
Recent Invaders in Small Mediterranean Islands: Wild Boars Impact Snakes in Port-Cros National Park

Abstract
:1. Introduction
2. Materials and Methods
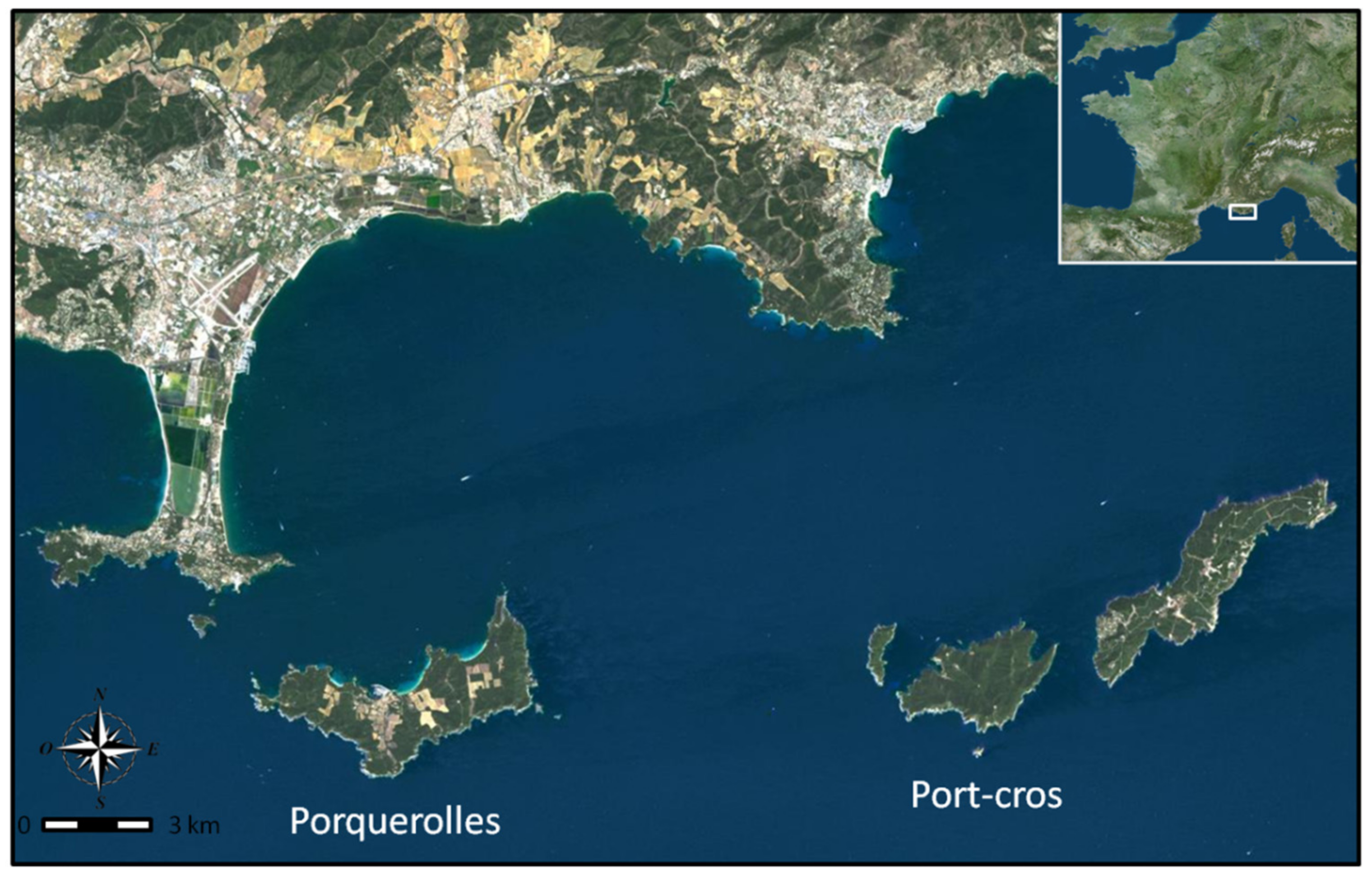
2.1. Study Areas
2.2. Wild Boar Arrival and Proliferation
2.3. Snake Population Monitoring
2.4. Analyses
2.5. Ethics Statement
3. Results
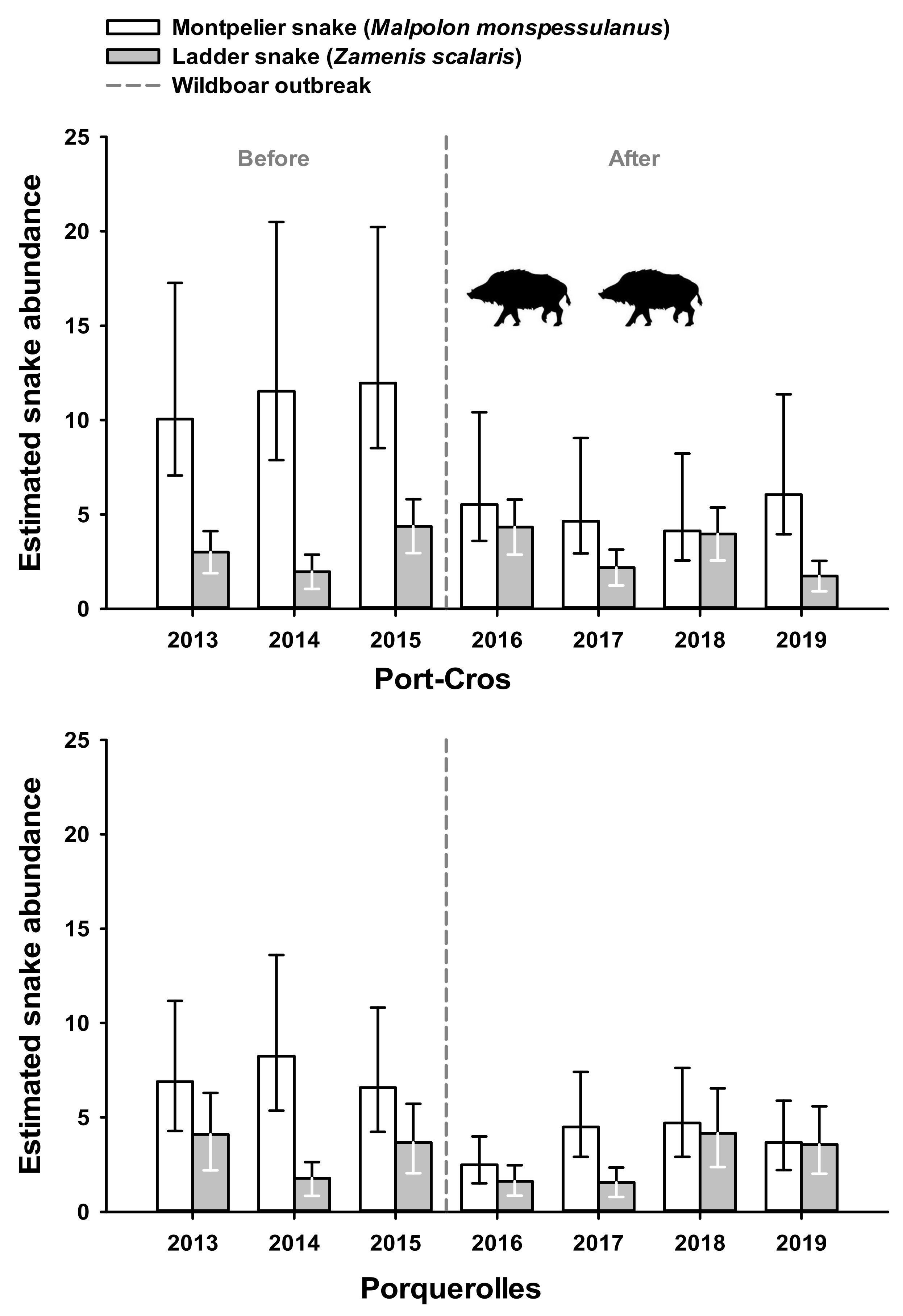
3.1. Index of Snake Abundance
3.2. Demographic Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Losos, J.B.; Ricklefs, R.E. Adaptation and diversification on islands. Nature 2009, 457, 830–836. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.C.; Kueffer, C. Island biodiversity in the Anthropocene. Annu. Rev. Environ. Resour. 2019, 44, 31–60. [Google Scholar] [CrossRef]
- Courchamp, F.; Hoffmann, B.D.; Russell, J.C.; Leclerc, C.; Bellard, C. Climate change, sea-level rise, and conservation: Keeping island biodiversity afloat. Trends Ecol. Evol. 2014, 29, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Courchamp, F.; Chapuis, J.L.; Pascal, M. Mammal invaders on islands: Impact, control and control impact. Biol. Rev. 2003, 78, 347–383. [Google Scholar] [CrossRef] [Green Version]
- Chapuis, J.L.; Boussès, P.; Barnaud, G. Alien mammals, impact and management in the French subantarctic islands. Biol. Conserv. 1994, 67, 97–104. [Google Scholar] [CrossRef]
- Spatz, D.R.; Zilliacus, K.M.; Holmes, N.D.; Butchart, S.H.; Genovesi, P.; Ceballos, G.; Tershy, B.R.; Croll, D.A. Globally threatened vertebrates on islands with invasive species. Sci. Adv. 2017, 3, e1603080. [Google Scholar] [CrossRef] [Green Version]
- Colautti, R.I.; MacIsaac, H.J. A neutral terminology to define ‘invasive’ species. Divers. Distrib. 2004, 10, 135–141. [Google Scholar] [CrossRef]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species: A Selection from the Global Invasive Species Database; ISSG: Auckland, New Zealand, 2000; Available online: www.issg.org/booklet.pdf (accessed on 21 June 2021).
- Ickes, K.; Paciorek, C.J.; Thomas, S.C. Impacts of nest construction by native pigs (Sus scrofa) on lowland Malaysian rain forest saplings. Ecology 2005, 86, 1540–1544. [Google Scholar] [CrossRef] [Green Version]
- Massei, G.; Kindberg, J.; Licoppe, A.; Gačić, D.; Šprem, N.; Kamler, J.; Baubet, E.; Hohmann, U.; Monaco, A.; Ozoliņš, J.; et al. Wild boar populations up, numbers of hunters down? A review of trends and implications for Europe. Pest Manag. Sci. 2014, 71, 492–500. [Google Scholar] [CrossRef]
- Tack, J. Wild boar (Sus scrofa) populations in Europe. In A Scientific Review of Population Trends and Implications for Management; European Landowners’ Organization: Brussels, Belgium, 2018; p. 56. [Google Scholar]
- Sáaez--Royuela, C.; Telleriia, J.L. The increased population of the wild boar (Sus scrofa L.) in Europe. Mammal. Rev. 1986, 16, 97–101. [Google Scholar] [CrossRef]
- Vetter, S.G.; Puskas, Z.; Bieber, C.; Ruf, T. How climate change and wildlife management affect population structure in wild boars. Sci. Rep. 2020, 10, 7298. [Google Scholar] [CrossRef]
- Barrios-Garcia, M.N.; Ballari, S.A. Impact of wild boar (Sus scrofa) in its introduced and native range: A review. Biol. Invasions 2012, 14, 2283–2300. [Google Scholar] [CrossRef]
- Graitson, E.; Barbraud, C.; Bonnet, X. Catastrophic impact of wild-boars: Insufficient hunting pressure pushes snakes to the brink. Anim. Conserv. 2018, 22, 165–176. [Google Scholar] [CrossRef]
- Kupfer, A.; Langel, R.; Scheu, S.; Himstedt, W.; Maraun, M. Trophic ecology of a tropical aquatic and terrestrial food web: Insights from stable isotopes (15N). J. Trop. Ecol. 2006, 22, 469–476. [Google Scholar] [CrossRef]
- Lewandowski, A.S.; Noss, R.F.; Parsons, D.R. The effectiveness of surrogate taxa for the representation of biodiversity. Conserv. Biol. 2010, 24, 1367–1377. [Google Scholar] [CrossRef]
- Jolley, D.B.; Ditchkoff, S.S.; Sparklin, B.D.; Hanson, L.B.; Mitchell, M.S.; Grand, J.B. Estimate of herpetofauna depredation by a population of wild pigs. J. Mammal. 2010, 91, 519–524. [Google Scholar] [CrossRef] [Green Version]
- Filippi, E.; Luiselli, L. Negative effect of the wild boar (Sus scrofa) on the populations of snakes at a protected mountainous forest in central Italy. Ecol. Mediterr. 2002, 28, 93–98. [Google Scholar] [CrossRef]
- Daltry, J.C.; Lindsay, K.; Lawrence, S.N.; Morton, M.N.; Otto, A.; Thibou, A. Successful reintroduction of the Critically Endangered Antiguan racer Alsophis antiguae to offshore islands in Antigua, West Indies. Int. Zoo Yearb. 2017, 51, 97–106. [Google Scholar] [CrossRef]
- Lillywhite, H.; Martins, M. (Eds.) Islands and Snakes: Isolation and Adaptive Evolution; Oxford University Press: Oxford, UK, 2019. [Google Scholar]
- Shine, R. The ecological impact of invasive cane toads (Bufo marinus) in Australia. Q. Rev. Biol. 2010, 85, 253–291. [Google Scholar] [CrossRef]
- Smokorowski, K.E.; Randall, R.G. Cautions on using the Before-After-Control-Impact design in environmental effects monitoring programs. Facets 2017, 2, 212–232. [Google Scholar] [CrossRef] [Green Version]
- Guareschi, S.; Laini, A.; England, J.; Johns, T.; Winter, M.; Wood, P.J. Invasive species influence macroinvertebrate biomonitoring tools and functional diversity in British rivers. J. Appl. Ecol. 2021, 58, 135–147. [Google Scholar] [CrossRef]
- Médail, F.; Quézel, P. Biodiversity hotspots in the Mediterranean Basin: Setting global conservation priorities. Conserv. Biol. 1999, 13, 1510–1513. [Google Scholar] [CrossRef]
- Migheli, Q.; Balmas, V.; Komoñ-Zelazowska, M.; Scherm, B.; Fiori, S.; Kopchinskiy, A.G.; Kubicek, C.P.; Druzhinina, I.S. Soils of a Mediterranean hot spot of biodiversity and endemism (Sardinia, Tyrrhenian Islands) are inhabited by pan—European, invasive species of Hypocrea/Trichoderma. Environ. Microbiol. 2009, 11, 35–46. [Google Scholar] [CrossRef]
- Ballouard, J.M.; Ferrari, T.; Bonnet, X.; Caron, S.; Maxime, L.; Garnier, G.; Gillet, P.; Ausanneau, M. Snakes of Port-Cros National Park islands: Capture-Mark-Recapture study of Malpolon monspessulanus and Rhinechis scalaris. Sci. Rep. Port-Cros Natl. Park 2016, 30, 23–44. [Google Scholar]
- Bonnet, X.; Naulleau, G.; Shine, R. The dangers of leaving home: Dispersal and mortality in snakes. Biol. Conserv. 1999, 89, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Cheylan, G.; Geoffroy, D. Colonisation des îles d’Hyères (Var, sud de la France) par le sanglier Sus scrofa. Sci. Rep. Port-Cros Natl. Park 2020, 34, 45–56. [Google Scholar]
- Pleguezuelos, J.M.; Fernández-Cardenete, J.R.; Honrubia, S.; Feriche, M.; Villafranca, C. Correlates between morphology, diet and foraging mode in the Ladder Snake Rhinechis scalaris (Schinz, (Schinz, 1822). Contrib. Zool. 2007, 76, 179–186. [Google Scholar] [CrossRef] [Green Version]
- Feriche, M.; Pleguezuelos, J.M.; Santos, X. Reproductive ecology of the Montpellier snake, Malpolon monspessulanus (Colubridae), and comparison with other sympatric colubrids in the Iberian Peninsula. Copeia 2008, 2, 279–285. [Google Scholar] [CrossRef]
- Deso, G.; Bonnet, X.; De Haan, C.; Garnier, G.; Dubos, N.; Ballouard, J.M. Snake overboard! Observations of marine swimming in Malpolon monspessulanus. Herpet. Notes 2021, 14, 593–596. [Google Scholar]
- Cheylan, M. Mise en évidence d’une activité nocturne chez le serpent méditerranéen Elaphe scalaris (Ophidia, Colubridae). Amphib.-Reptil. 1986, 7, 181–186. [Google Scholar] [CrossRef]
- Martinez-Freiria, F.; Lorenzo, M.; Lizana, M. Zamenis scalaris prefers abandoned citrus orchards in Eastern Spain. Ecological insights from a radio-tracking survey. Amphib.-Reptil. 2019, 40, 113–119. [Google Scholar] [CrossRef]
- Suehs, C.M.; Affre, L.; Médail, F. Invasion dynamics of two alien Carpobrotus (Aizoaceae) taxa on a Mediterranean island: I. Genetic diversity and introgression. Heredity 2004, 92, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Hulme, P.E.; Brundu, G.; Camarda, I.; Dalias, P.; Lambdon, P.; Lloret, F.; Medail, F.; Moragues, E.; Suehs, C.; Traveset, A.; et al. Assessing the risks to Mediterranean islands ecosystems from alien plant introductions. In Plant Invasions: Human Perception, Ecological Impacts and Management; Backhuys Publishers: Leiden, The Netherlands, 2008; pp. 39–56. [Google Scholar]
- Capizzi, D.; Baccetti, N.; Sposimo, P. Fifteen years of rat eradication on Italian islands. In Problematic Wildlife; Angelici, F.M., Ed.; Springer: Cham, Germany, 2016; pp. 205–227. [Google Scholar]
- Brécard, D.; De Luigi, C. Fréquentation touristique de Port-Cros et Porquerolles: Les enseignements de la base de données Bount îles. Sci. Rep. Port-Cros Natl. Park 2016, 30, 65–94. [Google Scholar]
- Brown, W.S.; Parker, W.S. A ventral scale clipping system for permanently marking snakes (Reptilia, Serpentes). J. Herpetol. 1976, 10, 247–249. [Google Scholar] [CrossRef]
- López-Calderón, C.; Feriche, M.; Alaminos, E.; Pleguezuelos, J.M. Loss of largest and oldest individuals of the Montpellier snake correlates with recent warming in the southeastern Iberian Peninsula. Cur. Zool. 2017, 63, 607–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, E.P.; Orvos, D.R.; Cairns, J., Jr. Impact assessment using the before-after-control-impact (BACI) model: Concerns and comments. Can. J. Fish. Aquat. Sci. 1993, 50, 627–637. [Google Scholar] [CrossRef]
- Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. Available online: https://www.R-project.org/ (accessed on 21 June 2021).
- Pradel, R. Utilization of capture-mark-recapture for the study of recruitment and population growth rate. Biometrics 1996, 52, 703–709. [Google Scholar] [CrossRef]
- Kendall, W.L.; Nichols, J.D.; Hines, J.E. Estimating temporary emigration using capture recapture data with Pollock’s robust design. Ecology 1997, 78, 563–578. [Google Scholar]
- Arsovski, D.; Olivier, A.; Bonnet, X.; Drilholle, S.; Tomovic, L.; Béchet, A.; Golubovic, A.; Besnard, A. Covariates streamline age-specific early life survival estimates of two chelonian species. J. Zool. 2018, 306, 223–234. [Google Scholar] [CrossRef] [Green Version]
- White, G.C.; Burnham, K.P. Program MARK: Survival estimation from populations of marked animals. Bird Study 1999, 46, 120–138. [Google Scholar] [CrossRef]
- Cruz, F.; Donlan, C.J.; Campbell, K.; Carrion, V. Conservation action in the Galapagos: Feral pig (Sus scrofa) eradication from Santiago Island. Biol. Conserv. 2005, 121, 473–478. [Google Scholar] [CrossRef]
- Hegel, C.G.Z.; Santos, L.R.; Marinho, J.R.; Marini, M.Â. Is the wild pig the real “big bad wolf”? Negative effects of wild pig on Atlantic Forest mammals. Biol. Invasions 2019, 21, 3561–3574. [Google Scholar] [CrossRef]
- Mori, E.; Lazzeri, L.; Ferretti, F.; Gordigiani, L.; Rubolini, D. The wild boar Sus scrofa as a threat to ground--nesting bird species: An artificial nest experiment. J. Zool. 2021, 314, 311–320. [Google Scholar] [CrossRef]
- Reading, C.J.; Luiselli, L.M.; Akani, G.C.; Bonnet, X.; Amori, G.; Ballouard, J.M.; Philippi, E.; Naulleau, G.; Pearson, D.; Rugiero, L. Are snake populations in widespread decline? Biol. Lett. 2010, 6, 777–780. [Google Scholar] [CrossRef] [Green Version]
- Ajtić, R.; Tomović, L.; Sterijovski, B.; Crnobrnja-Isailović, J.; Djordjević, S.; Djurakić, M.; Bonnet, X. Unexpected life history traits in a very dense population of dice snakes. Zool. Anz. 2013, 252, 350–358. [Google Scholar] [CrossRef]
- Folt, B.; Goessling, J.M.; Tucker, A.; Guyer, C.; Hermann, S.; Shelton--Nix, E.; McGowan, C. Contrasting patterns of demography and population viability among gopher tortoise populations in Alabama. J. Wildl. Manag. 2021, 85, 617–630. [Google Scholar] [CrossRef]
- Brischoux, F.; Bonnet, X.; Pinaud, D. Fine scale site fidelity in sea kraits: Implications for conservation. Biodivers. Conserv. 2009, 18, 2473–2481. [Google Scholar] [CrossRef]
- Wilcox, J.T. Implications of predation by wild pigs on native vertebrates: A case study. Calif. Fish Game 2015, 101, 72–77. [Google Scholar]
- Shine, R.; Bonnet, X. Snakes: A new ‘model organism’in ecological research? Trends Ecol. Evol. 2000, 15, 221–222. [Google Scholar] [CrossRef]
- Ward, R.J.; Griffiths, R.A.; Wilkinson, J.W.; Cornish, N. Optimising monitoring efforts for secretive snakes: A comparison of occupancy and N-mixture models for assessment of population status. Sci. Rep. 2017, 7, 18074. [Google Scholar] [CrossRef] [Green Version]
- Kaminski, G.; Brandt, S.; Baubet, E.; Baudoin, C. Life-history patterns in female wild boars (Sus scrofa): Mother–daughter postweaning associations. Can. J. Zool. 2005, 83, 474–480. [Google Scholar] [CrossRef] [Green Version]
- Lombardo, C.A.; Faulkner, K.R. Eradication of feral pigs (Sus scrofa) from Santa Rosa Island, Channel Islands National Park, California. In Proceedings of the Fifth California Islands Symposium; Santa Barbara Museum of Natural History: Santa Barbara, CA, USA, 2000; pp. 300–306. [Google Scholar]
- Servanty, S.; Gaillard, J.M.; Ronchi, F.; Focardi, S.; Baubet, E.; Gimenez, O. Influence of harvesting pressure on demographic tactics: Implications for wildlife management. J. Appl. Ecol. 2011, 48, 835–843. [Google Scholar] [CrossRef]
- Keuling, O.; Baubet, E.; Duscher, A.; Ebert, C.; Fischer, C.; Monaco, A.; Podgórski, T.; Prevot, C.; Ronnenberg, K.; Sodeikat, G.; et al. Mortality rates of wild boar Sus scrofa L. in central Europe. Eur. J. Wildl. Res. 2013, 59, 805–814. [Google Scholar] [CrossRef]
- Vajas, P.; Calenge, C.; Richard, E.; Fattebert, J.; Rousset, C.; Saïd, S.; Baubet, E. Many, large and early: Hunting pressure on wild boar relates to simple metrics of hunting effort. Sci. Total Environ. 2020, 698, 134251. [Google Scholar] [CrossRef] [PubMed]
- Donlan, C.J.; Howald, G.R.; Tershy, B.R.; Croll, D.A. Evaluating alternative rodenticides for island conservation: Roof rat eradication from the San Jorge Islands, Mexico. Biol. Conserv. 2003, 114, 29–34. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Model | Deviance | Num. Par | AICc | Delta AICc | AICc Weights |
|---|---|---|---|---|---|
| ϕ (sp) γ (sp + BACI cst PQL) p(island + sp + year) | 145.03 | 46 | 3059.25 | 0.00 | 0.54 |
| ϕ (sp) γ (sp + BACI) p(island + sp + year) | 144.99 | 47 | 3061.57 | 2.32 | 0.17 |
| ϕ (sp) γ (island + sp + BA) p(island + sp + year) | 145.59 | 47 | 3062.17 | 2.92 | 0.13 |
| ϕ (sp) γ (island + sp + BACI) p(island + sp + year) | 144.38 | 48 | 3063.32 | 4.07 | 0.07 |
| ϕ (island + sp) γ (island + sp + BACI) p(island + sp + year) | 144.34 | 49 | 3065.66 | 6.41 | 0.02 |
| ϕ (sp + BA) γ (island + sp + BACI) p(island + sp + year) | 144.37 | 49 | 3065.69 | 6.44 | 0.02 |
| ϕ (sp + BACI cst PQL) γ (island + sp + BACI) p(island + sp + year) | 144.38 | 49 | 3065.70 | 6.45 | 0.02 |
| ϕ (island + sp + BA) γ (island + sp + BA) p(island + sp + year) | 145.53 | 49 | 3066.85 | 7.60 | 0.01 |
| ϕ (island + sp + BA) γ (island + sp + BACI) p(island + sp + year) | 144.33 | 50 | 3068.04 | 8.79 | 0.01 |
| ϕ (sp + BACI) γ (island + sp + BACI) p(island + sp + year) | 144.37 | 50 | 3068.08 | 8.83 | 0.01 |
| S(sp) γ (sp + BA) p = c(island + sp + year) | 154.47 | 46 | 3068.70 | 9.45 | 0.00 |
| ϕ (island + sp + BACI) γ (island + sp + BACI) p(island + sp + year) | 144.26 | 51 | 3070.36 | 11.11 | 0.00 |
| ϕ (island + sp + BACI) γ (island + sp + BACI) p(island + year) | 149.69 | 50 | 3073.39 | 14.15 | 0.00 |
| ϕ (island + sp + BACI) γ (island + sp + BACI) p(sp + year) | 154.37 | 50 | 3078.08 | 18.83 | 0.00 |
| ϕ (island + sp + BACI) γ (island + sp + BACI) p(island + sp) | 180.85 | 44 | 3090.38 | 31.13 | 0.00 |
| Species | Factors | Estimate | Std. Error | Z Value | p |
|---|---|---|---|---|---|
| Zamenis scalaris + Malpolon monspessulanus | Time | 0.542270 | 0.134336 | 4.037 | <0.001 |
| Island | 0.007439 | 0.137389 | 0.054 | 0.9568 | |
| Time*island | −0.486746 | 0.201369 | −2.417 | <0.05 | |
| Zamenis scalaris | Time | 0.04709 | 0.23341 | 0.202 | 0.840 |
| Island | 0.09590 | 0.22207 | 0.432 | 0.666 | |
| Time*island | −0.22933 | 0.34120 | −0.672 | 0.502 | |
| Malpolon monspessulanus | Time | 0.78568 | 0.14436 | 5.443 | <0.001 |
| Island | −0.04636 | 0.15788 | −0.294 | 0.769 | |
| Time*island | −0.56400 | 0.22182 | −2.543 | <0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ballouard, J.-M.; Kauffman, C.; Besnard, A.; Ausanneau, M.; Amiguet, M.; Billy, G.; Caron, S.; Fosseries, G.; Ferrari, T.; Mariani, V.; et al. Recent Invaders in Small Mediterranean Islands: Wild Boars Impact Snakes in Port-Cros National Park. Diversity 2021, 13, 498. https://0-doi-org.brum.beds.ac.uk/10.3390/d13100498
Ballouard J-M, Kauffman C, Besnard A, Ausanneau M, Amiguet M, Billy G, Caron S, Fosseries G, Ferrari T, Mariani V, et al. Recent Invaders in Small Mediterranean Islands: Wild Boars Impact Snakes in Port-Cros National Park. Diversity. 2021; 13(10):498. https://0-doi-org.brum.beds.ac.uk/10.3390/d13100498
Chicago/Turabian StyleBallouard, Jean-Marie, Cécile Kauffman, Aurélien Besnard, Mathieu Ausanneau, Manon Amiguet, Gopal Billy, Sébastien Caron, Guillaume Fosseries, Thomas Ferrari, Vincent Mariani, and et al. 2021. "Recent Invaders in Small Mediterranean Islands: Wild Boars Impact Snakes in Port-Cros National Park" Diversity 13, no. 10: 498. https://0-doi-org.brum.beds.ac.uk/10.3390/d13100498