti, R.; Chamberlain, D.E.; Palestrini, C.; Motta, R.; Rolando, A. Effects of forest management on ground beetle diversity in alpine beech (Fagus sylvatica L.) stands. For. Ecol. Manag. 2014, 328, 300–309. [Google Scholar] [CrossRef] [Green Version]
- Elek, Z.; Kovács, B.; Aszalós, R.; Boros, G.; Samu, F.; Tinya, F.; Ódor, P. Taxon-specific responses to different forestry treatments in a temperate forest. Sci. Rep. 2018, 8, 16990. [Google Scholar] [CrossRef] [Green Version]
- Elek, Z.; Růžičková, J.; Ódor, P. Functional plasticity of carabids can presume better the changes in community composition than taxon-based descriptors. Ecol. Appl. 2021, e02460. [Google Scholar] [CrossRef]
- Bengtsson, J.; Nilsson, S.G.; Franc, A.; Menozzi, P. Biodiversity, disturbances, ecosystem function and management of European forests. For. Ecol. Manag. 2000, 132, 39–50. [Google Scholar] [CrossRef]
- Ewers, R.M.; Didham, R.K. Confounding factors in the detection of species responses to habitat fragmentation. Biol. Rev. 2006, 81, 117–142. [Google Scholar] [CrossRef] [PubMed]
- Schneider, G.; Krauss, J.; Steffan-Dewenter, I. Predation rates on semi-natural grasslands depend on adjacent habitat type. Basic Appl. Ecol. 2013, 14, 614–621. [Google Scholar] [CrossRef]
- González-Gómez, P.L.; Estades, C.F.; Simonetti, J.A. Strengthened insectivory in a temperate fragmented forest. Oecologia 2006, 148, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Belovsky, G.E.; Slade, J.B.; Stockhoff, B.A. Susceptibility to predation for different grasshoppers: An experimental study. Ecology 1990, 71, 624–634. [Google Scholar] [CrossRef]
- Pitt, W.C. Effects of multiple vertebrate predators on grasshopper habitat selection: Trade-offs due to predation risk, foraging, and thermoregulation. Evol. Ecol. 1999, 13, 499–515. [Google Scholar] [CrossRef]
- Bartholomew, A.; El Moghrabi, J. Seasonal preference of darkling beetles (Tenebrionidae) for shrub vegetation due to high temperatures, not predation or food availability. J. Arid Environ. 2018, 156, 34–40. [Google Scholar] [CrossRef]
- Pearson, D.L. The function of multiple anti-predator mechanisms in adult tiger beetles (Coleoptera: Cicindelidae). Ecol. Entomol. 1985, 10, 65–72. [Google Scholar] [CrossRef]
- Sam, K.; Remmel, T.; Molleman, F. Material affects attack rates on dummy caterpillars in tropical forest where arthropod predators dominate: An experiment using clay and dough dummies with green colourants on various plant species. Entomol. Exp. Appl. 2015, 157, 317–324. [Google Scholar] [CrossRef]
- Howe, A.G.; Nachman, G.; Lövei, G.L. Predation pressure in Ugandan cotton fields measured by a sentinel prey method. Entomol. Exp. Appl. 2015, 154, 161–170. [Google Scholar] [CrossRef]
- Růžičková, J.; Bérces, S.; Ackov, S.; Elek, Z. Individual movement of large carabids as a link for activity density patterns in various forestry treatments. Acta Zool. Acad. Sci. Hung. 2021, 67, 77–86. [Google Scholar] [CrossRef]
- Tinya, F.; Kovács, B.; Prättälä, A.; Farkas, P.; Aszalós, R.; Ódor, P. Initial understory response to experimental silvicultural treatments in a temperate oak-dominated forest. Eur. J. For. Res. 2019, 138, 65–77. [Google Scholar] [CrossRef] [Green Version]
- Kovács, B.; Tinya, F.; Guba, E.; Németh, C.; Sass, V.; Bidló, A.; Ódor, P. The short-term effects of experimental forestry treatments on site conditions in an oak–hornbeam forest. Forests 2018, 9, 406. [Google Scholar] [CrossRef] [Green Version]
- Kovács, B.; Tinya, F.; Németh, C.; Ódor, P. Unfolding the effects of different forestry treatments on microclimate in oak forests: Results of a 4-yr experiment. Ecol. Appl. 2020, 30, e02043. [Google Scholar] [CrossRef] [Green Version]
- Effect of Forestry Treatments on Forest Site, Regeneration and Biodiversity. An Experimental Study. Available online: https://piliskiserlet.ecolres.hu/en (accessed on 16 September 2021).
- Blender—A 3D Modelling and Rendering Package. Available online: http://www.blender.org (accessed on 16 September 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Bartoń, K. MuMIn: Multi-Model Inference. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 16 September 2021).
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information Theoretic Approach; Springer: New York, NY, USA, 2002; 488p. [Google Scholar]
- Low, P.A.; Sam, K.; McArthur, C.; Posa, M.R.C.; Hochuli, D.F. Determining predator identity from attack marks left in model caterpillars: Guidelines for best practice. Entomol. Exp. Appl. 2014, 152, 120–126. [Google Scholar] [CrossRef]
- Schley, L.; Roper, T.J. Diet of wild boar Sus scrofa in Western Europe, with particular reference to consumption of agricultural crops. Mammal Rev. 2003, 33, 43–56. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, J.G. The Diet and Habitat Utilisation of the Badger (Meles meles) in an Area to the South of Durham City. Ph.D. Thesis, Durham University, Durham, UK, 1992. [Google Scholar]
- Marassi, M.; Biancardi, C. Diet of the Eurasian badger (Meles meles) in an area of the Italian Prealps. Hystrix It. J. Mamm. 2002, 13, 19–28. [Google Scholar] [CrossRef]
- Ries, L.; Fagan, W.F. Habitat edges as a potential ecological trap for an insect predator. Ecol. Entomol. 2003, 28, 567–572. [Google Scholar] [CrossRef] [Green Version]
- Gray, J.S. Effects of environmental stress on species rich assemblages. Biol. J. Linn. Soc. Lond. 1989, 37, 19–32. [Google Scholar] [CrossRef]
- Eötvös, C.B.; Lövei, G.L.; Magura, T. Predation pressure on sentinel insect prey along a riverside urbanization gradient in Hungary. Insects 2020, 11, 97. [Google Scholar] [CrossRef] [Green Version]
- Eötvös, C.B.; Magura, T.; Lövei, G.L. A meta-analysis indicates reduced predation pressure with increasing urbanization. Landsc. Urban Plan. 2018, 180, 54–59. [Google Scholar] [CrossRef]
- Niemelä, J.; Spence, J.R.; Spence, D.H. Habitat associations and seasonal activity of ground-beetles (Coleoptera, Carabidae) in central Alberta. Can. Entomol. 1992, 124, 521–540. [Google Scholar] [CrossRef]
- Pearce, J.L.; Venier, L.A.; McKee, J.; Pedlar, J.; McKenney, D. Influence of habitat and microhabitat on carabid (Coleoptera: Carabidae) assemblages in four stand types. Can. Entomol. 2003, 135, 337–357. [Google Scholar] [CrossRef]
- Wehnert, A.; Wagner, S. Niche partitioning in carabids: Single-tree admixtures matter. Insect Conserv. Divers. 2019, 12, 131–146. [Google Scholar] [CrossRef]
- Murray, B.D.; Holland, J.D.; Summerville, K.S.; Dunning, J.B.; Saunders, M.R.; Jenkins, M.A. Functional diversity response to hardwood forest management varies across taxa and spatial scales. Ecol. Appl. 2017, 27, 1064–1081. [Google Scholar] [CrossRef] [PubMed]
- Shochat, E.; Warren, P.S.; Faeth, S.H.; McIntyre, N.E.; Hope, D. From patterns to emerging processes in mechanistic urban ecology. Trends Ecol. Evol. 2006, 21, 186–191. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Printed decoy of Carabus coriaceus in comparison with a real specimen (a). Decoys installed on the green cardboard (b) were checked for turns (c), broken or bent parts (d), predation attempts by birds (e), wild boars (f) or relocations (g).
Figure 1.
Printed decoy of Carabus coriaceus in comparison with a real specimen (a). Decoys installed on the green cardboard (b) were checked for turns (c), broken or bent parts (d), predation attempts by birds (e), wild boars (f) or relocations (g).

Figure 2.
The effect of treatment (a) and time of day (b) on the predation pressure. Vertical lines represent a 95% confidence interval and different capital letters above the bars indicate significant differences based on Tukey multiple comparisons of means.
Figure 2.
The effect of treatment (a) and time of day (b) on the predation pressure. Vertical lines represent a 95% confidence interval and different capital letters above the bars indicate significant differences based on Tukey multiple comparisons of means.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Environmental characteristics of forestry treatments represented as the cover of bare soil, leaf litter, herbal layer and shrubbery and the average number of trees per plot.
Table 1.
Environmental characteristics of forestry treatments represented as the cover of bare soil, leaf litter, herbal layer and shrubbery and the average number of trees per plot.
| Cover of (Mean ± SEM, in %) | No. of Trees per Plot (Mean) | ||||
|---|---|---|---|---|---|
| Treatment | Bare Soil | Leaf Litter | Herbal Layer | Shrubs | |
| Control | 2.51 ± 0.52 | 68.70 ± 2.68 | 28.25 ± 2.59 | 0.58 ± 0.37 | 4.67 |
| Clear cut | 3.63 ± 1.04 | 10.91 ± 1.68 | 36.10 ± 2.20 | 49.35 ± 2.90 | 0.50 |
| Preparation cut | 4.11 ± 0.97 | 33.36 ± 2.39 | 50.40 ± 2.76 | 12.10 ± 2.14 | 2.67 |
Table 2.
Summary of the model selection using estimations based on the calculated AICc value of the models, serving as the weight of evidence in favor of the different models. The most parsimonious model (delta < 2) is emphasized in bold.
Table 2.
Summary of the model selection using estimations based on the calculated AICc value of the models, serving as the weight of evidence in favor of the different models. The most parsimonious model (delta < 2) is emphasized in bold.
| Model | df | LogLik | AICs | Delta | Weight |
|---|---|---|---|---|---|
| Spatio-temporal scale | 5 | −179.890 | 370.1 | 0.00 | 0.833 |
| Spatial scale | 3 | −183.925 | 374.0 | 3.86 | 0.121 |
| Micro-spatial scale | 6 | −181.705 | 375.9 | 5.77 | 0.046 |
Table 3.
The effects of treatment, daytime, season and environmental variables on the predation pressure. Significant effects are in bold, marginal in italics.
Table 3.
The effects of treatment, daytime, season and environmental variables on the predation pressure. Significant effects are in bold, marginal in italics.
| Model | Explanatory Variables | χ2 | df | p |
|---|---|---|---|---|
| Spatio-temporal scale | treatment | 11.334 | 2 | 0.003 |
| daytime | 4.444 | 1 | 0.035 | |
| season | 3.625 | 1 | 0.056 | |
| Spatial scale | treatment | 11.286 | 2 | 0.004 |
| Micro-spatial scale | treatment | 0.203 | 2 | 0.903 |
| leaf litter | 0.973 | 1 | 0.323 | |
| bare soil | 2.404 | 1 | 0.121 | |
| tree | 0.010 | 1 | 0.918 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Růžičková, J.; Elek, Z. Unequivocal Differences in Predation Pressure on Large Carabid Beetles between Forestry Treatments. Diversity 2021, 13, 484. https://0-doi-org.brum.beds.ac.uk/10.3390/d13100484
AMA Style
Růžičková J, Elek Z. Unequivocal Differences in Predation Pressure on Large Carabid Beetles between Forestry Treatments. Diversity. 2021; 13(10):484. https://0-doi-org.brum.beds.ac.uk/10.3390/d13100484
Chicago/Turabian StyleRůžičková, Jana, and Zoltán Elek. 2021. "Unequivocal Differences in Predation Pressure on Large Carabid Beetles between Forestry Treatments" Diversity 13, no. 10: 484. https://0-doi-org.brum.beds.ac.uk/10.3390/d13100484
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.