
Habitat Selection by Brown Bears with Varying Levels of Predation Rates on Ungulate Neonates

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
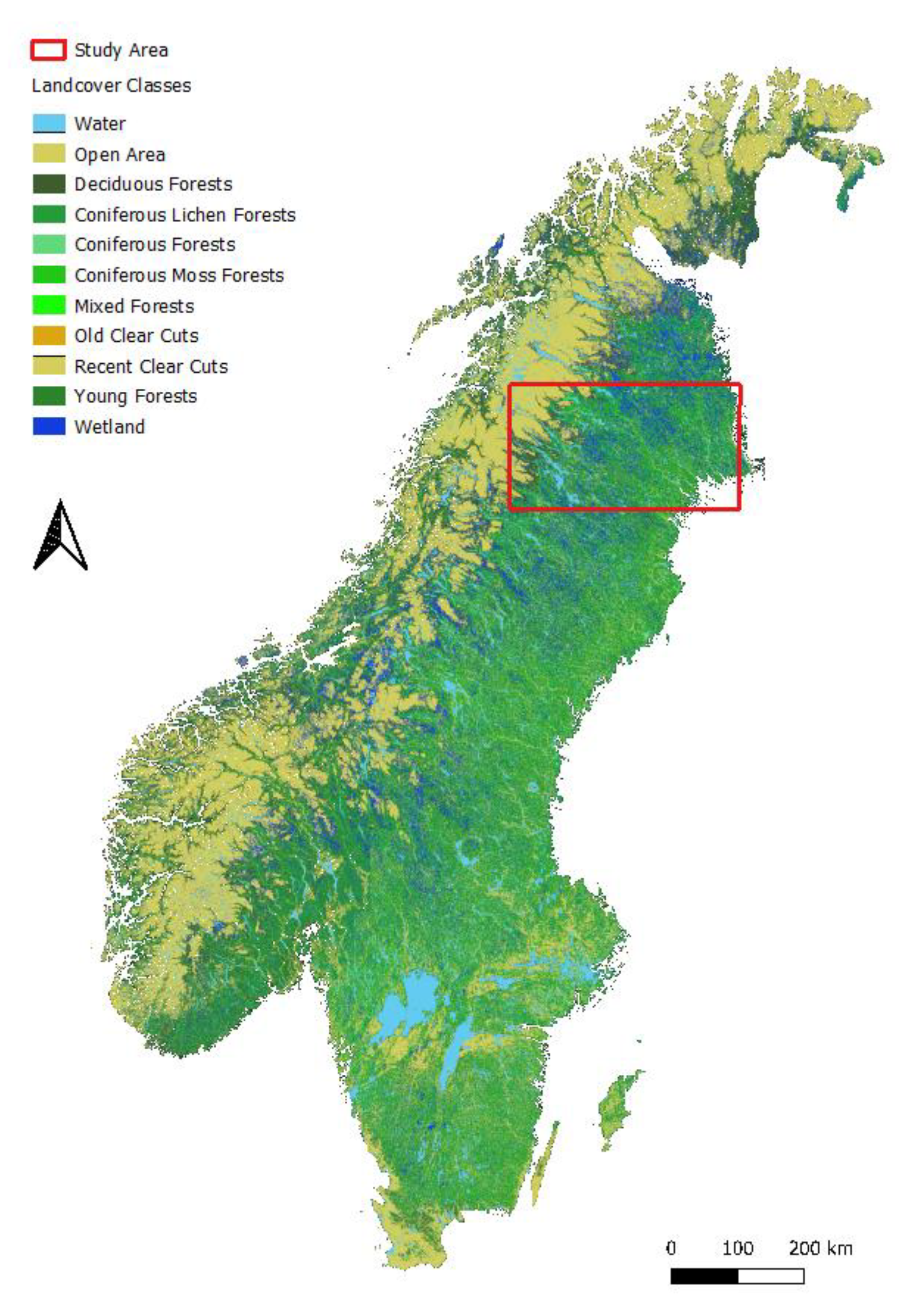
2.1. Study Area
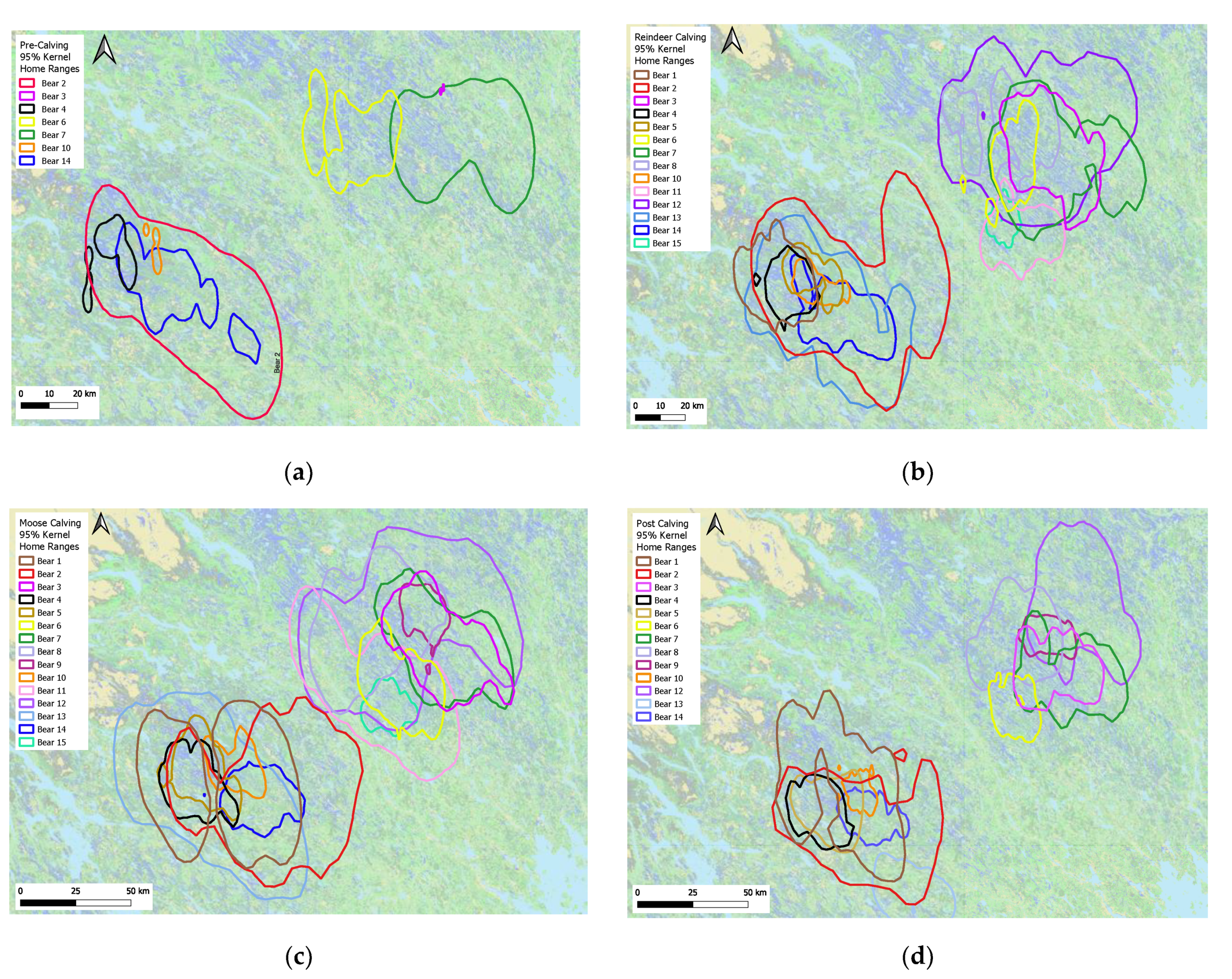
2.2. Study Period—The ‘Bear Year’
2.3. GPS Locations of Bears
2.4. Predation Data and Clusters of GPS Positions
2.5. Bear Habitat Selection
2.6. Statistical Analyses
3. Results
3.1. Bear Kill Rates
3.2. Bear Habitat Selection (Resource Selection Function)
3.3. Differences in Habitat Selection between Low and High Predatory Bears
4. Discussion
4.1. Changes in Habitat Selection during Predatory Periods
4.2. Differences in Habitat Selection between Low and High Predatory Bears
4.3. Consequences for Conservation and Management of Large Carnivores
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bruskotter, J.T.; Shelby, L.B. Human dimensions of large carnivore conservation and management: Introduction to the special issue. Hum. Dimens. Wildl. 2010, 15, 311–314. [Google Scholar] [CrossRef]
- Chapron, G.; Kaczensky, P.; Linnell, J.D.; Von Arx, M.; Huber, D.; Andren, H.; Boitani, L. Recovery of large carnivores in Europe’s modern human-dominated landscapes. Science 2014, 346, 1517–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ripple, W.J.; Estes, J.A.; Beschta, R.L.; Wilmers, C.C.; Ritchie, E.G.; Hebblewhite, M.; Wirsing, A.J. Status and ecological effects of the world’s largest carnivores. Science 2014, 343, 1241484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Distefano, E. Human-Wildlife Conflict Worldwide: Collection of Case Studies, Analysis of Management Strategies and Good Practices; FAO Corporate Document Repository; Food and Agricultural Organization of the United Nations (FAO), Sustainable Agriculture and Rural Development Initiative (SARDI): Rome, Italy, 2005; Available online: https://www.fao.org/publications/card/en/c/e21b6162-b3ad-4661-8c52-710f95ebeaf7 (accessed on 14 December 2021).
- May, R.; Van Dijk, J.; Wabakken, P.; Swenson, J.E.; Linnell, J.D.; Zimmermann, B.; Landa, A. Habitat differentiation within the large-carnivore community of Norway’s multiple-use landscapes. J. Appl. Ecol. 2008, 45, 1382–1391. [Google Scholar] [CrossRef] [Green Version]
- Inga, B. Reindeer (Rangifer tarandus tarandus) feeding on lichens and mushrooms: Traditional ecological knowledge among reindeer-herding Sami in northern Sweden. Rangifer 2009, 27, 93–106. [Google Scholar] [CrossRef] [Green Version]
- Furberg, M.; Evengård, B.; Nilsson, M. Facing the limit of resilience: Perceptions of climate change among reindeer herding Sami in Sweden. Glob. Health Action 2011, 4, 8417. [Google Scholar] [CrossRef] [Green Version]
- Tveraa, T.; Stien, A.; Brøseth, H.; Yoccoz, N.G. The role of predation and food limitation on claims for compensation, reindeer demography and population dynamics. J. Appl. Ecol. 2014, 51, 1264–1272. [Google Scholar] [CrossRef] [Green Version]
- Sivertsen, T.R.; Åhman, B.; Steyaert, S.M.; Rönnegård, L.; Frank, J.; Segerström, P.; Skarin, A. Reindeer habitat selection under the risk of brown bear predation during calving season. Ecosphere 2016, 7, e01583. [Google Scholar] [CrossRef]
- Hobbs, N.T.; Andren, H.; Persson, J.; Aronsson, M.; Chapron, G. Native predators reduce harvest of reindeer by Sami pastoralists. Ecol. Appl. 2012, 22, 1640–1654. [Google Scholar] [CrossRef] [Green Version]
- Ordiz, A.; Milleret, C.; Kindberg, J.; Månsson, J.; Wabakken, P.; Swenson, J.E.; Sand, H. Wolves, people, and brown bears influence the expansion of the recolonizing wolf population in Scandinavia. Ecosphere 2015, 6, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Rivrud, I.M.; Sivertsen, T.R.; Mysterud, A.; Åhman, B.; Støen, O.; Skarin, A. Reindeer green-wave surfing constrained by predators. Ecosphere 2018, 9, e02210. [Google Scholar] [CrossRef]
- Schwerdtner, K.; Gruber, B. A conceptual framework for damage compensation schemes. Biol. Conserv. 2007, 134, 354–360. [Google Scholar] [CrossRef]
- Rauset, G.R.; Kindberg, J.; Swenson, J.E. Modeling female brown bear kill rates on moose calves using global positioning satellite data. J. Wildl. Manag. 2012, 76, 1597–1606. [Google Scholar] [CrossRef]
- Dahle, B.; Wallin, K.; Cederlund, G.; Persson, I.; Selvaag, L.S.; Swenson, J.E. Predation on adult moose Alces alces by European brown bears Ursus arctos. Wildl. Biol. 2013, 19, 165–169. [Google Scholar] [CrossRef]
- Swenson, J.E.; Sandegren, F.; So-Derberg, A. Geographic expansion of an increasing brown bear population: Evidence for presaturation dispersal. J. Anim. Ecol. 1998, 67, 819–826. [Google Scholar] [CrossRef]
- Singh, N.J.; Börger, L.; Dettki, H.; Bunnefeld, N.; Ericsson, G. From migration to nomadism: Movement variability in a northern ungulate across its latitudinal range. Ecol. Appl. 2012, 22, 2007–2020. [Google Scholar] [CrossRef] [PubMed]
- Swenson, J.E.; Dahle, B.; Busk, H.; Opseth, O.; Johansen, T.; Söderberg, A.; Cederlund, G. Predation on moose calves by European brown bears. J. Wildl. Manag. 2007, 71, 1993–1997. [Google Scholar] [CrossRef]
- Tallian, A.; Ordiz, A.; Metz, M.C.; Milleret, C.; Wikenros, C.; Smith, D.W.; Stahler, D.R.; Kindberg, J.; Macnulty, D.R.; Wabakken, P.; et al. Competition between apex predators? Brown bears decrease wolf kill rate on two continents. Proc. R. Soc. B Biol. Sci. 2017, 284, 20162368. [Google Scholar] [CrossRef] [PubMed]
- Tallian, A.; Ordiz, A.; Zimmermann, B.; Sand, H.; Wikenros, C.; Wabakken, P.; Bergqvist, G.; Kindberg, J. The return of large carnivores: Using hunter observation data to understand the role of predators on ungulate populations. Glob. Ecol. Conserv. 2021, 27, e01587. [Google Scholar] [CrossRef]
- Ordiz, A.; Milleret, C.; Uzal, A.; Zimmermann, B.; Wabakken, P.; Wikenros, C.; Sand, H.; Swenson, J.E.; Kindberg, J. Individual Variation in Predatory Behavior, Scavenging and Seasonal Prey Availability as Potential Drivers of Coexistence between Wolves and Bears. Diversity 2020, 12, 356. [Google Scholar] [CrossRef]
- Gervasi, V.; Nilsen, E.B.; Sand, H.; Panzacchi, M.; Rauset, G.R.; Pedersen, H.C.; Linnell, J.D. Predicting the potential demographic impact of predators on their prey: A comparative analysis of two carnivore–ungulate systems in Scandinavia. J. Anim. Ecol. 2012, 81, 443–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauset, G.R.; Andrén, H.; Swenson, J.E.; Samelius, G.; Segerström, P.; Zedrosser, A.; Persson, J. National parks in northern Sweden as refuges for illegal killing of large carnivores. Conserv. Lett. 2016, 9, 334–341. [Google Scholar] [CrossRef] [Green Version]
- Andrén, H.; Linnell, J.D.; Liberg, O.; Andersen, R.; Danell, A.; Karlsson, J.; Segerström, P. Survival rates and causes of mortality in Eurasian lynx (Lynx lynx) in multi-use landscapes. Biol. Conserv. 2006, 131, 23–32. [Google Scholar] [CrossRef]
- Persson, J.; Ericsson, G.; Segerström, P. Human caused mortality in the endangered Scandinavian wolverine population. Biol. Conserv. 2009, 142, 325–331. [Google Scholar] [CrossRef]
- Leclerc, M.; Vander, E.; Zedrosser, A.; Swenson, J.E.; Kindberg, J.; Pelletier, F. Quantifying consistent individual differences in habitat selection. Oecologia 2016, 180, 697–705. [Google Scholar] [CrossRef] [Green Version]
- Toscano, B.J.; Gownaris, N.J.; Heerhartz, S.M. Personality, foraging behavior and specialization: Integrating behavioral and food web ecology at the individual level. Oecologia 2016, 182, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Milleret, C.; Ordiz, A.; Chapron, G.; Andreassen, H.P.; Kindberg, J.; Månsson, J.; Tallian, A.; Wabakken, P.; Wikenros, C.; Zimmermann, B.; et al. Habitat segregation between brown bears and gray wolves in a human-dominated landscape. Ecol. Evol. 2018, 8, 11450–11466. [Google Scholar] [CrossRef] [PubMed]
- Stenset, N.E.; Lutnæs, P.N.; Bjarnadóttir, V.; Dahle, B.; Fossum, K.H.; Jigsved, P.; Swenson, J.E. Seasonal and annual variation in the diet of brown bears Ursus arctos in the boreal forest of southcentral Sweden. Wildl. Biol. 2016, 22, 107–117. [Google Scholar] [CrossRef] [Green Version]
- Ordiz, A.; Kindberg, J.; Sæbø, S.; Swenson, J.E.; Støen, O.G. Brown bear circadian behavior reveals human environmental encroachment. Biol. Conserv. 2014, 173, 1–9. [Google Scholar] [CrossRef]
- Kindberg, J.; Swenson, J.E.; Ericsson, G.; Bellemain, E.; Miquel, C.; Taberlet, P. Estimating population size and trends of the Swedish brown bear Ursus arctos population. Wildl. Biol. 2011, 17, 114–123. [Google Scholar] [CrossRef] [Green Version]
- Lavsund, S.; Nygrén, T.; Solberg, E.J. Status of moose populations and challenges to moose management in Fennoscandia. Alces 2003, 39, 109–130. [Google Scholar]
- Evans, A.L.; Sahlén, V.; Støen, O.; Fahlman, Å.; Brunberg, S.; Madslien, K.; Fröbert, O.; Swenson, J.E.; Arnemo, J.M. Capture, anesthesia, and disturbance of free-ranging brown bears (Ursus arctos) during hibernation. PLoS ONE 2012, 7, e40520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahle, B.; Swenson, J.E. Home ranges in adult Scandinavian brown bears (Ursus arctos): Effect of mass, sex, reproductive category, population density and habitat type. J. Zool. 2003, 260, 329–335. [Google Scholar] [CrossRef]
- Ordiz, A.; Støen, O.; Sæbø, S.; Sahlén, V.; Pedersen, B.E.; Kindberg, J.; Swenson, J.E. Lasting behavioural responses of brown bears to experimental encounters with humans. J. Appl. Ecol. 2013, 50, 306–314. [Google Scholar] [CrossRef]
- Hertel, A.G.; Steyaert, S.M.; Zedrosser, A.; Mysterud, A.; Lodberg-Holm, H.K.; Swenson, J.E. Bears and berries: Species-specific selective foraging on a patchily distributed food resource in a human-altered landscape. Behav. Ecol. Sociobiol. 2016, 70, 831–842. [Google Scholar] [CrossRef] [Green Version]
- Frank, J.; Støen, O.G.; Segerström, P.; Stokke, R.; Persson, L.T.; Stokke, L.H.; Persson, S.; Stokke, N.; Persson, A.; Segerström, E.; et al. Björnpredation på ren och potentiella effekter av tre förebyggande åtgärder. 2012, 6. [In Swedish]. Available online: https://pub.epsilon.slu.se/13047/ (accessed on 14 December 2021).
- Friebe, A.; Swenson, J.E.; Sandegren, F. Denning chronology of female brown bears in central Sweden. Ursus 2001, 12, 37–45. [Google Scholar]
- Arnemo, J.M.; Evans, A. Biomedical Protocols for Free-Ranging Brown Bears, Wolves, Wolverines and Lynx; Hedmark University College Evenstad: Koppang, Norway, 2017. [Google Scholar]
- Frank, S.C.; Leclerc, M.; Pelletier, F.; Rosell, F.; Swenson, J.; Bischof, R.; Zedrosser, A. Sociodemographic factors modulate the spatial response of brown bears to vacancies created by hunting. J. Anim. Ecol. 2018, 87, 247–258. [Google Scholar] [CrossRef] [Green Version]
- Worton, B.J. Kernel methods for estimating the utilization distribution in home-range studies. Ecology 1989, 70, 164–168. [Google Scholar] [CrossRef]
- Martin, J. Habitat selection and movement by brown bears in multiple-use landscapes. Ph.D. Thesis, Norwegian University of Life Sciences, Department of Ecology and Natural Resource Management, Ås, Norway, 2009. [Google Scholar]
- Moe, T.; Kindberg, J.; Jansson, I.; Swenson, J. Importance of diel behaviour when studying habitat selection: Examples from female Scandinavian brown bears (Ursus arctos). Can. J. Zool. 2007, 85, 518–525. [Google Scholar] [CrossRef] [Green Version]
- Pape, R.; Löffler, J. Ecological dynamics in habitat selection of reindeer: An interplay of spatial scale, time, and individual animal’s choice. Polar Biol. 2015, 38, 1891–1903. [Google Scholar] [CrossRef]
- Bjørneraas, K.; Solberg, E.J.; Herfindal, I.; Moorter, B.V.; Rolandsen, C.M.; Tremblay, J.; Astrup, R. Moose Alces alces habitat use at multiple temporal scales in a human-altered landscape. Wildl. Biol. 2011, 17, 44–54. [Google Scholar] [CrossRef] [Green Version]
- Sappington, J.M.; Longshore, K.M.; Thompson, D.B. Quantifying landscape ruggedness for animal habitat analysis: A case study using bighorn sheep in the Mojave Desert. J. Wildl. Manag. 2007, 71, 1419–1426. [Google Scholar] [CrossRef]
- Environmental Systems Resource Institute. ArcMap 10.3.1; Environmental Systems Research Institute: Redlands, CA, USA, 2015. [Google Scholar]
- Zuur, A.F.; Hilbe, J.M.; Ieno, E.N. A Beginner’s Guide to GLM and GLMM with R: A Frequentist and Bayesian Perspective for Ecologists; Highland Statistics Limited: Newburgh, UK, 2013. [Google Scholar]
- Milleret, C. Is moose (Alces alces) habitat selection affected by wolf (Canis lupus) re-establishment in south-central Sweden. Ph.D. Thesis, Université Claude Bernard, Villeurbanne, France, 2012. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Calenge, C. The package “adehabitat” for the R software: A tool for the analysis of space and habitat use by animals. Ecol. Model. 2006, 197, 516–519. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Herfindal, I.; Tremblay, J.P.; Hansen, B.B.; Solberg, E.J.; Heim, M.; Sæther, B.E. Scale dependency and functional response in moose habitat selection. Ecography 2009, 32, 849–859. [Google Scholar] [CrossRef]
- Ordiz, A.; Støen, O.G.; Delibes, M.; Swenson, J.E. Staying cool or staying safe in a human-dominated landscape: Which is more relevant for brown bears? Oecologia 2017, 185, 191–194. [Google Scholar] [CrossRef] [Green Version]
- Sahlén, E.S. Indirect Effects of Predation in Human-Modified Landscapes. Ph.D. Thesis, Swedish University of Agricultural Sciences, Faculty of Forest Sciences, Department of Wildlife, Fish and Environmental Studies, Umeå, Sweden, 2016. [Google Scholar]
- Dussault, C.; Ouellet, J.; Courtois, R.; Huot, J.; Breton, L.; Jolicoeur, H. Linking moose habitat selection to limiting factors. Ecography 2005, 28, 619–628. [Google Scholar] [CrossRef]
- Melin, M.; Matala, J.; Pusenius, J.; Packalen, T. Calving and post-calving habitat use of female moose in two contrasting landscapes. Wildl. Biol. 2019, 2019, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Iversen, M.; Fauchald, P.; Langeland, K.; Ims, R.A.; Yoccoz, N.G.; Bråthen, K.A. Phenology and cover of plant growth forms predict herbivore habitat selection in a high latitude ecosystem. PLoS ONE 2014, 9, e100780. [Google Scholar]
- Mårell, A.; Edenius, L. Spatial heterogeneity and hierarchical feeding habitat selection by reindeer. Arct. Antarct. Alp. Res. 2006, 38, 413–420. [Google Scholar] [CrossRef]
- Frank, J.; Eklund, A. Poor construction, not time, takes its toll on subsidised fences designed to deter large carnivores. PLoS ONE 2017, 12, e0175211. [Google Scholar] [CrossRef] [PubMed]
- Ordiz, A.; Sæbø, S.; Kindberg, J.; Swenson, J.E.; Støen, O.-G. Seasonality and human disturbance alter brown bear activity patterns; implications for circumpolar carnivore conservation? Anim. Conserv. 2017, 20, 51–60. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Variable | Description | Area (and Percentage Area) of Variable within ‘Study Area’ | Source |
|---|---|---|---|
| Coniferous Forest | Minimum Euclidian distance to conifer forests (m) (mixed ground flora and understory) | 12,258 km2 (30.49%) | Geographical Data Sweden [9] |
| Coniferous Lichen Forest | Minimum Euclidian distance to conifer lichen forests (m) (ground flora dominated by lichens) | 4470 km2 (11.12%) | Geographical Data Sweden [9] |
| Coniferous Moss Forest | Minimum Euclidian distance to conifer moss forests (m) (ground flora dominated by mosses, includes coniferous forests on bog) | 850 km2 (2.11%) | Geographical Data Sweden [9] |
| Deciduous Forest | Minimum Euclidian distance to deciduous forests (m) | 1574 km2 (3.92%) | Geographical Data Sweden [9] |
| Mixed Forest | Minimum Euclidian distance to mixed forests (m) | 2871 km2 (7.14%) | Geographical Data Sweden [9] |
| Open Area | Minimum Euclidian distance to open areas, including mountainous areas, bare ground, grasslands, etc. (m) | 1389 km2 (3.45%) | Geographical Data Sweden [9] |
| Water | Minimum Euclidian distance to water bodies, including lakes and rivers (m) | 3197 km2 (7.95%) | Geographical Data Sweden [9] |
| Wetland | Minimum Euclidian distance to wetlands, including peatlands and bogs (m) | 7177 km2 (17.85%) | Geographical Data Sweden [9] |
| Terrain Ruggedness Index Score (TRI) | A variable calculated as the total length (m) of the linear development of level curves within a 1.56 ha. | N/A | Geographical Data Sweden [9] |
| Old Clear Cuts | Minimum Euclidian distance to clear cuts that are between 6–12 years old or <2 m in the year 2000 (m) (derived from 1:10,000 satellite images, taken from 2000 to 2012) | 41 km2 (0.10%) | Utförd avverkning, Swedish Forest Agency 2015 [9] |
| Recent Clear Cuts | Minimum Euclidian distance to clear cuts that are between 0 and 5 years old in the year 2015. | 2610 km2 (6.49%) | Utförd avverkning, Swedish Forest Agency 2015 [9] |
| Young Forest | Minimum Euclidian distance to previous clear cuts that are 2–5 m in the year 2000 (m) | 3767 km2 (9.37%) | Utförd avverkning, Swedish Forest Agency 2015 [9] |
| Public Roads with Regular Traffic | Minimum Euclidian distance to public roads with regular traffic (m) | N/A | 1:100,000 Vägkartan, Geographical Data Sweden, National Land Survey Sweden, [9] |
| Gravel Roads | Minimum Euclidian distance to gravel roads (m) | N/A | 1:100,000 Vägkartan, Geographical Data Sweden, National Land Survey Sweden, [9] |
| Bear ID | Sex | Age | Total Kills per Period 2011 | Total Kills per Period 2012 | Average Kills per Day, during Calving | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Pre-Calving (Early Spring) | Reindeer Calving (May) | Moose Calving (June) | Post-Calving (July–September) | Pre-Calving (Early Spring) | Reindeer Calving (May) | Moose Calving (June) | Post-Calving (July–September) | ||||
| 1 | Male | unknown | 13 (10R/3M) | 4 (4M) | 0.48 | ||||||
| 2 | Male | 7 | 1 (1M) | 11 (7R/4M) | 8 (8M) | 1 (1M) | 0.30 | ||||
| 3 | Female | 3 | 1 (1M) | 26 (23R/3M) | 6 (1R/5M) | 1 (1R) | 5 (4R/1M) | 2 (2R) | 0.35 | ||
| 4 | Female | 13 | 2 (2R) | 38 (35R/3M) | 18 (1R/17M) | 2 (2M) | 5 (2R/3M) | 0.62 | |||
| 5 | Female | 11 | 37 (37R) | 2 (2M) | 1.39 | ||||||
| 6 | Female | unknown | 2 (1R/1M) | 11 (11M) | 0.31 | ||||||
| 7 | Male | 3 | 29 (26R/3M) | 6 (6M) | 1 (1M) | 24 (24R) | 2 (2R) | 0.87 | |||
| 8 | Male | 10 | 14 (14R) | 1 (1M) | 0.54 | ||||||
| 9 | Female | 5 | 1 (1R) | 0 | |||||||
| 10 | Female | 3 | 15 (15R) | 8 (5R/3M) | 14 (14R) | 3 (3M) | 0.44 | ||||
| 11 | Male | 10 | 3 (3R) | 0.21 | |||||||
| 12 | Male | 6 | 7 (6R/1M) | 1 (1M) | 0.38 | ||||||
| 13 | Male | 14 | 4 (3R/1M) | 1 (1M) | 1 (1M) | 0.21 | |||||
| 14 | Female | 8 | 4 (2R/2M) | 6 (6M) | 1 (1R) | 38 (36R/2M) | 3 (3M) | 0.52 | |||
| 15 | Female | 8 | 13 (11R/2M) | 5 (4R/1M) | 0.43 | ||||||
| Period | ||||
|---|---|---|---|---|
| Pre-Calving (Early Spring) | Reindeer Calving (May) | Moose Calving (June) | Post-Calving (Hyperphagia) (July to September) | |
| Water | ||||
| Wetlands | ||||
| Open Areas | ||||
| Deciduous Forest | ||||
| Coniferous Forest | ||||
| Coniferous Lichen Forests | ||||
| Coniferous Moss Forests | ||||
| Mixed Forests | ||||
| Old Clear Cuts | ||||
| Young Forests | ||||
| Public Roads with Regular Traffic | ||||
| Gravel Roads | ||||
| Terrain Ruggedness Index | ||||
| Period | ||||||||
|---|---|---|---|---|---|---|---|---|
| Pre-Calving (Early Spring) | Reindeer Calving (May) | Moose Calving (June) | Post-Calving (July to September) | |||||
| Interaction | High Predatory | Low Predatory | High Predatory | Low Predatory | High Predatory | Low Predatory | High Predatory | Low Predatory |
| Wetlands × behaviour | + | - | + | ++ | ns(+) | + | - | |
| Open Areas × behaviour | + | ++ | - | + | - | -- | - | -- |
| Deciduous Forest × behaviour | + | - | + | - | ns(+) | ns(+) | ||
| Coniferous Forest × behaviour | ++ | + | + | - | + | ++ | ns(+) | |
| Coniferous Lichen Forests × behaviour | -- | - | ns(−) | + | - | - | + | |
| Coniferous Moss Forests × behaviour | - | + | ns(−) | - | + | -- | - | |
| Mixed Forests × behaviour | - | + | - | -- | - | - | - | + |
| Old Clear Cuts × behaviour | -- | - | - | + | + | ++ | ++ | + |
| Young Forests × behaviour | + | - | - | -- | - | + | - | -- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Twynham, K.; Ordiz, A.; Støen, O.-G.; Rauset, G.-R.; Kindberg, J.; Segerström, P.; Frank, J.; Uzal, A. Habitat Selection by Brown Bears with Varying Levels of Predation Rates on Ungulate Neonates. Diversity 2021, 13, 678. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120678
Twynham K, Ordiz A, Støen O-G, Rauset G-R, Kindberg J, Segerström P, Frank J, Uzal A. Habitat Selection by Brown Bears with Varying Levels of Predation Rates on Ungulate Neonates. Diversity. 2021; 13(12):678. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120678
Chicago/Turabian StyleTwynham, Kate, Andrés Ordiz, Ole-Gunnar Støen, Geir-Rune Rauset, Jonas Kindberg, Peter Segerström, Jens Frank, and Antonio Uzal. 2021. "Habitat Selection by Brown Bears with Varying Levels of Predation Rates on Ungulate Neonates" Diversity 13, no. 12: 678. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120678