
Good Things Come in Larger Packages: Size Matters for Adult Fruit-Feeding Butterfly Dispersal and Larval Diet Breadth

 , , , and
, , , and
Abstract
:1. Introduction
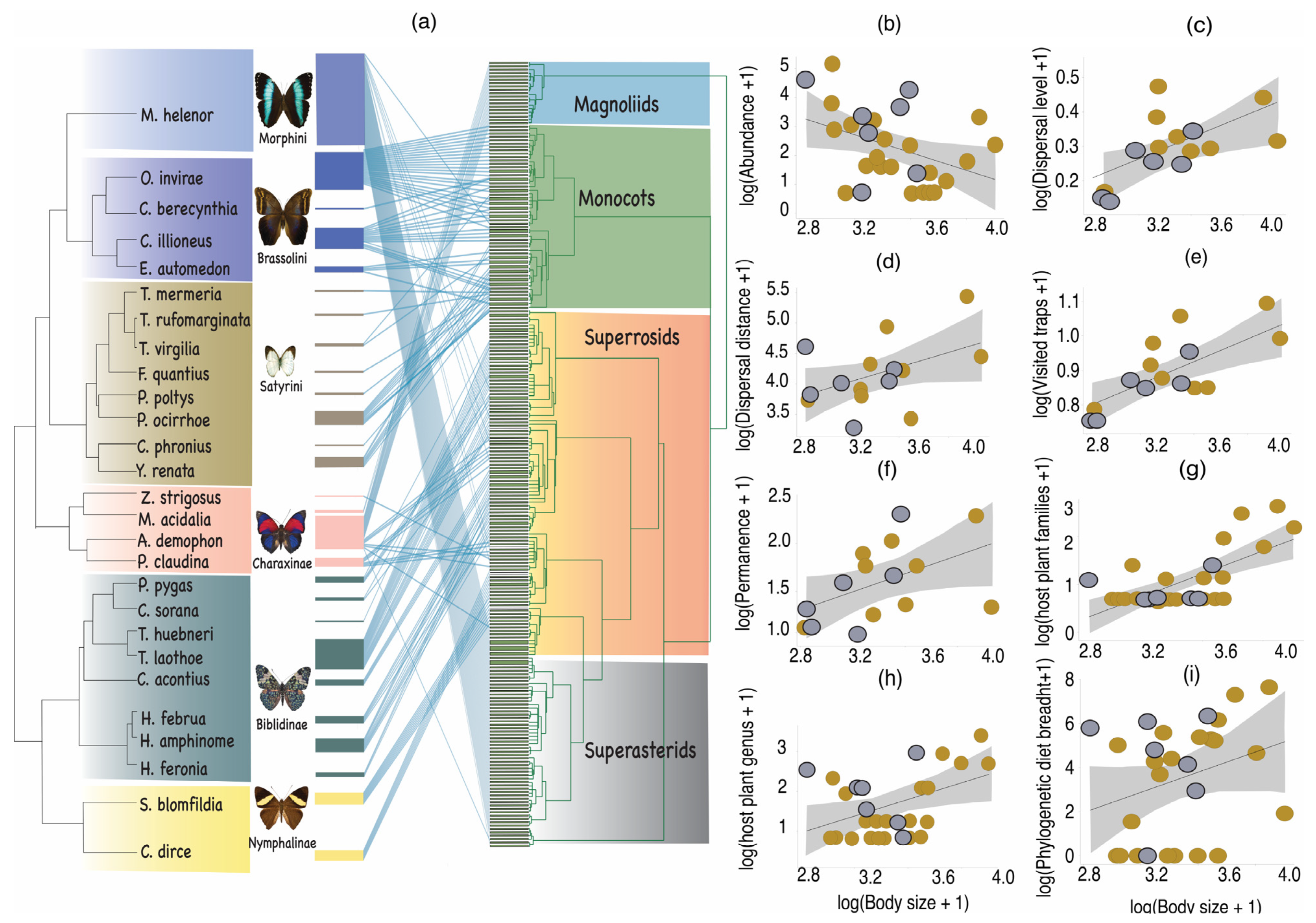
2. Materials and Methods
2.1. Study Site and Sampling Design
2.2. Fruit Feeding Butterflies
2.3. Phylogenetic, Morphological and Ecological Traits
2.4. Statistical Analysis
3. Results
4. Discussion
Trait-Dispersal Relationships
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Subfamily-Tribe/Species | FWL—mm | N | Disp. Level (%) | Distance—m Max (Mean ± Sd) | Perm.—Days Max (Mean ± Sd) | Diet Breadth Fam/Gen/Spp-Phy |
|---|---|---|---|---|---|---|
| Biblidinae | 30.5 ± 4.4 | 179 | 30 | 356.1 (50.7 ± 72.5) | 26 (5.8 ± 7.1) | - |
| Callicore sorana (Godart, [1824]) | 26.9 ± 1.6 | 16 | 30 | 123.1 (24.6 ± 48.3) | 6 (3.5 ± 3.6) | 1/3/3-121 |
| Catagramma pygas (Godart, [1824]) | 26.01 ± 1.4 | 4 | - | - | - | 1/2/4-21 |
| Catonephele acontius (Hübner, [1823]) | 32.9 ± 2.6 | 24 | 30 | 356.1 (140.5 ± 56.7) | 26 (6.3 ± 6.3) | 1/1/4-416 |
| Hamadryas amphinome (Linnaeus, 1767) | 39.01 ± 2.1 | 1 | - | - | 1/1/13-718 | |
| Hamadryas februa (Hübner, [1823]) | 33.4 ± 1.2 | 38 | 20 | 341.1 (55.8 ± 54.8) | 22 (5.5 ± 5.9) | 1/2/8-335 |
| Hamadryas feronia (Linnaeus, 1758) | 34.7 ± 2.1 | 70 | 40 | 266.2 (69.0 ± 88.7) | 11 (5.5 ± 6.3) | 1/1/6-618 |
| Temenis laothoe (Cramer, 1777) | 24.7 ± 1.6 | 25 | 30 | 157.1 (54.7 ± 49.1) | 20 (3.2 ± 4.3) | 1/6/21-616 |
| Temenis huebneri (Fruhstorfer, 1907) | 24.7 ± 1.3 | 1 | - | - | - | 1/6/21-na |
| Charaxinae | 28.1 ± 1.6 | 14 | 55 | 105.1 (47.8 ± 38.3) | 17(5.1 ± 5.9) | - |
| Archaeoprepona demophon (Hübner, [1814]) | 46.3 ± 1.4 | 2 | - | - | - | 11/16/46-1480 |
| Memphis acidalia (Hübner, [1819]) | 27.7 ± 1.4 | 6 | 50 | 105.1 (50.6 ± 47.3) | 17 (4.2 ± 4.5) | 2/2/3-na |
| Prepona claudina (Godart, [1824]) | 41.01 ± 1.4 | 1 | - | - | - | 6//6/7-471 |
| Zaretis strigosus (Gmelin, 1790) | 28.6 ± 1.6 | 5 | 60 | 69.1 (43.5 ± 33.5) | 11 (2.3 ± 2.9) | 1/1/2-231 |
| Nymphalinae | 30.5 ± 9.04 | 14 | 30 | 183.2 (75.8 ± 76.3) | 10 (3.5 ± 3.5) | - |
| Colobura dirce (Linnaeus, 1758) | 29.5 ± 2.3 | 12 | 30 | 183.2 (75.8 ± 76.3) | 10 (3.5 ± 3.5) | 1/1/7-699 |
| Smyrna blomfildia (Fabricius, 1781) | 40.01 ± 2.3 | 1 | - | - | - | 1/5/11-391 |
| Tigridia acesta (Linnaeus, 1758) | 22.01 ± 2.3 | 1 | - | - | - | 3/5/8-370 |
| Satyrinae-Brassolini | 52.6 ± 11.8 | 23 | 36 | 422 (124.7 ± 142.1) | 17 (3.8 ± 4.9) | - |
| Caligo illioneus (Cramer, 1775) | 65.9 ± 4.0 | 10 | 20 | 75.6 (60.4 ± 21.4) | 5 (3.2 ± 3.7) | 8//12/16-583 |
| Catoblepia berecynthia (Cramer, 1777) | 41.01 ± 2.2 | 3 | - | - | - | 1/2/2-na |
| Eryphanis automedon (C. Felder & R. Felder, 1867) | 53.9 ± 2.2 | 5 | 60 | 422.8 (237.3 ± 160.8) | 17 (4.8 ± 4.6) | 4/9/12-351 |
| Opsiphanes invirae (Hübner, [1808]) | 38.1 ± 2.0 | 5 | 30 | 40.2 (30.1 ± 12.5) | 12 (3.2 ± 3.5) | 3/17/29-566 |
| Satyrinae-Morphini | 59.1 ± 4.1 | 27 | 30 | 142.8 (86.5 ± 44.6) | 10 (3.1 ± 3.4) | - |
| Morpho helenor (C. Felder & R. Felder, 1867) | 59.1 ± 4.1 | 27 | 30 | 142.8 (86.5 ± 44.6) | 10 (3.1 ± 3.4) | 11/30/58-2094 |
| Satyrinae-Satyrini | 22.3 ± 7.1 | 282 | 15 | 272.1 (49.2 ± 53.3) | 21 (2.3 ± 3.1) | - |
| Fosterinaria quantius (Godart, [1824]) | 23.1 ± 0.8 | 20 | - | - | - | 1/1/1-na |
| Pareuptychia ocirrhoe (Fabricius, 1776) | 19.9 ± 1.4 | 174 | 10 | 151.9 (41.7 ± 38.5) | 18 (3.4 ± 4.3) | 1/8/11-359 |
| Cissia phronius (Godart, [1824]) | 19.7 ± 1.1 | 47 | 10 | 272.1 (100.9 ± 78.7) | 21 (3.3 ± 4.6) | 1/1/1-na |
| Paryphthimoides poltys (Prittwitz, 1865) | 20.2 ± 0.8 | 17 | 10 | 111.2 (45.1 ± 29.5) | 13 (2.5 ± 3.2) | 1/1/1-na |
| Taygetis virgilia (Cramer, 1776) | 36.01 ± 1.6 | 10 | - | - | - | 2/2//2-216 |
| Taygetis laches (Fabricius, 1793) | 35.7 ± 1.4 | 10 | 30 | 171.1 (66.9 ± 52.1) | 8 (2.1 ± 2.6) | 2/2/2-216 |
| Taygetis mermeria (Cramer, 1776) | 30.01 ± 1.4 | 4 | - | - | - | 1/2/2-200 |
| Total | - | 539 | 30 | 422.8 (237.3 ± 160.8) | 26 (6.3 ± 6.3) | - |
References
- Phillips, B.L.; Brown, G.P.; Shine, R. Evolutionarily accelerated invasions: The rate of dispersal evolves upwards during the range advance of cane toads. J. Evol. Biol. 2010, 23, 2595–2601. [Google Scholar] [CrossRef]
- Stevens, V.M.; Trochet, A.; Dyck, H.V.; Colbert, J.; Baguete, M. How is dispersal integrated in life histories: A quantitative analysis using butterflies. Ecol. Lett. 2012, 15, 74–86. [Google Scholar] [CrossRef]
- Fountain-Jones, N.M.; Baker, S.C.; Jordan, G.J. Moving beyond the guild concept: Developing a practical functional trait framework for terrestrial beetles. Ecol. Entomol. 2015, 40, 1–13. [Google Scholar] [CrossRef]
- Spaniol, R.L.; Duarte, L.S.; Mendonça, M.S., Jr.; Iserhard, C.A. Combining functional traits and phylogeny to disentangling Amazonian butterfly assemblages on anthropogenic gradients. Ecosphere 2019, 10, e02837. [Google Scholar] [CrossRef] [Green Version]
- Faleiro, F.; Machado, R.B.; Loyola, R.D. Defining spatial conservation priorities in the face of land-use and climate change. Biol Conserv. 2013, 158, 248–257. [Google Scholar] [CrossRef]
- Brown, J.H.; Gillooly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Toward a metabolic theory of ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Marini–Filho, O.J.; Martins, R.P. Nymphalid butterfly dispersal among forest fragments at Serra da Canastra National Park, Brazil. J. Insect Conserv. 2010, 14, 401–411. [Google Scholar] [CrossRef]
- Gillespie, M.A.K.; Birkemoe, T.; Sverdrup-Thygeson, A. Interactions between body size, abundance, seasonality and phenology in forest beetles. Ecol. Evol. 2017, 7, 1091–1100. [Google Scholar] [CrossRef] [PubMed]
- Molleman, F.; Zwaan, B.J.; Brakefield, P.M.; Carey, J.R. Extraordinary long life spans in fruit-feeding butterflies can provide window on evolution of life span and aging. Exp. Gerontol. 2007, 42, 472–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, J. Phylogenetic and ecological correlates with male adult life span of rainforest butterflies. Evol. Ecol. 2008, 22, 507–517. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, D.B.; Freitas, A.V.L. Large-sized insects show stronger seasonality than small-sized ones: A case study of fruit-feeding butterflies. Biol. J. Linn. Soc. 2011, 104, 820–827. [Google Scholar] [CrossRef]
- Uehara–Prado, M.; Brown, K.S., Jr.; Freitas, A.V.L. Species richness, composition and abundance of fruit-feeding butterflies in the Brazilian Atlantic Forest: Comparison between a fragmented and a continuous landscape. Glob. Ecol. Biogeogr. 2007, 16, 43–54. [Google Scholar] [CrossRef]
- Shahabuddin, G.; Ponte, C.A. Frugivorous butterfly species in tropical forest fragments: Correlates of vulnerability to extinction. Biodivers. Conserv. 2005, 14, 1137–1152. [Google Scholar] [CrossRef]
- Skórka, P.; Kudlek, J.; Pepkowska, A.; Sliwinska, E.B.; Settele, J.; Woyciechowski, M. Movements and flight morphology in the endangered large blue butterflies. Cent. Eur. J. Biol. 2013, 7, 662–669. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, P.R.; Raven, P.H. Butterflies and plants: A study in coevolution. Evolution 1964, 18, 586–608. [Google Scholar] [CrossRef]
- Forister, M.L.; Dyer, L.A.; Singer, M.S.; Stireman, J.O.; Lill, J.T. Revisiting the evolution of ecological specialization, with emphasis on insect–plant interactions. Ecology 2012, 93, 981–991. [Google Scholar] [CrossRef]
- MacArthur, R.H. Geographical Ecology: Patterns in the Distribution of Species; Harper & Row: New York, NY, USA, 1972. [Google Scholar]
- Davis, R.B.; Õunap, E.; Javois, J.; Gerhold, P.; Tammaru, T. Degree of specialization is related to body size in herbivorous insects: A phylogenetic confirmation. Evolution 2012, 67, 583–589. [Google Scholar] [CrossRef]
- Hjalmarsson, A.; Bergsten, J.; Monaghan, M.T. Dispersal is linked to habitat use in 59 species of water beetles (Coleoptera: Adephaga) on Madagascar. Ecography 2015, 38, 732–739. [Google Scholar] [CrossRef]
- Sudta, C.; Salcido, D.M.; Forister, M.L.; Walla, T.; Villarín-Cortez, S.; Dyer, L.A. Jack-of-all trades paradigm meets long-term data: Generalist herbivores are more widespread and locally less abundant. bioRxiv 2020. [Google Scholar] [CrossRef]
- Gaston, K.J. Patterns in local and regional dynamics of moth populations. Oikos 1988, 53, 49–57. [Google Scholar] [CrossRef]
- Reavey, D. Egg size, first instar behavior and the ecology of Lepidoptera. J. Zool. 1992, 227, 277–297. [Google Scholar] [CrossRef]
- Pavoine, S.; Baguette, M.; Stevens, V.M.; Leibold, M.A.; Turlure, C.; Bonsall, M.B. Life history traits, but not phylogeny, drive compositional patterns in a butterfly metacommunity. Ecology 2014, 95, 3304–3313. [Google Scholar] [CrossRef] [Green Version]
- Duarte, L.D.S.; Debastiani, V.J.; Carlucci, M.B.; Diniz-Filho, J.A.F. Analyzing community-weighted trait means across environmental gradients: Should phylogeny stay or should it go? Ecology 2014, 99, 385–398. [Google Scholar] [CrossRef] [PubMed]
- Iserhard, C.A.; Duarte, L.; Seraphim, N.; Freitas, A.V.L. How urbanization affects multiple dimensions of biodiversity in tropical butterfly assemblages. Biodivers. Conserv. 2019, 28, 621–638. [Google Scholar] [CrossRef]
- Graça, M.B.; Pequeno, P.A.C.L.; Frankling, E.; Morais, J.W. Coevolution between flight morphology, vertical stratification and sexual dimorphism: What can we learn from tropical butterflies? J. Evol. Biol. 2017, 30, 1862–1871. [Google Scholar] [CrossRef] [Green Version]
- Santos, A.C.; Sales, P.C.L.; Ribeiro, D.B. Habitat conversion affects beta diversity in frugivorous butterfly assemblages. Stud. Neotrop. Fauna Environ. 2020, 55, 180–192. [Google Scholar] [CrossRef]
- Lourenço, G.M.; Soares, G.R.; Santos, T.P.; Dáttilo, W.; Freitas, A.V.L.; Ribeiro, S.P. Equal but different: Natural ecotones are dissimilar to anthropic edges. PLoS ONE 2019, 14, e0213008. [Google Scholar] [CrossRef] [PubMed]
- Freire, G.B.F., Jr.; Diniz, I.R. Temporal dynamics of fruit-feeding butterflies (Lepidoptera: Nymphalidae) in two habitats in a Brazilian environment. Fla. Entomol. 2015, 98, 1207–1216. [Google Scholar]
- De Vries, P.J. Stratification of fruit-feeding nymphalid butterflies in a Costa Rican rainforest. J. Res. Lepid. 1988, 26, 98–108. [Google Scholar]
- Freitas, A.V.L.; Iserhard, C.A.; Santos, J.P.; Carreira, J.Y.O.; Ribeiro, D.B.; Melo, D.H.A.; Rosa, A.H.B.; Marini-Filho, O.J.; Accacio, G.M.; Uehara-Prado, M. Studies with butterfly bait traps: An overview. Rev. Colomb. Entomol. 2014, 40, 209–218. [Google Scholar]
- Espírito-Santo, M.M.; Leite, M.E.; Silva, J.O.; Barbosa, R.S.; Rocha, A.M.; Anaya, F.C.; Dupin, M.G.V. Understanding patterns of land-cover change in the Brazilian Cerrado from 2000 to 2015. Philos. Trans. R. Soc. 2016, 371, 20150435. [Google Scholar] [CrossRef] [Green Version]
- Revell, L.J. Phytools: An R package for phylogenetic comparative biology (and other things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Wahlberg, N.; Leneveu, J.; Kodandaramaiah, U.; Pena, C.; Nylin, S.; Freitas, A.V.L.; Brower, A.V.Z. Nymphalid butterflies diversify following near demise at the Cretaceous/Tertiary boundary. Proc. Biol. Sci. 2009, 276, 4295–4302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH image to ImageJ: 25 years of image analysis. Focus Bioim. Inf. 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Beccaloni, G.W.; Hall, S.K.; Viloria, A.L.; Robinson, G.S. Catalogue of the hostplants of the Neotropical Butterflies/Catálogo de las plantas huésped de las mariposas Neotropicales. In RIBES-CYTED; Monografias, T.M., Zaragoza, S.E.A., Eds.; The Natural History Museum, Instituto Venezolano de Investigaciones Científicas: Zaragosa, Spain, 2008; Volume 8, 536p. [Google Scholar]
- Zanne, A.E.; Tank, D.C.; Cornwell, W.K.; Eastman, J.M.; Smith, A.S.; FitzJohn, R.G.; McGlinn, D.J.; O’Meara, B.C.; Moles, A.T.; Reich, P.B.; et al. Three keys to the radiation of angiosperms into freezing environments. Nature 2014, 506, 89–92. [Google Scholar] [CrossRef]
- Blomberg, S.P.; Garland, T.; Ives, A.R. Testing for phylogenetic signal in comparative data: Behavioral traits are more labile. Intern. J. Org. Evol. 2013, 57, 717–745. [Google Scholar] [CrossRef] [PubMed]
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- Kembel, S.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef] [Green Version]
- Freire, G.D.B., Jr.; Ribeiro, D.B.; Santos, A.C.; Silva, T.; Dias, J.P.; Rodrigues, H.P.; Diniz, I.R. Horizontal and vertical variation in the structure of fruit-feeding butterfly (Nymphalidae) assemblages in the Brazilian Cerrado. Insect Conserv. 2021, 14, 1–10. [Google Scholar] [CrossRef]


| Traits | Blomberg’s K | p-Value | |
|---|---|---|---|
| Body size | 1.1 | 0.001 | *** |
| Abundance | 0.39 | 0.27 | ns |
| Dispersal level | 0.76 | 0.07 | ns |
| Dispersal distance | 0.53 | 0.53 | ns |
| Visited traps | 0.39 | 0.66 | ns |
| Permanence (days) | 0.39 | 0.78 | ns |
| Diet breadth (Family of plants) | 1.08 | 0.001 | *** |
| Diet breadth (Genera of plants) | 0.84 | 0.002 | ** |
| Diet breadth (Species of plants) | 0.79 | 0.01 | ** |
| Phylogenetic Diet breadth | 0.48 | 0.06 | ns |
| Traits | F | Beta | p-Value | |
|---|---|---|---|---|
| Abundance | 7.2 | −0.09 | 0.01 | ** |
| Dispersal level | 9.69 | 1.17 | 0.01 | ** |
| Dispersal distance | 9.09 | 0.22 | 0.01 | ** |
| Visited traps | 9.69 | 0.13 | 0.01 | ** |
| Permanence (days) | 4.64 | 0.13 | 0.04 | * |
| Diet Breadth (Family of plants) | 16.23 | 3.8 | 0.001 | *** |
| Diet Breadth (Genera of plants) | 4.64 | 0.79 | 0.04 | * |
| Diet Breadth (Species of plants) | 1.83 | −0.03 | 0.81 | ns |
| Phylogenetic Diet Breadth | 4.83 | 0.11 | 0.001 | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Freire, G.B., Jr.; Silva, T.; Oliveira, H.; Collier, C.; Rodrigues, H.P.; Dias, J.P.; Santos, J.P.; Marini-Filho, O.J.; Freitas, A.V.L.; Smilanich, A.M.; et al. Good Things Come in Larger Packages: Size Matters for Adult Fruit-Feeding Butterfly Dispersal and Larval Diet Breadth. Diversity 2021, 13, 664. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120664
Freire GB Jr., Silva T, Oliveira H, Collier C, Rodrigues HP, Dias JP, Santos JP, Marini-Filho OJ, Freitas AVL, Smilanich AM, et al. Good Things Come in Larger Packages: Size Matters for Adult Fruit-Feeding Butterfly Dispersal and Larval Diet Breadth. Diversity. 2021; 13(12):664. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120664
Chicago/Turabian StyleFreire, Geraldo B., Jr., Thayane Silva, Hernani Oliveira, Chloe Collier, Hanna P. Rodrigues, Joao P. Dias, Jessie P. Santos, Onildo J. Marini-Filho, André V. L. Freitas, Angela M. Smilanich, and et al. 2021. "Good Things Come in Larger Packages: Size Matters for Adult Fruit-Feeding Butterfly Dispersal and Larval Diet Breadth" Diversity 13, no. 12: 664. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120664