
Appendicularia (Tunicata) in an Antarctic Glacial Fjord–Chaotic Fjordic Structure Community or Good Indicators of Oceanic Water Masses?


1
Department of Marine Plankton Research, Faculty of Oceanography and Geography, University of Gdańsk, Al. Marszałka J. Piłsudskiego 46, 81-378 Gdynia, Poland
2
Department of Aquatic Ecology, Maritime Institute, Gdynia Maritime University, Al. Grunwaldzka 311, 80-309 Gdańsk, Poland
*
Author to whom correspondence should be addressed.
Diversity 2021, 13(12), 675; https://0-doi-org.brum.beds.ac.uk/10.3390/d13120675
Submission received: 28 October 2021
/
Revised: 9 December 2021
/
Accepted: 14 December 2021
/
Published: 16 December 2021
(This article belongs to the Special Issue Linking Plankton Diversity with Ecosystem Functioning and Services)
Abstract
:Appendicularians are important but remain poorly studied groups of zooplankton in polar regions. The present research is based on samples collected in Admiralty Bay (King George Island) during a year-long period. Six larvacean species were noted, among which Fritillaria borealis and Oikopleura gaussica were found to be the most numerous, while the other species were relatively rare. Fritillaria borealis was a dominant part of the late summer (warm water) community, while O. gaussica had the highest presence in the winter (cold water) community. The abundance of appendicularians recorded in the bay was less numerous than that described by other authors. The most important factors influencing annual changes in the larvaceans in the bay was season, but only in the case of the two species. These facts were probably linked to the very dynamic changes in the abiotic conditions in the fjord, and the influx of specific masses of water.
1. Introduction
Appendicularians in some marine regions can be considered as one of the most important groups of epipelagic mesozooplankton immediately after copepods [1,2]. However, they play a different role in the marine food web [3], being an important group of marine pelagic filter feeders, especially in regard to very small organic particles [4,5]. They are producers of large sinking aggregates thanks to their extremely time-effective ability of building filter structures, “houses” [6,7,8], which can be considered as an important fraction of vertical carbon flux in the water column [9,10]. Appendicularians can also be regarded as an important factor in the diet of some economically important fishes, e.g., the anchovy Engraulis anchoita; in springtime, adults feed on meso- and macrozooplanktons, including appendicularians [11]. Capitanio et al. [12,13] studied the stomach contents of anchovy adults collected at the La Plata and El Rincón estuaries and at the Valdés Peninsula tidal front and found great quantities of Oikopleura fusiformis and O. dioica appendicularians in their guts.
Larvaceans are not well studied by using in situ collection methods, e.g., plankton nets or bottles [14]. This is mainly due to their patchy distribution [15] and fragility, which makes them very difficult to identify [14,16] and leads to underestimations of their abundance and biomass [17,18]. In the Southern Ocean area, including the Antarctic, studies on larvacean taxonomy, distribution and abundance are very rare. Hunt and Hosie [19] recorded high appendicularian abundances in the seasonal ice zone (SIZ). Similar observations were made by Tsujimoto et al. [20]. Daponte and Esnal [21] validated the Antarctic species of the “Oikopleura gaussica group” in the regions near South Georgia and in the Bellingshausen Sea, and Kalarus and Panasiuk [16] researched the zonal distribution of these animals in the Drake Passage. Most other studies conducted in recent decades have examined entire zooplankton communities and have examined appendicularians only as a single group or by focusing on the dominant species [22,23,24,25]. In Antarctic or sub-Antarctic bays or fjords, studies on larvaceans are even rarer. Fuentes et al. [26] recorded Fritillaria borealis and Oikopleura gaussica in Potter Cove (King George Island), while Freire et al. [27] found F. borealis in Admiralty Bay (King George Island), and Potocka et al. [28] noted Appendicularia in Herve Cove (Admiralty Bay).
Gelatinous zooplankton, which includes appendicularians, may play an important role in the functioning of pelagic food webs and biogeochemical cycles due to their important role in the circulation of organic matter [8,16,29,30,31,32]. Increased melting of glaciers has been observed in Antarctic shelf regions in recent decades [28,33,34,35]. The current state of knowledge allows us to suggest that plankton communities in fjordic and shelf areas such as Admiralty Bay will react rapidly to environmental changes. This was previously confirmed by the results of research conducted by, e.g., Garcia et al. [36,37]. Hydrological conditions in Admiralty Bay are the result of many factors: variability in the inflow of waters and their characteristics from the Bransfield Strait, tidal forces, wind, and the influence of glaciers. Based on a work by Antezana [38], who researched plankton in southern Chilean fjords, we may assume that plankton communities in this type of fjord area may be chaotic (can be influenced by many dynamically variable factors, in time and space), therefore, it may be difficult to determine both certain trends in the occurrence of plankton animals and the most important factors determining their presence and abundance.
The main goal of our work was to thoroughly investigate the species composition and abundance of appendicularians in an Antarctic glacial fjord and their annual variability in relation to selected environmental factors. The present research can make a significant contribution to a more detailed understanding of some aspects of polar appendicularian ecology, offering a good source of data on their species diversity and abundance, especially in this period of changing climatic conditions in Antarctic ecosystems.
2. Material and Methods
2.1. Study Area
The South Shetland Island archipelago is located in West Antarctica, north of the Antarctic Peninsula. The archipelago is separated from the Antarctic Peninsula by the narrow Bransfield Strait and from South America by the Drake Passage. Admiralty Bay (King George Island, South Shetland Islands) is one of the largest bays in the entire archipelago, with a coastline length of 83.4 km [39]. The bay is a fjord opening to the south toward the Bransfield Strait. The central part of the bay has a depth of 400–500 m (maximum 530 m) [40]. The northern part of Admiralty Bay branches into three smaller fjords: Ezcurra, Mackellar and Martel [40]. The hydrological conditions of Admiralty Bay result from the influence of Bransfield Strait waters, as well as from the inflow of freshwater from the land and local fjord processes [41]. Tides are an important factor generating the inflow and mixing of water masses in the bay [42]. A complete exchange of surface waters (up to 100 m deep) takes from one to two weeks [43]. Glaciers have the greatest cooling effect on bay waters: in the immediate vicinity of meltwater, salinity may decrease to as low as 20 [44]. Because a significant part of the coast of Admiralty Bay (46%) is covered with ice forms [42], reduced salinity and increased concentrations of suspended solids are often observed in this zone. The latter is almost five times higher than that recorded in the open waters of the Antarctic [45]. The western Antarctic Peninsula region, including the South Shetland Islands, is impacted by warmer and saltier waters from the Antarctic Circumpolar Current (ACC) [46]. One of these water masses includes Upper Circumpolar Deep Water, which is typical for the Antarctic Shelf, with a temperature between 1.70 and 2.13 °C and salinity between 34.5 and 34.75 [47,48]. The physical environment of the shelf also includes typically cold water from the Weddell Sea [49].
2.2. Sample Collection and Laboratory Studies
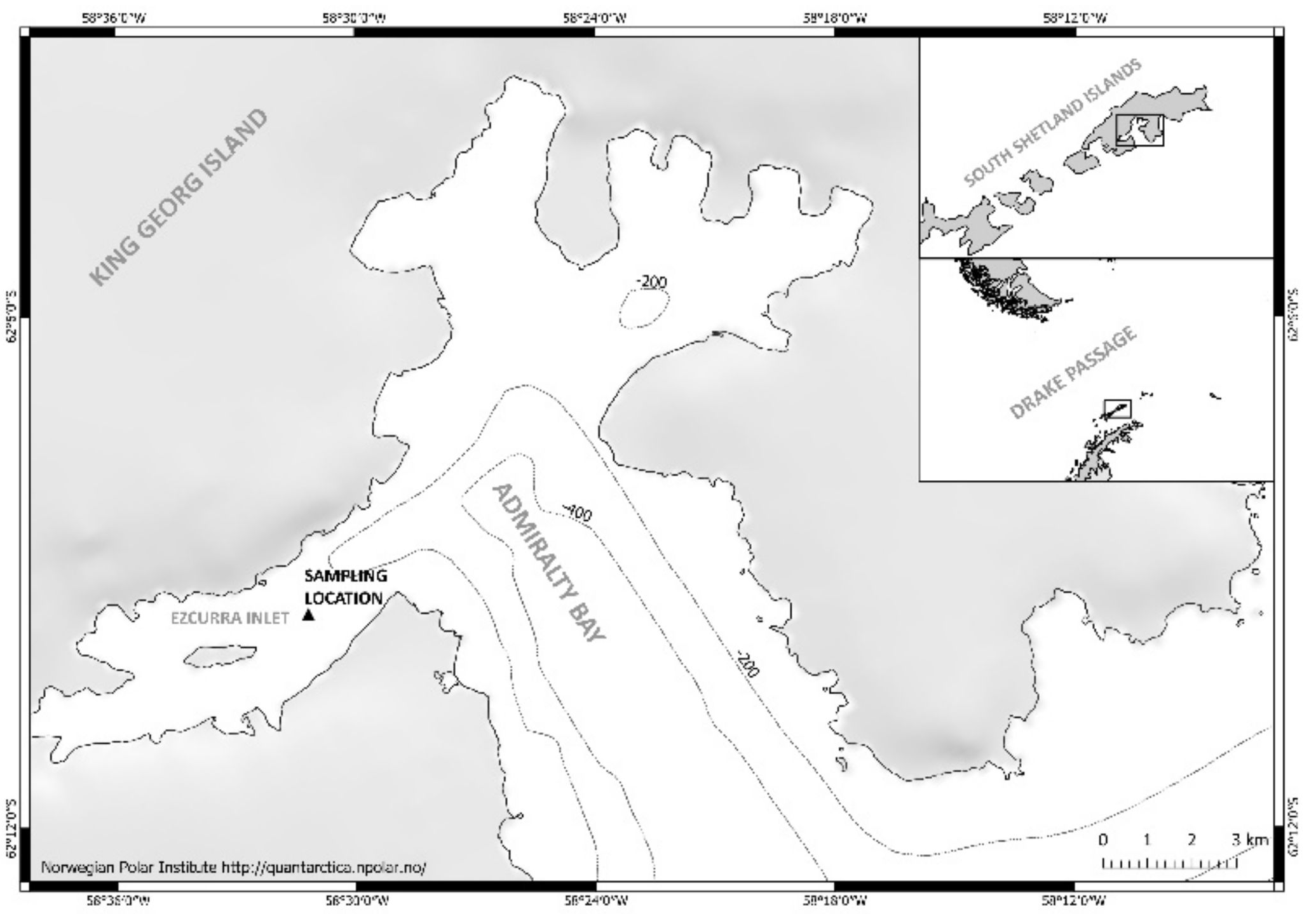
Plankton samples used in this study were collected during the 28th (2003–2004) Antarctic Expedition to the Polish Henryk Arctowski Antarctic Station (King George Island, South Shetland Islands). Favorable ice conditions in Admiralty Bay (no icing) allowed for continuous sampling for 11 months, i.e., from December 2003 to November 2004. Zooplankton material was collected twice per month at approximately 10-day intervals, from a single research station located in the Ezcurra Inlet, in the western part of Admiralty Bay (62°08′90″ S, 58°29′40″ W). Samples were collected from two depth layers—50–0 m and 100–50 m (Figure 1)—using WP2 net with a mesh size of 200 µm. At the same time, CTD probe was used to record temperature and salinity. The research material—a total of 96 samples—was used to determine the species composition and abundance of appendicularians in Admiralty Bay. Taxonomic identification was performed to the lowest possible taxonomic level based on Esnal [50,51], Fenaux [52] and O’Sullivan [53], specimens in samples were counted, and the obtained numbers were calculated into m−3 based on the volume of filtered water.
2.3. Statistical Analyses
Appendicularian abundance was expressed as the number of individuals per 103 m−3 assuming 100% efficiency of the net. Multivariate statistical analyses of appendicularian community abundance data were carried out using the PRIMER 7 software package [54,55] and RStudio using R version 4.0.3 [56]. Multivariate statistical analyses were performed using the vegan package version 2.5–7 in R [57]. Square root transformation was used for the pretreatment of the obtained abundance data. Similarities between samples were examined using the Bray–Curtis index [58] (vegdist function). ANOSIM (analysis of similarities) was used to check differences between specific groups of appendicularians. Nonmetric multidimensional scaling (NMDS, function metaMDS) was carried out with dissimilarity matrices calculated for the appendicularian abundances and selected environmental factors. The NMDS plot was made using environmental vectors representing temperature, and salinity (functions envfit and ordisurf). We used generalized additive models and ordisurf function to check how the variability of temperature can affect Appendicularia communities. Ordisurf function plots fit environmental variables on ordination diagram. Permutational multivariate analysis of variance (PERMANOVA) was performed to identify the most important environmental factors among temperature, salinity, depth and season for larvacean abundance. Interpretation of differences and similarities between the groups, as well as identification of the species responsible for group differences, were carried out using the SIMPER procedure.
3. Results
3.1. Hydrological Conditions
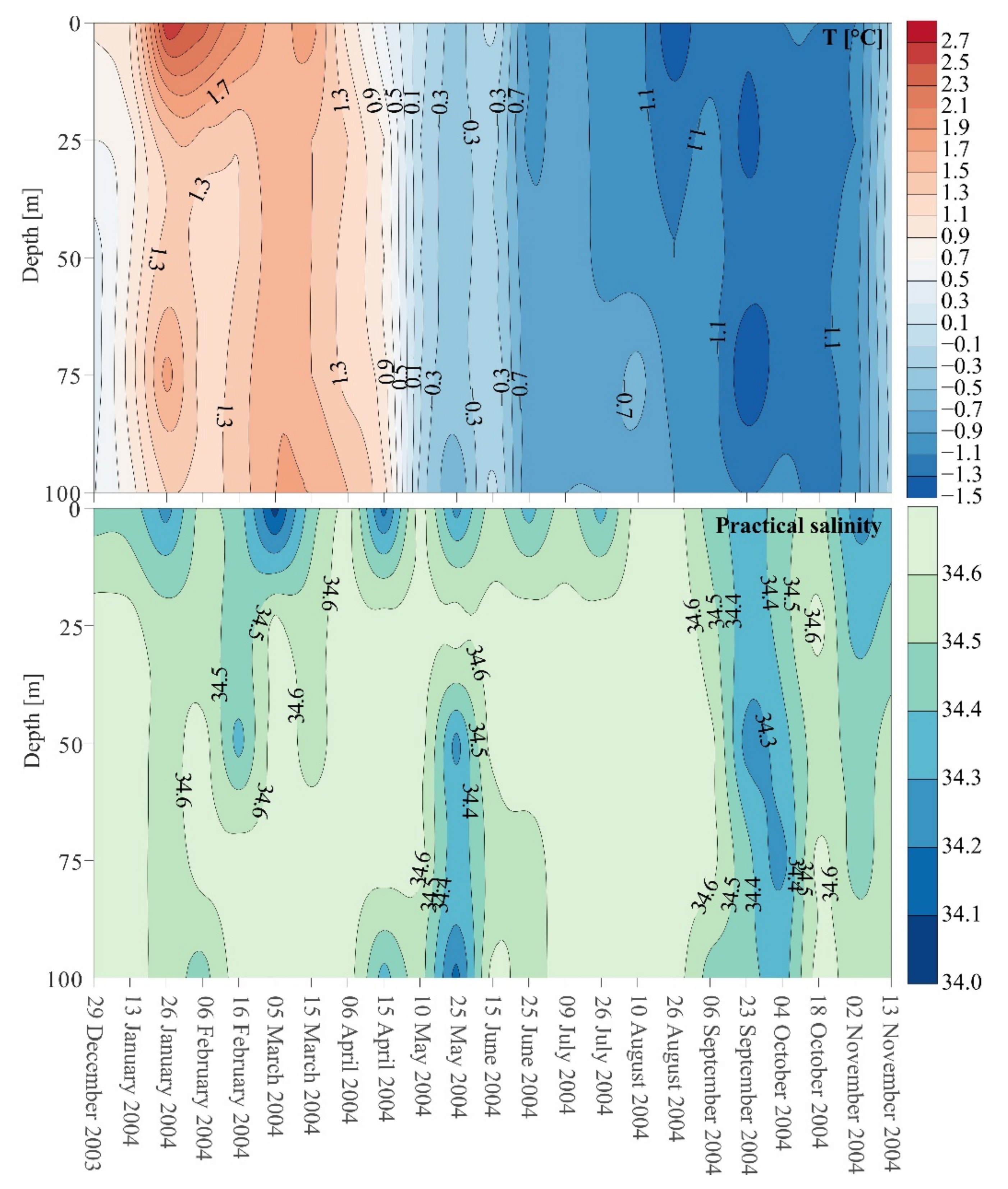
The highest temperature (+2.7 °C) was recorded at the end of January at a depth of 1 m, and the lowest temperature (−1.5 °C) was recorded at the end of August, also at a depth of 1 m (Figure 2). The change from Antarctic summer to winter occurred from April to May, when temperature decreased from +0.5 °C to −0.9 °C in the surface water layer and from +1.2 °C to −0.9 °C at a depth of 100 m (Figure 2). During Antarctic summer (December–April), the temperature of the upper water layer changed from +2.7 °C in January to +0.5 °C in April. In May–June, isothermal conditions were observed in the investigated water column (Figure 2). The salinity of the waters of Admiralty Bay varied slightly, from 34.0 to a maximum value of 34.6 (Figure 2). The greatest variability of this parameter was recorded near the surface, where it reached a minimum value of 34.0 (Figure 2) in March. A slight decrease in salinity in the entire water column, i.e., from 34.2 to 34.4 (Figure 2), was recorded in September and October. Based on hydrological data the research period was divided into seasons, summer (S) lasting from December to the end of March, transition period (TP) from April to May, and winter (W) from June to November.
3.2. Appendicularian Diversity and Abundance—Annual Changes
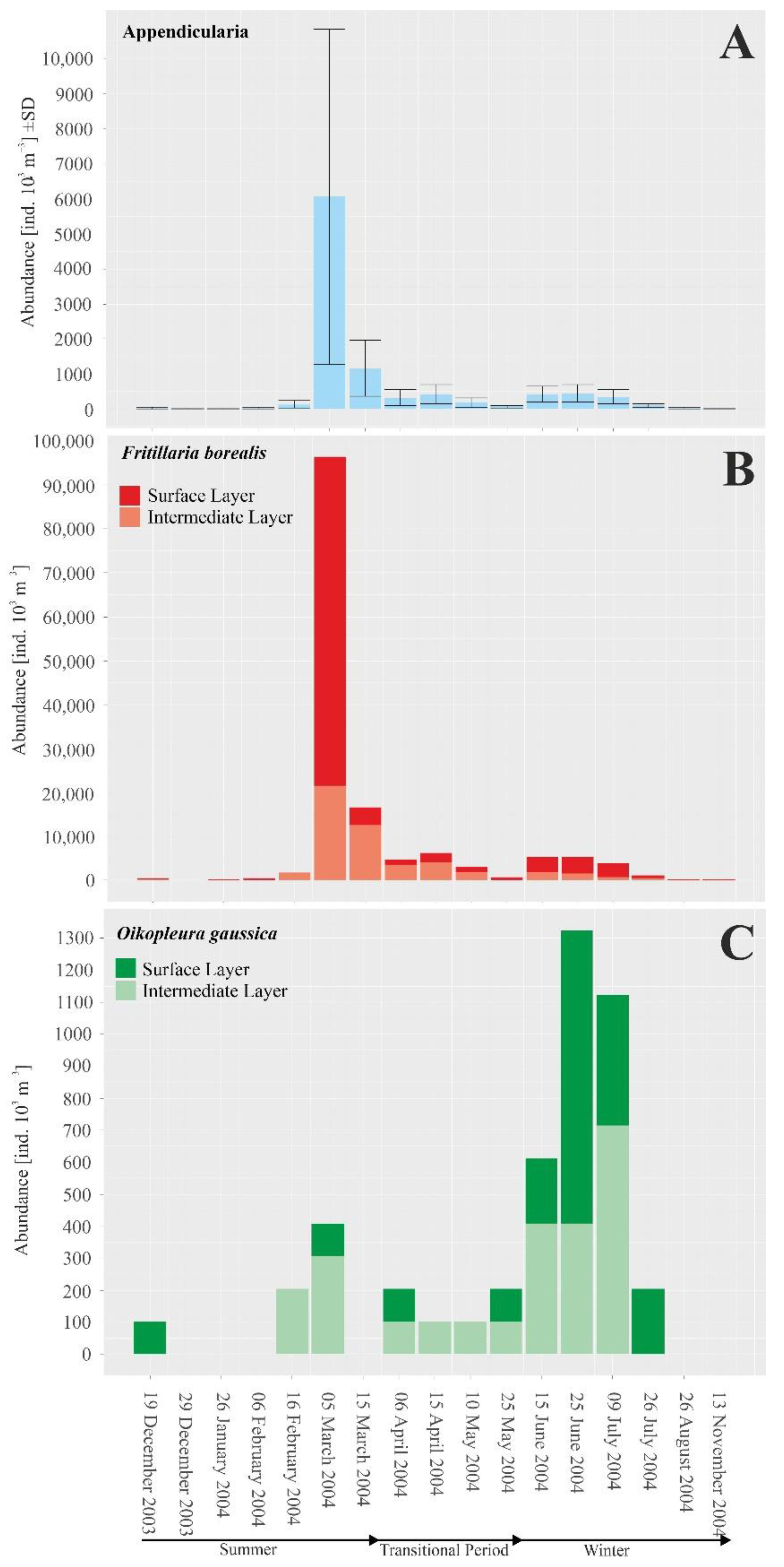
In Admiralty Bay, six species of Appendicularia were found: three from the genus Fritillaria, namely, F. borealis typica, F. antarctica and F. aberrans, and three from the genus Oikopleura, i.e., O. fusiformis, O. gaussica and O. parva. The highest density of larvaceans was recorded from March to July (late summer–early winter), with the maximum abundance observed in March (more than 40,000 ind. 103 m−3) (Table 1, Figure 3).
Throughout the research period, Fritillaria borealis was the most abundant species, and maximum density values were observed at the end of the Antarctic summer in March (Figure 3). During wintertime, the abundance of this species decreased, but individuals were still observed until September (Figure 3). The second most numerous species was Oikopleura gaussica, whose density peak was recorded at the beginning of winter (June–July) (Figure 3). The highest recorded mean abundance in winter was approximately 250 ind. 103 m−3 (maximum abundance more than 1000 ind. 103 m−3 was recorded in June) (Table 1, Figure 3). Other species were recorded sporadically, and their mean numbers reached no more than 50 ind. 103 m−3 (Table 1).
3.3. Environmental Factors Influencing the Annual Changes in the Abundance of Appendicularia
The results of the ANOSIM analysis showed statistically significant differences (p < 0.05) between samples collected in different seasons (summer, transition period, and winter). The SIMPER analysis indicated that the species that determined most similarity between the samples was Fritillaria borealis (Table 2). This larvacean accounted for over 80% of the similarity between the samples collected in the summer and winter periods (Table 2). During the winter season, a noticeable increase in the importance of Oikopleura gaussica was observed (Table 2).
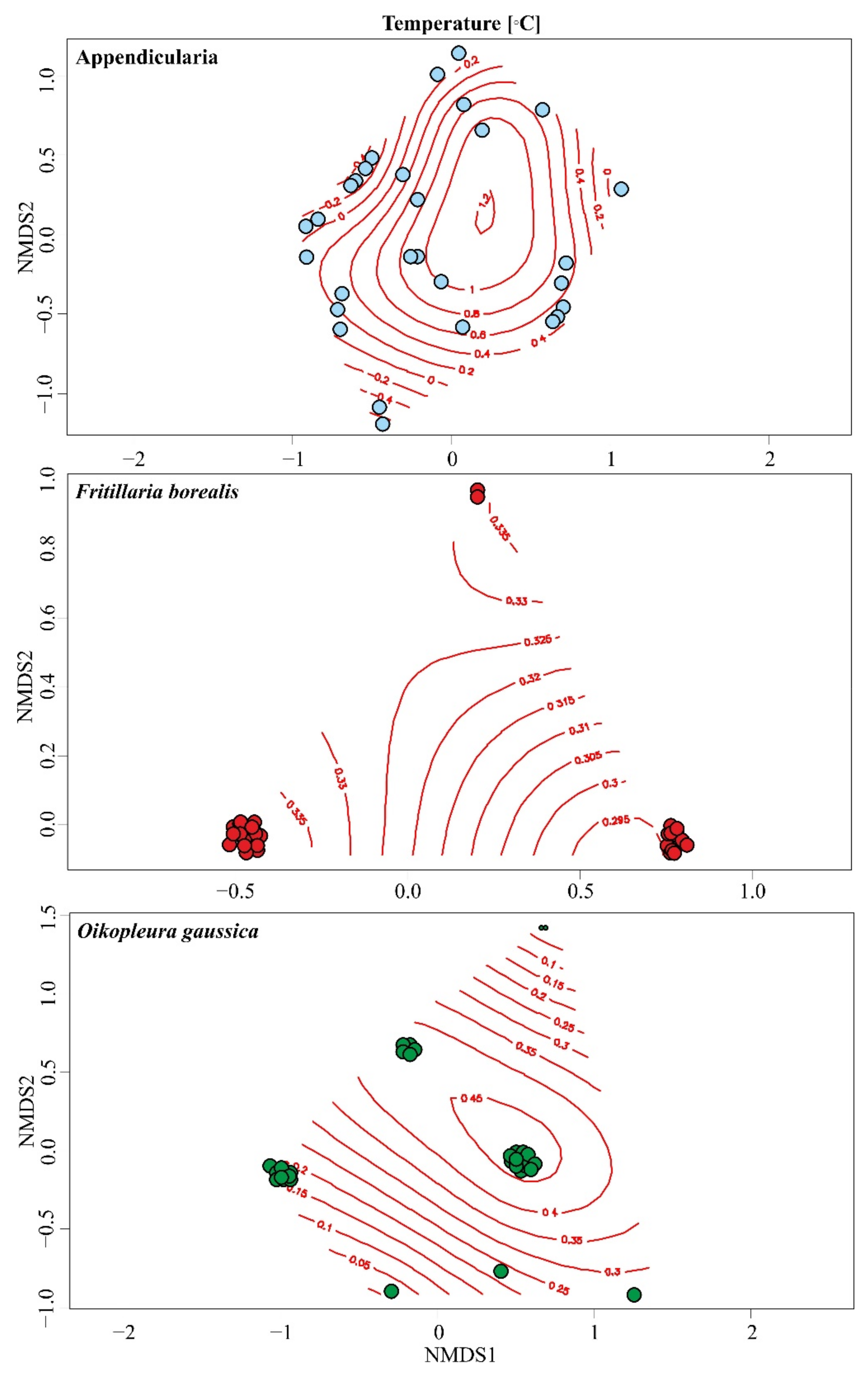
The influence of environmental factors on the abundance of larvaceans from Admiralty Bay was investigated using the Permanova test. The results showed that the factors that had the greatest influence on the observed appendicularian variability were temperature (T) and season (Table 3). Our studies showed that only Fritillaria borealis correlated with temperature and season variables. In the case of the remaining analyzed taxa, the studied environmental factors were not statistically significant. The ordisurf models for temperature showed that Appendicularia were most abundant at temperatures above 0.2 °C or slightly higher (Figure 4). Fritillaria borealis preferred slightly higher temperatures, above 0.3 °C, while Oikopleura gaussica was most numerous in water with a temperature above 0 °C (Figure 4). The ordisurf models for salinity showed that in the case of all investigated species, a value above 35.5 was the most favorable (Figure 4).
4. Discussion
Appendicularians are a very important component of the mesozooplankton, they also play an important role in the production of “sea snow”, and thus they can be an important link in the vertical carbon flux [59]. In addition, larvaceans can play a crucial role in the diet of copepods, fish or jellyfish [60]. Nonetheless, these animals are still very poorly studied, especially in polar regions, which is often due to the use of inadequate plankton nets, as well as difficulties in taxonomic identification [16,61]. Considering the above facts, the results of the research presented in this paper may offer a significant contribution to a better understanding of this group of animals in polar fjord environments, where the effects of ongoing climate change are most clearly and readily visible [28,35,62].
4.1. Appendicularians Diversity and Abundance
During our research, we found Fritillaria borealis and Oikopleura gaussica to be the most numerous species in the investigated area. These two species were also previously recorded in the bay by Freire et al. [27]. Other species, i.e., F. antarctica, F. aberrans, O. fusiformis and O. parva had not been previously noted in the investigated region or in the entire area of the Antarctic Peninsula. However, we observed the presence of these species in the Drake Passage [16]. The larvacean abundance recorded in this study was higher than that found by Freire et al. [27], i.e., 1200 ind. 102 m−3 (March 1989) in the central part of Admiralty Bay, and only 2 ind. 102 m−3 in the Ezcurra Inlet. Freire et al. [27] used nets with a relatively large mesh size, i.e., 384 µm, which probably induced an underestimation of the number of individuals. The values obtained by Kittel et al. [63] were also much lower than the ones recorded in the present study. Generally, high differences obtained by several authors in the same area of research can be partially explained by large spatial and periodic fluctuations in the abundance of larvaceans, which may be presumably conditioned by the hydrological specificity of the waters [42,63]. However, the differences in abundances obtained during the previous studies could be a consequence of use of nets with different mesh sizes. It should also be mentioned that the most important factor determining the occurrence of larvaceans is the availability of food, i.e., the composition and density of phytoplankton (pico- and nanoplankton) [64]. Studies conducted by other researchers in the bay indicated a relatively low phytoplankton biomass within the investigated area, with high values of suspended organic matter that limit the range of the euphotic zone [65,66,67]. However, while the mean abundance of F. borealis recorded in the bay was higher than that in the Drake Passage, it was relatively similar to that reported for the Antarctic Zone [16].
The results obtained during our studies indicated seasonal changes in the qualitative and quantitative composition of appendicularians in Admiralty Bay, including the presence of species succession, i.e., Fritillaria borealis (summer/warmer water masses) → Oikopleura gaussica (winter/colder water masses). It should be emphasized, however, that in the case of Fritillaria species, we observed one large abundance peak in March 2004. In our opinion, this was probably the result of an intrusion of warmer water masses, as these animals were later observed in the bay but in much smaller numbers. In March very high temperatures (>1.5 °C) were recorded in the bay, which might have been related to the periodic advection of circumpolar deep water [48]. During wintertime, the influence of Weddell Sea waters increases, which may be the source of the high O. gaussica abundance in the bay.
4.2. Environmental Factors Influencing the Abundance of Appendicularia
In fact, with some exceptions, it was very difficult to determine the environmental factors (statistically significant) conditioning the structure of the community of the studied group of animals. For most species that we studied, their presence in the bay as well as their density were not related to environmental factors. However, based on our results we can conclude that seasonality had the most significant impact on the community structure of larvaceans in the bay, while season and salinity (analyzed independently) may have some effect on the occurrence of rare species. This observation may suggest a strong relationship between the occurrence of these animals and the dynamics of bay water masses, primarily—as mentioned above—the inflow of waters from the surrounding areas, probably from the Bellingshausen Sea during a summer period and from the Weddell Sea during a wintertime.
4.3. Appendicularia (Tunicata) in an Antarctic Glacial Fjord vs. Open Oceanic Waters
In Admiralty Bay, we found six species of larvaceans, and two of them, Fritillaria borealis and Oikopleura gaussica, were the most numerous. Kalarus and Panasiuk [16], in the Drake Passage in 2010, found eight species and the most abundant were F. borealis, F. fraudax, O. fusiformis and O. gaussica. In the Drake Passage, F. borealis was a common species in the entire research area, while O. gaussica was a typical species in the polar front and in the polar front zone areas [16]. The results of Lindsay and Williams [68], who conducted research in open oceanic waters in East Antarctica, showed that Oikopleura species had a more northern occurrence than Fritillaria species. Based on Esnal’s [51] and our results, we can state that F. borealis is very abundant in all Antarctic waters; however, it is probably related with the warmer water mass community, and O. gaussica is a typical species that occurs from 50° S to the south in relatively colder waters. In Admiralty Bay we did not record F. pellucida and F. fraudax specimens (these species were previously noted by us in the Drake Passage), which is probably related to the northern distribution of both species [50].
5. Conclusions
(1) In Admiralty Bay six species of Appendicularia were found, which indicates a much higher species diversity than previously recorded. Fritillaria borealis and Oikopleura gaussica were most numerous. However, the former was a significant part of the warm water mass community of this group of animals, while O. gaussica was an indicator of the colder water mass community.
(2) The dynamics of larvacean species observed in Admiralty Bay suggest the presence of succession, probably with relation to the inflow of specific water masses from the Bransfield Strait (e.g., summer–Bellingshausen Sea–warmer waters, winter–Weddell Sea–colder waters).
Author Contributions
A.P. was responsible for organizing the dataset, statistical analysis and for the management and preparation of the manuscript. M.K. managed the laboratory analysis, dataset preparation, and results analysis and interpretation. Moreover, he was also responsible for the management and preparation of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by research grant KBN No. PBZ-KBN-108/P04/2004 and internal funds of the The Department of Antarctic Biology, Polish Academy of Sciences and The Department of Marine Plankton Research, Faculty of Oceanography and Geography, University of Gdańsk (Poland).
Institutional Review Board Statement
This article does not contain any studies with human partic-ipants or animals performed by any of the authors. This is an observational study and no ethical approval is required.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The datasets analyzed during the current study are available from the corresponding author on reasonable request.
Acknowledgments
We would like to thank Ryszard Kuczyński, MSc and the crew of the Henryk Arctowski Polish Antarctic Station for their assistance in collecting samples. In particular, we would like to thank Maria Iwona Żmijewska for her invaluable help and supervision of our research. The zooplankton samples and data used in the paper were collected at the Henryk Arctowski Polish Antarctic Station.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Purcell, J.E. Predation by the hydromedusa Aequorea victoria on fish larvae and eggs at a herring spawning ground in British Columbia. Can. J. Fish. Aquat. Sci. 1989, 46, 1415–1427. [Google Scholar] [CrossRef]
- Purcell, J.E.; Sturdevant, M.V. Prey selection and dietary overlap among zooplanktivorous jellyfish and juvenile fishes in Prince William Sound, Alaska. Mar. Ecol. Prog. Ser. 2001, 210, 67–83. [Google Scholar] [CrossRef] [Green Version]
- Di Mauro, R.; Capitanio, F.; Viñas, M.D. Capture efficiency for small dominant mesozooplankters (Copepoda, Appendicularia) off Buenos Aires Province (34° S–41° S), Argentine Sea, using two plankton mesh sizes. Braz. J. Oceanogr. 2009, 57, 205–214. [Google Scholar] [CrossRef]
- Landry, M.R.; Peterson, W.K.; Fagerness, V.L. Mesozooplankton grazing in the Southern California Bight. I. Population abundances and gut pigment contents. Mar. Ecol. Prog. Ser. 1994, 115, 55–71. [Google Scholar] [CrossRef]
- Fernández, D.; López-Urrutia, A.; Fernández, A.; Acuña, J.L.; Harris, R. Retention efficiency of 0.2 to 6 μm particles by the appendicularians Oikopleura dioica and Fritillaria borealis. Mar. Ecol. Prog. Ser. 2004, 266, 89–101. [Google Scholar] [CrossRef] [Green Version]
- López-Urrutia, A.; Acuña, J.L. Gut throughput dynamics in the appendicularian Oikopleura dioica. Mar. Ecol. Prog. Ser. 1999, 191, 195–205. [Google Scholar] [CrossRef]
- Alldredge, A.L. The contribution of discarded appendicularian houses to the flux of particulate organic carbon from oceanic surface waters. In Response of Marine Ecosystems to Global Change: Ecological Impact of Appendicularians; Gorsky, G., Youngbluth, M.J., Deibel, D., Eds.; GB Scientific Publisher: Paris, France, 2005; pp. 309–326. [Google Scholar]
- Winder, M.; Bouquet, J.-M.; Bermúdez, J.R.; Berger, S.A.; Hansen, T.; Brandes, J.; Sazhin, A.F.; Nejstgaard, J.C.; Båmstedt, U.; Jakobsen, H.H.; et al. Increased appendicularian zooplankton alter carbon cycling under warmer more acidified ocean conditions. Limnol. Oceanogr. 2017, 62, 1541–1551. [Google Scholar] [CrossRef]
- Vargas, C.A.; Tönnesson, K.; Sell, A.; Maar, M.; Friis Møller, E.; Zervoudaki, T.; Giannakourou, A.; Christou, E.; Satapoomin, S.; Petersen, J.K.; et al. Importance of copepods versus appendicularians in vertical carbon fluxes in a Swedish fjord. Mar. Ecol. Prog. Ser. 2002, 241, 125–138. [Google Scholar] [CrossRef] [Green Version]
- Robison, B.H.; Reisenbichler, K.R.; Sherlock, R.E. Giant larvacean houses: Rapid carbon transport to the deep seafloor. Science 2005, 308, 1609–1611. [Google Scholar] [CrossRef]
- Capitanio, F.L.; Spinelli, M.L.; Presta, M.L.; Aguirre, G.E.; Cervetto, G.; Pájaro, M.; Derisio, C.M. Ecological Role of Common Appendicularian Species from Shelf Waters Off Argentina. In Plankton Ecology of the Southwestern Atlantic; Hoffmeyer, M.S., Sabatini, M.E., Brandini, F.P., Calliari, D.L., Santinelli, N.H., Eds.; Springer International Publishing AG, Springer Nature: Cham, Switzerland, 2018; pp. 201–218. [Google Scholar]
- Capitanio, F.; Pájaro, M.; Esnal, G.B. Appendicularians (Chordata, Tunicata) in the diet of anchovy Engraulis anchoita in the Argentine sea. Sci. Mar. 1997, 61, 9–15. [Google Scholar]
- Capitanio, F.; Pájaro, M.; Esnal, G.B. Appendicularians: An important food supply for the Argentine anchovy Engraulis anchoita in coastal waters. J. Appl. Ichthyol. 2005, 21, 414–419. [Google Scholar] [CrossRef]
- Lombard, F.; Legendre, L.; Picheral, M.; Sciandra, A.; Gorsky, G. Prediction of ecological niches and carbon export by appendicularians using a new multispecies ecophysiological model. Mar. Ecol. Prog. Ser. 2010, 98, 109–125. [Google Scholar] [CrossRef]
- Fenaux, R. Anatomy and functional morphology of the Appendicularia. In The Biology of Pelagic Tunicates; Bone., Q., Ed.; Oxford University Press: New York, NY, USA, 1998; pp. 25–34. [Google Scholar]
- Kalarus, M.; Panasiuk, A. Spatial Distribution Patterns of Appendicularians in the Drake Passage: Potential Indicators of Water Masses? Diversity 2021, 13, 286. [Google Scholar] [CrossRef]
- Hopcroft, R.R.; Roff, J.C.; Chavez, F. Size paradigms in copepod communities: A re-examination. Hydrobiologia 2001, 453, 133–141. [Google Scholar] [CrossRef]
- Harris, R.P.; Lopez-Urrutia, A.; Acuna, J.L.; Båmstedt, U.; Flood, P.R.; Fhyn, H.J.; Gorsky, G.; Youngbluth, M.J.; Irigoien, I.; Martinussen, M. A comparison of appendicularian seasonal cycles in four distinct European coastal environments. In Response of Marine Ecosystems to Global Change: Ecological Impact of Appendicularians; Gorsky, G., Youngbluth, M.J., Deibel, D., Eds.; Editions des Archives Contemporaires; GB Scientific Publisher: Paris, France, 2005; pp. 255–276. [Google Scholar]
- Hunt, B.P.V.; Hosie, G.W. Zonal structure of zooplankton communities in the Southern Ocean South of Australia: Results from a 2150 km continuous plankton recorder transect. Deep-Sea Res. Part I 2005, 52, 1241–1271. [Google Scholar] [CrossRef]
- Tsujimoto, M.; Takahashi, K.T.; Hirawake, T.; Mitsuo, F. Unusual abundance of appendicularians in the seasonal ice zone (140° E) of the Southern Ocean. Polar Biosc. 2006, 19, 133–141. [Google Scholar]
- Daponte, M.; Esnal, G. The classification of Antarctic appendicularians: The Oikopleura gaussica group. Ant. Sci. 2003, 15, 476–482. [Google Scholar]
- Hosie, G.W.; Schultz, M.B.; Kitchener, J.A.; Cochran, T.G.; Richards, K. Macrozooplankton community structure off East Antarctica (80–150° E) during the Austral summer of 1995/1996. Deep Sea Res. Part II 2000, 47, 2437–2463. [Google Scholar] [CrossRef]
- Hosie, G.W.; Fukuchi, M.; Kawaguchi, S. Development of the Southern Ocean Continuous Plankton Recorder survey. Prog. Oceanogr. 2003, 58, 263–283. [Google Scholar] [CrossRef]
- Takahashi, K.T.; Hosie, G.W.; Kitchener, J.A.; McLeod, D.J.; Odate, T.; Fukuchi, M. Comparison of zooplankton distribution patterns between four seasons in the Indian Ocean sector of the Southern Ocean. Polar Sci. 2010, 4, 317–331. [Google Scholar] [CrossRef] [Green Version]
- Ojima, M.; Takahashi, K.T.; Tanimura, A.; Odate, T.; Fukuchi, M. Spatial distribution of micro- and meso-zooplankton in the seasonal ice zone of east Antarctica during 1983–1995. Polar Sci. 2015, 9, 319–326. [Google Scholar] [CrossRef]
- Fuentes, V.L.; Schnack-Schiel, S.B.; Schloss, I.R.; Esnal, G.G. Mesozooplankton of Potter Cove: Community composition and seasonal distribution in 2002 and 2003, In The Antarctic Ecosystem of Potter Cove, King-George Island (Isla 25 de Mayo), Synopsis of Research Performed 1999–2006 at the Dallmann Laboratory and Jubany Station; Wiencke, C., Ferreyra, G.A., Abele, D., Marenssi, S., Eds.; Reports on Polar and Marine Research; Alfred-Wegener-Institut Für Polar- und Meeresforschung: Bremerhaven, Germany, 2008; Volume 571, pp. 75–84. [Google Scholar]
- Freire, A.S.; Coelho, M.J.C.; Bonecker, S.L.C. Short term spatial—Temporal distribution patterns of zooplankton in Admiralty Bay (Antarctica). Polar Biol. 1993, 13, 433–439. [Google Scholar] [CrossRef]
- Potocka, M.; Kidawa, A.; Panasiuk, A.; Bielecka, L.; Wawrzynek-Borejko, J.; Patuła, W.; Wójcik, K.A.; Plenzler, J.; Janecki, T.; Bialik, R.J. The effect of glacier recession on benthic and pelagic communities: Case study in Herve Cove, Antarctica. J. Mar. Sci. Eng. 2019, 7, 285. [Google Scholar] [CrossRef] [Green Version]
- Mańko, M.K.; Panasiuk-Chodnicka, A.A.; Żmijewska, M.I. Pelagic coelenterates in the Atlantic sector of the Arctic Ocean: Species diversity and distribution as water mass indicators. Oceanol. Hydrobiol. Stud. 2015, 44, 466–479. [Google Scholar] [CrossRef]
- Słomska, A.W.; Panasiuk-Chodnicka, A.A.; Żmijewska, M.I.; Mańko, M.K. Variability of Salpa thompsoni population structure in the Drake Passage in summer season 2010. Pol. Polar Res. 2015, 36, 391–404. [Google Scholar] [CrossRef]
- La, H.S.; Park, K.; Wåhlin, A.; Arrigo, K.R.; Kim, D.S.; Yang, E.J.; Atkinson, A.; Fielding, S.; Im, J.; Kim, T.-W.; et al. Zooplankton and micronekton respond to climate fluctuations in the Amundsen Sea polynya, Antarctica. Sci. Rep. 2019, 9, 10087. [Google Scholar] [CrossRef]
- Panasiuk, A.; Grzonka, L.; Prątnicka, P.; Wawrzynek-Borejko, J.; Szymelfenig, M. Zonal variability of pelagic Siphonophora (Cnidaria) in the Atlantic sector of the southern ocean. J. Sea Res. 2020, 165, 101951. [Google Scholar] [CrossRef]
- Birkenmajer, K. Retreat of Ecology Glacier, Admiralty Bay. King George Island (South Shetland Islands, West Antarctica), 1956–2001. Bull. Pol. Acad. Sci. 2002, 50, 15–30. [Google Scholar]
- Simões, J.C.; Dani, N.; Bremer, U.F.; Aquino, F.E.; Arigony-Neto, J. Small cirque glaciers retreat on Keller Peninsula, Admiralty Bay, King George Island, Antarctica. Pesqui. Antart. Bras. 2004, 4, 49–56. [Google Scholar]
- Pudełko, R.; Angiel, P.J.; Potocki, M.; Jędrejek, A.; Kozak, M. Fluctuation of Glacial Retreat Rates in the Eastern Part of Warszawa Icefield, King George Island, Antarctica, 1979–2018. Remote Sens. 2018, 10, 892. [Google Scholar] [CrossRef] [Green Version]
- Garcia, M.D.; Fernández Severini, M.D.; Spetter, C.; López Abbatea, M.C.; Tartara, M.N.; Nahuelhual, E.G.; Marcovecchio, J.E.; Schloss, I.R.; Hoffmeyer, M.S. Effects of glacier melting on the planktonic communities of two Antarctic coastal areas (Potter Cove and Hope Bay) in summer. Reg. Stud. Mar. Sci. 2019, 30, 100731. [Google Scholar] [CrossRef]
- Garcia, M.D.; Dutto, M.S.; Chazarreta, C.J.; Berasategui, A.A.; Schloss, I.R.; Hoffmeyer, M.S. Micro-mesozooplankton successions in an Antarctic coastal environment during a warm year. PLoS ONE 2020, 15, e0232614. [Google Scholar] [CrossRef] [PubMed]
- Antezana, T. Plankton of Southern Chilean fjords: Trends and linkages. Sci. Mar. 1999, 63 (Suppl. S1), 69–80. [Google Scholar] [CrossRef]
- Rakusa-Suszczewski, S. The Maritime Antarctic Coastal Ecosystem of Admiralty Bay; Department of Antarctic Biology, Polish Academy of Sciences: Warsaw, Poland, 1993; pp. 1–216. [Google Scholar]
- Rakusa-Suszczewski, S. The hydrography of Admiralty Bay and its inlets, coves and lagoons (King George Island, Antarctica). Pol. Polar Res. 1995, 17, 29–42. [Google Scholar]
- Wright, F.F. Fiord circulation and sedimentation Southeast Alaska. In Proceedings of the First International Conference of Port Ocean Engineering Under Arctic Conditions, Trondheim, Norway, 23–30 August 1971; Volume 1, pp. 279–289. [Google Scholar]
- Rakusa-Suszczewski, S. The role of near shore research in gaining an understanding of the functioning of the Antarctic ecosystem. Pol. Arch. Hydrobiol. 1980, 27, 229–233. [Google Scholar]
- Tokarczyk, R. Classification of water masses in the Bransfield Strait and southern part of the Drake Passage using a method of statistical multidimensional analysis. Pol. Polar Res. 1987, 8, 333–366. [Google Scholar]
- Szafrański, Z.; Lipski, M. Characteristics of water temperature and salinity at Admiralty Bay (King George Island, South Shetlands Islands, Antarctic) during the austral summer 1978/1979. Pol. Polar Res. 1982, 3, 7–24. [Google Scholar]
- Pęcherzewski, K. Distribution and quantity of suspended matter in Admiralty Bay (king George Island, South Shetland Islands). Pol. Polar Res. 1980, 1, 75–82. [Google Scholar]
- Orsi, H.; Whitworth, T.; Nowlin, W.D., Jr. On the meridional extent and fronts of the Antarctic Circumpolar Current. Deep Sea Res. Part I 1995, 42, 641–673. [Google Scholar] [CrossRef]
- Martinson, D.G.; Stammerjohn, S.E.; Iannuzzi, R.A.; Smith, R.C.; Vernet, M. Western Antarctic Peninsula physical oceanography and spatio–temporal variability. Deep Sea Res. Part II Top. Stud. Oceanogr. 2008, 55, 1964–1987. [Google Scholar] [CrossRef]
- Cape, M.R.; Vernet, M.; Pettit, E.C.; Wellner, J.; Truffer, M.; Akie, G.; Domack, E.; Leventer, A.; Smith, C.R.; Huber, B.A. Circumpolar Deep Water Impacts Glacial Meltwater Export and Coastal Biogeochemical Cycling Along the West Antarctic Peninsula. Front. Mar. Sci. 2019, 6, 144. [Google Scholar] [CrossRef]
- Moffat, C.; Meredith, M. Shelf–ocean exchange and hydrography west of the Antarctic Peninsula: A review. Philos. Trans. A Math. Phys. Eng. Sci. 2018, 376, 20170164. [Google Scholar] [CrossRef]
- Esnal, G. Apendicularia. In Atlas del Zooplancton del Atlántico Sudoccidental y Métodosde Trabajo con el Zooplancton Marino; Boltovskoy, D., Ed.; Publicación Especial, Instituto Nacional de Investigacióny Desarrollo Pesquero: Mar del Plata, Argentina, 1981; pp. 809–820. [Google Scholar]
- Esnal, G. Appendicularia. In South Atlantic Zooplankton; Boltovskoy, D., Ed.; Backhuys Publication: Leiden, The Netherlands, 1999; pp. 1375–1399. [Google Scholar]
- Fenaux, R. The Classification of Appendicularia (Tunicata): History and Current State. Mem. Inst. Oceanogr. 1993, 17, 1–123. [Google Scholar]
- O’Sullivan, D. A Guide to the Pelagic Tunicates of the Southern Ocean and Adjacent Waters; ANARE Research Notes; Antarctic Division: Kingston, Australia, 1983.
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 2nd ed.; PRIMER-E: Plymouth, UK, 2001; pp. 1–176. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2006; pp. 1–190. [Google Scholar]
- Vanables, W.N.; Smith, D.M.; R Core Team. An Introduction to R, Notes on R: A Programming Environment for Data Analysis and GraphicsVersion 4.1.0. Available online: https://cran.r-project.org/doc/manuals/r-release/R-intro.pdf (accessed on 10 November 2021).
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Henry, M.; Stevens, H. The vegan package. Community Ecol. Package 2013, 10, 631–637. [Google Scholar]
- Bray, J.B.; Curtis, J.T. An ordination of the upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Spinelli, M.L.; Franzosi, C.; Olguin Salinas, H.; Capitanio, F.L.; Alder, V.A. Appendicularians and copepods from Scotia Bay (Laurie island, South Orkney, Antarctica): Fluctuations in community structure and diversity in two contrasting, consecutive summers. Polar Biol. 2017, 41, 663–678. [Google Scholar] [CrossRef]
- Gorsky, G.; Youngbluth, M.J.; Deibel, D. Response of Marine Eco-Systems to Global Change: Ecological Impact of Appendicularians; Editions Scientifiques; GB Scientific Publisher: Paris, France, 2005; pp. 1–440. [Google Scholar]
- Atkinson, A.; Ward, P.; Hunt, B.P.V.; Pakhomov, E.A.; Hosie, G.W. An overview of South-ern Ocean zooplankton data: Abundance, biomass, feeding and functional relationships. CCAMLR Sci. 2012, 19, 171–218. [Google Scholar]
- Krajewska, M.; Szymczak-Żyła, M.; Tylmann, W.; Kowalewska, G. Climate change impact on primary production and phytoplankton taxonomy in Western Spitsbergen fjords based on pigments in sediments. Glob. Planet. Chang. 2020, 189, 103158. [Google Scholar] [CrossRef]
- Kittel, W.; Siciński, J.; Żmijewska, M.I.; Bielecka, L.; Błachowiak-Samołyk, K. Antarctic neritic zooplankton community (Admiralty Bay, King George Island, South Shetland Island). Pol. Polar Res. 2001, 22, 11–33. [Google Scholar]
- Acuña, J.-L.; Anadón, R. Appendicularian assemblages in a shelf area and their relationship with temperature. J. Plankton Res. 1992, 14, 1233–1250. [Google Scholar] [CrossRef]
- Ligowski, R. Net phytoplankton of the Admiralty Bay (King George Island, South Shetland Island) in 1983. Pol. Polar Res. 1986, 7, 127–154. [Google Scholar]
- Woźniak, B.; Hapter, R.; Maj, B. The inflow of solar Energy and the irradiance of the euphotic zone in the region of Ezcurra Inlet during the antarctica summer of 1977/78. Oceanologia 1983, 15, 141–173. [Google Scholar]
- Wasiłowska, A.; Kopczyńska, E.E.; Rzepecki, M. Temporal and spatial variation of phytoplankton in Admiralty Bay, South Shetlands: The dynamics of summer blooms shown by pigment and light microscopy analysis. Polar Biol. 2015, 38, 1249–1265. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, M.C.M.; Williams, G.D. Distribution and abundance of Larvaceans in the Southern Ocean between 30 and 80° E. Deep Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 905–915. [Google Scholar] [CrossRef]
Figure 1.
Geographic position of the sampling station in Admiralty Bay, Ezcurra Inlet (King George Island), December 2003–November 2004.
Figure 1.
Geographic position of the sampling station in Admiralty Bay, Ezcurra Inlet (King George Island), December 2003–November 2004.

Figure 2.
Variabilities in temperature (T) (°C) and salinity, in the surface water layer (50–0 m) and in the intermediate water layer (100–50 m) of Admiralty Bay from December 2003 to November 2004.
Figure 2.
Variabilities in temperature (T) (°C) and salinity, in the surface water layer (50–0 m) and in the intermediate water layer (100–50 m) of Admiralty Bay from December 2003 to November 2004.

Figure 3.
Monthly changes in abundances of Appendicularia in Admiralty Bay, December 2003–November 2004: (A)—all recorded individuals and species (means ± SDs) (ind. 103 m−3), (B)—Fritillaria borealis, (C)—Oikopleura gaussica.
Figure 3.
Monthly changes in abundances of Appendicularia in Admiralty Bay, December 2003–November 2004: (A)—all recorded individuals and species (means ± SDs) (ind. 103 m−3), (B)—Fritillaria borealis, (C)—Oikopleura gaussica.

Figure 4.
Ordisurf models showing the distribution of temperature over the NMDS clusters of Appendicularia (all identified species), Fritillaria borealis and Oikopleura gaussica. Circles represent samples and predicted temperatures for the community or selected species were plotted as a smooth surface on ordination diagrams.
Figure 4.
Ordisurf models showing the distribution of temperature over the NMDS clusters of Appendicularia (all identified species), Fritillaria borealis and Oikopleura gaussica. Circles represent samples and predicted temperatures for the community or selected species were plotted as a smooth surface on ordination diagrams.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Abundance (mean ± SDs) (ind. 103 m−3) of Appendicularia in Admiralty Bay in summer (S), transition period (TP), and winter (W) seasons in the surface layer; App—Appendicularia, Fb—Fritillaria borealis, Fa—F. aberrans, Fan—F. antarctica, F—Fritillaria sp., Of—Oikopleura fusiformis, Op—O. parva, Og—O. gaussica, and O—Oikopleura sp.
Table 1.
Abundance (mean ± SDs) (ind. 103 m−3) of Appendicularia in Admiralty Bay in summer (S), transition period (TP), and winter (W) seasons in the surface layer; App—Appendicularia, Fb—Fritillaria borealis, Fa—F. aberrans, Fan—F. antarctica, F—Fritillaria sp., Of—Oikopleura fusiformis, Op—O. parva, Og—O. gaussica, and O—Oikopleura sp.
| Period | Fb | Fa | Fan | F | Of | Op | Og | O | App |
|---|---|---|---|---|---|---|---|---|---|
| S | 7541.4 ± 18,917.9 | 6.4 ± 25.5 | 0 | 127.4 ± 296.5 | 0 | 6.4 ± 25.5 | 57.3 ± 90.9 | 12.7 ± 34.8 | 968.9 ± 6964.4 |
| TP | 1630.6 ± 1392.9 | 0 | 0 | 84.9 ± 163.3 | 0 | 0 | 67.9 ± 52.6 | 33.9 ± 52.6 | 227.2 ± 705.8 |
| W | 1293.5 ± 1370.1 | 7.8 ± 28.3 | 7.8 ± 28.3 | 62.7 ± 78.2 | 47.1 ± 114.8 | 7.8 ± 28.3 | 250.9 ± 304.6 | 47.1 ± 98.6 | 215.6 ± 637.5 |
Table 2.
Values of SIMPER analysis of appendicularian taxa accumulating 90% similarity for abundance, in Admiralty Bay; S—summer, TP—transition period, and W—winter.
Table 2.
Values of SIMPER analysis of appendicularian taxa accumulating 90% similarity for abundance, in Admiralty Bay; S—summer, TP—transition period, and W—winter.
| Season | S | TP | W | |||
|---|---|---|---|---|---|---|
| Av. Sim. | 36.14 | 59.70 | 48.05 | |||
| Av. Sim. and Contrib. [%] | ||||||
| Fritillaria borealis | 30.19 | 83.53 | 48.94 | 81.99 | 35.97 | 81.99 |
| Seasons | Summer and TP | Summer and Winter | TP and Winter | |||
| Av. Diss. | 56.14 | 58.56 | 46.36 | |||
| Av. Diss. and Contrib. [%] | ||||||
| Fritillaria borealis | 36.76 | 80.20 | 31.68 | 54.10 | 21.49 | 46.34 |
| Oikopleura gaussica | 7.38 | 13.14 | 10.17 | 17.36 | 10.17 | 21.93 |
Table 3.
The results of the 1- and 2-way PERMANOVA tests are reported as pseudo-F values; significance levels are marked as follows: * p < 0.05 (Ef –environmental factors, T—temperature, S—salinity, depth, season, App—Appendicularia, Fb—Fritillaria borealis, Fa—F. aberrans, Fan—F. antarctica, F—Fritillaria sp., Of—Oikopleura fusiformis, Op—O. parva, Og—O. gaussica, and O—Oikopleura sp.).
Table 3.
The results of the 1- and 2-way PERMANOVA tests are reported as pseudo-F values; significance levels are marked as follows: * p < 0.05 (Ef –environmental factors, T—temperature, S—salinity, depth, season, App—Appendicularia, Fb—Fritillaria borealis, Fa—F. aberrans, Fan—F. antarctica, F—Fritillaria sp., Of—Oikopleura fusiformis, Op—O. parva, Og—O. gaussica, and O—Oikopleura sp.).
| Ef | Df R | Fb | Fa | Fan | F | Of | Op | Og | O | App |
|---|---|---|---|---|---|---|---|---|---|---|
| Season | 1 33 | 0.196 | 1 | 0.37 | 0.345 | 0.14 | 1 | 0.189 | 0.439 | 0.099 |
| Season + T | 1 1 32 | 0.165 0.046 * | 0.798 0.534 | 0.345 0.947 | 0.345 0.431 | 0.034 * 0.359 | 0.766 0.491 | 0.223 0.889 | 0.458 0.756 | 0.066 0.012 * |
| T | 1 33 | 0.148 | 0.823 | 0.422 | 0.257 | 0.336 | 0.861 | 0.359 | 0.65 | 0.069 |
| S | 1 33 | 0.605 | 0.323 | 1 | 0.515 | 0.095 | 0.344 | 0.318 | 0.495 | 0.311 |
| T + S | 1 1 32 | 0.170 0.626 | 0.799 0.323 | 0.411 0.816 | 0.273 0.629 | 0.323 0.076 | 0.812 0.321 | 0.370 0.239 | 0.649 0.437 | 0.063 0.386 |
| Depth | 1 33 | 0.997 | 0.207 | 0.477 | 0.53 | 0.239 | 0.242 | 0.943 | 0.407 | 0.964 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Panasiuk, A.; Kalarus, M. Appendicularia (Tunicata) in an Antarctic Glacial Fjord–Chaotic Fjordic Structure Community or Good Indicators of Oceanic Water Masses? Diversity 2021, 13, 675. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120675
AMA Style
Panasiuk A, Kalarus M. Appendicularia (Tunicata) in an Antarctic Glacial Fjord–Chaotic Fjordic Structure Community or Good Indicators of Oceanic Water Masses? Diversity. 2021; 13(12):675. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120675
Chicago/Turabian StylePanasiuk, Anna, and Marcin Kalarus. 2021. "Appendicularia (Tunicata) in an Antarctic Glacial Fjord–Chaotic Fjordic Structure Community or Good Indicators of Oceanic Water Masses?" Diversity 13, no. 12: 675. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120675
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.