
Diversity of Flower Visiting Beetles at Higher Elevations on the Yulong Snow Mountain (Yunnan, China)


1
College of Plant Protection, Yunnan Agricultural University, Kunming 650201, China
2
Kunming Natural History Museum of Zoology, Kunming Institute of Zoology, Chinese Academy of Sciences, Kunming 650201, China
3
Key Laboratory for Plant Diversity and Biogeography of East Asia, Kunming Institute of Botany, Chinese Academy of Sciences, Kunming 650201, China
4
Lijiang Forest Biodiversity National Observation and Research Station, Lijiang 674100, China
*
Authors to whom correspondence should be addressed.
Diversity 2021, 13(11), 604; https://0-doi-org.brum.beds.ac.uk/10.3390/d13110604
Submission received: 17 October 2021
/
Revised: 16 November 2021
/
Accepted: 19 November 2021
/
Published: 21 November 2021
(This article belongs to the Section Animal Diversity)
Abstract
:Background: Flowers are one of the important microhabitats promoting beetle diversity, but little is known about variation in the diversity of these insects at higher elevations. We do not know how divergent habitats influence the distribution of beetles among montane flora. Methods: We sampled beetles systematically in angiosperm flowers at 12 sites at two elevations (2700 m and 3200 m) and in two habitats (meadows and forests) for two consecutive years (2018 and 2019) on the Yulong Snow Mountain in Yunnan, southwestern China. Beetle diversity among sites were compared. Their interactions with flowers of identified plant species were analyzed using bipartite networks approach. Results: We collected 153 species of beetles from 90 plant species recording 3391 interactions. While plant species richness was lower at the higher, 3200 m elevation regardless of habitat type, beetle species richness was not significantly different among sites. Plant-beetle interaction networks were strongly modular and specialized. The structure of networks showed greater differences between elevations than between habitats. The turnover of networks was determined by species composition showing a weak influence by interaction rewiring. Conclusion: Our study showed a high diversity of beetles in flowers at higher elevations within this mountain complex. The role of beetles in plant–insect interactions within some sections of temperate, montane sites appear to be underestimated and warrant further study.
1. Introduction
It is estimated that the majority of flowering plant species (87.5%) are pollinated by invertebrates and vertebrates [1]. The Coleoptera include over 400,000 species, comprising the largest order of insects [2]. Therefore, beetles constitute almost 40% of described insects and 25% of all known species in Kingdom Animalia [3]. Beetles are the second most diverse group of pollinators after Lepidoptera, as an expected 77,000 species are found visiting flowers as many adults are florivores and/or eat pollen [4,5].
Fossil evidence indicates that beetles were part of the guild that visited flowers during the Cretaceous. Some species that belong to modern lineages remain essential pollinators today. They are especially important pollinators of the basal eumagnoliids with their chamber-like flowers including members of Magnoliaceae, Annonaceae and Eupomatiaceae. However, plant species pollinated primarily by beetles have evolved repeatedly in unrelated families of eudicotyledons and monocotyledons producing flowers showing great variation in pigmentation and discernible scents [6]. Most beetle-pollinated flowers are salver-bowl shaped with easy access to pollen [6,7,8,9,10,11]. While beetles often devour portions of a flower, some may be able to disperse pollen grains sufficient distances to effect cross-pollination.
The diversity of flower visiting beetles, both florivores and true pollinators, remains underestimated, although they are often the second most common group of flower visitors in tropical ecosystems [5,12,13,14,15]. For example, in a lowland dipterocarp forest in Malaysia, Sakai et al. (1999) [11] found that beetles accounted for 74% of visitors during the flowering period of a single emergent tree. In an Australian dry rainforest, 22% of plants were found to be beetle-pollinated [16]. Momose et al. (1998) [17] similarly found that 20% of the 270 plant species in an Asian dipterocarp forest were beetle-pollinated, representing the second most abundant pollination system after social bees in that forest. In an Australian tropical rainforest, a detailed analysis of the beetle fauna revealed that flowers were utilized by approximately 41% of the species [18]. Therefore, it has been suggested that beetle pollination in tropical forests represents a more important contribution to the reproduction of indigenous angiosperms compared to those plant species comprising temperate forests [19].
In fact, the effectiveness of beetles as pollinators is still poorly understood in most cases, as there are few studies on how these insects pick up and transfer pollen. However, flowers are especially important microhabitats and supported a disproportionately large number of beetle species [20], while beetle densities (per unit biomass of microhabitat) were 10–10,000 times greater on flowers than on adjacent foliage [18,20]. Beetle visiting flowers may not contribute to pollination, however, their visitation may indirectly change the nectar and/or pollen attributes and consequently influence plant–pollinator interaction.
The diversity of flower visiting beetles in temperate, montane regions has received less attention. Mountains support an estimated one-third of all terrestrial species diversity [21,22,23] and make up half of all 34 regions classified currently as global biodiversity hotspots [24]. However, most studies of flower-visiting insects in alpine/subalpine regions are focused on plant–pollinator interactions at species or community levels [25,26,27,28,29,30,31,32,33]. In these studies, beetles are usually reduced to minor roles within pollinator assemblages [34,35]. Lefebvre et al. (2018) [31] studied the flower-visitor variation along elevation gradients in communities in the Alps. Results showed that both the abundance and richness of anthophilous beetles decreased as elevation increased above 2000 m. Additionally, Zhao et al. (2016, 2019) [32,33] studied flower-visiting interactions in communities in the Himalaya–Hengduan Mountains. They found that only 7.3% of putative pollinator species were beetles.
Beetle-pollination has also been shown to be strongly dependent on plant phylogeny showing a strong phylogenetic signal and/or phylogenetic conservatism [36,37]. With the exception of the beetle-pollinated basal lineages of angiosperms [6], recent studies showed that flowers produced in several families of eudicotyledons including Asteraceae and Ranunculaceae were hotspots for beetle visitations [37,38]. Floral presentation in these plant families is usually indicative of a generalized pollination system visited by a wide variety of insects representing several orders and many families. However, although these two families are common in montane regions of the Northern Hemisphere, the diversity and density of beetles on their flowers have not been investigated broadly at high elevations, because these are the elevations in which pollination systems tend to be dominated by eusocial bees in the genera Apis and Bombus [33]. Therefore, we still do not know whether the divergence of habitats at higher elevations into forests and meadows effect flower visiting beetle diversity and how they interact with the host plant species at the community level forming networks.
Previous studies on plant–pollinator interactions are based on the identification of insects collected in the flowers, and their contact with floral reproductive organs as they foraged for floral resources, i.e., nectar and pollen [33,39,40]. Therefore, most of these studies showed a bias towards the large-bodied beetles, often ignoring smaller specimens found in the flowers. To fully explore flower visiting beetle diversity we propose a field survey. In this study, we examined how the abundance and species richness of flower visiting beetles varied at different elevation gradients, and how abundance and species richness of flower visiting beetles varied in flowers based on vegetation types restricted to different habitats. We also used a bipartite network analysis to characterize the properties of flower–beetle interactions. We address the following questions: (1) Is beetle diversity in flowers influenced by elevation gradients and habitat types? (2) What are the network properties of flower visiting beetles at two high elevations? (3) Do the properties of flower–beetle interaction networks vary among different sites on the same mountain?
2. Material and Methods
2.1. Study Sites and Plot Design
All fieldwork was conducted on the Yulong Snow Mountain in the Hengduan mountain range through southwestern Yunnan Province, China. Most of the sites were located at the Lijiang Forest Biodiversity National Observation and Research Station operated by the Kunming Institute of Botany (KIB), Chinese Academy of Sciences. We selected two elevations (2700 m and 3200 m), and each elevation was subdivided into three sites. The linear distance between two sites at the same elevation gradient was about 2 km. The lower elevation locations included: Yuhu (YH), 27°01′41.61″ N, 100°12′52.27″ E, 2864 m; Yushuizhai (YS), 26°59′58.05″ N, 100°11′57.44″ E, 2706 m; Juxianzhuang (JX), 26°59′22.13″ N, 100°11′50.94″ E, 2635 m. The higher elevation locations included: Shuiku (SK), 27°00′07.20″ N, 100°10′54.72″ E, 3238 m; Luxia (LX), 26°59′21.73″ N, 100°10′24.13″ E, 3233 m; Lubian (LB), 26°58′42.65″ N, 100°10′45.44″ E, 3180 m. See Table S1 for sampling site details and sampling times. The meadows YS and SK are the same as described in Zhao et al. (2016, 2019) [32,33]. The sites at high elevation are the same as in Xu et al. (2021) [41] and Liang et al. (2021) [42]. These previous publications provided us with the list of flowering plant species used in this study with herbarium specimens deposited previously in KIB. Weather records and seasonal climate follow Xu et al. (2021) [41] and Liang et al. (2021) [42]. Based on previous publications [32,33,41,42], flower visitors collected previously at our sites were dominated by bumblebees, honeybees (Apis cerana), and solitary bees. While beetles were components of minor pollinator groups following the standard pollinator survey technique by observing and recording whenever an insect was observed contacting the stamen(s) and/or the stigma(s) in each flower.
At each site, we setup two paralleled transects (plots). One was the meadow transect and the second was the forest transect following Xu et al. (2021) [41]. Each meadow transect was 50 m in length following a linear route, and 3 m in width. Each forest transect was 100 m in length following a linear route, and 3 m in width. We tried to set the meadow transect in the meadow’s center to avoid the margin effect. The linear distance between the meadow transect and the forest transect was about 100 m. The plant assemblage of flowering herbs in the meadow and forest understory at high elevation followed Xu et al. (2021) [41], but we added the flowers of native shrub species in this study. Conifers dominated the forest vegetation at the low sites. At the higher elevation the forest was a Pinus-Quercus assembly.
2.2. Data Collection
Sampling took place between the 18 July–15 October in 2018 and 21 May–4 October in 2019. We conducted one survey per month, with 20–31 days intervals. Each site we sampled 10 times over two years with four samplings in 2018 followed by six samplings in 2019 (Table S1). Sampling was restricted to sunny or partly cloudy days. We could not sample on rainy days. Each sampling round consisted of walking at a slow pace for a 15–60 min bout from 9:00–17:00. The average collection time was 30 min in each transect depending on the abundance of flowering plants. The minimum collecting time along a transect was 15 min if no insect was found and collected. Specifically, we recorded no beetles present for seven sampling periods five times at our high elevation sites in 2019 during the last sampling in October, which is the end of the flowering season (Table S1).
We adopted a selected beetle sampling strategy collecting only specimens found in the flowers while avoiding beetles on leaves, stems or on the ground. We did not collect insects that did not belong to the order Coleoptera. We used a beetle specific collection method following Wardhaugh (2013) [43] by gently beating each flower or inflorescence 3 times with a wooden stick while holding an insect net under said flower or inflorescence. Beetles that fell into the net were retrieved, and each specimen was stored in a small bottled containing 100% ethanol. The plant species on which the beetle was collected was recorded. Bottled beetles were brought to the laboratory for identification.
Voucher specimens was either identified by the first author or sent to other entomologists in China (see Acknowledgement). Many collections were novel and could not be identified to species. When we could not identify a specimen to species we assigned it to a morphotype. Taxonomic treatment to family followed Zheng and Gui (1999) [44]. Voucher specimens were pinned and deposited in the Kunming Natural History Museum of Zoology, Kunming Institute of Zoology (KIZ), Chinese Academy of Sciences. Plant specimens were identified by the second author in the field or from pressed vouchers that were also deposited in KIZ.
The completeness of sampling was estimated by the Hill numbers method (species richness, Shannon diversity and Simpson diversity). Using an iNEXT package on R [45], we estimated the species coverage by specimen numbers for different elevations and habitats.
2.3. Network Construction and Analysis
Flower-visitor networks were constructed using the package bipartite [46] in R (version 3.6.2) [47]. As we focused on the influence of elevation and habitat type (forest and meadow) on the network properties and turnover, we combined all the sampling data of each survey accumulated over two years to fully represent beetle diversity for each site. We did not consider the effects of season and year on plant-beetle interactions. We treated meadow and forest transects as separate sites at each locality. To understand the structure of the plant–flower visiting beetle network, we built a total network combining all data to provide a brief view of the interactive topology between plants and beetles. Then, we built 12 networks for each site (transect) to do the comparison.
Network and species-level indices commonly used in plant–insect networks, considered to provide ecologically relevant information about the structure and function of these networks, were calculated using bipartite package. In addition to the basic information of networks including network size, the number of interactions and the number of species at high and low levels (beetles and plants), the following network-level indices were calculated for the total network and each plant-beetle network. We used the network level function to calculate the following indices.
- H2′: Network generalization describes the degree of specialization among plants and pollinators within the network [48] and ranges between 0 (extreme generalization) and 1 (extreme specialization).
- Weighted connectance refers to the realized proportion of possible links weighted by network size.
- Nestedness is based on overlap and decreasing fill (NODF): the extent to which specialists interact with a subset of species that also interact with generalists.
- Niche overlap of beetles refers to mean similarity in interaction patterns between flower visiting beetles.
- Extinction slope of beetles refers to the simulated secondary loss of beetle species following the extinctions of plant species.
- Robustness of beetles refers to the “fragility” of beetles to losses in the other level (plant).
- Functional complementarity of beetles refers to the extent of the sharing of interactions between beetles.
Bees and other anthophilous insects belonging to other orders (Lepidoptera and Diptera) are regarded as the most common pollinators of indigenous plant species at our sites [33,42]. Therefore, we assumed that the reproductive dependency of plants on beetles was much lower than the dependency of foraging beetles on the plants they visited. Consequently, we used a beetle-centered sampling approach to build the interacting networks. This explains why we only calculated niche overlap, extinction slope, robustness and functional complementarity for beetles, but not for plants.
We also calculated modularity and species roles for the total network. Modularity Q described the degree of compartmentalization in a network and ranged from 0 (the network did not have more links within modules than expected by chance) to a maximum value of 1 (all links are within modules) [49,50]. We calculated the modularity metric Q using the DIRTLPAwb + algorithm, which searches for the maximum modularity possible [50]. As this is an optimization algorithm, the maximum modularity may vary among runs, so we repeated them five times and accepted the highest Q value. Species roles for beetles and plants of the total network were calculated following Olesen et al. (2007) [49]. The within-module degree (z) is a measure of the number of connections a species had within its own module relative to other species in that module. In contrast, the among-module connectivity (c) informed us about how well a given species is connected to species that belong to other modules [49]. According to their c and z-values, species were classified as: peripherals (low values of both c and z), connectors (high c and low z values), module hubs (high z and low c values), or network hubs (high values of both c and z).
2.4. Network Dissimilarity (β-Diversity)
We followed Poisot et al. (2012) [51] to examine the turnover of networks among elevations and habitat types. The following indices were calculated, βS, dissimilarity in the species composition of communities; βOS, dissimilarity of interactions established between species common to both sets of interactions; βWN, dissimilarity of interactions and βST, dissimilarity of interaction due to species turnover. These indices were calculated using the bipartite package ver. 2.14 in R [46,52] as quantitative Jaccard dissimilarities between networks.
2.5. Statistical Analyses
We used a series of two-way ANOVAs to compare the mean differences of network indices between elevation and habitat types. The elevation (low or high) and habitat type (meadow or forest) were analyzed as two independent variables (factors). We used Pearson product–moment correlation coefficient to test the correlations among plant species richness for each angiosperm family, beetle species richness and beetle species abundance. As network properties varied mainly between elevations rather than between habitats, we compared network turnover (β-diversity) between elevation and within elevation (between habitat types) using Mann–Whitney rank sum tests.
3. Results
3.1. Species Identification and Community Composition
A total of 3391 flower-visiting beetles were collected at our 12 sites over two years. The collection represented 153 morpho-species subdivided into 24 different families (Table 1 and Table S2, Figure 1, Figure 2 and Figure 3). These specimens were caught foraging on the flower of 90 different species representing 30 families in 22 angiosperm orders (Table 2 and Table S3, Figure 1). The species richness of beetles varied with family. Specimens identified as members of the Chrysomelidae accounted for 27.45% (42 species) of the catch. The Nitidulidae made up 22.22% (34 species), the Curculionidae with 6.54% (10 species), and the Coccinellidae with 6.54% (10 species). In contrast, the abundance of beetle specimens was related to species richness (r = 0.782, p < 0.001), raising the Nitidulidae to 48.86% (1657 specimens), the Chrysomelidae to 28.16% (955), the Crioceridae to 5.1% (173), and the Cantharidae to 4.16% (141) as the major families. The dominant beetle species were Nonarthra variabilis Baly (706 specimens; Chrysomelidae) and Lamiogethes sp.1 (672 specimens; Nitidulidae).
Species in the Asteraceae (23.33%, 21 species), Ericaceae (2 species), Rosaceae (5 species), Apiceae (3 species) and Ranuculaceae (9 species, Table 2) were the dominant plant families harboring the highest beetle diversity in their individual flowers and/or compound inflorescences (involucre). We found that 21 species within the Asteraceae were visited by 57 species of beetles representing 13 families. However, while two plant families, Caprifoliaceae and Celastraceae, were represented by only one species each (Dipsacus asper Wallich ex Candolle and Parnassia wightiana Wall. ex Wight et Arn.), respectively, both showed the highest beetle specimen abundance. Additionally, the Hypericaceae represented by two species in the genus Hyperium were visited by 14 beetle species representing 307 specimens. At the species level, Rhododendron decorum Franch. (Ericaceae) was visited by the highest number of beetle species (36), followed by Dipsacus asper Wallich ex Candolle (18). In addition, we collected and identified 17 beetle species from flowers of Physospermopsis delavayi (Franch.) Wolff (Apicaceae) and 16 from Pyracantha fortuneana (Maxim.) Li (Rosaceae).
The species coverage of beetles at all sites was > 90%, but species diversity was at a middle level (Table S4 and Figures S1, S2). The number of plant species for each family was highly correlated with the number of beetle species per plant family (r = 0.758, p < 0.001), but plant species per family showed no correlation with the sheer number of beetle specimens removed from their flowers (r = 0.351, p > 0.05).
3.2. Network Structure of Plant-Flower Visiting Beetles
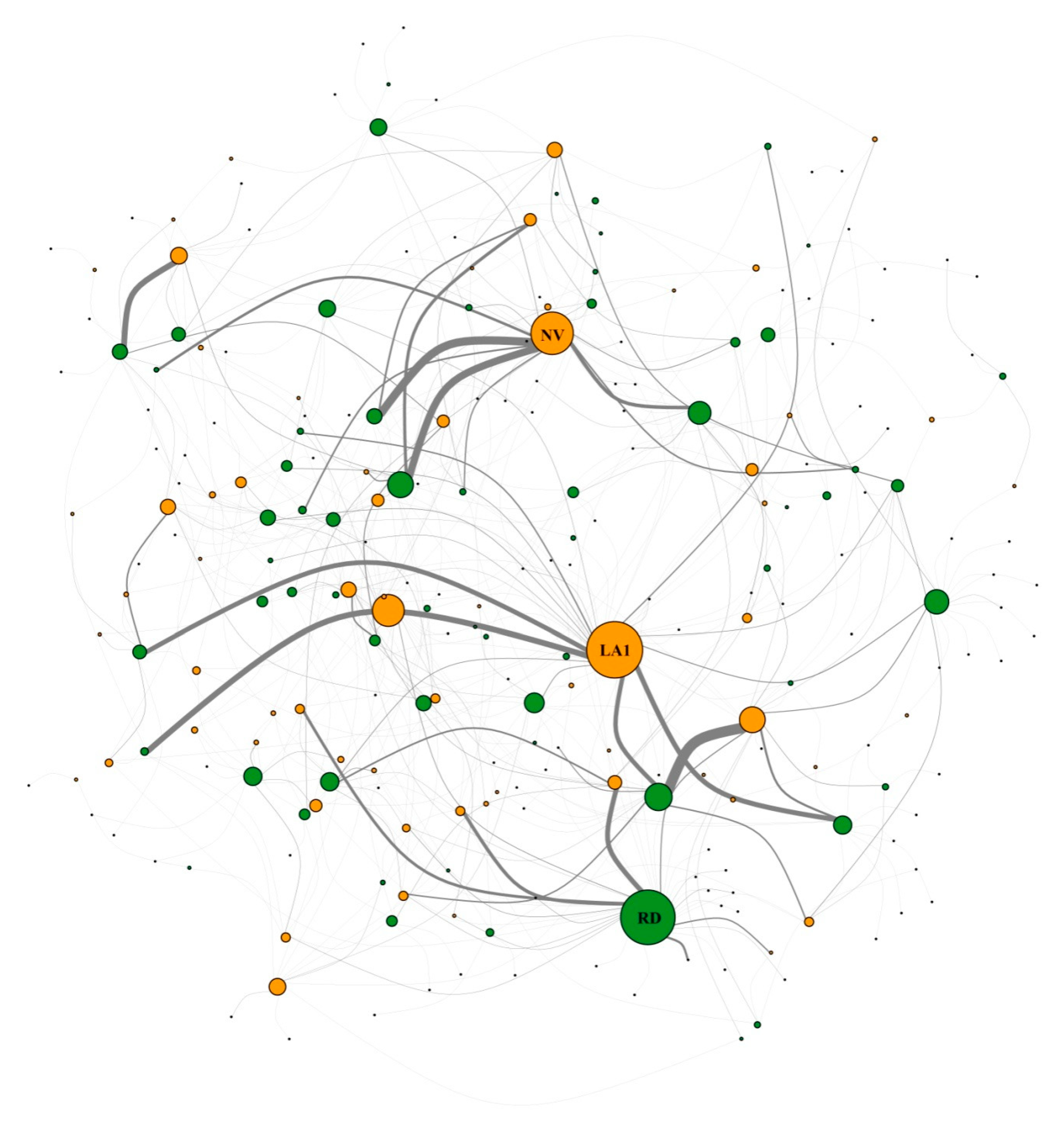
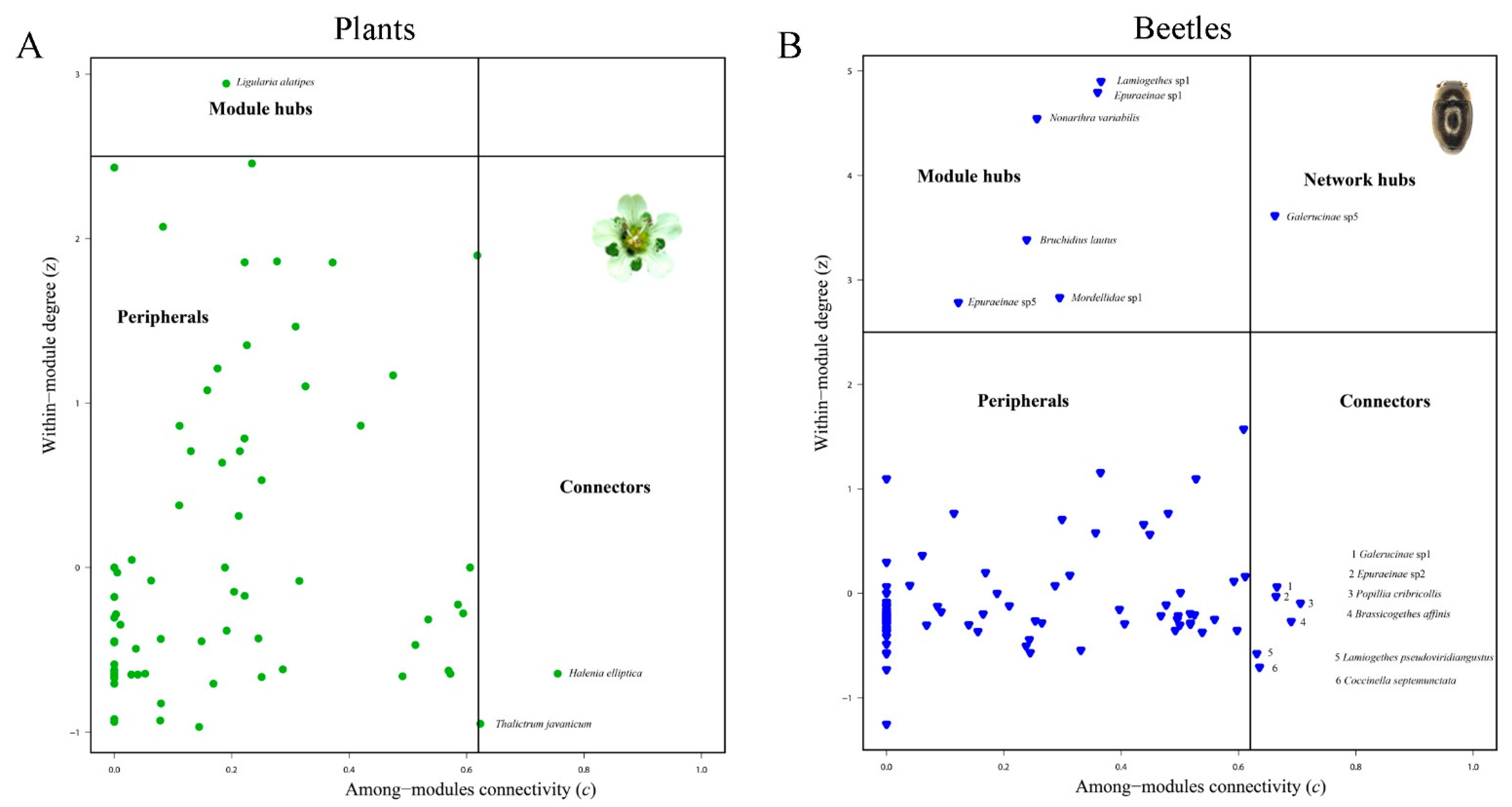
The total network showed strong modularity (Q = 0.632) with a Z-score of 138.29, which was significantly different from the null model (p = 0; Figure 2). There were 13 modules detected for the total network. The total network also showed a nested structure. The NODF was 11.9651 with a Z-score of -22.38, which was significantly different from the null model (p = 0). One beetle species Galerucinae sp.5 (Chrysomelidae) was detected as the network hub. Six additional beetle species were module hubs, and six species were connectors. For plants, no network hub was detected. Ligularia alatipes Hand.-Mazz. (Asteraceae), Halenia elliptica D. Don (Gentianaceae) and Thalictrum javanicum Bl. (Ranunculaceae; Figure 4) were detected as module hubs.
Network size of 12 plant–beetle networks varied from 30 to 46 with the network size at the low elevation being higher than that at the higher elevation (F = 8.329, p = 0.02; Figure 3). Similarly, the number of interactions were also slightly higher at the lower elevation (F = 5.472, p = 0.047), but no habitat difference was shown (F = 4.704, p = 0.062). The interaction effects between elevation and habitat were also significant for the number of interactions (F = 9.991, p = 0.013, Table 3). The number of beetle species did not show significant differences between elevations and between the two habitat types (p > 0.05). The numbers of plant species with flowers and/or inflorescences inhabited by beetles were lower at the higher elevation sites compared with the lower sites (F = 36. 698, p < 0.001; Table 3).
For the network structures, we detected a significant effect of elevation on weighted connectance, H2, niche overlap of beetles, and functional complementarity of beetles. The specialization of interacting (H2) was higher at lower elevation (F = 7.191, p < 0.05), and there was also a significant interacting effect for elevations and habitat types (p < 0.05), but no difference among habitat types. Niche overlap of beetle species showed differences among both elevations (F = 100. 330, p < 0.001) and habitat types (F = 34.279, p < 0.001; Table 3).
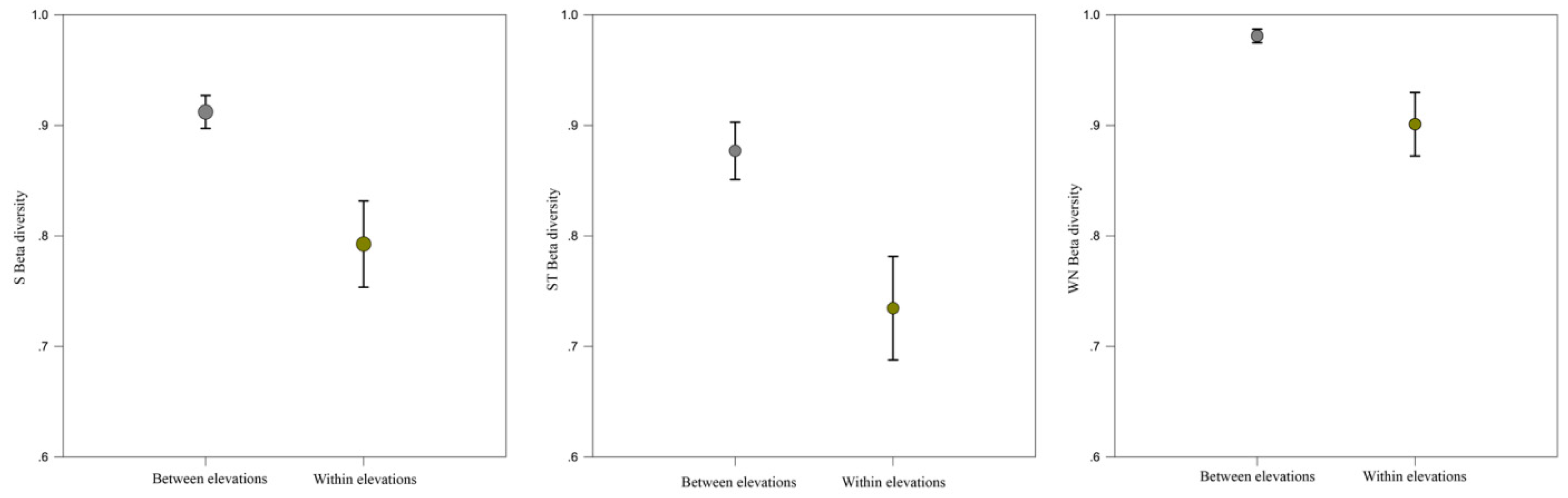
The dissimilarity of species composition (βS) among different sites were high in general (all values > 0.86), except for three forest sites at higher elevation (0.34–0.39; Table S5). Consequently, dissimilarity of interactions established between species common to sites (βOS) was extremely low due to the lower levels of species sharing. Network turnover among sites was determined by species turnover (Table S5). Network turnover between elevation was always higher than within elevation (among habitat types; Figure 5).
4. Discussion
4.1. Comparative Diversity of Flower Visiting Beetles in Sub-Alpine Versus Tropical Versus Mediterranean Communities
In this study, a high diversity of flower-visiting beetles are associated with plants of sub-alpine communities at previously studied sites in the Hengduan Mountain. Despite increased elevation flower-visiting beetle diversity was high and comparable to beetle pollinator diversity proposed previously for temperate, montane ecosystems in the Mediterranean babitats [36,37]. We found 153 species of beetles visiting 90 plant species in our 12 sites. The previous plant–pollinator interaction study by Zhao et al. (2019) [33] on Yulong Snow Mountain, based on multiple samplings in one year, found 26 beetle species accounting for only 7.3% of the guild of pollinator species. However, they also found that most of the flower visiting beetles carried deposits of pollen grains of their host flowers on their bodies. Our findings (above) dramatically increase the biodiversity of beetles on flowers on the same mountain and at some of the same sites used by Zhao et al. (2019) [33]. We presume that the number of beetle species that act as pollen vectors (potential pollinators) will increase when palynological stains and microscopy are employed [9].
Evidence here suggests that flowers of plant species restricted to higher elevations may actually offer a broad range niches producing beetle diversity hotspots that may parallel the floral niches restricted to lowland tropical forests. Wardhaugh (2013, 2015) [4,43] and Wardhaugh et al. (2012, 2015) [18,20] found that flowers offer an important microhabitat encouraging beetle diversity in Australian tropical forest. A primary difference in our study is that the majority of flowers preferred by our beetles belong to herbaceous species excluding shrubs in the Ericaceae and other plant families. This is significant, as Yulong Snow Mountain meadows are dominated by wind-pollinated grasses and its forests are dominated by wind-pollinated gymnosperms and Quercus. The reproductive organs of these plants lack nectar and colorful scented petals where beetles may conceal themselves while they forage and mate.
4.2. Coevolution or Foraging Opportunism of Flower Visiting Beetles and Plant and Plant Lineages?
Flower visiting/pollinating beetle species and the flowers of the plant species, with which they interact, may show a strong phylogenetic conservatism at both a macro-evolutionary scale [6] and at the community level [36]. A particularly strong co-phylogenetic signal between beetles and the native Asteraceae was detected in this study and within the plant–pollinator network in a coastal thermo-Mediterranean sclerophyllous community [38]. Using flower visitation by nitidulid beetles (Nitidulidae) for 251 plant species from Mediterranean montane habitats of southeastern Spain, Herrera and Otero (2021) [37] found that flower visitation by nitidulids was related significantly to plant phylogeny. Specifically, these beetles clustered on flowers belonging to families in the Ranuculales, Malvales, Rosales and Asterales. However, these Mediterranean nitidulids were remarkably absent on flowers of plants classified in the Fabales and Lamiales. As in the work of Herrera and Otero (2021) [37] our beetle collections also showed that their greatest diversity on Yulong Snow Mountain was found on or in flowers belonging to families in the Asterales (Asteraceae), Ranuculales (Ranuculaceae) and Rosales (Rosaceae), suggesting a potential co-evolutionary relationship between flower visiting beetles and these plant lineages.
Of course, there are important differences. First, our two Hypericum species (Hypericaceae; Malphigiales) attracted 12 beetle species. Their dish-shaped, yellow flowers are multi-staminate (polyandrous), and this trait is known to attract some beetles [6,7,8]. Second, in contrast, this study showed that many small beetles were found in flowers of the Fabaceae, Lamiaceae, Ericaceae and Campanulaceae. Previous studies indicated that a combination of pollen features, including scent biochemistry and nutrients in the cytoplasm of grains, may drive beetle visitation to flowers belonging to atypical, angiosperm lineages [37,53,54]. The flowers in these four families have tubular, funnel-bell shaped corollas concealing nectar. This is an additional reward for pollen-eating beetles visiting some angiosperms [6,9] and we did observe these insects crawling towards or congregating at the bases of floral tubes and bells. Some beetles may be nectar robbers. Rhododendron decorum Franch. secretes high volumes of nectar at the base of its ovary and is pollinated by large Bombus species [55], but it was also visited by 36 beetles species, feeding on nectar, in this study.
4.3. Effect of Elevation and Habitat on Beetle Diversity on Montane Flora
As discussed above, the diversity of flower visiting beetles was strongly associated with specific plant species. We did not detect either a strong elevation or habitat type effect on beetle diversity and their interaction with plants. The modularity of the plant–beetle network appears to depend more on preferred, resident, plant species. Herrera and Otero (2021) [37] also found that environmental factors (habitat type), elevation and/or macroscopic floral features (perianth type, floral color, and flower massing on stems) did not explain nitidulid visitation after statistically accounting for the effect of plant phylogeny. The turnover of networks was more dependent on changes to beetle composition at different site. We are reminded here, that our study compared habitats at only two elevations both with the highest plant diversities on Yulong Snow Mountain. On this mountain, complex topology of mountain building and vegetation turnover made the biodiversity variation along elevation did not following the linear decreasing with the increasing of elevation [32,33]. Zhao et al. (2016) [32] found a correlation between decreasing pollinator diversity as elevation increased, but this trend did not apply to plant diversity. Therefore, if in the future, when sampling sites incorporate floras above the tree line (>3900 m) we may expect to find a decrease or a middle domain effect in the diversity of flower visiting species of beetles.
4.4. Uncertain Roles of Flower Visiting Beetles
One of the limits of this study is that we did not discriminate among the many roles different beetles may play in the same flowers as pollinators, or florivores, or pollen and/or nectar thieves, or seed parasites, etc. In fact, one beetle species may play different roles visiting flowers of different species in the same network. We preserved our insect specimens in 100% alcohol in case we needed to do DNA extractions for species identification. This mode of preservation washed pollen off bodies. We do not have observations of which beetles foraged on pollen or nectar while contacting receptive stigmas. Therefore, the role of each beetle species as potential pollinators of any of the plant species at either elevation remains unknown. However, previous studies by other authors indicate that some beetle families (e.g., Nitidulidae) are greater contributors to the pollination of their host flowers compared to species in other families [37].
We understand that a large proportion of the beetle species we collected may play no role in the pollination of the flowers in which they were collected. Some florivorous beetles, including the one with a large number, Nonarthra variabilis Baly (Chrysmelidae), were found to feed heavily on the shiny staminodes of Parnassia wightiana Wall. ex Wight et Arn.. Nonarthra variabilis is probably attracted to the nectar-mimicking staminodes in this flower, but we never saw them eating the ovaries [56], and Chen et al. unpublished data]. It’s also unlikely that these tiny beetles pollinate the bilaterally symmetrical and complex flowers in the Yulong species of in Fabaceae and Lamiaceae. They are probably nectar/pollen robbers, as they were not observed eating floral organs. However, in a study on microbial communities in flower nectar, de Vega et al. (2021) [57] found that plants visited by beetles supported the highest richness and phylogenetic diversity of yeasts, and bacteria communities compared to flowers visited by pollinators like bees. This suggests that the inhabiting of these tiny beetles in flowers may change the nectar attributes, and consequently influence interactions between flowers and their legitimate pollinators.
5. Conclusions
In this study, our specific collections showed a surprisingly high diversity of beetles in flowers at two relatively high elevations. As consistent with previous publications, we found that flower visiting beetles are habitat specific and their diversity on flowers appears to be constrained, at least in part, by plant phylogeny. Subsequently, the beetle–flower interaction networks were highly modular, and network variation among different sites was determined by species composition turnover. We must also add that previous research indicates that beetle flower visitors are also sensitive to land use change. Millard et al. (2021) [58] found a decrease in beetle abundance in the agricultural landscape relative to the primary vegetation baseline. Beetle showed low visitation probabilities in disturbed habitats in contrast to high visitation rates in native habitats [36]. Furthermore, climate change is also expected to strongly affect mountain ecosystems [31,59] and this will include beetle diversity. Suitable climatic conditions and habitat for alpine biota is declining and the trend is expected to continue in the future [60]. Therefore, to protect this underestimated biodiversity for future generations, conservation of natural environments and the plants that attract these beetle species are very important.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/d13110604/s1, Table S1. Sample sites, transects, GPS information and sampling date. Table S2. Beetle species and their families. The number of specimens for each species was given. Table S3. Plant species and their families. A plant marked as inflorescence refers to a large flower display in a more or less condensed inflorescence with many tiny flowers, including inflorescence type of Apiaceae (Umbel) and Asteraceae (Head). Table S4. Sampling completeness estimations for different elevations and habitats using the iNEXT method. Table S5. Network turnover among different transects (n = 12). H and L refer to high elevation and low elevation, F and M refer to forest and meadow. Figure S1. An example of sampling completeness (species coverage) for three forest transects at the high elevation. Figure S2. An example of sampling completeness (species diversity) for three forest transects at the high elevation.
Author Contributions
Conceptualization, K.-Q.L., Z.-X.R. and Q.L.; methodology, K.-Q.L., Z.-X.R.; software: Z.-X.R.; formal analysis: K.-Q.L. and Z.-X.R.; investigation: K.-Q.L.; writing: K.-Q.L. and Z.-X.R.; review and editing: Z.-X.R. and Q.L. All authors have read and agreed to the published version of the manuscript.
Funding
This study is supported by grants from the National Natural Science Foundation of China (Grant No. 31702047), the Biological Resources Program, Chinese Academy of Sciences (KFJ-BRP-017-21) to KQL, and a Talent Young Scientist Program of Yunnan Province (YNWR-QNBJ-2019-055) to ZXR.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in article and Supplementary Materials.
Acknowledgments
We are grateful to Ningnian Xiao, Yuxia Yang, Yongying Ruan, Zhiliang Wang, Wangang Liu, Yuanyuan Lu, Meike Liu, Qilong Lei, You Li, and Haitian Song for identification of beetles. We also thank Xin Xu of Kunming Institute of Botany, Chinese Academy of Sciences for helping with the statistical analyses, Lijiang Forest Biodiversity National Observation and Research Station for allowing us access to sites and for their logistic support. We thank Peter Bernhardt of the Missouri Botanical Garden for helping us revise grammar and sentence structure.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ollerton, J.; Winfree, R.; Tarrant, S. How many flowering plants are pollinated by animals? Oikos 2011, 120, 321–326. [Google Scholar] [CrossRef]
- Slipinski, S.A.; Leschen, R.A.B.; Lawrence, J.F. Order Coleoptera Linneaus, 1758. In Animal Biodiversity: An Outline of Higher-Level Classification and Survey of Taxonomic Richness; Zhang, Z.Q., Ed.; Magnolia Press: Auckland, New Zealand, 2011; Volume 3148, pp. 203–208. [Google Scholar]
- Bao, T.; Walczyska, K.S.; Moody, S.; Wang, B.; Rust, J. New family Apotomouridae fam. nov. (Coleoptera: Tenebrionoidea) from lower Cenomanian amber of Myanmar. Cretac. Res. 2018, 91, 14–19. [Google Scholar] [CrossRef]
- Wardhaugh, C.W. How many species of arthropods visit flowers? Arthropod-Plant Interact. 2015, 9, 547–565. [Google Scholar] [CrossRef]
- Ollerton, J. Pollinator Diversity: Distribution, Ecological Function, and Conservation. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 353–376. [Google Scholar] [CrossRef] [Green Version]
- Bernhardt, P. Convergent evolution and adaptive radiation of beetle-pollinated angiosperms. Plant Syst. Evol. 2000, 222, 293–320. [Google Scholar] [CrossRef]
- Prance, G.; Arias, J. A study of the floral biology of Victoria amazonica (Poepp.) Sowerby (Nymphaeaceae). Acta Amaz. 1975, 6, 163–170. [Google Scholar] [CrossRef] [Green Version]
- Schneider, E.L.; Buchanan, J. Morphological studies of the Nymphaeaceae. XI. The floral biology of Nelumbo pentapetala. Amer. J. Bot. 1980, 67, 182–193. [Google Scholar] [CrossRef]
- Goldblatt, P.; Bernhardt, P.; Manning, J.C. Pollination of petaloid geophytes by monkey beetles (Scarabaeidae: Rutelinae: Hopliini) in southern Africa. Ann. Missouri Bot. Gard. 1998, 85, 215–230. [Google Scholar] [CrossRef]
- Dieringer, G.; Cabrera, R.L.; Lara, M.; Loya, L.; Resyes-Castillo, P. Beetle pollination and floral thermogenicity in Magnolia tamaulipana (Magnoliaceae). Int. J. Plant Sci. 1999, 160, 64–71. [Google Scholar] [CrossRef]
- Sakai, S.; Kuniyasu, M.; Ymoto, T.; Kato, M.; Inoue, T. A new pollination system: Dung-beetle pollination discovered in Orchidantha inouei (Lowiaceae, Zingiberales) in Sarawak, Malaysia. Amer. J. Bot. 1999, 86, 56–61. [Google Scholar] [CrossRef]
- Corlett, R.T. Pollination in a degraded tropical landscape: A Hong Kong case study. J. Trop. Ecol. 2001, 17, 155–161. [Google Scholar] [CrossRef] [Green Version]
- Sayers, T.D.J.; Steinbauer, M.J.; Miller, R.E. Visitor or vector? The extent of rove beetle (Coleoptera: Staphylinidae) pollination and floral interactions. Arthropod-Plant Interact. 2019, 13, 685–701. [Google Scholar] [CrossRef]
- Kirmse, S.; Chaboo, C.S. Flowers are essential to maintain high beetle diversity (Coleoptera) in a Neotropical rainforest canopy. J. Nat. Hist. 2020, 54, 1661–1696. [Google Scholar] [CrossRef]
- Armstrong, J.E.; Irvine, A.K. Floral Biology of Myristica-Insipida (Myristicaceae), a Distinctive Beetle Pollination Syndrome. Am. J. Bot. 1989, 76, 86–94. [Google Scholar] [CrossRef]
- Hansman, D.J. Floral biology of dry rainforest in north Queensland and a comparison with adjacent savanna woodland. Aust. J. Bot. 2001, 49, 137–153. [Google Scholar] [CrossRef]
- Momose, K.; Yumoto, T.; Teruyoshi, N.; Kato, M.; Nagamasu, H.; Sakai, S.; Harrison, R.; Ttioka, T.; Hamid, A.; Inoue, T. Pollination biology in a lowland dipterocarp forest in Sarawak, Malaysia. I. Characteristics of the plant-pollinator community in a lowland dipterocarp forest. Am. J. Bot. 1998, 85, 1477–1501. [Google Scholar] [CrossRef]
- Wardhaugh, C.W.; Stork, N.E.; Edwards, W.; Grimbacher, P.S. The overlooked biodiversity of flower-visiting invertebrates. PLoS ONE 2012, 7, e45796. [Google Scholar] [CrossRef] [Green Version]
- Gottsberger, G. The structure and function of the primitive angiosperm flower-a discussion. Acta Bot. Neerl. 1974, 23, 461–471. [Google Scholar] [CrossRef]
- Wardhaugh, C.W.; Edwards, W.; Stork, N.E. The specialization and structure of antagonistic and mutualistic networks of beetles on rainforest canopy trees. Biol. J. Linn. Soc. 2015, 114, 287–295. [Google Scholar] [CrossRef]
- Körner, C. Mountain biodiversity, its causes and function. AMBIO 2004, 33, 11–17. [Google Scholar] [CrossRef]
- Körner, C. Coldest places on earth with angiosperm plant life. Alp. Bot. 2011, 121, 11–22. [Google Scholar] [CrossRef]
- Körner, C.; Jetz, W.; Paulsen, J.; Payne, D.; Rudmann-Maurer, K.; Spehn, E.M. A global inventory of mountains for bio-geographical applications. Alp. Bot. 2017, 127, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Chape, S.; Spalding, M.; Jenkins, M. The World’s Protected Areas: Status, Values and Prospects in the 21st Century; University of California Press: Berkeley, CA, USA, 2008. [Google Scholar]
- Arroyo, M.T.K.; Primack, R.; Armesto, J. Community studies in pollination ecology in the high temperate Andes of central Chile. I. Pollination mechanisms and altitudinal variation. Am. J. Bot. 1982, 69, 82–97. [Google Scholar] [CrossRef]
- Müller, H. The fertilisers of alpine flowers. Nature 1880, 21, 275. [Google Scholar] [CrossRef] [Green Version]
- Heithaus, E.R. The role of plant-pollinator interactions in determining community structure. Ann. Mo. Bot. Gard. 1974, 61, 675–691. [Google Scholar] [CrossRef]
- Moldenke, A.R. California pollination ecology and vegetation types. Phytologia 1976, 34, 305–361. [Google Scholar]
- Warren, S.D.; Harper, K.T.; Booth, G.M. Elevational distribution of insect pollinators. Am. Midl. Nat. 1988, 120, 325–330. [Google Scholar] [CrossRef]
- Medan, D.; Montaldo, N.H.; Devoto, M.; Maniese, A.; Vasellati, V.; Roitman, G.G.; Bartoloni, N.H. Plant-pollinator Relationships at Two Altitudes in the Andes of Mendoza, Argentina. Arct. Antarct. Alp. Res. 2002, 34, 233–241. [Google Scholar] [CrossRef]
- Lefebvre, V.; Villemant, C.; Fontaine, C.; Daugeron, C. Altitudinal, temporal and trophic partitioning of flower-visitors in Alpine communities. Sci. Rep. 2018, 8, 4706. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.H.; Ren, Z.X.; Lázaro, A.; Wang, H.; Bernhardt, P.; Li, H.D.; Li, D.Z. Floral traits influence pollen vectors’ choices in higher elevation communities in the Himalaya-Hengduan Mountains. BMC Ecol. 2016, 16, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.H.; Lázaro, A.; Ren, Z.X.; Zhou, W.; Li, H.D.; Tao, Z.B.; Xu, K.; Wu, Z.K.; Wolfe, L.M.; Li, D.Z.; et al. The topological differences between visitation and pollen transport networks: A comparison in species rich communities of the Himalaya-Hengduan Mountains. Oikos 2019, 128, 551–562. [Google Scholar] [CrossRef]
- Doré, M.; Fontaine, C.; Thébault, E. Relative effects of anthropogenic pressures, climate, and sampling design on the structure of pollination networks at the global scale. Glob. Chang. Biol. 2021, 27, 1266–1280. [Google Scholar] [CrossRef] [PubMed]
- Adedoja, O.A.; Kehinde, T.; Samways, M.J. Insect-flower interaction networks vary among endemic pollinator taxa over an elevation gradient. PLoS ONE 2018, 13, e0207453. [Google Scholar] [CrossRef] [PubMed]
- Herrera, C.M. Flower traits, habitat, and phylogeny as predictors of pollinator service: A plant community perspective. Ecol. Monogr. 2020, 90, e01402. [Google Scholar] [CrossRef]
- Herrera, C.M.; Otero, C. Plant phylogeny as a major predictor of flower visitation by nitidulid beetles, a lineage of ancestral angiosperm pollinators. J. Pollinat. Ecol. 2021, 28, 179–188. [Google Scholar] [CrossRef]
- Kantsa, A.; Raguso, R.A.; Dyer, A.G.; Olesen, J.M.; Tscheulin, T.; Petanidou, T. Disentangling the role of floral sensory stimuli in pollination networks. Nat. Commun. 2018, 9, 1041. [Google Scholar] [CrossRef]
- Memmott, J. The structure of a plant-pollinator food web. Ecol. Lett. 1999, 2, 276–280. [Google Scholar] [CrossRef]
- Alarcón, R. Congruence between visitation and pollen-transport networks in a California plant–pollinator community. Oikos 2010, 119, 35–44. [Google Scholar] [CrossRef]
- Xu, X.; Ren, Z.X.; Trunschke, J.; Kuppler, J.; Zhao, Y.H.; Knop, E.; Wang, H. Bimodal activity of diurnal flower visitation at high elevation. Ecol. Evol. 2021, 11, 13487–13500. [Google Scholar] [CrossRef]
- Liang, H.; Zhao, Y.H.; Rafferty, N.E.; Ren, Z.X.; Zhong, L.; Li, H.D.; Li, D.Z.; Wang, H. Evolutionary and ecological factors structure a plant-bumblebee network in a biodiversity hotspot, the Himalaya-Hengduan Mountains. Funct. Ecol. 2021, 35, 2523–2535. [Google Scholar] [CrossRef]
- Wardhaugh, C.W. The importance of flowers for beetle biodiversity and abundance. In Treetops at Risk: Challenges of Global Canopy Ecology and Conservation; Lowman, M., Devy, M., Ganesh, S.T., Eds.; Springer: New York, NY, USA, 2013; pp. 275–288. [Google Scholar]
- Zheng, L.Y.; Gui, H. Insect Classification; Nanjing Normal University Press: Nanjing, China, 1999. [Google Scholar]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Dormann, C.F.; Gruber, B.; Fründ, J. Introducing the bipartite package: Analysing ecological networks. R News 2008, 8, 8–11. [Google Scholar]
- R Core Team. R: A language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Blüthgen, N.; Menzel, F.; Blüthgen, N. Measuring specialization in species interaction networks. BMC Ecol. 2006, 6, 9. [Google Scholar] [CrossRef] [Green Version]
- Olesen, J.M.; Bascompte, J.; Dupont, Y.L.; Jordano, P. The modularity of pollination networks. Proc. Natl. Acad. Sci. USA 2007, 104, 19891–19896. [Google Scholar] [CrossRef] [Green Version]
- Beckett, S.J. Improved community detection in weighted bipartite networks. R Soc. Open Sci. 2016, 3, 140536. [Google Scholar] [CrossRef] [Green Version]
- Poisot, T.; Canard, E.; Mouillot, D.; Mouquet, N.; Gravel, D. The dissimilarity of species interaction networks. Ecol. Lett. 2012, 15, 1353–1361. [Google Scholar] [CrossRef]
- Schwarz, B.; Dormann, C.F.; Vázquez, D.P.; Fründ, J. Within-day dynamics of plant–pollinator networks are dominated by early flower closure: An experimental test of network plasticity. Oecologia 2021, 196, 781–794. [Google Scholar] [CrossRef]
- Cook, S.M.; Bartlet, E.; Murray, D.A.; Williams, I.H. The role of pollen odour in the attraction of pollen beetles to oilseed rape flowers. Entomol. Exp. Appl. 2002, 104, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Jürgens, A.; Dötterl, S. Chemical composition of anther volatiles in Ranunculaceae: Genera-specific profiles in Anemone, Aquilegia, Caltha, Pulsatilla, Ranunculus, and Trollius species. Am. J. Bot. 2004, 91, 1969–1980. [Google Scholar] [CrossRef]
- Pyke, G.H.; Ren, Z.X.; Trunschke, J.; Lunau, K.; Wang, H. Salvage of floral resources through re-absorption before flower abscission. Sci. Rep. 2020, 10, 15960. [Google Scholar] [CrossRef]
- Lunau, K.; Ren, Z.X.; Fan, X.Q.; Pyke, G.H.; Trunschke, J.; Wang, H. Nectar mimicry: A new phenomenon. Sci. Rep. 2020, 10, 7039. [Google Scholar] [CrossRef] [PubMed]
- De Vega, C.; Álvarez-Pérez, S.; Albaladejo, R.G.; Steenhuisen, S.-L.; Lachance, M.-A.; Johnson, S.D.; Herrera, C.M. The role of plant–pollinator interactions in structuring nectar microbial communities. J. Ecol. 2021, in press. [Google Scholar]
- Millard, J.; Outhwaite, C.L.; Kinnersley, R.; Freeman, R.; Gregory, R.D.; Adedoja, O.; Gavini, S.; Kioko, E.; Kuhlmann, M.; Ollerton, J.; et al. Global effects of land-use intensity on local pollinator biodiversity. Nat. Commun. 2021, 12, 2902. [Google Scholar] [CrossRef] [PubMed]
- Spehn, E.M.; Rudmann-Maurer, K.; Körner, C.; Maselli, D. Mountain Biodiversity and Global Change; GMBA-DIVERSITAS: Basel, Switzerland, 2010. [Google Scholar]
- Ernakovich, J.G.; Hopping, K.A.; Berdanier, A.B.; Simpson, R.T.; Kachergis, E.J.; Steltzer, H.; Wallenstein, M.D. Predicted responses of arctic and alpine ecosystems to altered seasonality under climate change. Glob. Chang. Biol. 2014, 20, 3256–3269. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Flowers and their beetles on Yulong. Living, uncollected insects were not easy to identify to species and most here are identified to family and/or genus. (A): Trollius yunnanensis (Franch.) Ulbr. (Ranunculaceae) and a species in the Tenebrionoidea; (B): Hypericum bellum Li (Hypericaceae) visited by one species in the Rutelidae, and at least one species in the Chrysomelidae; (C): Potentilla fulgens Wall. ex Hook (Rosaceae). visited by a species in the Melolonthidae; note it is eating the petals; (D): Verbascum thapsus L. (Scrophulariaceae) visited by a species in the Alticinae (Chrysomelidae); (E): Ligularia alatipes Hand.-Mazz. (Asteraceae) visited by Northora sp. (Chrysomelidae: Alticinae); (F): Polygonum paleaceum Wall. ex Hook. f. visited by a species in the Tenebrionoidea; (G): Thalictrum delavayi Franch. (Ranunculaceae) visited by a species of Galerucinae (Chrysomelidae); (H): Adenophora coelestis Diels (Campanulaceae) with pendant flowers that were always inhabited/visited by small beetles in the family Nitidulidae; (I): Rhododendron decorum Franch. (Ericaceae) was visited frequently by members of the family Nitidulidae; (J): Parnassia wightiana Wall. ex Wight et Arn. (Celastraceae) visited by Northora sp. in the Alticinae (Chrysomelidae), note that its staminodes were eaten by these beetles; (K): Juncus allioides Franch. (Juncaceae) visited by Northora sp. in the Alticinae (Chrysomelidae); (L): Pyracantha fortuneana (Maxim.) Li (Rosaceae) visited by Popillia sp. (Rutelidae); (M): Dipsacus asper Wallich ex Candolle (Caprifoliaceae) visited by a species in the Galerucinae (Chrysomelidae); (N): Leontopodium calocephalum (Franch.) Beauv. (Asteraceae) visited by Northora sp. in the Alticinae (Chrysomelidae); (O): Physospermopsis delavayi (Franch.) Wolff (Apiaceae) visited by a species in the Cantharidae; (P): Pimpinella yunnanensis (Franch.) Wolff (Apiaceae) visited by an unidentified beetle.
Figure 1.
Flowers and their beetles on Yulong. Living, uncollected insects were not easy to identify to species and most here are identified to family and/or genus. (A): Trollius yunnanensis (Franch.) Ulbr. (Ranunculaceae) and a species in the Tenebrionoidea; (B): Hypericum bellum Li (Hypericaceae) visited by one species in the Rutelidae, and at least one species in the Chrysomelidae; (C): Potentilla fulgens Wall. ex Hook (Rosaceae). visited by a species in the Melolonthidae; note it is eating the petals; (D): Verbascum thapsus L. (Scrophulariaceae) visited by a species in the Alticinae (Chrysomelidae); (E): Ligularia alatipes Hand.-Mazz. (Asteraceae) visited by Northora sp. (Chrysomelidae: Alticinae); (F): Polygonum paleaceum Wall. ex Hook. f. visited by a species in the Tenebrionoidea; (G): Thalictrum delavayi Franch. (Ranunculaceae) visited by a species of Galerucinae (Chrysomelidae); (H): Adenophora coelestis Diels (Campanulaceae) with pendant flowers that were always inhabited/visited by small beetles in the family Nitidulidae; (I): Rhododendron decorum Franch. (Ericaceae) was visited frequently by members of the family Nitidulidae; (J): Parnassia wightiana Wall. ex Wight et Arn. (Celastraceae) visited by Northora sp. in the Alticinae (Chrysomelidae), note that its staminodes were eaten by these beetles; (K): Juncus allioides Franch. (Juncaceae) visited by Northora sp. in the Alticinae (Chrysomelidae); (L): Pyracantha fortuneana (Maxim.) Li (Rosaceae) visited by Popillia sp. (Rutelidae); (M): Dipsacus asper Wallich ex Candolle (Caprifoliaceae) visited by a species in the Galerucinae (Chrysomelidae); (N): Leontopodium calocephalum (Franch.) Beauv. (Asteraceae) visited by Northora sp. in the Alticinae (Chrysomelidae); (O): Physospermopsis delavayi (Franch.) Wolff (Apiaceae) visited by a species in the Cantharidae; (P): Pimpinella yunnanensis (Franch.) Wolff (Apiaceae) visited by an unidentified beetle.

Figure 2.
The total network combining all interaction data from all sites. Green cycles representing plant species, orange cycles representing beetle species. The largest circles are labeled RD = Rhododendron decorum Franch. (RD), LA1 = Lamiogethes sp.1 (LA1) and NV = Nonarthra variabilis Baly (NV).
Figure 2.
The total network combining all interaction data from all sites. Green cycles representing plant species, orange cycles representing beetle species. The largest circles are labeled RD = Rhododendron decorum Franch. (RD), LA1 = Lamiogethes sp.1 (LA1) and NV = Nonarthra variabilis Baly (NV).

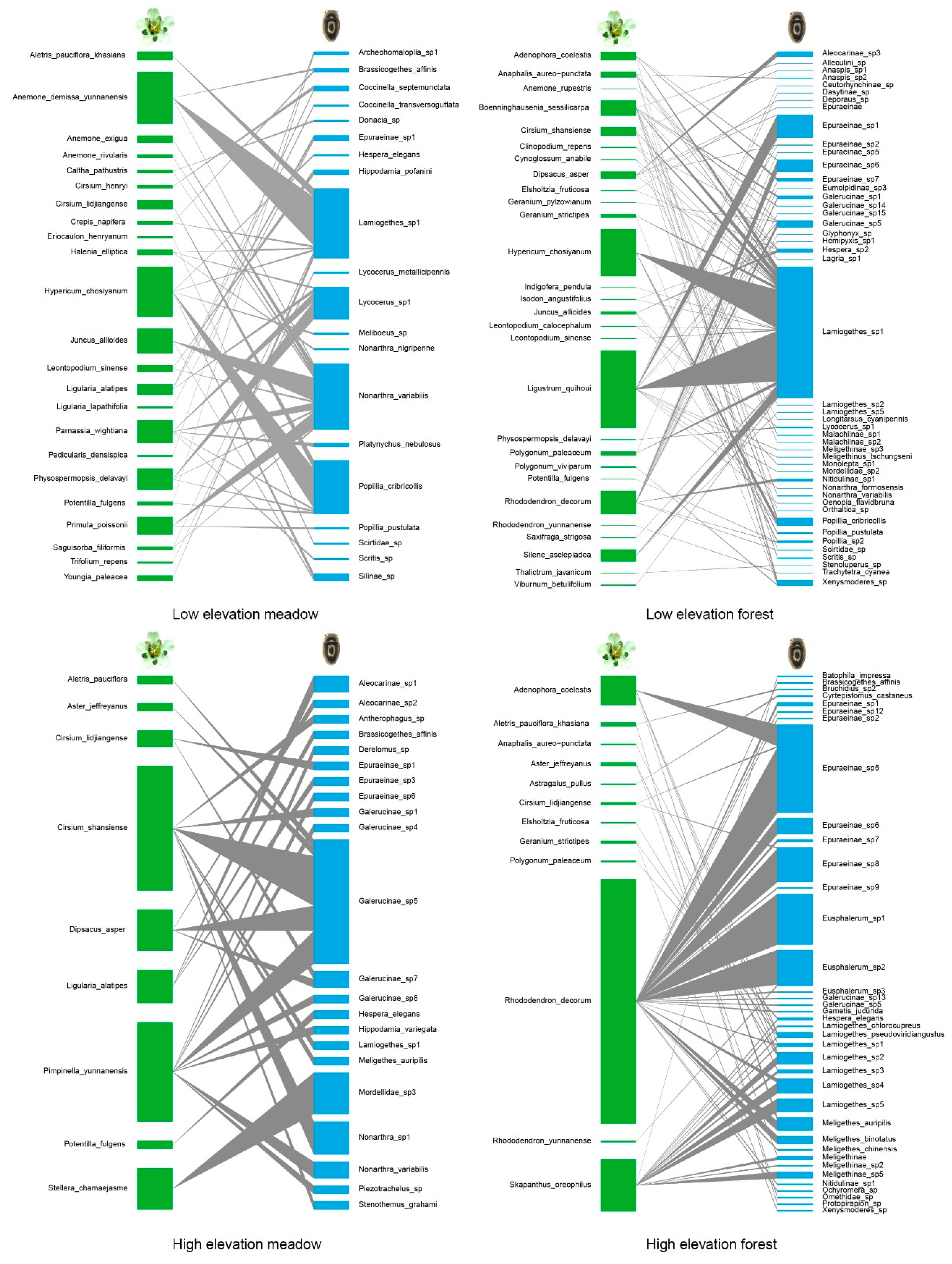
Figure 3.
Four networks representing plant and their flower visiting beetles. Green left side = plant species and blue right side = beetles. High elevation, LB; low elevation, YH.
Figure 3.
Four networks representing plant and their flower visiting beetles. Green left side = plant species and blue right side = beetles. High elevation, LB; low elevation, YH.

Figure 4.
Species roles in the total network. (A). No network hub was detected for plants, while three species Ligularia alatipes Hand.-Mazz. (Asteraceae), Halenia elliptica D. Don (Gentianaceae) and Thalictrum javanicum Bl. (Ranunculaceae) were detected as module hubs. (B). One beetle species Galerucinae sp.5 (Chrysomelidae) was detected as a network hub. Six species were detected as module hubs, and six species (marked as 1–6) as connectors.
Figure 4.
Species roles in the total network. (A). No network hub was detected for plants, while three species Ligularia alatipes Hand.-Mazz. (Asteraceae), Halenia elliptica D. Don (Gentianaceae) and Thalictrum javanicum Bl. (Ranunculaceae) were detected as module hubs. (B). One beetle species Galerucinae sp.5 (Chrysomelidae) was detected as a network hub. Six species were detected as module hubs, and six species (marked as 1–6) as connectors.

Figure 5.
Network turnover comparison between elevation and within elevation (among habitats). From left to right. S Beta diversity (βS), ST Beta diversity (βST) and WN Beta diversity (βWN) were always higher between elevation than within elevation (all p < 0.05).
Figure 5.
Network turnover comparison between elevation and within elevation (among habitats). From left to right. S Beta diversity (βS), ST Beta diversity (βST) and WN Beta diversity (βWN) were always higher between elevation than within elevation (all p < 0.05).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Classification and number of specimens representing each beetle family.
| Family | No. of Species | No. of Specimens | Family | No. of Species | No. of Specimens |
|---|---|---|---|---|---|
| Chrysomelidae | 42 | 955 | Eumolpididae | 3 | 6 |
| Nitidulidae | 34 | 1657 | Melyeridae | 3 | 6 |
| Curculionidae | 10 | 45 | Melolonthidae | 2 | 5 |
| Coccinellidae | 10 | 28 | Brentidae | 2 | 2 |
| Cantharidae | 7 | 141 | Scirtidae | 2 | 7 |
| Staphylinidae | 7 | 115 | Cetoniidae | 1 | 4 |
| Crioceridae | 5 | 173 | Omethidae | 1 | 4 |
| Elateridae | 5 | 55 | Attelabidae | 1 | 1 |
| Tenebrionidae | 5 | 12 | Buprestidae | 1 | 1 |
| Rutelidae | 3 | 72 | Corylophidae | 1 | 1 |
| Mordellidae | 3 | 66 | Cryptophagidae | 1 | 1 |
| Scraptiidae | 3 | 33 | Lycidae | 1 | 1 |
Table 2.
Classification and identification of plant species and the relative abundance of beetle taxa visiting their flowers.
Table 2.
Classification and identification of plant species and the relative abundance of beetle taxa visiting their flowers.
| Order | Family | No. of Plant Genus | No. of Plant Species | No. of Beetle Species | No. of Specimens | No. of Visiting Beetle Family |
|---|---|---|---|---|---|---|
| Asterales | Asteraceae | 12 | 21 | 57 | 442 | 13 |
| Ericales | Ericaceae | 1 | 2 | 37 | 462 | 10 |
| Rosales | Rosaceae | 5 | 5 | 30 | 264 | 12 |
| Apiales | Apiaceae | 2 | 3 | 27 | 336 | 14 |
| Fabales | Fabaceae | 6 | 9 | 21 | 34 | 7 |
| Dipsacales | Caprifoliaceae | 1 | 1 | 18 | 463 | 4 |
| Lamiales | Lamiaceae | 4 | 4 | 17 | 54 | 4 |
| Sapindales | Rutaceae | 1 | 2 | 14 | 39 | 5 |
| Ranunculales | Ranunculaceae | 4 | 9 | 15 | 84 | 5 |
| Lamiales | Oleaceae | 2 | 2 | 12 | 183 | 6 |
| Malpighiales | Hypericaceae | 1 | 2 | 12 | 307 | 7 |
| Gentianales | Gentianaceae | 3 | 3 | 12 | 25 | 6 |
| Asterales | Campanulaceae | 1 | 1 | 12 | 87 | 2 |
| Celastrales | Celastraceae | 1 | 1 | 10 | 246 | 9 |
| Dioscoreales | Nartheciaceae | 1 | 2 | 8 | 20 | 7 |
| Caryophyllales | Polygonaceae | 2 | 4 | 8 | 16 | 5 |
| Lamiales | Scrophulariaceae | 1 | 1 | 7 | 73 | 3 |
| Geraniales | Geraniaceae | 1 | 3 | 6 | 17 | 4 |
| Poales | Juncaceae | 1 | 1 | 5 | 47 | 3 |
| Dipsacales | Adoxaceae | 1 | 2 | 5 | 19 | 2 |
| Cornales | Hydrangeaceae | 1 | 1 | 5 | 8 | 2 |
| Poales | Eriocaulaceae | 1 | 1 | 4 | 41 | 4 |
| Caryophyllales | Caryophyllaceae | 1 | 1 | 4 | 34 | 2 |
| Ericales | Primulaceae | 1 | 1 | 3 | 65 | 2 |
| Boraginales | Boraginaceae | 1 | 1 | 3 | 4 | 3 |
| Saxifragales | Saxifragaceae | 1 | 2 | 2 | 10 | 2 |
| Lamiales | Orobanchaceae | 1 | 2 | 2 | 4 | 2 |
| Malvales | Thymelaeaceae | 1 | 1 | 1 | 5 | 1 |
| Asparagales | Orchidaceae | 1 | 1 | 1 | 1 | 1 |
| Commelinales | Commelinaceae | 1 | 1 | 1 | 1 | 1 |
Table 3.
Network-level properties of beetle-plant networks (n = 12) from two elevations and two habitats. HL (high level of network) refers to beetles while LL (low level of network) refers to plants.
Table 3.
Network-level properties of beetle-plant networks (n = 12) from two elevations and two habitats. HL (high level of network) refers to beetles while LL (low level of network) refers to plants.
| Index | Overall (n = 12) | High Elevation Forest (n = 3) | High Elevation Meadow (n = 3) | Low Elevation Forest (n = 3) | Low Elevation Meadow (n = 3) | between Elevations (F Value) | between Habitats (F Value) | Interaction |
|---|---|---|---|---|---|---|---|---|
| Network size | 43.08 ± 4.25 | 37 ± 6.93 | 30.33 ± 1.76 | 58.67 ± 7.75 | 46.33 ± 7.69 | F = 8.329, p = 0.020 | F = 2.119, p = 0.184 | F = 0.189, p = 0.676 |
| No. of interaction | 282.58 ± 45.44 | 188.00 ± 40.99 | 244.33 ± 101.15 | 500.33 ± 17.27 | 197.68 ± 26.27 | F = 5.472, p = 0.047 | F = 4.704, p = 0.062 | F = 9.991, p = 0.013 |
| No. of species HL | 26.25 ± 3.11 | 27 ± 5.77 | 18 ± 2.08 | 36 ± 5.51 | 24 ± 7.77 | F = 1.753, p = 0.222 | F = 3.436, p = 0.101 | F = 0.0701, p = 0.798 |
| No. of species LL | 16.83 ± 1.90 | 10 ± 1.16 | 12.33 ± 2.03 | 22.67 ± 2.91 | 22.33 ± 0.33 | F = 36.698, p < 0.001 | F = 0.286, p = 0.608 | F = 0.508, p = 0.496 |
| NODF | 21.93 ± 2.74 | 26.10 ± 2.89 | 27.75 ± 9.58 | 16.44 ± 2.29 | 17.42 ± 3.07 | F = 3.482, p = 0.099 | F = 0.0598, p = 0.813 | F = 0.00397, p = 0.951 |
| Weighted connectance | 0.08 ± 0.01 | 0.11 ± 0.01 | 0.11 ± 0.01 | 0.05 ± 0.00 | 0.06 ± 0.01 | F = 39.214, p < 0.001 | F = 0.471, p = 0.512 | F = 1.340, p = 0. 280 |
| H2′ | 0.56 ± 0.03 | 0.56 ± 0.04 | 0.44 ± 0.05 | 0.57 ± 0.06 | 0.68 ± 0.01 | F = 7.191, p = 0.028 | F = 0.00320, p = 0.956 | F = 5.668, p = 0.044 |
| Niche overlap HL | 0.21 ± 0.04 | 0.38 ± 0.03 | 0.23 ± 0.02 | 0.15 ± 0.01 | 0.07 ± 0.01 | F = 100.330, p < 0.001 | F = 34.279, p < 0.001 | F = 3.113, p = 0.116 |
| Extinction slope HL | 1.96 ± 0.12 | 2.20 ± 0.24 | 1.67 ± 0.18 | 1.87 ± 0.08 | 2.09 ± 0.38 | F = 0.0337, p = 0.859 | F = 0.397, p = 0.546 | F = 2.34, p = 0.165 |
| Robustness HL | 0.65 ± 0.01 | 0.68 ± 0.02 | 0.62 ± 0.03 | 0.65 ± 0.01 | 0.66 ± 0.03 | F = 0.038, p = 0.850 | F = 0.909, p = 0.368 | F = 2.229, p = 0.174 |
| Functional complementarity HL | 264.91 ± 51.33 | 134.12 ± 15.63 | 244.10 ± 112.07 | 511.58 ± 32.44 | 169.84 ± 19.99 | F = 6.448, p = 0.035 | F = 3.768, p = 0.088 | F = 14.313, p = 0.005 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, K.-Q.; Ren, Z.-X.; Li, Q. Diversity of Flower Visiting Beetles at Higher Elevations on the Yulong Snow Mountain (Yunnan, China). Diversity 2021, 13, 604. https://0-doi-org.brum.beds.ac.uk/10.3390/d13110604
AMA Style
Li K-Q, Ren Z-X, Li Q. Diversity of Flower Visiting Beetles at Higher Elevations on the Yulong Snow Mountain (Yunnan, China). Diversity. 2021; 13(11):604. https://0-doi-org.brum.beds.ac.uk/10.3390/d13110604
Chicago/Turabian StyleLi, Kai-Qin, Zong-Xin Ren, and Qiang Li. 2021. "Diversity of Flower Visiting Beetles at Higher Elevations on the Yulong Snow Mountain (Yunnan, China)" Diversity 13, no. 11: 604. https://0-doi-org.brum.beds.ac.uk/10.3390/d13110604
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.