
Morphogenetic Polyvariance in the Colonial Hydroid Dynamena pumila (L.)

Department of Invertebrate Zoology, Lomonosov Moscow State University, 119991 Moscow, Russia
Diversity 2021, 13(12), 683; https://0-doi-org.brum.beds.ac.uk/10.3390/d13120683
Submission received: 12 November 2021
/
Revised: 14 December 2021
/
Accepted: 15 December 2021
/
Published: 20 December 2021
(This article belongs to the Special Issue Diversity, Ecology, and Evolution of Hydrozoans)
Abstract
:The formation of aberrant modules (internodes) of the shoot was found sporadically in the colonial hydroid Dynamena pumila from the family Sertulariidae, which is distinguished by the symmetrical two-row position of the hydrotheca on the shoot stem. Despite rare significant deviations from the norm in the structure of some modules, the following modules in the shoots are formed normally. All variants of deviation from the norm (called morphovariations) are classified and, in general, are represented by four groups according to the possible reasons for their occurrence. The morphotypes are the most interesting morphological variations and look like similar modules in other genera of hydroids. Considering the position of the aberrant modules on the shoots of one colony, we can confidently infer that they were formed at different times, which undermines the assumption that deviations from the norm in morphogenesis were caused by environmental factors. All morphovariations are characterized by certain occurrences. Consequently, the described phenomenon is not limited to phenotypic plasticity, polymorphism, or sequential changes of phenotypes in the life cycle and, therefore, is singled out as a special phenomenon called morphogenetic polyvariance.
1. Introduction
In one organism, a colony in hydroids [1], along with the normal forms of shoot structure, various deviations from the norm can sometimes be found. They are of great interest when studying the mechanisms of morphogenesis. They offer potential resources of variability in morphogenesis—the realization of parallel ways of development of the organism.
The phenomenon of “morphogenetic polyvariance” was described in hydroids for the first time 45 years ago in Dynamena pumila (L., 1758), belonging to the Sertulariidae family of the order Leptothecata [2]. Since that time, the study of this phenomenon has continued, but never before the results of the work were published in English.
Morphogenetic polyvariance is the ability of an organism to produce some alternative phenotypes besides the normal (canonical). Alternative phenotypes are called “morphovariations”.
The majority of species have more than one canonical phenotype; for example, hydroids have polyps (or colonial stage of development) and medusa phenotypes. A lot of species of insects’ life cycles include the stages of the worm, bunch, and winged forms. Among flatworms, there are some examples of complex life cycles with more than two phenotypes.
Between canonic and alternative phenotypes, there is a principal difference. Canonic phenotypes manifest a natural regularity. Each phenotype arises at a definite stage of individual development.
Morphogenetic polyvariance emerges in a fundamentally different way. First of all, morphovariations arise: (1) occasionally (stochastically), and (2) without obvious connection with exogenous factors.
Morphogenetic polyvariance is not easy to recognize in a unitary organism. Comparing individuals of the same species, it is difficult to determine to what extent the detected deviation is a simple consequence of population variability or genetic mutation.
It is much easier to detect and study morphogenetic polyvariance in modular organisms, which include colonial invertebrates, many plants and fungi [3,4,5,6]. Modular organisms are characterized by reproduction of the same form (module—zooid, leaf, branch, etc.) many times. Among the many stereotypical modules, there are abnormal ones, but all of them, both normal and abnormal modules, are implemented according to the same genetic program, based on a single genotype of a given organism.
Nevertheless, among the whole set of modular organisms, only in one species, D. pumila, the phenomenon of morphogenetic polyvariance has been studied in sufficient detail. This is due to two circumstances. First, D. pumila forms dense populations at the boundary between the littoral and the upper sublittoral, which makes it easy to collect colonies in any quantity, in any season and from different habitats. Secondly, any deviations from the norm in the structure of D. pumila shoots are easy to recognize.
Among all Leptothecata, D. pumila is distinguished by the particularly symmetrical form of its shoots, in which the hydrants (and their perisarcal sheaths—hydrotheca) are located opposite one another in two rows. The slightest distortion in the formation is immediately noticeable by the deviation of the shape of the shoot from the ideally symmetric.
This is why morphogenetic polyvariance was found easily in D. pumila. For this species, research methods were developed based mainly on the classification of morphovariations and their quantitative accounting.
By summarizing all the results obtained over many years of research, for the first time in this article the phenomenon of morphogenetic polyvariance will be described, focusing on the following issues:
- (a)
- How diverse are morphological aberrations?
- (b)
- Are they deformities?
- (c)
- Is there a limit to deviations from the norm of the structure?
- (d)
- Are there any regular differences between morphovariations in terms of their frequency of occurrence?
- (e)
- How often do morphological aberrations appear in the colony?
2. Materials and Methods
2.1. Characteristics of the Studied Species
The colonial hydroid D. pumila consists of many shoots, arising from a smooth, tubular stolon. The hydrants inside hydrothecae are located on the shoot stem directly opposite in two rows, connected by the common coenosarc [7,8,9,10]. All this is covered by an external chitinous perisarc, whose shape is quite complex and species-specific (Figure 1). The shoot and its lateral branches are arranged in the same way, differing neither in the size and shape of the hydrotheca, nor in their mutual arrangement.
Repeated (cyclic) morphogenesis [11] means that in the process of differentiation, in addition to the parts characteristic of a given shoot module, a new primordium is also formed, from which the formation of the next module will begin (Figure 2). That is, cyclic morphogenesis is characterized by the formation of a rudiment at the end of a series of transformations of the same size and shape as the one from which the growth of the next module began.
The escape modules of D. pumila have highly stable, accurate and rich characteristic features such as the shape of the opening of the hydrothecae; the aperture at the base of hydrotheca; the defined length ratio of adjacent and distant parts of the hydrotheca; the strict alternation of wide and narrow parts of the shoot. Therefore, any deviations from the norm are clearly distinguishable when studying shoots under a stereomicroscope.
The cyclical morphogenesis in D. pumila makes it possible to track the process of the multiple formations of stereotypical colony modules and compare them with each other, helping determine the nature and degree of environmental influence on the development of the organism, or setting the limit of endogenous phenotypic variability.
Deviations from the usual (canonical) form of the module are called aberrant or anomalous modules. All anomalies must be subdivided into those that are capable of performing their vital functions, and others that are not capable of this. The latter are essentially aberrant. Aberrant modules are very diverse, and therefore difficult to classify, while viable anomalous modules have characteristic forms and their morphogenesis has clear signs of stability. Therefore, such anomalous modules are called morphovariations. Some of the morphovariations turn out to be so similar to the modules of shoots of hydroids from other genera that it would be correct to call them morphotypes. If the morphovariations are rarely repeated in sequentially formed modules, then the morphotypes are often repeated, thus demonstrating certain morphogenetic stability.
2.2. Collection of Material
The material was collected near the White Sea biological station of Moscow State University in Kandalaksha Bay. To determine the degree of homogeneity in the D. pumila population, several samples were taken at a distance of 40 km from Sidorov Island near Chupa Bay, as well as at 840 km out into the Barents Sea, near the station of the Murmansk Marine Biological Institute in the village of Dalnie Zelentsyi. Colonies of hydroids together with algae on which they lived were collected in summer 1992, 1993, 1994, 2000, 2005.
2.3. Methodology for Accounting for the Composition and Frequency of Occurrence of Deformities, Morphogenetic Variations and Morphotypes
To determine the composition and frequency of occurrence of alternative morphovariations in D. pumila shoots, the following parameters were taken into account:
(1) The number of modules in the shoot stem; (2) the number of modules in each of the branches; (3) morphovariations in the structure of the shoot modules, identifying them according to the classifications published earlier (Marfenin, 1995) or adding new ones, if necessary; (4) the places of the unusual modules on the branch, counting from the apex.
The unusually shaped shoot modules were drawn and photographed. All anomalies were selected and stored separately at the Department of Invertebrate Zoology, Lomonosov Moscow State University (Moscow, Russia).
In total, 40 samples were examined, including 280,000 modules in the White Sea population and 140,000 modules in the Barents Sea population of D. pumila. These included 11,000 and 4000 shoots, respectively.
For each sample, it was possible to indicate:
- the cumulative proportion (%) of all morphovariations, hereinafter referred to as the cumulative frequency of occurrence of morphovariations (CFOM);
- the absolute value of the occurrence of each morphovariation (FOM) in % of the total number of studied modules.
- Legend:
Roman numerals are the alleged reasons for the aberration of the module and its location, namely:
Group I: Deviations in the morphogenesis of the shoot stem without stopping its further growth;
Group II: Deviations in the morphogenesis of the shoot tip, preventing its further growth;
Group III: Juvenile ugly shoots;
Group IV: Deviations from the norm of the structure of the hydrotheca;
Group V: Deviations from the norm of the location of the lateral branches of the shoot;
Group VI: Deviations from the norm of the shape and position of the gonotheca;
Group VII: Deviations from the norm in the position of stolons in the colony;
Group VIII: Deviations in shoot morphogenesis, possibly caused by damage to it, as well as incomprehensible abnormalities of unknown origin.
Arabic numerals are the registration numbers of the morphovariations within the groups.
Arabic numerals in brackets are the ordinal numbers in the first classification [2], in which there were only 27 morphovariations.
2.4. Valid Sample Size
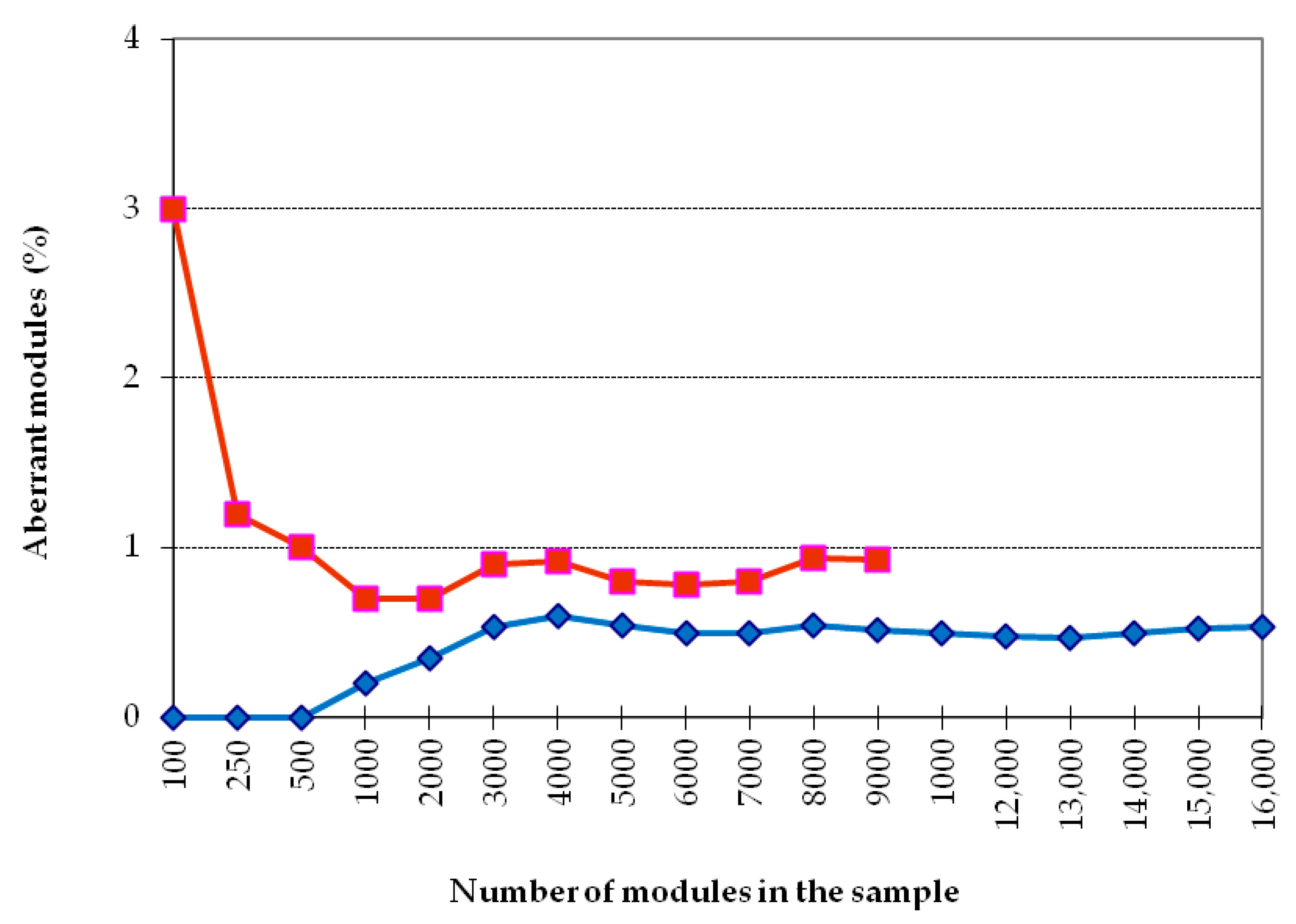
First of all, it was necessary to establish the size of a reliable sample for the analysis of the cumulative frequency of occurrence of morphovariations (CFOM). Figure 3 shows the dependence of CFOM on the sample size, divided into two portions according to the size of shoots (<20 and >20 modules). The same calculations were carried out for a number of other samples. CFOM stabilization occurred in all cases after the sample reached approximately 2000–3000 modules. Stabilization was not necessarily accompanied by a gradual increase in the CFOM value as the sample increased; there were examples of gradual decreases in CFOM to a stable level. Thus, to determine the long exponent, the CFOM was limited to the processing of the modules of 3000 shoot’s modules of D. pumila. Since the cumulative frequency of occurrence mainly comprises several dominant morphovariations, a separate analysis of the most widespread varieties of deviation in the structure of the shoots using the same sample size is reliable.
However, for the analysis of the occurrence of more rare morphovariations, the established sample is insufficient. Having increased the sample to 10,000 modules, we created conditions for the comparative assessment of not only the indicator of the total frequency of occurrence of all abnormal modules but also the occurrence of most of the usual morphovariations individually. Thus, a sample of 10,000 modules was taken as a standard.
Only in some cases, lacking material, I used samples with less than 10,000 modules—we used as little as 3000, and then only for analyzing the indicator of the cumulative frequency of occurrence of all morphovariations (CFOM).
3. Results
3.1. Diversity and Classification of Aberrant Modules (Morphovariations) in D. pumila Shoots
All detected deviations from the norm in the structure of shoot modules were systematized, and each anomaly (morphovariation) received its own serial number. Initially, 27 anomalies in the structure of D. pumila shoot modules were described [2].
With the accumulation of information about new, mostly rare, morphovariations, the classification had to be expanded and revised. In the second version of the classification [12], 47 morphovariations are described, and this list continues to be replenished with new descriptions, such that, to date, 98 varieties are included in the working list of all detected morphovariations of D. pumila.
In this article, 16 basic morphovariations, listed in Table 1 under the numbers corresponding to the 1995 classification to demonstrate the breadth of morphogenetic variations, as well as draw attention to some of them that cannot be explained by simple deformities, injuries, or failures of morphogenetic programs.
Below are the first examples of the morphovariations in the typical morphogenesis of D. pumila shoots, as well as those that differ from the norm for this species.
3.2. Variants of Typical Morphogenesis of D. pumila
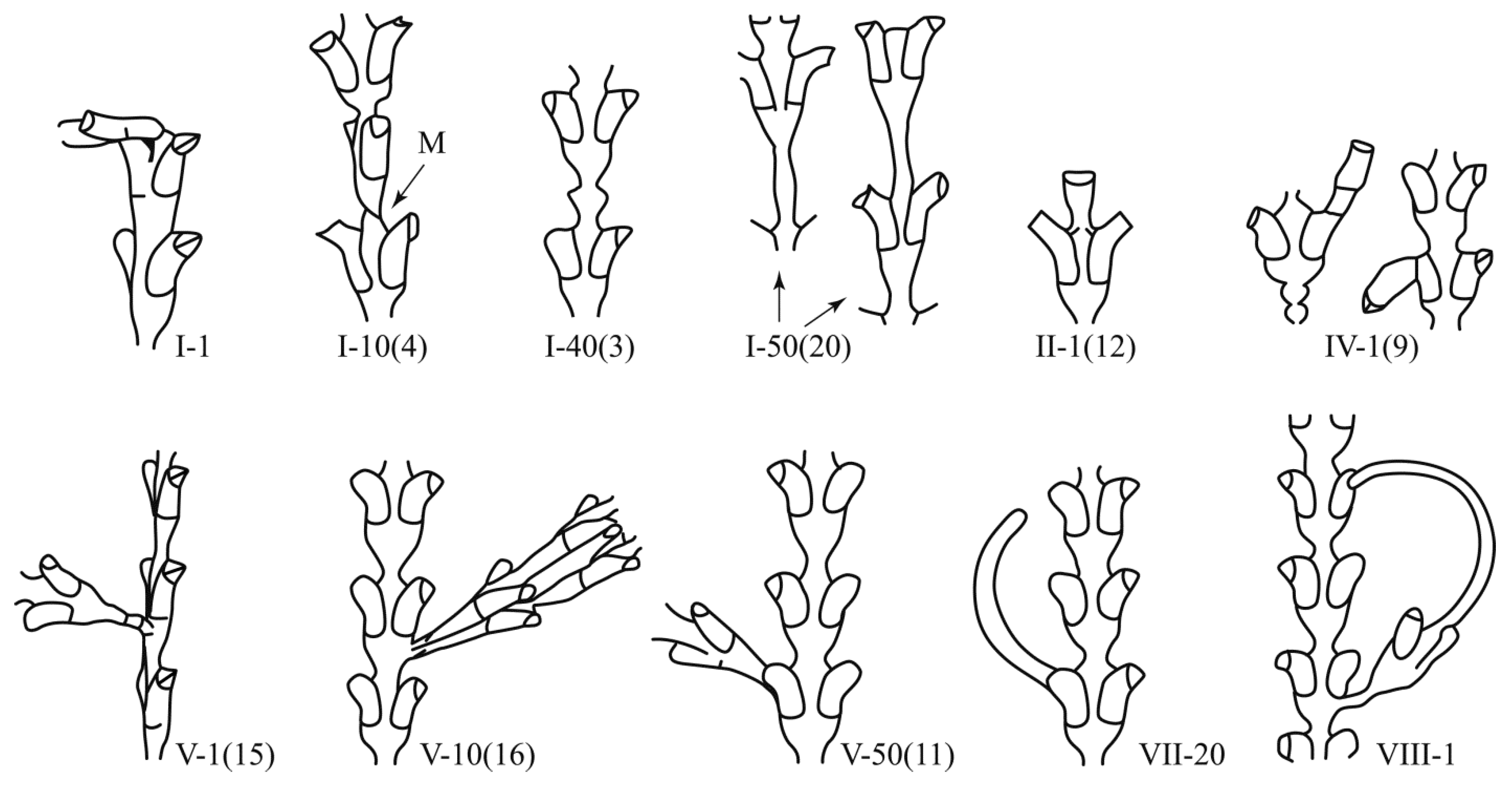
I-1(1). Bending without deformation of the shoot of the hydroid can occur in different directions relative to the longitudinal axis of the shoot (Figure 4). The largest deflection angle is 90°. The place of bending is always the stretch between two successive modules. Presumably, the bending of the shoot or its branch occurs after the suspension of growth when the tip of the cenosarc at the beginning of the next growth period dissolves the chitinous perisarc laterally instead of apically.
1-10(4).The plane of the upper part of the shoot does not coincide with the lower (Figure 4). The turning point is a thin waist between the modules. Rotation can occur at different angles up to 90°. This morphovariation occurs after the suspension of growth and is often accompanied by the formation of an oblique constriction after the beginning of the next growth period. Normally, an oblique constriction is only formed at the shoot rudiment when it appears on the stolon.
1-40(3).Thickening on the trunk (Figure 4). Thickening of the perisarc between two pairs of hydrants. The diameter of the thickening is 1.25–2.0 times the diameter of the constriction. Usually, there is one bead, which is much thinner than two. Thickening, according to its shape and morphological features, is a consequence of the suspension of growth of the shoot tip (not yet divided into three primordia in the upper part), followed by the resumption of growth of only the central portion of the tip.
1-50(20). Cylindrical long constriction between hydrothecae pairs (Figure 4). Normally, the perisarc of the shoot in the area between adjacent pairs of the hydrothecae narrows in the direction of growth, and then immediately expands (see Figure 2). In this case, the shoot remains unchanged for a certain length, forming an elongated cylindrical constriction, which is unusual for a normal shoot module. The shoot may grow like a stolon at first (see below VII-1 (17)).
II-1(12). One hydrant forms at the tip of the shoot (Figure 4). It will be abnormal in shape, with either an increased size or an unusual curvature of the hydrotheca. As a result of this anomaly, the shoot stops growing.
IV-1(9). Elongated hydrotheca (Figure 4). This is not prolonged growth, but a hydrotheca growing on a hydrotheca. The anomaly is the result of the re-development of a hydrant over the mouth of an existing hydrotheca. This is possible due to the periodic resorption of hydrants and their reformation. Normally, this process takes place in D. pumila inside the former hydrotheca. Sometimes, the new rudiment of a hydrant can extend beyond the limits of the old hydrotheca, as a result of which a second hydrotheca is grown partially or completely over it (in the form of a continuation).
V-1(15). The lateral shoot grows perpendicular to the frontal plane of the main shoot (Figure 4). Normally, lateral branches grow only on the lateral sides of the shoot below the bottom of the hydrotheca. Branching in other places should be considered as a separate morphovariation, since both systematic and functional indicators of a shoot adapted to life in a strong current can change at the same time.
V-10(16). Two lateral branches side by side (Figure 4). Both branches are normal in shape and grow from the correct location; however, the formation of two branches in the same location is abnormal. Normally, two branches from one module can only form opposite one another.
V-50(11). A lateral branch grows from the mouth of the hydrotheca (Figure 4). On the hydrotheca, from which a lateral branch has grown, the border of its opening (operculum) is visible. This means that the lateral branch emerged after the hydrant was resorbed. With the continuous growth of the rudiment of the hydrant, a lateral branch of the shoot forms.
VII-20(10). Stolon grows out of a hydrotheca (Figure 4). Clear traces of the upper edge are visible on the hydrotheca. This means that there was a normal hydrant that was then dedifferentiated. With the continuous growth of the rudiment, a stolon was formed.
VII-1(17). The tip of the shoot is transformed into a stolon (Figure 4). The stolon, as a rule, grows forward, bending smoothly to one side, which increases the likelihood of its contact with the substrate. In Diphasia fallax (Johnston, 1847), from the same family, such a change in the differentiation of the shoot tip is common.
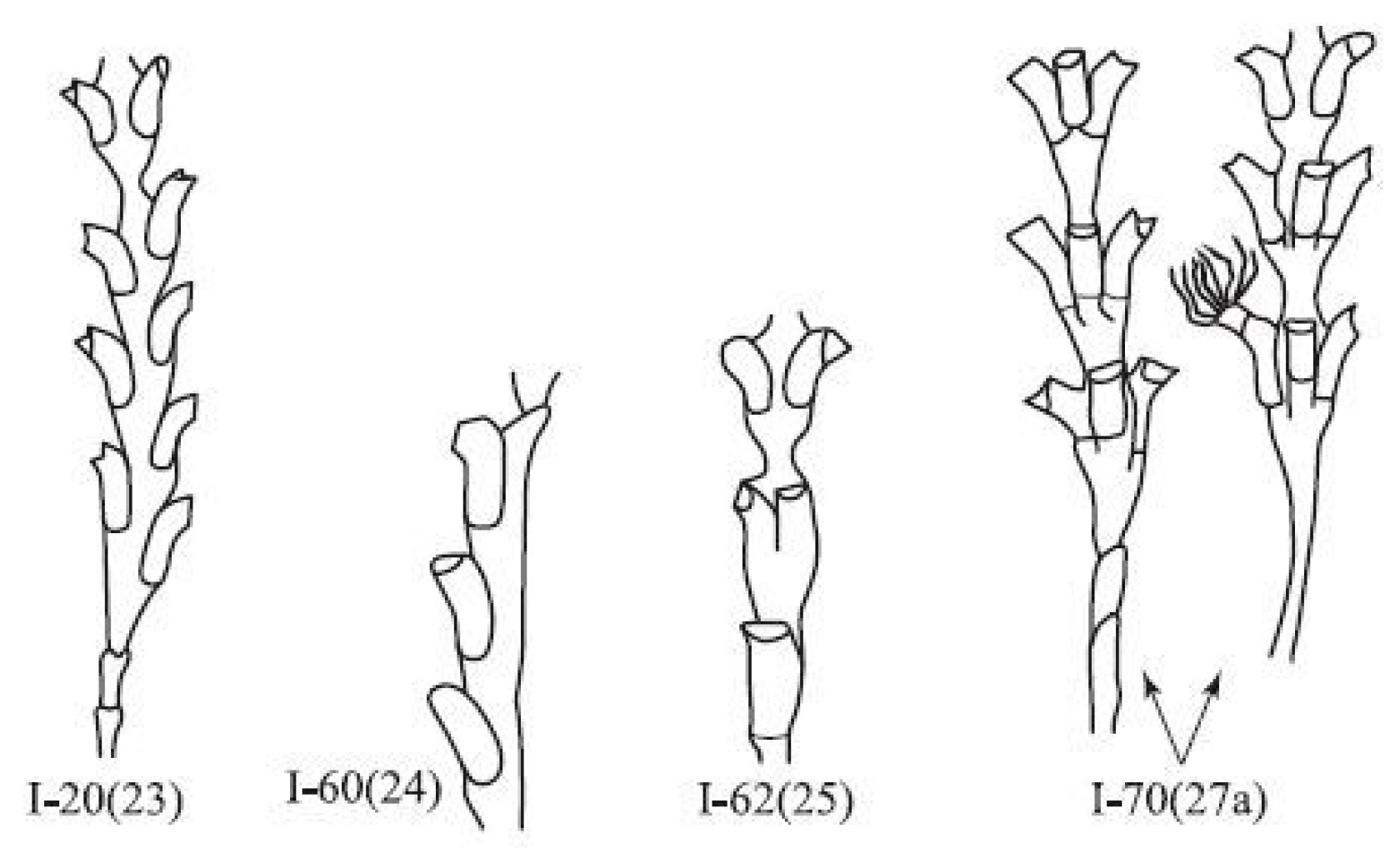
I-20(23). The asymmetric arrangement of the guide-welts (Figure 5). This morphotype involves the vertical displacement of hydrants of one module in relation to each other. Most often, the initial displacement is insignificant; however, if several pairs of hydrants in a row are separated from each other by wide constrictions, then the displacement increases in each successive pair. After the formation of a narrow waist, the displacement disappears. This morphotype is the most common deviation from the canonical structure of D. pumila. It may account for 1/3 to 1/2 of anomalies, which is 0.6–0.7% of all normal modules. Such a seemingly insignificant deviation in the structure of the shoot has, however, a deep morphogenetic significance, since it arises as a result of the asymmetry in the tip of the growth, which is the main path of the evolutionary transformation of the shape of the shoot.
I-60(24). Single-row arrangement of hydrothecae (Figure 5). This anomaly arises in the form of a lateral or, less commonly, main shoot, which consists of one row of hydrants and a cenosarc. The hydrothecae are located on one side of the branch, alternately deployed with their openings to the left and right. Usually, the anomaly repeats over several modules. A similar one-row arrangement of hydrothecae is normally characteristic of another genus (Hydrallmania) in that family. There are no stretches between modules. After the formation of the constriction, the shoot continues to grow normally, forming a two-row arrangement of hydrothecae.
I-62(25). The transition of a single-row arrangement of hydrothecae to a two-row one. First, an unusually large hydrant is formed, then the next module contains a bifurcated hydrant, in the next—two adjacent hydrants and, finally, two normal hydrants opposite one another. The transition can be realized even faster if one of the intermediate stages drops out.
I-70(27). Three-row arrangement of hydrothecae (Figure 5). Here, three normal hydrothecae are located in the modules at the same level, with the same angle (120°) between them. Sometimes, one hydrant is in a different location, which is reflected in a reduction in the contact of the side of one of the hydrants with the coenosarc. The mechanism of this anomaly is not clear. By its recurrence and viability, this anomaly can be classified as a rare morphotype of D. pumila.
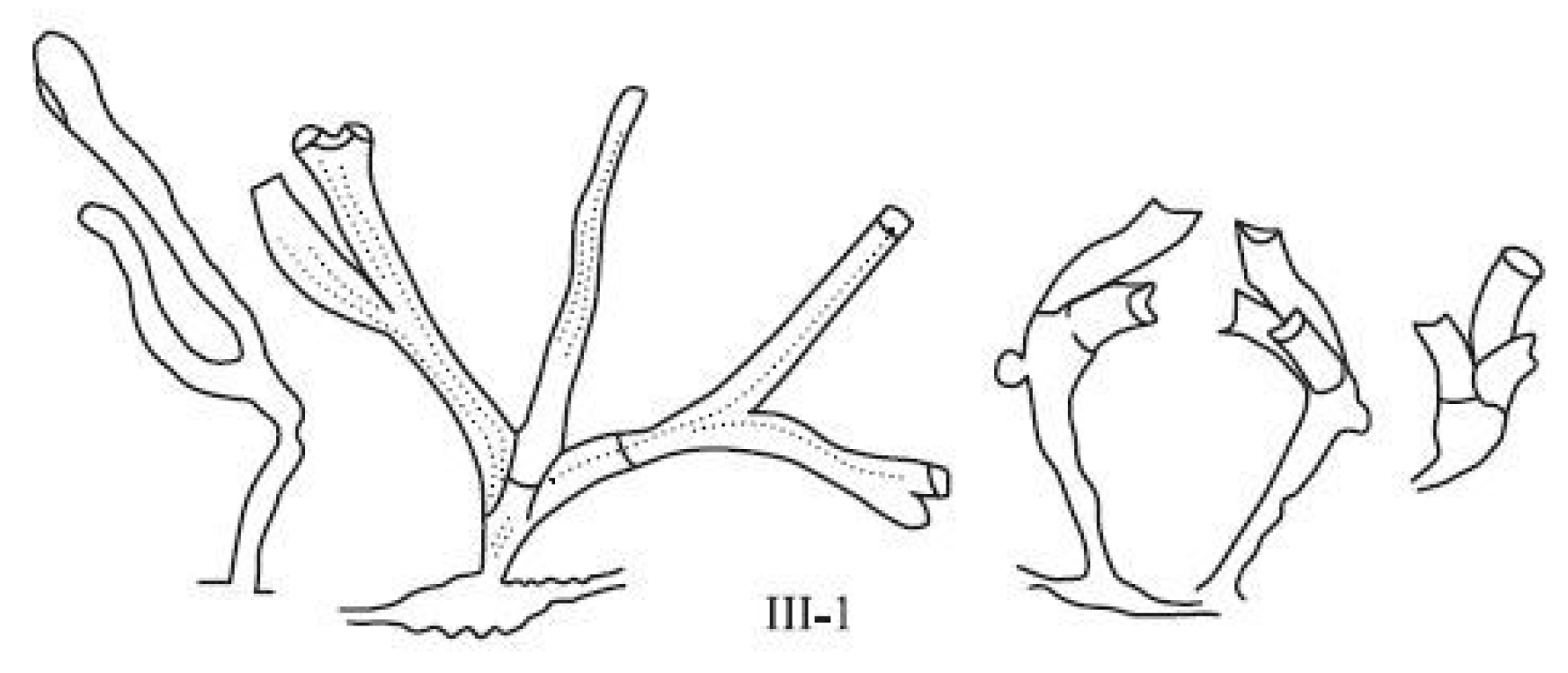
III-1. The aberrant development of the whole shoot (Figure 6). It is difficult to classify developmental anomalies and shoot forms belonging to this group, since they are very diverse, and do not differ in the stability of their multiplication. This group contains a whole set of interrelated deviations in the development of various elements of the first shoot module (Figure 6). The apex is dissected into two, or less often three, primordia.
3.3. Diversity of the Morphovariations
I used phenotypic variations in the structure of the D. pumila shoot for three purposes: (1) quantitative analysis of population variability of the species; (2) studying the reaction of the species to various environmental factors; (3) searching for the limits of the morphogenetic capabilities of the species.
The first two problems allow for a rather formal systematization of anomalies, as long as the classification is not laborious, and the clusters are clearly delineated from each other such that it is easy to apply quantitative methods of comparison. Therefore, excessive detailing is contraindicated to achieving this goal.
For the third task—analysis of the breadth of the morphogenetic capabilities of the species—it is actually more expedient to scrupulously select all the many variants of structure in the spectrum of morphotypes and to carry out the classification in accordance with the proposed mechanisms of morphogenesis. In this case, the number of D. pumila anomalies described above should be approximately doubled.
I chose the first variant for classification and tried to limit the variety of morphogenetic variations, focusing on the defining characteristics indicated in the description of morphogenetic variations. Therefore, when using the classification, it is recommended to reduce many fundamentally indistinguishable varieties to the key morphovariation, thus artificially narrowing the morphogenetic variability.
3.4. Frequency of Occurrence of Aberrant Modules in Shoots of D. pumila
Aberrant modules of the D. pumila shoot occur with an average frequency of 1–2%. The frequency of deviations varies insignificantly from sample to sample, although, within one sample, aberrant modules within the shoot are not very common.
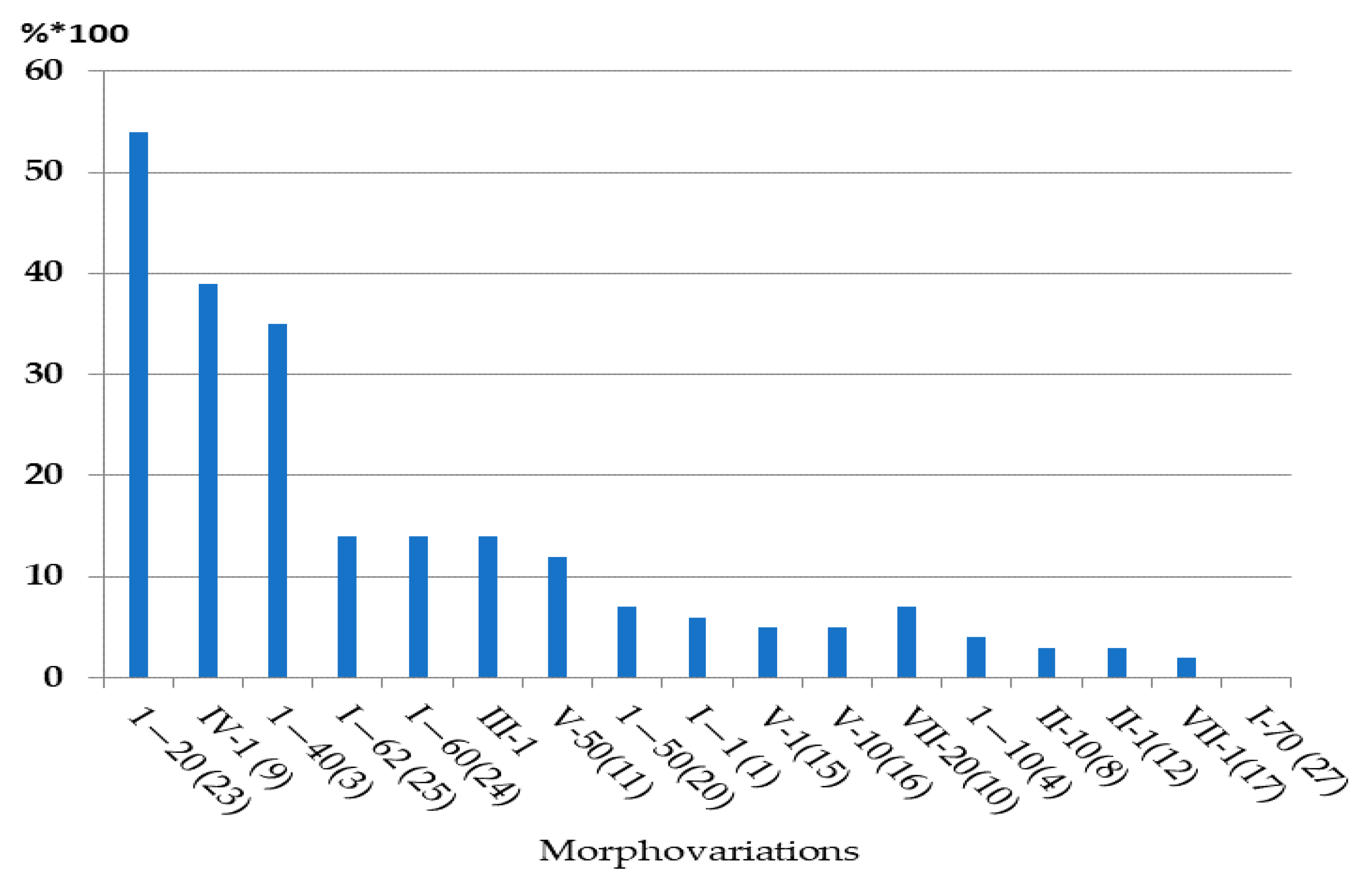
Table 1 shows the averaged values for the frequency of occurrence of the main varieties of aberrant modules, calculated for all samples together. The hierarchical positions of morphovariations in terms of their frequency of occurrence are practically unchanged, as illustrated in Figure 7, taking into account the standard deviations in the frequencies of occurrence of each morphovariation obtained by processing 30 samples.
The most interesting morphovariations are also found at very different frequencies; a single-row arrangement of hydrotheca belongs to the rank of very common aberrations, and a three-row arrangement belongs to the category of very rare aberrations. The difference between them is two orders of magnitude, although these morphotypes differ only in the number of rows, and both appear equally harmonious.
3.5. Location of Aberrant D. pumila Modules in the Shoot
In the colony, a multitude of shoots with dozens and even hundreds of growth tips grow simultaneously at constant rates. If the appearance of morphological aberrations depends on environmental factors, then aberrant modules should appear simultaneously on the tips that are currently forming. Their arrangement on the shoot should, provided they grow at a constant rate, take the appearance of a belt, i.e., they should all appear at the same distance from the tips of the shoot. If the formation of aberrant modules does not depend on environmental factors, then their locations on the shoots should appear random in relation to the tips.
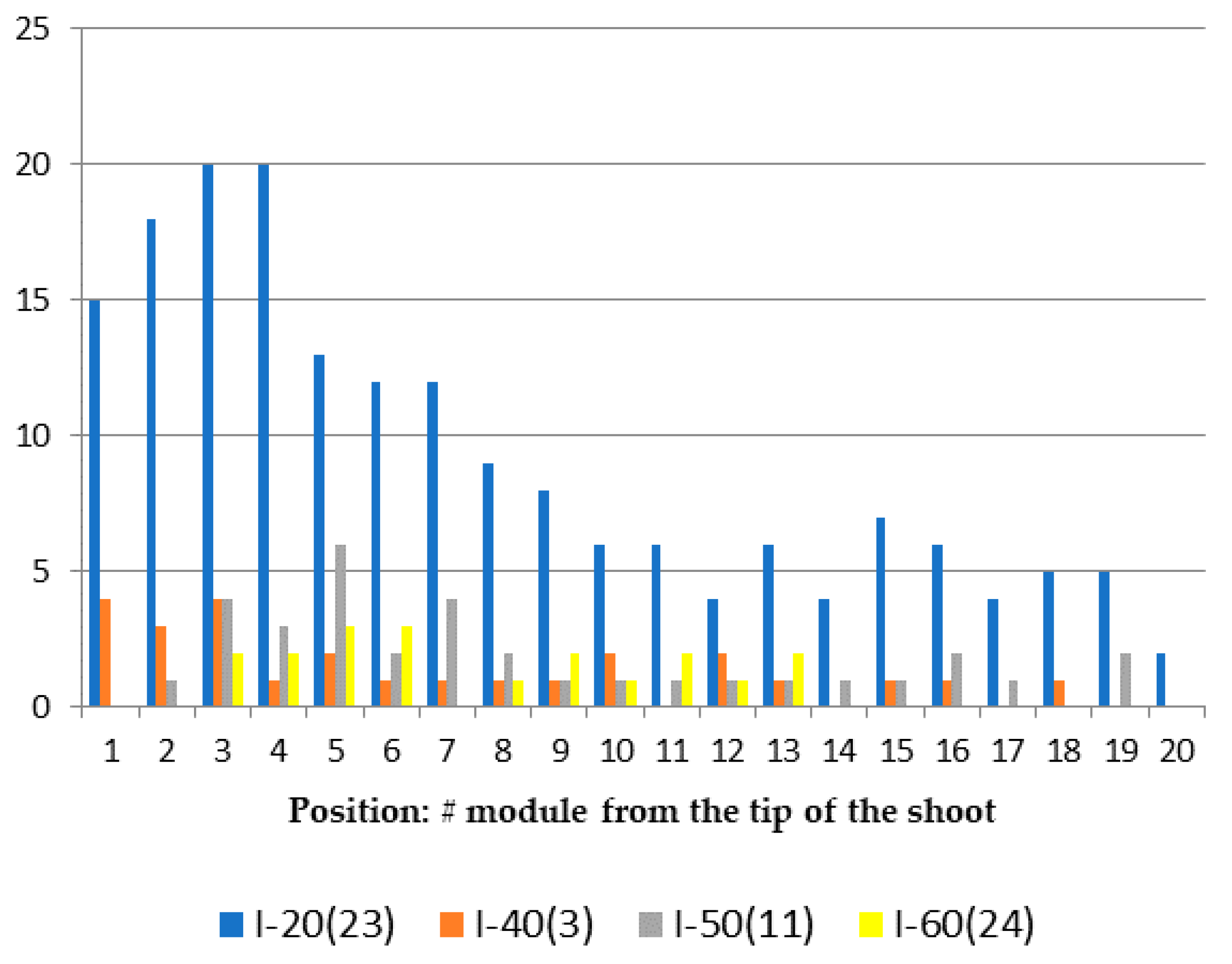
In order to check this assumption, when processing some samples, the positions of aberrant modules on the shoot were noted, counting from the tip. Figure 8 shows the distribution of dominant morphovariations in each position on the shoot. It turned out that morphovariations tend to be located fairly evenly along the shoot, with the exception of one—the asymmetric arrangement of hydrothecae on modules No. I-20(23).
A more detailed analysis revealed that an “asymmetric arrangement of the hydrotheca” is more common at the bases of lateral branches, i.e., in the first modules of the side branches. In the first module from the bottom of the lateral branches, the frequency of occurrence of this morphovariation was 1.16%, while in the second module from the bottom, the frequency of this anomaly was 0.59%, and in the third, it was 0.18% (sample of 7324 modules). This is probably due to the asymmetry in the original bud of a lateral branch that extends from the parent trunk at an angle of about 60°. Therefore, the bilateral symmetry of the apex of the lateral branch at the beginning of its formation is often slightly distorted.
As the morphogenesis of one module is completed and the morphogenetic cycle of the next one begins, some compensation for the asymmetry in the growth apex occurs, since the next cycle begins with a small sphere-like bud, meaning the asymmetry of the previous module is negated. This means that the asymmetry in the growth apex could be a consequence of the initial asymmetry in the primordium arising on the lateral side of the shoot, i.e., due to endogenous causes, most likely epigenetic in nature.
Since the number of short branches in the sample of branched shoots of D. pumila is greater than the number of long branches, the modules located closest to the apex are far greater in number than the modules more distant from the apex, because the latter can be found only on the long lateral branches and main stems of shoots. Therefore, in the diagram of the distribution in the shoot (or branch) of the four most common morphogenetic anomalies and morphotypes, only the asymmetric arrangement of hydrotheca (anomaly No. I-20(23)) is characterized by its frequent occurrence at the first modules (Figure 8).
In general, the data obtained indicate the typically random occurrence of all anomalies (except for No. I-20(23)) on the shoot. This means that the morphovariations can hardly be associated with any external influence, since if they were, more aberrant modules would emerge at the time of occurrence of an external factor, i.e., simultaneously in different shoots and branches. All of them should have arisen at the same distance from the growth tips of the shoots, but this was not found.
4. Discussion
Morphogenetic polyvariance is of great use for understanding the mechanisms of morphogenesis. When abnormalities are found in single organisms, there is usually no doubt that they are the result of genetic mutations. Rare deviations from the norm in a colonial organism are perceived in a fundamentally different way. The abnormal module is preceded by completely normal ones, and after it, the same normal modules occur—in our case, the normal segments of the shoots of the D. pumila colony. Therefore, morphogenetic aberrations cannot be the result of genetic mutations, but at the same time, they do not adhere to the usual ideas about morphological variability. At least six types of morphological variability in organisms are known:
- Sexual dimorphism;
- Polymorphism—the presence of several variants in the zooid’s morphology in the colony (Hydrozoa, Bryozoa);
- Sequential change in phenotypes in the life cycle—for example, in the classes Hydrozoa and Scyphozoa, the formation of a medusoid stage after a polypoid one may occur. In many parasitic organisms, we see phenotype changes during the transition from one stage of the life cycle to the next; in the Insecta class, individual development usually includes metamorphosis. There are many similar examples. Moreover, ontogeny in any species can be represented by a series of phenotypes;
- Intrapopulation morphological variability— this reflects the breadth of variations in morphological characteristics that accumulate in the gene pool of a species during the crossing and exchanging of genetic information;
- Phenotypic plasticity—a change in the shape of an organism or its individual parts in response to a change in the habitat;
- Deformities—deviations in the shape and structure of the body from the norm as a result of genetic mutations, or other causes. Depending on the nature of the mutations, deformities can be inherited or appear in the same generation. In the latter case, one speaks of somatic mutations.
Sexual dimorphism and polymorphism are obligatory and stable manifestations of phenotypic polyvariance fixed in the genotype. Additionally, successive changes in the phenotypes in the life cycle are fixed genetically. In hydroids, this is expressed firstly in a change in polypoid and medusoid generations of many species, and secondly, in the presence of several zooids, such as gastrozooids, gonozooids, dactilozooids, etc. Morphogenetic polyvariance, such as that in D. pumila, does not correspond to the natural manifestation of polymorphism, since morphovariations arise out of connection with the stages of development of the colony or its life cycle [13,14,15].
The intrapopulation variability of morphological characteristics manifests only when comparing organisms with one another. In each individual organism, the phenotype is predetermined by the individual characteristics of the genotype. This does not correspond to cases of aberrant modules being detected in one colony.
Phenotypic plasticity is a consequence of the broad reaction rate of a species. Morphological variations generated by phenotypic plasticity always occur in response to external factors. Phenotypic plasticity has always attracted the attention of researchers, and there is a huge number of publications on this subject.
The phenomenon of morphogenetic plasticity, which we found in the colonial hydroid D. pumila in the form of a deviation from the norm in the structures of shoot modules, cannot be attributed to either polymorphism or phenotypic plasticity. The results of our study of morphovariations in D. pumila indicate that anomalies appear regardless of the state of the environment; they are not sufficiently genetically fixed to manifest in each individual at a certain stage of its individual development. At the same time, morphological abnormalities occur rarely, but with a certain stable frequency. Among the variations in the structure of the module, there are unconditional deformities, but most of the variations are distinguished by a harmonious structure, and in some cases the morphovariations are similar to the phenotypes of the shoot modules of other genera of the same family (Sertulariidae).
Therefore, morphogenetic plasticity is not reducible to the known forms of morphological variations but is an independent phenomenon. Its main feature is that morphogenesis occurs only in some of the many simultaneously forming modules of one organism, while in all the others, morphogenesis proceeds according to the usual program.
To date, it has been established that the presence of alternative morphovariations in D. pumila is not limited to samples collected in the White and Barents Seas, i.e., on the borders of the range of this species. The same morphovariations were also found in colonies collected in the North Sea near Helgoland [16].
After examining samples that include several hundreds of thousands of modules and developing a more complete classification of morphovariations, I came to two general conclusions:
- Despite the dozens of described morphogeneses, the real diversity of deviations in the morphogenesis of modules of the D. pumila shoot is very small; and
- most morphovariations derive from the morphogenesis typical of this type.
At the same time, individual morphovariations are fundamentally different from the norm in terms of the structure and order of module formation. They are very much resembling the structure of the shoot in hydroids from other genera, which may be a manifestation of the species’ ability to implement an alternative morphogenetic program.
In general, all deviations from the norm in D. pumila fit into four groups: (a) deformities; (b) morphogenetic abnormalities resulting from growth arrest; (c) morphogenetic failures in the position of the module; (d) alternative morphogenetic programs or morphotypes.
Such forms have no obvious adaptive value but look viable. If deformity occurs at the tip of the shoot, then the next shoot cannot grow. If the hydrant turns out to be aberrant, then it cannot fulfill its food-producing function.
Morphogenetic anomalies that appear after the suspension of shoot growth look quite viable. These include the bulbous expansion of the trunk, a sharp curvature of the trunk, or a change in the orientation of the plane of the trunk, etc. Resumption of growth can occur on a limited surface of the apex, rather than across its entire working surface, which leads to the formation of unusual shapes, such as bulbous expansions and the bending of the trunk. However, there may be other reasons for the latter anomaly. With a chronic lack of food, a distortion in the proportions of the module is possible; for example, the lengthening of its lower thinnest part, which is associated with an increase in the period of expansion of the apex in the morphogenetic cycle (personal observations).
Morphogenetic mistakes include all cases of incorrect body position when its structure is basically normal, for example, an offshoot or stolon grows instead from the hydrothecae (see morphovariation VII-20(10)); the entire growth tip turns into one hydrant (I-1(12)); the lateral branch departs from the trunk not in its frontal plane, but perpendicular to it (V-1(15)), etc. Often, the growth of an organ in the “wrong” place is associated with growth arrest or the process of secondary morphogenesis after hydrant resorption. Therefore, morphogenetic disruptions can be considered special cases of morphogenetic anomalies. However, for the subsequent comparison of the spectra of morphovariations, it is convenient to consider them separately.
Morphotypes are of particular interest, since, firstly, the appearance of alternative morphotypes is associated with a radical change in the shape of the growth apex and the order of its differentiation into rudiments (buds), and, secondly, the transition to a different type of morphogenesis turns out to be temporary even after several cycles, since in the same shoot normal morphogenesis is restored, and so after unusual modules, completely normal ones emerge.
If the appearance of aberrant modules does not lead to the cessation of shoot growth, then subsequent modules on the same shoots are, as a rule, completely normal. This feature is important to the search for mechanisms of morphogenetic polyvariance.
Deviations from the norm in the development of the organism are associated, first of all, with mutations—genetic or somatic. The appearance of alternative phenotypes in a colony of hydroids is difficult to explain via mutations since along with abnormal modules, there are always completely normal ones in the colony. Usually, in a colony, several tens, or even hundreds, of shoot tips grow at the same time. While in one of these the morphogenesis turned out to differ from the canonical one, in all the others it did not differ. Since the entire colony is genetically identical, there is no reason to assume that the main cause of the appearance of abnormal modules is genetic mutations. Somatic mutations are also not easily defined as the main mechanism of morphogenetic polyvariance. In the apical part of the growth apex—in the zone of morphogenetic transformations—mitoses are absent. The zone of mitotic activity, i.e., cell proliferation, is located at the base of the growth apex. Even if we assume that a somatic mutation occurs in any cell of the growth apex, and its multiplication affects the subsequent process of morphogenesis, then, consequently, the rudiment of the next module, isolated on the apical surface of the growth apex, should consist of the same mutant cells, which means that the next module must be aberrant. However, this is very rare.
Thus, the return to normal morphogenesis after the realization of a different morphogenetic program should be taken to indicate the nonmutational causes of the morphogenetic polyvariance—supporting the hypothesis of the epigenetic origin of deviations in the structures of modules.
So far, three morphotypes have been discovered that resemble the structures of the shoots of three species belonging to different genera of the Sertulariidae family.
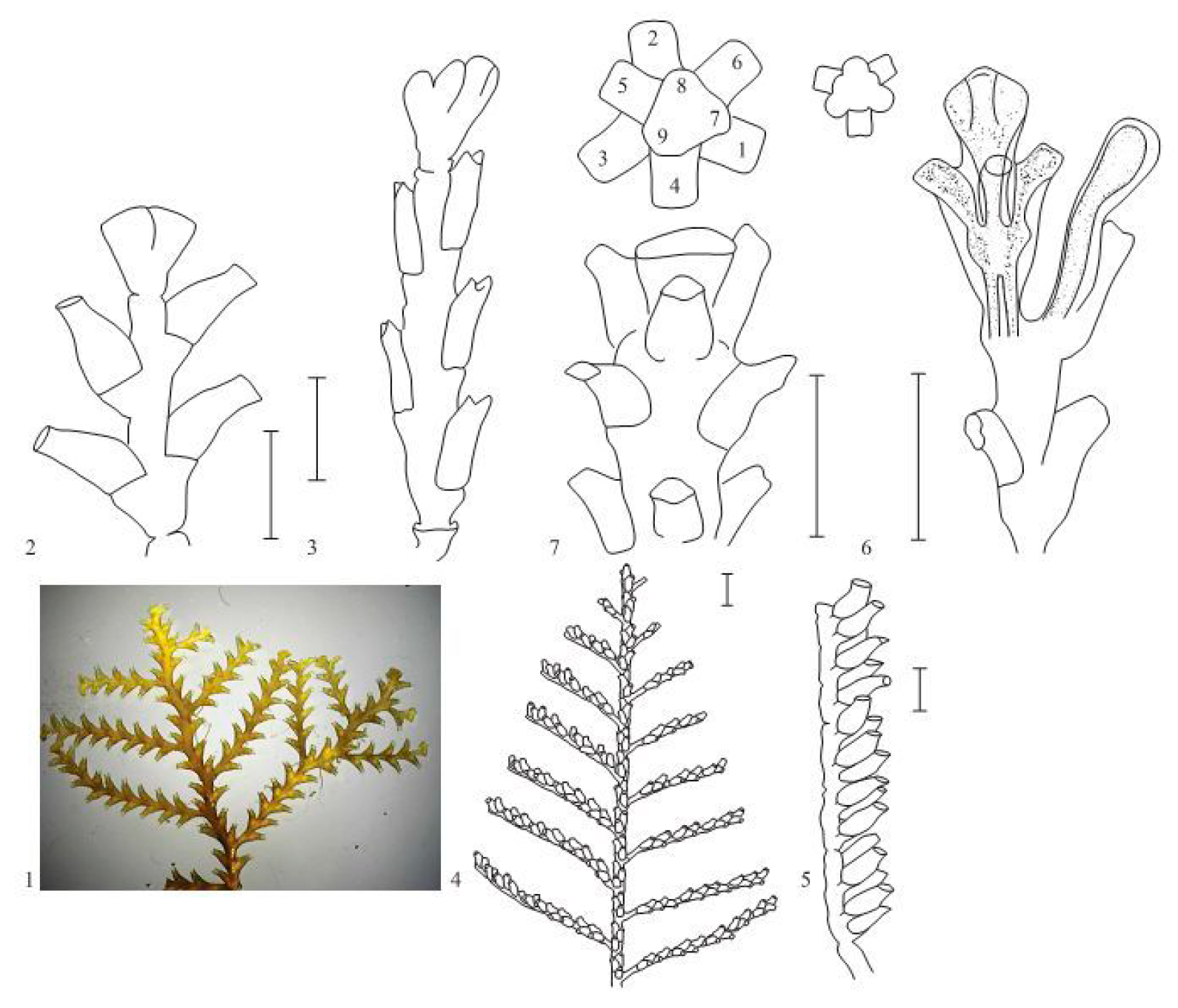
Among all morphovariations, morphotype No. I-20(23) (asymmetric arrangement of hydrothecae) occurs most frequently. This deviation from the norm is very similar to the structures of the shoots in hydroids from the genus Abietinaria. The hydrothecae are located in two rows, similar to in D. pumila, but alternately, i.e., with an offset relative to each other in height. In species from the genus Abietinaria, during the growth of the shoot tip, the primordium of one hydrant is first laid, and then the second primordium is laid on the opposite side of the shoot stem (Figure 9). Similarly, morphogenesis occurs in D. pumila when the strictly opposing arrangement of the location of the hydrothecae is deviated from. Asymmetry in the position of the hydrothecae entails the multiplication of this error over and over again during the formation of the following modules. As a result, a significant part of the shoot may be aberrant, i.e., not typical for D. pumila. Nevertheless, the alternate arrangement of the hydrothecae is usually replaced with an opposite one forming after a narrow constriction (“thin waist”) forms between successive shoot modules. In addition to the position of the hydrothecae, the other distinguishing features of D. pumila, as a rule, do not change. For example, the opercular apparatus consists of two lids, and not one, as in species from the genus Abietinaria. The lateral branches are laid not at the growth tips, as in Abietinaria or Sertularia suensoni Levinsen, 1913 (Figure 9), but instead, emerge after some time under the previously formed hydrothecae. The high frequency of occurrence of morphotype No. I-20(23) is due not only to objective differences from the norm, but also to the difficulty of determining the boundary between the norm and deviations from it since such deviations are not discrete, but gradual.
The situation is completely different for morphotype No. I-60(24) (single-row position of the hydrothecae). This is a pronounced discrete anomaly. In this case, there is no difficulty in distinguishing the anomaly from the norm. Several hydrothecae are formed in a row, on only one side of the shoot. The hydrothecae are alternately oriented with their openings facing opposite directions. This structure is typical for the genus Hydrallmania, in particular for H. falcata. The morphogenesis of the shoot tip with a single-row structure is fundamentally different from the morphogenesis of a two-row shoot. With a single-row arrangement, the division of the growth apex into two primordia occurs not by a transverse partition, but by an oblique one, which divides the apex into two parts of different sizes, related to each other as 1:2. In morphotype I-20(23), the transverse septum is laid either on one side of the apex or on the other, while in Hydrallmania, the hydrothecae are all laid on the same side of the shoot apex (Figure 9), but at alternating angles of tilt (conventionally 45° and 135° relative to the sagittal plane). Notably, in H. falcata, at the beginning of the growth of a young shoot, a two-row arrangement of hydrothecae was identified [17,18,19]. Consequently, the two-row and single-row phenotypes can be programmed in the genotypes of both Hydralmania and Dynamena, although alternative variants are still not species-specific, and occur rarely. The opercular apparatus in the aberrant modules of D. pumila remains typical of this genus.
Apparently, the three-row arrangement of hydrothecae is also programmed in the D. pumila genotype. The combination of the extreme rarity of this morphotype with its striking harmony of form is surprising. This is manifested in the fact that an angle of 120° is strictly maintained between the planes of the hydrothecae, and they are not deformed, retaining the shape typical of D. pumila. Among the species of the Sertulariidae family, we can find some with a three-row arrangement of hydrothecae—if not everywhere, then at least in those with thin branches, such as Pericladium mirabilis (Verrill, 1873) [14,15].
Upon interpreting these facts, we believe that the D. pumila genotype includes additional information on the construction of a viable “spare” form of shoot module. Such phenotypic plasticity within the same organism can be a reserve of adaptive radiation in biological evolution [20].
It can be assumed that the phenomenon of morphogenetic polyvariance should also manifest itself in other modular organisms: corals, siphonophores, bryozoans, camptozoas, colonial ascidians, as well as outside the animal kingdom in many plants and fungi. Of these colonial invertebrates, only corals and bryozoans have been the subject of morphological studies. There are few studies on intracolonial morphovariations in the structure of zooids [21]. The shape variability of corallites is a well-known phenomenon [22,23,24,25]. The structure of the colony itself in corals varies even more—not only in its size and branching sites but also in other features, such as the relationship between the length and thickness of the branch [26,27,28]. However, this phenomenon is always explainable by one of two mechanisms: it is either a reaction to environmental factors or the effect of positional regulation, namely, a variation in the shape of an organ depending on its location in the body.
It would seem that in plants, this phenomenon should have attracted the attention of researchers. However, phenotypic plasticity is traditionally interpreted as the result of a reaction to a change in the external environment, to which hundreds of articles are devoted (of which I cite only a few to illustrate the diversity of research angles [29,30,31,32]). Unfortunately, none of the authors of these works have suggested that deviations in the structures of stereotypical modules in plants can occur on their own without connection with environmental factors or genetic changes. We know of one very significant study of leaf morphovariations, published only in Russian almost a century ago [33].
Indeed, in plants, it is difficult to distinguish changes in the shape of a module, for example, a leaf, resulting from endogenous causes, from those caused by exogenous causes, for example, the intensity or spectral characteristics of light [34,35,36,37,38,39].
Until now, no dependence of the morphovariation frequency in the structure of the shoot module of D. pumila on environmental factors was found. As such, it is important to note that aberrant modules never appear in D. pumila shoots in simultaneous series on parallel-growing shoots, but are instead located “at different levels” from the tips of growth, i.e., form at different times.
The second important feature of morphogenetic polyvariance is the significant consistency in the frequencies of occurrence of different morphogenetic variations. The cumulative frequency of all morphogenetic anomalies together varies within narrow limits, from 0.5 to 4% of the total number of counted modules, but mainly within 0.9–1.5%. The frequencies of occurrence of each morphovariation have also turned out to be limited to narrow intervals. The order of dominance of morphovariations changes little from sample to sample, i.e., the sequence of morphovariation, arranged in decreasing order of their frequency of occurrence within the analyzed sample. Altogether, the CFOA, FOA, and the order of dominance of morphovariations represent a kind of “morphogenetic portrait of the species”, carrying information about its internal morphogenetic potency.
The stability of the frequency of occurrence of the morphovariations found in D. pumila is similar to that in the morphological abnormalities found in Drosophila caused by mutation of the Hsp90 protein. In the mutant heterozygous line, in which the action of the protein is suppressed, various deviations from the norm in the structures of various organs arise [40]. The authors believe that the Hsp90 protein suppresses (buffers) the manifestation of natural morphogenetic variations in the developmental pathways of the organism that remain hidden (Hsp90 buffers the variation affecting the morphogenic pathways that exist in nature, but it is usually silent). With a reduction in the activity of this protein, latent morphogenetic polyvariance occurs, and in the future, under the influence of selection, it can gain a foothold. The frequency of the manifestation of deviations from the norm in the Hsp90 mutant line was 1–2%, and the frequency of occurrence in the offspring indicated genetic fixation. The authors came to the conclusion of the existence of silent genes that predetermine abnormal morphological signs through variations in developmental pathways, using the example of a unitary [41] rather than a modular organism, in which this phenomenon would be easier to detect. The results of studies on morphovariations in the colonial hydroid D. pumila confirm this hypothesis.
Funding
The research was carried out with the financial support of grants from the Russian Foundation for Basic Research (No. 07-04-00736-a, No. 01-04-48798), and also as part of the Scientific Project of the State Order of the Government of Russian Federation to Lomonosov Moscow State University No. 121032300118-0.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The author thanks Elena M. Mayer, Rivva J. Margulis, Denis Nikishin, Tanya Orlova, Anna Sergeeva, Boris V. Yuzenkov for their great help in processing the material; Ninel N. Panteleev from the Murmansk Marine Biological Institute; Yuri B. Burykin from the White Sea Biological Station of Moscow State University for collecting several samples used in this study; Igor A. Kosevich for help at all stages of the study and the discussion of the results; Tomas C.G. Bosch (University Kiel, Germany) for valuable advice while discussing the study results.
Conflicts of Interest
The author declares no conflict of interest.
References
- Marfenin, N.N. Decentralized organism exemplified with colonial hydroid species. Biosphere 2016, 8, 315–337. [Google Scholar] [CrossRef] [Green Version]
- Marfenin, N.N. Abnormalities of the stem shape in the colony of Dynanema pumila (Hydrozoa, Leptolida). In Complex Investigation of an Ocean Nature; Moscow University Press: Moscow, Russia, 1975; Volume 5, pp. 230–239. (In Russian) [Google Scholar]
- Rosen, B.R. Modules, members and communes: A postscript introduction to social organisms. In Biology and Systematics of Colonial Organisms; Larwood, G.P., Rosen, B.R., Eds.; Syst. Ass. Spec. Vol. 11; Academic Press: London, UK, 1979; Volume 11, pp. XII–XXXV. [Google Scholar]
- Ryland, J.S.; Warner, G.F. Growth and form in modular animals: Ideas on the size and arrangement of zooids. In The Growth and Form of Modular Organisms; Harper, J.L., Rosen, B.R., White, J., Eds.; The Royal Society: London, UK, 1986; Volume B 313, pp. 53–76. [Google Scholar]
- Jackson, J.B.C.; Winston, J.E. Modular growth and longevity in bryozoans. In Recent and Fossil Bryozoa; Larwood, G.P., Nielsen, C., Eds.; Olsen and Olsen: Fredensborg, Denmark, 1981; pp. 121–126. [Google Scholar]
- Jackson, J.B.C.; Coates, A.G. Life cycles and evolution of clonal (modular} animals. In The Growth and Form of Modular Organisms; Harper, J.L., Rosen, B.R., White, J., Eds.; The Royal Society: London, UK, 1986; pp. 7–22. [Google Scholar]
- Hyman, L.H. The Invertebrates: Protozoa through Ctenophora; McGraw-Hill: New York, NY, USA, 1940; pp. 400–497. [Google Scholar]
- Naumov, D.V. Hydroids and Hydromedusae of the USSR; Translated from Russian Edition, 1960; Israel Program for Scientific Translations: Jerusalem, Israel, 1969; 660p. [Google Scholar]
- Cornelius, P.F.S. North-West European Thecate hydroids and their medusae. Part 2. Sertulariidae to Campanulariidae. In Synopses of the British Fauna; Barnes, R.S.K., Crothers, J.H., Eds.; New Series; Linnean Society of London and Estuarine and Coastal Sciences Association: London, UK, 1995; pp. 1–386. [Google Scholar]
- Marfenin, N.N. Functional Morphology of the Colonial Hydroids; Zoological Institute RAS: St.-Petersburg, Russia, 1993; 151p. (In Russian) [Google Scholar]
- Marfenin, N.N. Development of the concept of modular organization. J. Obs. Biol. 1999, 60, 6–17. Available online: https://istina.msu.ru/publications/article/2094944/ (accessed on 11 January 2010). (In Russian).
- Marfenin, N.N.; Margulis, R.I.; Majer, E.M. Morphological variability in colonial hydroid Dynamena pumila and classification of observed morphotypes. In Cnidaria. Resent State and Prospects of Research II; Proc. Zool. Inst. RAS: St. Petersburg, Russia, 1995; Volume 261, pp. 71–89. Available online: https://istina.msu.ru/publications/article/2095204/ (accessed on 11 January 2010). (In Russian)
- Agrawal, A.A. Phenotypic plasticity in the interactions and evolution of species. Science 2001, 294, 321–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nijhout, H.F. Development and evolution of adaptive polyphenisms. Evol. Dev. 2003, 5, 9–18. [Google Scholar] [CrossRef] [PubMed]
- West-Eberhard, M.J. Developmental Plasticity and Evolution; Oxford University Press: New York, NY, USA, 2003; 814p. [Google Scholar]
- Berking, S.; Hesse, M.; Herrmann, K. A shoot meristem-like organ in animals. Monopodial and sympodial growth in Hydrozoa. Int. J. Dev. Biol. 2002, 46, 301–308. [Google Scholar] [PubMed]
- Kosevich, I. Branching in colonial hydroids. Branching Morphogenesis; Davis, J., Ed.; Springer US: New York, NY, USA, 2006; pp. 91–112. [Google Scholar]
- Pyataeva, S.V.; Kosevich, I.A. Morphological and anatomical characteristics of the colonial hydroid Sertularia mirabilis (Sertulariidae). Zool. J. 2008, 87, 3–19. (In Russian) [Google Scholar]
- Kosevich, I. Morphogenetic foundations for increased evolutionary complexity in the organization of thecate hydroids shoots (Cnidaria, Hydroidomedusa, Leptomedusae). Biol. Bull. 2012, 39, 172–185. [Google Scholar] [CrossRef]
- Marfenin, N.N. Morphological anomalies as reserve of adaptive radiation in colonial hydroids. In Signs of Vital Activity and Environment’s Dynamic in the Ancient Biotops; Transact. XXX Session of the Soviet Paleontological Society: Kiev, Ukraine, 1988; pp. 149–156. (In Russian) [Google Scholar]
- Boardman, R.S.; Cheetham, A.H.; Cook, P.L. Intracolony variation and the genus concept in Bryozoa. Proc. Natl. Am. Paleontol. Conv. Chicago 1969, Part C, 294–320. [Google Scholar]
- Brakel, W.H. Corallite variation in Porites and the species problem in corals. In Proceedings of the Third International Coral Reef Symposium; University of Miami: Miami, FL, USA, 1977; Volume 1, pp. 457–462. [Google Scholar]
- Foster, A.B. Phenotypic plasticity in the reef corals Montastraea annularis (Ellis & Solander) and Siderastrea siderea (Ellis & Solander). J. Exp. Mar. Biol. Ecol. 1979, 39, 25–54. [Google Scholar] [CrossRef]
- Foster, A.B. The relationship between corallite morphology and colony shape in some massive reef-corals. Coral Reefs 1983, 2, 19–25. [Google Scholar] [CrossRef]
- Shaish, L.; Abelson, A.; Rinkevich, B. How plastic can phenotypic plasticity be? The Branching coral Stylophora pistillata as a model system. PLoS ONE 2007, 2, e644. [Google Scholar] [CrossRef]
- Foidart, J.L. Rapport scientifique de l’expédition belge à la Grande barrière d’Australie en 1967. Madrépores: II. Variations norphologiques intracoloniaires et al.térations du squelette coralliaire. Ânn. Soc. R. Zool. Belg. 1970, 100, 115–128. [Google Scholar]
- Veron, J.E.N.; Pichon, M. Scleractinia of Eastern Australia. Part 1; Monograph Series; Australian Institute of Marine Science: Canberra, Australia, 1976; 86p.
- Veron, J. Overview of coral taxonomy. Zool. J. Linn. Soc. 2013, 169, 485–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, P.A.; Kruuk, L.E.B.; Nicotra, A.B. How to analyse plant phenotypic plasticity in response to a changing climate. New Phytol. 2019, 222, 1235–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laitinen, R.A.E.; Nikoloski, Z. Genetic basis of plasticity in plants. J. Exp. Bot. 2019, 70, 739–745. [Google Scholar] [CrossRef]
- Stotz, G.C.; Salgado-Luarte, C.; Escobedo, V.M.; Valladares, F.; Gianoli, E. Global trends in phenotypic plasticity of plants. Ecol. Lett. 2021, 24, 2267–2281. [Google Scholar] [CrossRef]
- Wells, C.; Pigliucci, M. Adaptive phenotypic plasticity: The case of heterophylly in aquatic plants. Perspect. Plant Ecol. Evol. Syst. 2000, 3, 1–18. [Google Scholar] [CrossRef]
- Krenke, N.P. (Ed.) Phenogenetic Variability. Volume 1; K.A. Timiryazev Biology Institute: Moscow, Russia, 1933; 752p. [Google Scholar]
- Corré, W.J. Growth and morphogenesis of sun and shade plants. I. The influence of light intensity. Acta Bot. Neerl. 1983, 32, 49–62. [Google Scholar] [CrossRef]
- Corré, W.J. Growth and morphogenesis of sun and shade plants II. The influence of light quality. Acta Bot. Neerl. 1983, 32, 185–202. [Google Scholar] [CrossRef]
- Caswell, H. Phenotypic plasticity in life-history traits: Demographic effects and evolutionary consequences. Am. Zool. 1983, 23, 35–46. [Google Scholar] [CrossRef]
- Richards, J.H.; Lee, D.W. Light effects on leaf morphology in Water hyacinth (Eichhornia crassipes). Am. J. Bot. 1986, 73, 1741–1747. [Google Scholar] [CrossRef]
- Goldstein, J.; Wyatt, R.; Richaardson, J. Physiological ecology lab exercise: Phenotypic plasticity in sun and shade leaves. In Tested Studies for Laboratory Teaching; Proceedings of the 21st Workshop/Conference of the Association for Biology Laboratory Education, 1–5 June 1999; Association for Biology Laboratory Education: Plymouth, UK, 2000; Volume 21, pp. 478–481. [Google Scholar]
- Gratani, L. Plant phenotypic plasticity in response to environmental factors. Adv. Bot. 2014, 2014, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Rutherford, S.L.; Lindquist, S. Hsp90 as a capacitor for morphological evolution. Nature 1998, 396, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Schrader, L.; Winter, M.; Errbii, M.; Delabie, J.; Oettler, J.; Gadau, J. Inhibition of HSP90 causes morphological variation in the invasive ant Cardiocondyla obscurior. J. Exp. Zool. B Mol. Dev. Evol. 2021, 336, 333–340. [Google Scholar] [CrossRef] [PubMed]
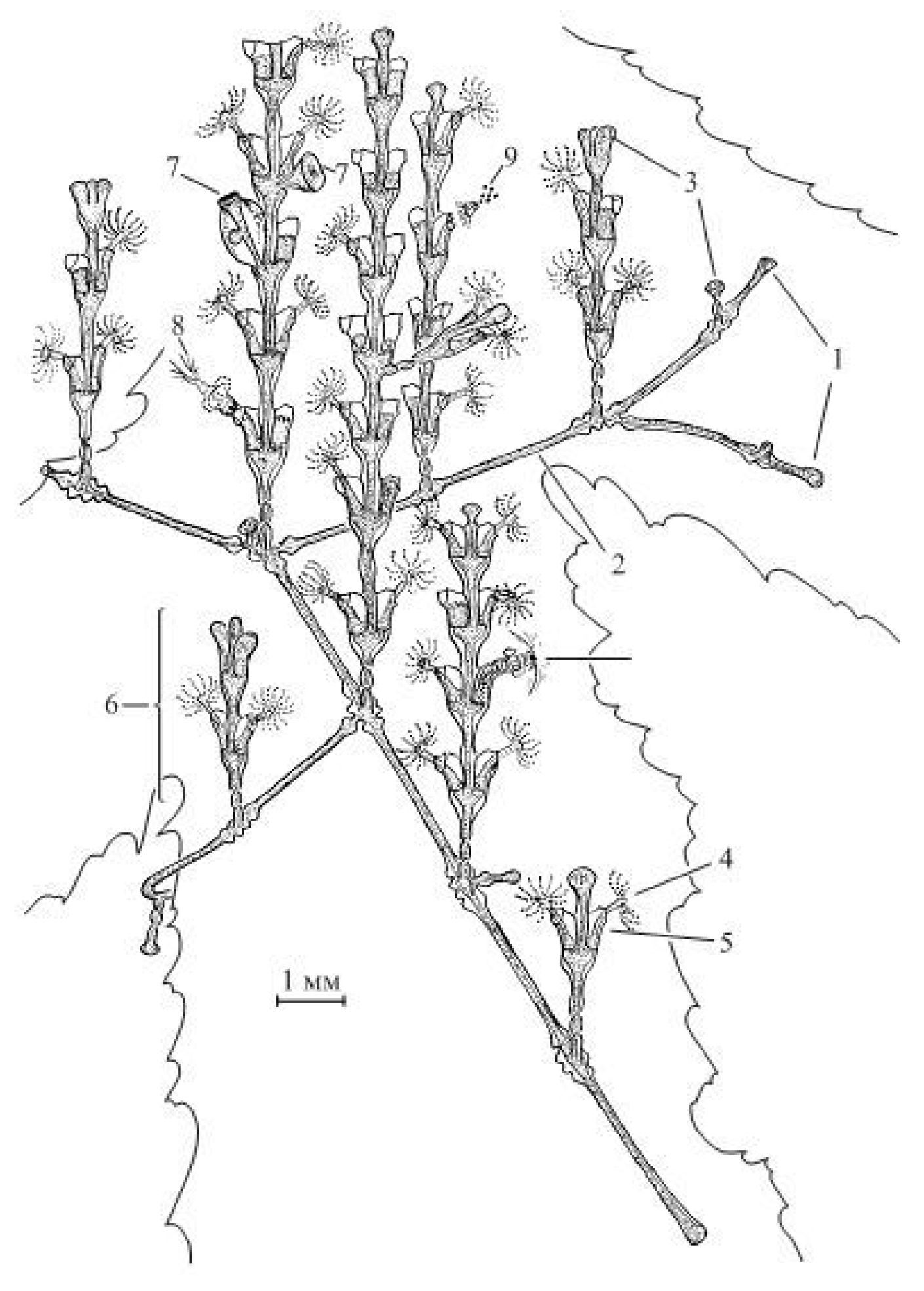
Figure 1.
Colony of the hydroid Dynamena pumila (L.) Legend: 1—tips of the stolon growth; 2—stolons; 3—tips of the shoot growth; 4—hydrant with an open rim of tentacles; 5—hydrotheca; 6—young shoot; 7—gonangium (gonotheca with blastostyle inside); 8—hydrants swallowing the victim; 9—excretion of undigested food debris from the hydrant.
Figure 1.
Colony of the hydroid Dynamena pumila (L.) Legend: 1—tips of the stolon growth; 2—stolons; 3—tips of the shoot growth; 4—hydrant with an open rim of tentacles; 5—hydrotheca; 6—young shoot; 7—gonangium (gonotheca with blastostyle inside); 8—hydrants swallowing the victim; 9—excretion of undigested food debris from the hydrant.

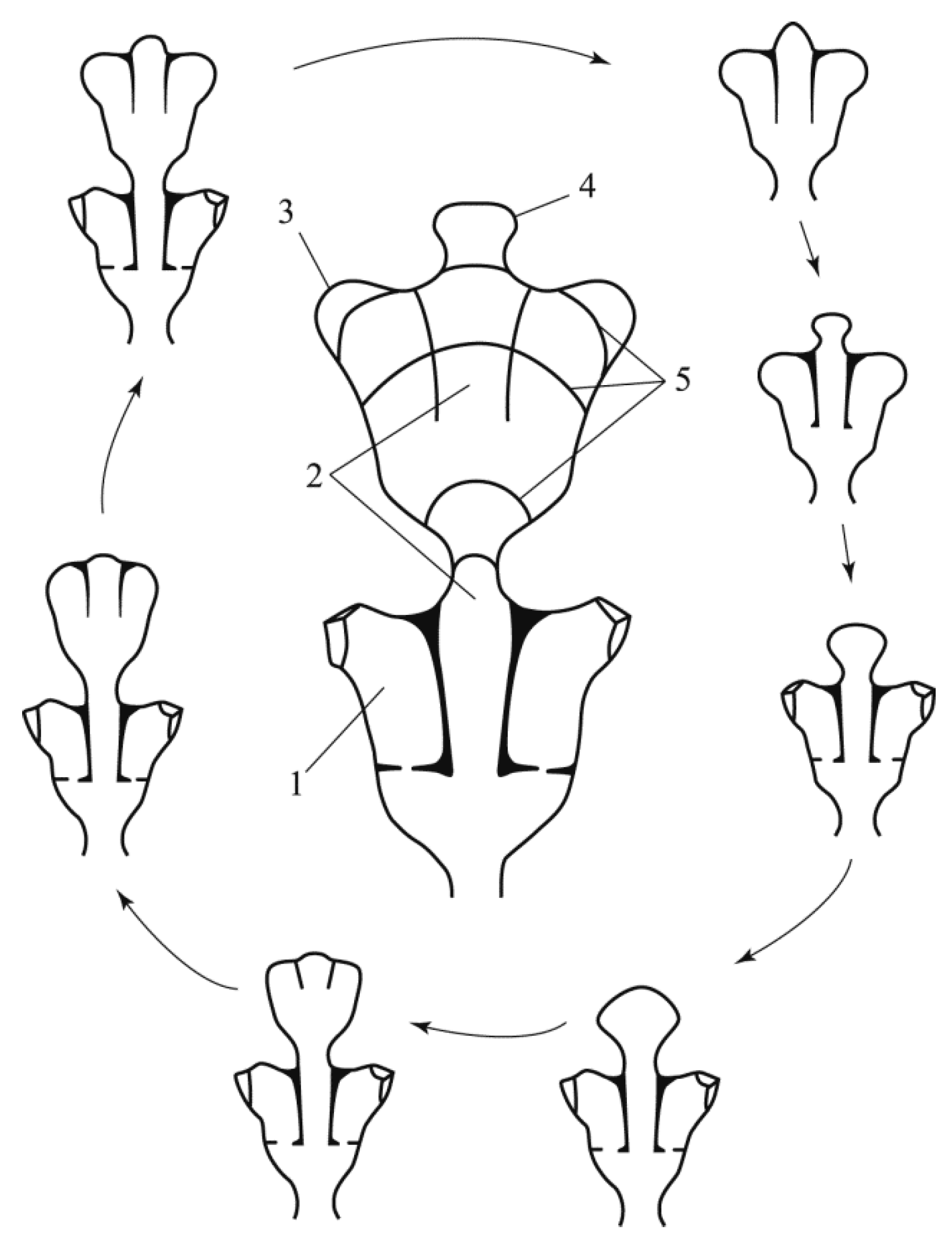
Figure 2.
The morphogenetic cycle of the formation of the Dynamena pumila (L.) shoot module, presented in two equivalent variants: in the central part of the figure, the stages of formation are marked by isolines; in the periphery of the figure, the same stages are shown in a clockwise sequence. Legend: 1—hydrotheca; 2—the central part of the shoot, in which the cenosarc is located; 3—the rudiment (bud) of a hydrant; 4—the tip of the shoot growth at an early stage of formation; 5—isolines marking the growth stages of the shoot apex, and the formation of a module with two oppositely located hydrants in the hydrothecae; the dotted arrow symbolizes the transition to the initial stage of module formation, that is, the closure of the morphogenetic cycle.
Figure 2.
The morphogenetic cycle of the formation of the Dynamena pumila (L.) shoot module, presented in two equivalent variants: in the central part of the figure, the stages of formation are marked by isolines; in the periphery of the figure, the same stages are shown in a clockwise sequence. Legend: 1—hydrotheca; 2—the central part of the shoot, in which the cenosarc is located; 3—the rudiment (bud) of a hydrant; 4—the tip of the shoot growth at an early stage of formation; 5—isolines marking the growth stages of the shoot apex, and the formation of a module with two oppositely located hydrants in the hydrothecae; the dotted arrow symbolizes the transition to the initial stage of module formation, that is, the closure of the morphogenetic cycle.

Figure 3.
Dependence of the proportion (%) of aberrant modules on the sample size (out of the counted modules of Dynamena pumila shoots). Two samples of big shoots (>20 modules each).
Figure 3.
Dependence of the proportion (%) of aberrant modules on the sample size (out of the counted modules of Dynamena pumila shoots). Two samples of big shoots (>20 modules each).

Figure 4.
Variants of deviation from typical shoot morphogenesis of D. pumila.

Figure 5.
Shoot morphovariations of D. pumila, resembling the shoot structure in other genera of the family Sertulariidae.
Figure 5.
Shoot morphovariations of D. pumila, resembling the shoot structure in other genera of the family Sertulariidae.

Figure 6.
Juvenile deformed shoots of D. pumila (group III-1).

Figure 7.
Average frequencies of occurrence of the basic morphovariations of D. pumila’s stem module (30 samples; 339,629 modules).
Figure 7.
Average frequencies of occurrence of the basic morphovariations of D. pumila’s stem module (30 samples; 339,629 modules).

Figure 8.
Dependence of the frequency of occurrence of the most common morphovariations on the position on the D. pumila shoot in a sample of 10,008 modules. The Y-axis shows the number of morphovariations in this position. Different colors are representing four morphovariation (according to nomenclature of Section 3.2).
Figure 8.
Dependence of the frequency of occurrence of the most common morphovariations on the position on the D. pumila shoot in a sample of 10,008 modules. The Y-axis shows the number of morphovariations in this position. Different colors are representing four morphovariation (according to nomenclature of Section 3.2).

Figure 9.
The most interesting morphological variations which looks like similar modules in other genera of hydroids. 1—Alternate hydrotheca pattern in the shoots and branches of Abietinaria abietina (L., 1758) (bar—1 mm); 2—division of the growth tip of the A. abietina shoot into two rudiments (bar—1 mm); 3—laying of a lateral branch at the growth tip of the shoot of Sertularia suensoni Levinsen, 1913 (bar—0.5 mm); 4—lateral branch of Hydrallmania falcata (L., 1758) (bar—1 mm); 5—a fragment of the H. falcata branch, lateral view (bar—0.5 mm); 6—the morphotype I-70(27) of D. pumila (bar—1 mm); 7—the apical part of the lateral branch of Pericladium mirabilis (Verrill, 1873) (bar—1 mm).
Figure 9.
The most interesting morphological variations which looks like similar modules in other genera of hydroids. 1—Alternate hydrotheca pattern in the shoots and branches of Abietinaria abietina (L., 1758) (bar—1 mm); 2—division of the growth tip of the A. abietina shoot into two rudiments (bar—1 mm); 3—laying of a lateral branch at the growth tip of the shoot of Sertularia suensoni Levinsen, 1913 (bar—0.5 mm); 4—lateral branch of Hydrallmania falcata (L., 1758) (bar—1 mm); 5—a fragment of the H. falcata branch, lateral view (bar—0.5 mm); 6—the morphotype I-70(27) of D. pumila (bar—1 mm); 7—the apical part of the lateral branch of Pericladium mirabilis (Verrill, 1873) (bar—1 mm).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The frequency of occurrence of the main morphovariation (FOM, %) in the shoot modules of the colonial hydroid Dynamena pumila (L., 1758), and the frequency of occurrence of each morphovariation (%) in 32 samples from the White and Barents Seas (1991–1994), including 329,629 modules.
Table 1.
The frequency of occurrence of the main morphovariation (FOM, %) in the shoot modules of the colonial hydroid Dynamena pumila (L., 1758), and the frequency of occurrence of each morphovariation (%) in 32 samples from the White and Barents Seas (1991–1994), including 329,629 modules.
| Index (1995) | Name of Morphovariation | The Number of Morphovariations among 329,629 Modules | % of Samples in Which This Morphovariation Was Found | FOM (%) |
|---|---|---|---|---|
| I-20 (23) | Alternating arrangement of hydrants | 1719 | 100 | 0.521 |
| I-40 (3) | Thickening on the trunk | 1186 | 100 | 0.360 |
| IV-1 (9) | Extended hydrotheca | 645 | 97 | 0.196 |
| I-60 (24) | Single-row arrangement of hydrants | 314 | 81 | 0.095 |
| V-50 (11) | Side shoot grows out of the hydrotheca | 299 | 78 | 0.091 |
| I-50 (20) | Cylindrical long constriction | 227 | 78 | 0.069 |
| V-1 (15) | The lateral shoot grows perpendicular to the frontal plane of the main shoot | 174 | 88 | 0.053 |
| VII-20 (10) | Stolon grows from the hydrotheca | 119 | 50 | 0.036 |
| I-62 (25) | Transition of a single-row arrangement of hydrants to a double-row arrangement | 109 | 28 | 0.033 |
| I-1 (1) | Bending of the shoot | 88 | 34 | 0.027 |
| I-10 (4) | The plane of the upper part of the shoot does not coincide with the lower | 76 | 56 | 0.023 |
| II-1 (12) | At the tip of the shoot is a hydrant | 61 | 69 | 0.019 |
| V-10 (16) | Two side shoots side by side | 39 | 19 | 0.012 |
| IV-20a (13) | Instead of a side escape, a hydrant | 35 | 31 | 0.011 |
| IV-10 (21) | One module has two pairs of hydrants | 32 | 13 | 0.010 |
| VII-1 (17) | Lateral shoot goes into stolon | 8 | 22 | 0.002 |
| I-70 (27) | Three-row arrangement of hydrants | 7 | 3 | 0.002 |
| V-30 (7) | Three trunks of the main shoot | 6 | 13 | 0.002 |
| I-71 (14) | The hydrant grows perpendicular to the frontal plane of the shoot | 4 | 13 | 0.001 |
| I-80 (6) | Independent formation of hydrants and cenosarc | 1 | 3 | 0.000 |
| VI-1 (19) | Side shoot looks like a triad of blastostyles | 1 | 3 | 0.000 |
| V-61 (22) | Escape grows from the gonotheca | 1 | 3 | 0.000 |
| I-63 (26) | The transition of a two-row arrangement of hydrants to a single row | 1 | 3 | 0.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Marfenin, N.N. Morphogenetic Polyvariance in the Colonial Hydroid Dynamena pumila (L.). Diversity 2021, 13, 683. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120683
AMA Style
Marfenin NN. Morphogenetic Polyvariance in the Colonial Hydroid Dynamena pumila (L.). Diversity. 2021; 13(12):683. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120683
Chicago/Turabian StyleMarfenin, Nikolay N. 2021. "Morphogenetic Polyvariance in the Colonial Hydroid Dynamena pumila (L.)" Diversity 13, no. 12: 683. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120683
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.