
Genome-Wide Analysis and Expression Profiling of HD-ZIP III Genes in Three Brassica Species

1
Jiangsu Key Laboratory for Biofunctional Molecules, College of Life Science and Chemistry, Jiangsu Second Normal University, Nanjing 211200, China
2
School of Life Sciences and Biotechnology, Shanghai Jiao Tong University, Shanghai 200240, China
*
Author to whom correspondence should be addressed.
Diversity 2021, 13(12), 684; https://0-doi-org.brum.beds.ac.uk/10.3390/d13120684
Submission received: 9 October 2021
/
Revised: 7 December 2021
/
Accepted: 16 December 2021
/
Published: 20 December 2021
(This article belongs to the Special Issue Phylogenetics of Stress Regulators in Plants)
Abstract
:Class III homeodomain-leucine zipper (HD-ZIP III) genes encode plant-specific transcription factors that play pivotal roles in plant growth and development. There is no systematic report on HD-ZIP III members in Brassica plants and their responses to stress are largely unknown. In this study, a total of 10, 9 and 16 HD-ZIP III genes were identified from B. rapa, B. oleracea and B. napus, respectively. The phylogenetic analysis showed that HD-ZIP III proteins were grouped into three clades: PHB/PHV, REV and CNA/HB8. Genes in the same group tended to have similar exon–intron structures. Various phytohormone-responsive elements and stress-responsive elements were detected in the promoter regions of HD-ZIP III genes. Gene expression levels in different tissues, as well as under different stress conditions, were investigated using public transcription profiling data. The HD-ZIP III genes were constitutively expressed among all the tested tissues and were highly accumulated in root and stem. In B. rapa, only one BrREV gene especially responded to heat stress, BrPHB and BrREV members were downregulated upon cold stress and most HD-ZIP III genes exhibited divergent responses to drought stress. In addition, we investigated the genetic variation at known miR165/166 complementary sites of the identified HD-ZIP III genes and found one single nucleotide polymorphism (SNP) in PHB members and two SNPs in REV members, which were further confirmed using Sanger sequencing. Taken together, these results provide information for the genome-wide characterization of HD-ZIP III genes and their stress response diversity in Brassica species.
1. Introduction
HD-ZIP genes encode plant-specific transcription factors that are involved in various biological processes. The HD-ZIP transcription factor is mainly composed of two conserved domains. The homeodomain (HD) is responsible for DNA binding, and a closely linked leucine zipper (LZ) domain is related to protein dimerization [1]. The HD-ZIP family is divided into four subfamilies: HD-ZIP I, HD-ZIP II, HD-ZIP III and HD-ZIP IV based on conserved domains, their gene structures and biological functions [2,3]. HD-ZIP III proteins possess a START domain for N-terminal steroid binding and a C-terminal MEKHLA (Met-Glu-Lys-His-Leu-Ala) domain associated with several chemical and physical stimuli [4,5].
HD-ZIP III genes are evolutionarily ancient and present in all major land plant lineages [6,7]. Studies showed that HD-ZIP III mRNAs share a conserved microRNA binding sequence and are post-transcriptionally regulated by miR165/166 [8,9,10]. Mutations that abolish the miRNA binding sites without changing the amino acid sequence result in dominant phenotypes [8,11,12]. HD-ZIP III proteins are known to control key developmental processes, such as embryogenesis, apical meristem formation, organ polarity establishment and vascular development [13,14,15]. In Arabidopsis thaliana, HD-ZIP III transcription factors comprise five genes: PHABULOSA (PHB)/ATHB14, PHAVOLUTA (PHV)/ATHB9, REVOLUTA (REV), ATHB8 and CORONA (CNA)/ATHB15 [1,16]. These genes play overlapping and antagonistic yet distinct roles during development [8,16,17,18,19]. REV, PHB and PHV are closely related and play partially overlapping functions in the post-embryonic shoot, whereas ATHB8 and CNA antagonize these functions [16]. HD-ZIP III genes determine the xylem type in a dose-dependent fashion and there is no metaxylem in the root vasculature when four of the five genes are mutated [9]. The expression of PHB, PHV and REV are restricted to the adaxial domain of developing leaf primordia and the gain-of-function mutants phb-d and phv-d result in the formation of adaxialized leaves [8,17]. RLD1 encodes an HD-ZIP III protein in maize and a semi-dominant mutation causes the adaxialization or partial reversal of leaf polarity [20]. In rice, OSHB1/OSHB2/OSHB3/OSHB4 are expressed in the adaxial domain of leaves, and the ectopic expression of miR166-resistant OSHB1m, OSHB3m and OSHB5m result in rod or filamentous leaves [21]. The single-nucleotide substitutions in miRNA165/166 binding sites in curly leaf-1 (cul-1) and curly leaf-2 (cul-2) mutants result in a higher expression level of CsPHB and display an upward-rolled leaf phenotype in cucumber [22]. HD-ZIP III genes regulate the wood formation in trees. The Populus CNA ortholog, namely, POPCORONA (PCN), is expressed during secondary growth and plays a role in regulating xylem cell differentiation [23]. PtrHB4 encodes a member of the HD-ZIP III genes and functions in the regulation of interfascicular cambium development [24].
Although HD-ZIP III genes are primarily known to act as core developmental factors, recent studies accumulated evidence showing that they also play an important role in the abiotic stress response [14]. Arabidopsis REV can directly regulate the expression of the auxin biosynthetic enzymes genes TAA1 and YUCCA5 (YUC5), and influence the free auxin levels, which are required for the shade-avoidance response [25]. STTM165/166 plants are more resistant to drought and cold stress in which the expression level of miR165/166 is reduced and target HD-ZIP III genes are elevated [26,27]. miR166 mediates the response to drought stress via leaf rolling and altering stem xylem development in rice [28]. A subset of HD-ZIP III genes in soybean, cassava, wheat, potato, Medicago truncatula and cucumber also respond to various abiotic stress treatments including cold, salt and drought [29,30,31,32,33,34]. These results suggest that HD-ZIP III may also participate in the abiotic stress response.
The Brassica genus contains economically important crops with valuable products that provide vegetables, oil, condiments and vitamin C [35]. B. napus (AACC, 2n = 38) is a typical allotetraploid of the Brassica genus, which was formed via natural hybridization and polyploidization of B. rapa (AA, 2n = 20) and B. oleracea (CC, 2n = 18) [36]. The production and quality of Brassica crops are affected by adverse environmental conditions. Through genome-wide analysis, many HD-ZIP families were identified in various plant species [29,30,31,32,33,34,37,38,39,40,41,42,43,44]. However, little is known of the HD-ZIP III members in Brassica plants and their responses to stress are less explored.
In this study, a comprehensive investigation was performed in three Brassica species and the HD-ZIP III gene members were identified in B. rapa, B. oleracea and B. napus. The protein properties, chromosomal localization, phylogenetic relationships, gene structures, conserved motifs and cis-elements in the promoter region were analyzed, and the gene expression levels in various tissues and different stress conditions were investigated. Moreover, the genetic variation of miR165/166 binding sequences at the HD-ZIP III genes was surveyed and verified using Sanger sequencing in B. rapa. Analysis of the HD-ZIP III genes provides useful clues to the functional research of its members in Brassica crops.
2. Materials and Methods
2.1. Identification of HD-ZIP III Family Genes in B. rapa, B. oleracea and B. napus
HD-ZIP III proteins of Arabidopsis were downloaded in the TAIR database (http://www.arabidopsis.org/, accessed on 7 July 2021). HD-ZIP III proteins of rice, maize and tomato were obtained from previous studies [37,38,40]. BLASTP was made using these protein sequences as a query with an e-value < 1 × 10−10 against the B. rapa (version 3.0), B. oleracea (version 1.1) and B. napus (version 5.0) genomes in the BRAD database (http://brassicadb.cn/#/BLAST/, accessed on 20 November 2021) [45]. The obtained protein sequences were further examined using Pfam (https://pfam.xfam.org/, accessed on 20 November 2021) and CD search (https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/Structure/cdd/wrpsb.cgi, accessed on 20 November 2021) to detect the conserved domains. The candidates were used to search against the GenBank non-redundant protein database (nr). Finally, the incorrect sequences were removed manually. Coding sequences and protein sequences that were homologous in B. rapa, B. oleracea and B. napus were retrieved from the BRAD. The ExPASy proteomics server database (https://web.expasy.org/protparam/, accessed on 14 August 2021) was used to predict the protein molecular weights, theoretical isoelectric point, grand average of hydropathicity (GRAVY) and protein stability of the HD-ZIP III proteins.
2.2. Chromosomal Location, Gene Structure, Conserved Motif and Domain Analyses
The gene positions and structural information were extracted from the GFF3 file (http://www.brassicadb.cn/#/Download/, accessed on 27 July 2021). The chromosomal location of HD-ZIP III genes was constructed with MapGene2chromosome V2 (http://mg2c.iask.in/mg2c_v2.0/, accessed on 30 September 2021) based on their position information. Gene structure was analyzed using TBtools according to the information from the GFF file [46]. Conserved motifs of the HD-ZIP III proteins were determined using the MEME program (https://meme-suite.org/meme/index.html, accessed on 25 September 2021) and visualized with TBtools.
2.3. Multiple Sequence Alignment and Phylogenetic Analysis of HD-ZIP III genes
HD-ZIP III amino acid sequences were subjected to multiple sequence alignments using Clustal W with default parameters. The evolutionary relationships were analyzed using MEGA (version 7.0) [47]. A phylogenetic tree was constructed with the value of the 1000 bootstrap replications using the neighbor-joining (NJ) method.
2.4. Cis-Elements Analysis in the Promoters of HD-ZIP III Genes
For the identification of cis-elements in the promoters of HD-ZIP III genes, the 2 kb regions upstream of the start codon were retrieved from the Brassica database (BRAD). The regulatory elements were analyzed using the PlantCARE webtool (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/, accessed on 20 August 2021) [48] and were presented with TBtools.
2.5. Expression Profiling of HD-ZIP III Genes in Different Tissues and Stress Conditions
The RNA-Seq expression data of B. rapa and B. oleracea were retrieved from the Gene Expression Omnibus (GEO) database via accession numbers GSE43245 and GSE42891, respectively [49,50]. The expression values of the HD-ZIP III genes of B. rapa and B. oleracea across six tissues (callus, root, stem, leaf, flower and silique) were obtained and analyzed. The expression data of B. napus were obtained from the Brassica Expression Database (BrassicaEDB) using bolting stage tissues [51]. Heatmaps were constructed using TBtools. The expression profiling of HD-ZIP III genes under abiotic stress was analyzed based on the transcriptome data of B. rapa under a high temperature of 45 °C [52], low temperature of 4 °C [53] and drought [54]. Heatmaps were generated with the pheatmap package in R using the log2-based fold change values of the treatment/control.
2.6. Prediction of miR165/166 and Comparison of miR165/166 Targeting Sites of HD-ZIP III Genes
The precursors of miR165/166 sequences in B. rapa were obtained from a previous study [55] and subjected to the RNAfold web server to predict secondary stem-loop structures with default parameters (http://rna.tbi.univie.ac.at/cgi-bin/RNAWebSuite/RNAfold.cgi, accessed on 6 December 2021). Multiple alignments of HD-ZIP III genes at the miR165/166 targeting sites were performed using DNAMAN software.
2.7. Plant Materials, Growth Conditions and RT-PCR
Three cultivars of B. rapa, namely, one heading Chinese cabbage (Bre) and two non-heading Chinese cabbage (HQ and ZCT), were used. Plants were grown in a greenhouse (22 °C) under long-day conditions (16 h light/day). Leaves were collected from 6-week-old plants. The samples were frozen in liquid nitrogen. Total RNA was extracted from the collected samples using the Trizol reagent (Invitrogen) according to the manufacturer’s instructions. Contaminating genomic DNA was removed with DNase I. The gene-specific primers were used to amplify the HD-ZIP III gene fragments. All the primers are listed in Table S1 in Supplementary Materials. PCR was carried out using KOD-Plus-Neo (TOYOBO). The reaction mixture included 5 μL PCR buffer, 5 μL dNTP, 3 μL MgSO4, 1.5 μL forward primer, 1.5 μL reverse primer, 2 μL cDNA template, 1 μL KOD-Plus-Neo and 31 μL nuclease-free H2O in a total volume of 50 μL. The PCR cycling conditions were as follows: pre-denature at 94 °C for 2 min, followed by 30 cycles of 98 °C for 10 s, 54 °C for 30 s and 68 °C for 10 s. The PCR products were purified and sequenced.
3. Results
3.1. Identification and Characterization of HD-ZIP III Genes in B. rapa, B. oleracea and B. napus
HD-ZIP III coding sequences from Arabidopsis, rice, maize and tomato were obtained and used as a query against the Brassica database using BLASTP. Thirty-five genes were identified, namely, 10 from B. rapa, 9 from B. oleracea and 16 from B. napus, and they were named after their Arabidopsis homologs. Except for BoPHV and BrPHV, there were at least two orthologs for each Arabidopsis HD-ZIP III gene. The physical and chemical properties of the HD-ZIP III proteins are summarized in Table 1. The lengths of the proteins ranged from 630 amino acids (aa) to 865 aa, with molecular weights from 69.27 kDa to 94.65 kDa. The theoretical isoelectric point (PI) ranged from 5.58 to 6.46. The GRAVY value for all studied genes was below −0.068 to −0.276, indicating all the studied proteins were hydrophilic. All the proteins had an instability index exceeding 40, implying that they could be unstable.
3.2. Chromosome Localization of the HD-ZIP III Genes
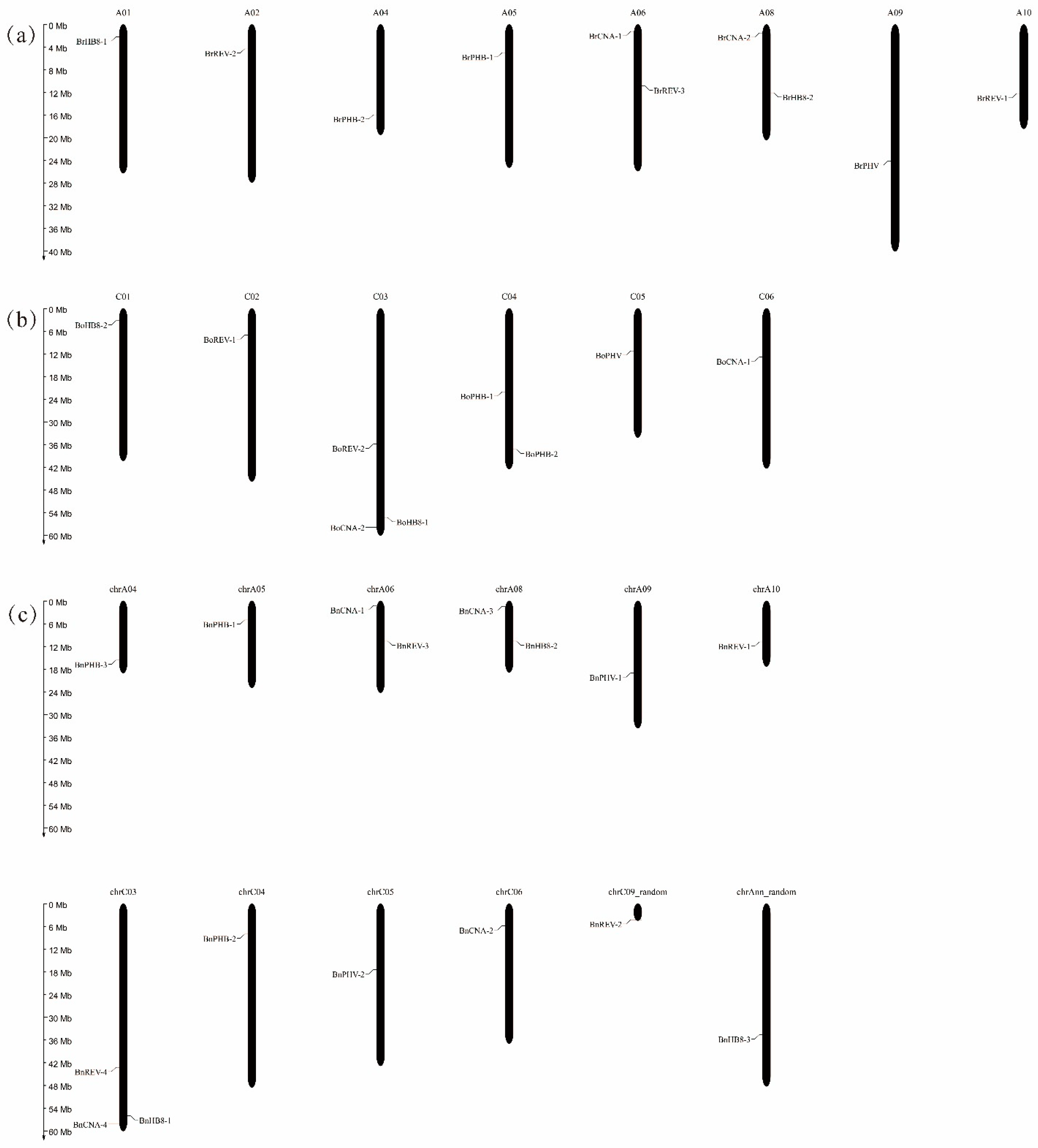
The 35 HD-ZIP III genes from B. rapa, B. oleracea and B. napus were mapped to chromosomes based on their physical position in the GFF3 files (Figure 1). The numbers of these genes in specific chromosomes ranged from one to three. In B. rapa, the chromosomes A06 and A08 each contained two genes, while the remaining six genes were distributed on individual chromosomes. In B. oleracea, nine genes were positioned on six chromosomes. Fourteen genes of B. napus were mapped to ten chromosomes and the other two genes were assigned to the chrC09 random chromosome and Ann (a random A-genome chromosome). Among them, nine genes were positioned in the A sub-genome and seven genes were positioned in the C sub-genome. This result suggested that these HD-ZIP III genes had uneven distributions in the three species.
3.3. Phylogenetic Analysis of HD-ZIP III Genes
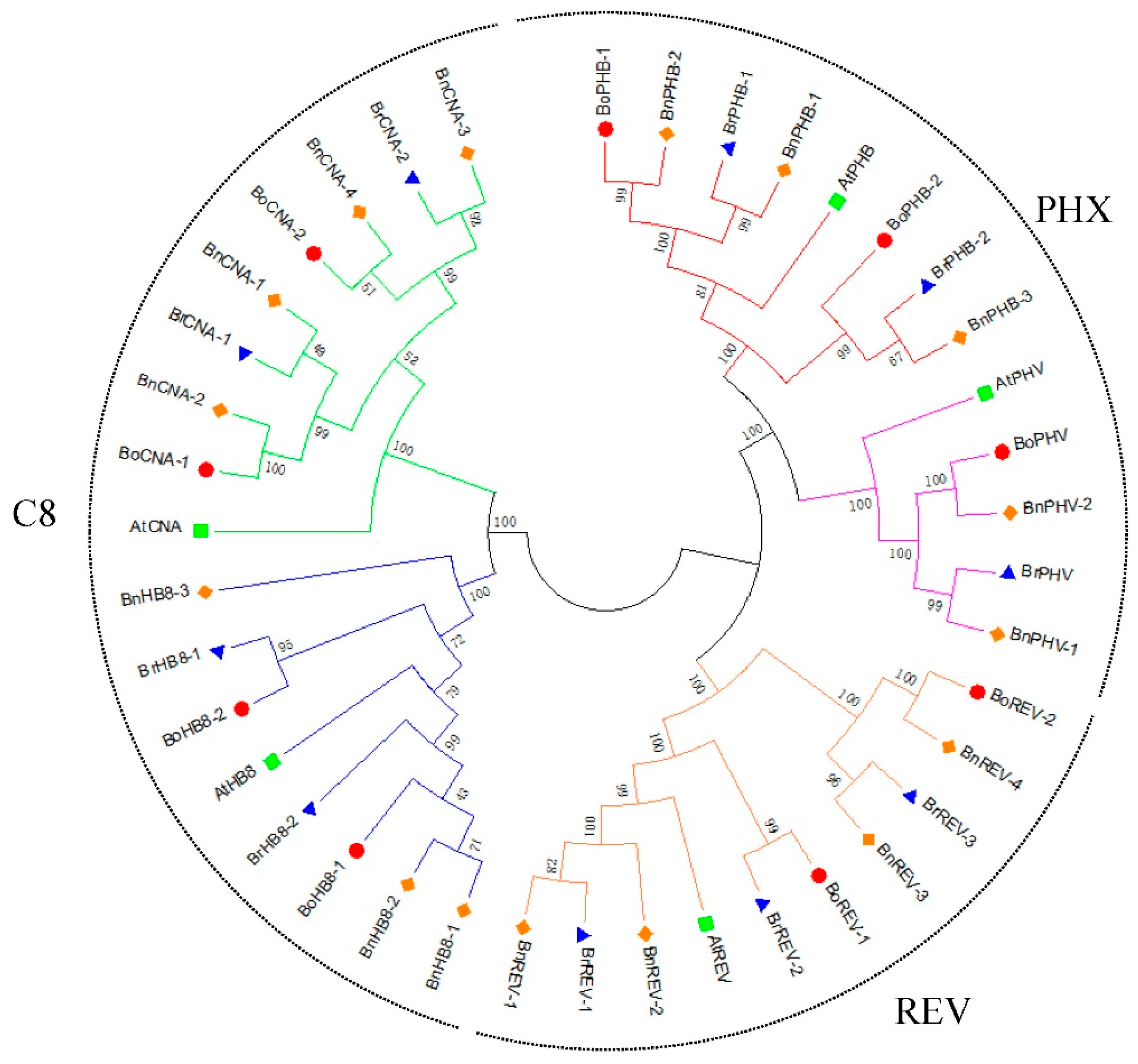
To gain insight into the phylogenetic relationships of the HD-ZIP III genes from A. thaliana and the three Brassica species, a phylogenetic tree was built based on protein sequences using the neighbor-joining method. The phylogenetic tree divided these proteins into three clades (Figure 2). One clade contained all the REV-related proteins. The PHB and PHV proteins had a close relationship and made a PHX (PHB/PHV-related) clade. Members from HB8 and CNA were closer and they made a C8 (CNA/HB8-related) clade. Genes from B. napus were closely associated with the corresponding genes in B. rapa and B. oleracea in each clade. The results correspond with previous studies on plant HD-ZIP III genes [7,23].
3.4. Gene Structure, Conserved Motifs and Domains of HD-ZIP III Genes
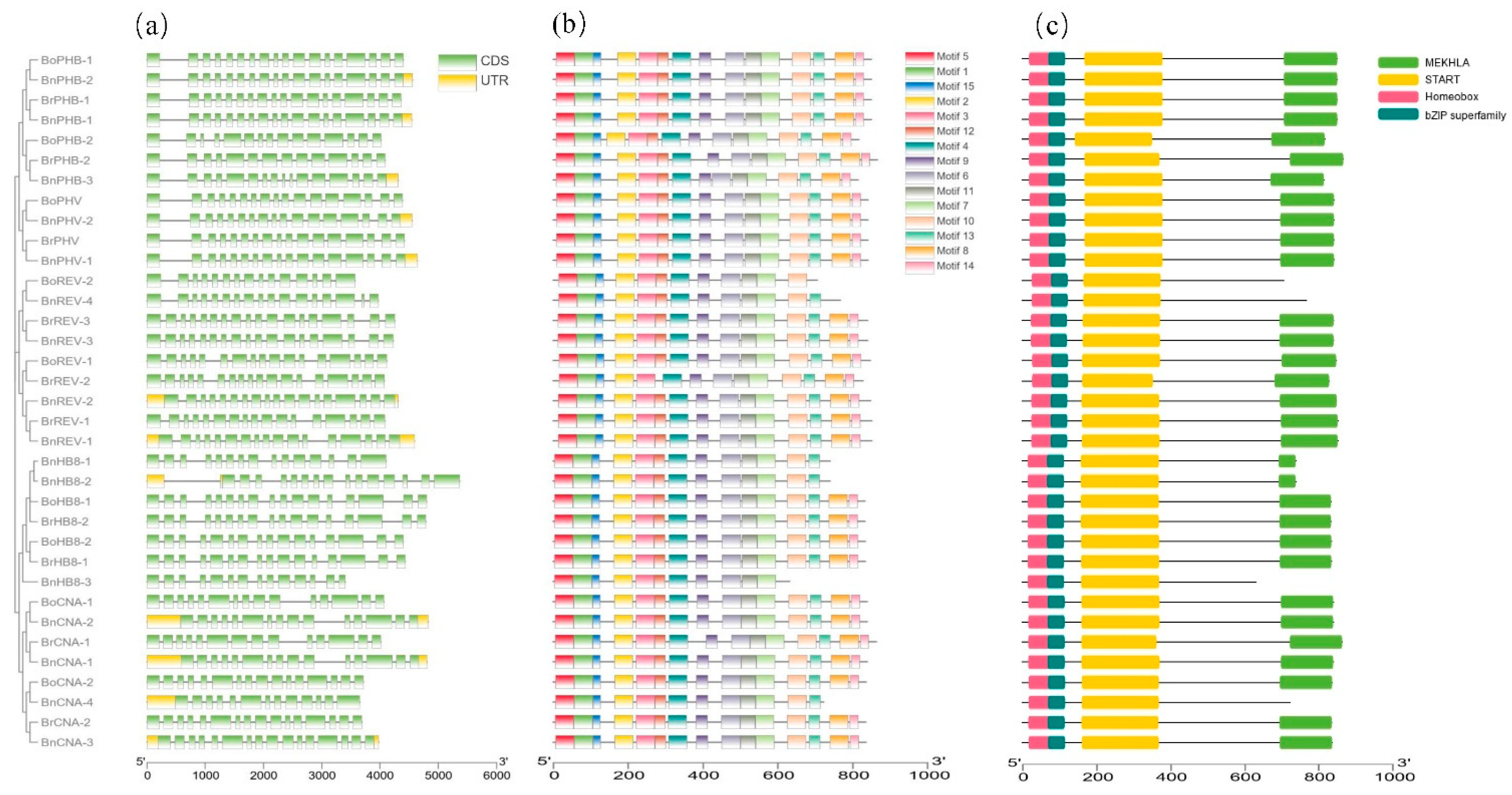
To investigate the structural features of HD-ZIP III genes, the exon–intron organization was analyzed. As shown in Figure 3a, the number of exons in the HD-ZIP III genes varied from 13 to 18. The majority of the homologous genes shared the same gene structure. Genes that were clustered together had similar exon–intron structures. Then, the protein sequences were subjected to the MEME tool and 15 conserved motifs were analyzed, which were designated as motifs 1 to 15 (Figure 3b and Figure S1). BrREV-2 missed motif12; BnREV-4, BnHB8-1, BnHB8-2 and BnCNA-4 missed motif8/14; BoREV-2 missed three motifs; and BnHB8-3 missed four motifs. This was probably due to the relatively shorter length of these protein sequences. The remaining proteins shared all the 15 conserved motifs. Conserved domain analysis showed that 31 HD-ZIP III proteins shared four domains, namely, homeobox, bZIP, START and MEKHLA domains, except that BoREV-2, BnREV-4, BnHB8-3 and BnCNA-4 lacked the MEKHLA domain, which was consistent with the motif analysis (Figure 3c). These findings indicated that the HD-ZIP III genes were conserved in the three Brassica species.
3.5. Analysis of the Cis-Elements in the HD-ZIP III Promoters
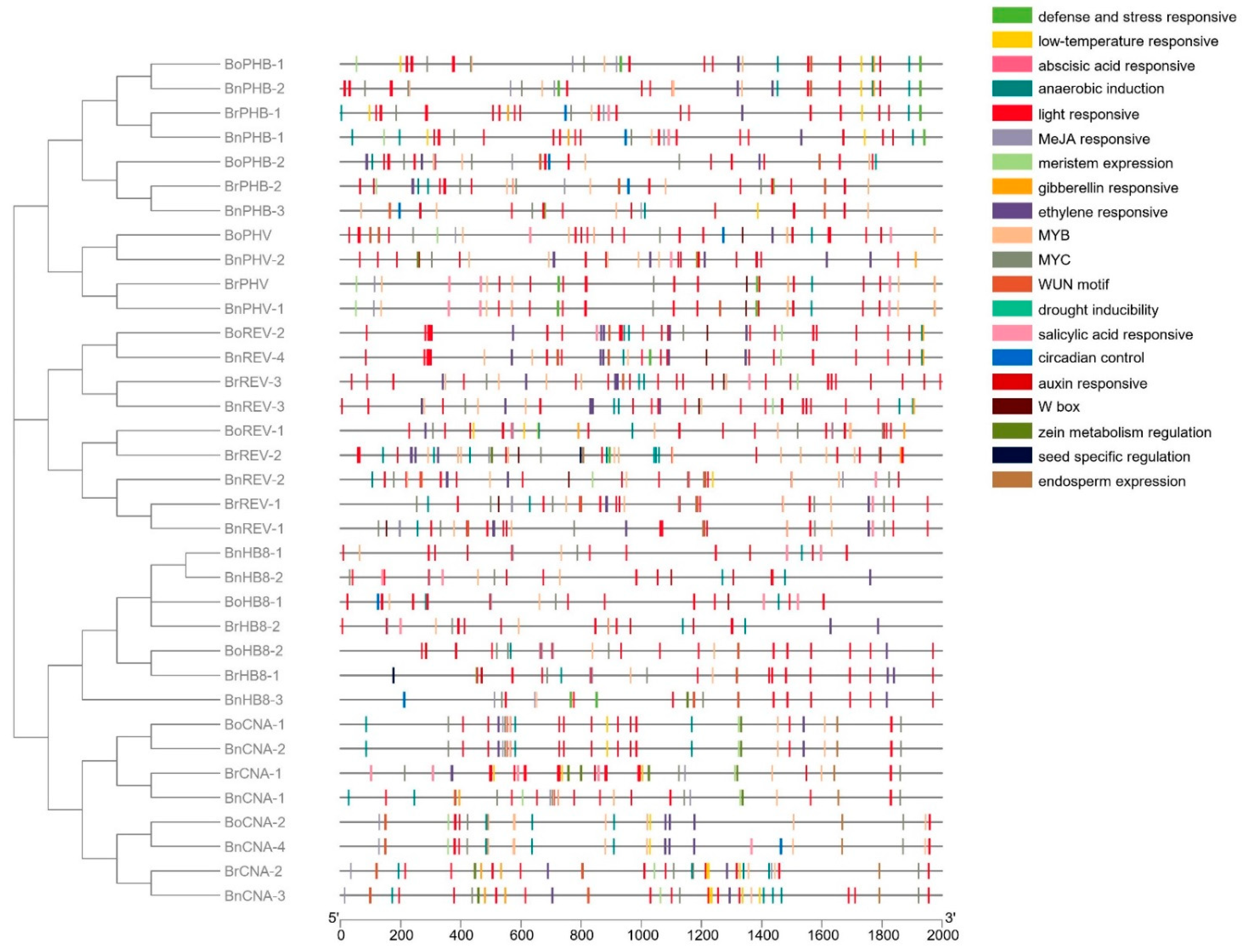
The cis-elements of promoters are involved in the regulation of gene expression and controlling various biological processes [56]. The 2 kb genomic sequences upstream of the start codon were retrieved from BRAD and the cis-regulatory elements were identified using the PlantCARE database (Figure 4, Table S2). The most predominant cis-elements were related to light responsiveness and a total of 427 elements were associated with light responsiveness, suggesting the significant role of HD-ZIP III genes in response to light stress. Development-related cis-elements were discovered, including those related to meristem expression, zein metabolism regulation, endosperm expression and differentiation of palisade mesophyll cells, providing clues that the Brassica HD-ZIP III genes may also participate in developmental processes. Hormone-correlated responsive elements related to methyl jasmonate (MeJA), ethylene, abscisic acid (ABA), salicylic acid (SA), gibberellin (GA) and auxin were identified. There were more MeJA-related elements (88) than other hormones and 45 ABA-related elements located in the promoters of 22 genes. Various kinds of stress-related cis-elements were discovered. The most predominant elements were related to anaerobic induction with a total of 74 sites. Others included the cis-elements related to low-temperature, defense and stress, drought, wounding and pathogen responses (W box and WUN motif), indicating that they may respond to stress stimulation. The MYB and MYC families of transcription factors were functionally characterized for their involvement in stress. There were 130 and 84 binding sites of MYB and MYC proteins, respectively. These results suggested that HD-ZIP III may function in both plant development and stress responses.
3.6. Expression Profiling of HD-ZIP III Genes in Different Tissues
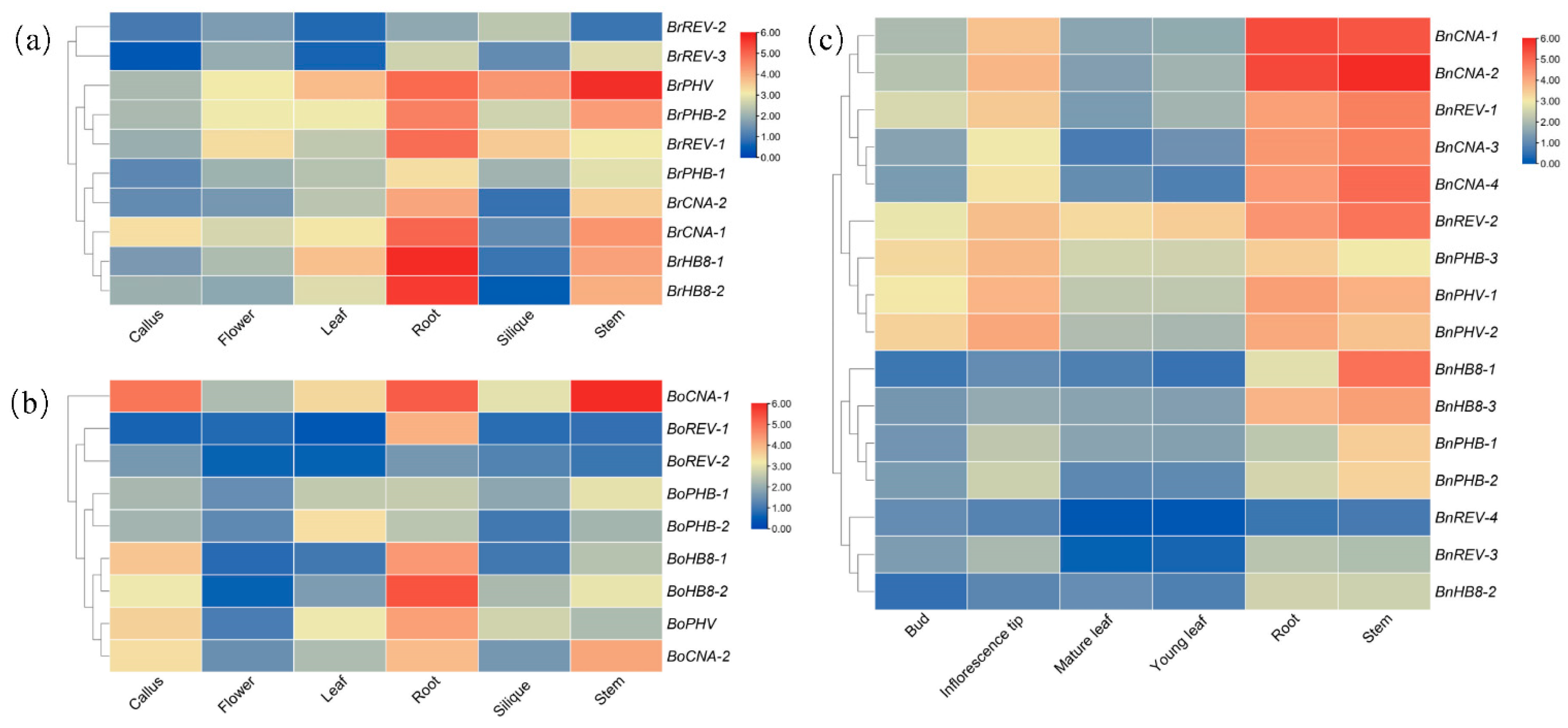
To characterize the expression profiling of HD-ZIP III genes in different tissues, the abundance of their transcripts in the callus, flower, root, stem, leaf and silique in B. rapa and B. oleracea were analyzed using publicly available RNA-Seq data (B. rapa: GSE43245; B. oleareaca: GSE42891) and the heatmaps were built based on the log2-transfomed FPKM values (Figure 5a,b; Tables S3 and S4). These genes exhibited broad expression patterns in various tissues. In most conditions, different copies exhibited the same expression tendency and were clustered in a clade, such as BrHB8-1 and BrHB8-2, as did members of BoREV, BoPHB and BoHB8. However, we also observed that homologs of BrREV and BoCNA possessed different expression levels. For example, BrREV-1 showed markedly higher expression than BrREV-2 and BrREV-3, as well as BoCNA-1 compared to BoCNA-2. Furthermore, HD-ZIP III genes showed various expression patterns in different tissues. It was notable that a majority of them had a high expression level in the root and stem, especially BrHB8-1 and BrHB8-2 in the root and BrPHV in the stem, as well as BoHB8-2 in the root and BoCNA-1 in the stem. The expression levels of BrHB8-1, BrHB8-2 and BrCNA-1 were much lower in the silique but the value was much high for BrPHV. BoHB8-1 and BoHB8-2 had a low expression in the flower. BrPHB-1 and BoPHB-1 exhibited moderate expression levels in each tissue compared with other genes.
The expression levels of HD-ZIP III genes in B. napus were retrieved from Brassica EDB with six tissues, namely, the bud, inflorescence tip, mature leaf, young leaf, root and stem at the bolting stage (Figure 5c, Table S5). Most genes were highly expressed in the root and stem, which was the same as that in B. rapa and B. oleracea. BnPHV-2 showed the highest expression level in the bud and inflorescence tip, while BnREV-4 expressed at a very low level in each tissue. These results indicated that HD-ZIP III genes may play roles in various developmental processes and provide an important basis for predicting their functions.
3.7. Expression Profiling of B. rapa HD-ZIP III Genes under Abiotic Stresses
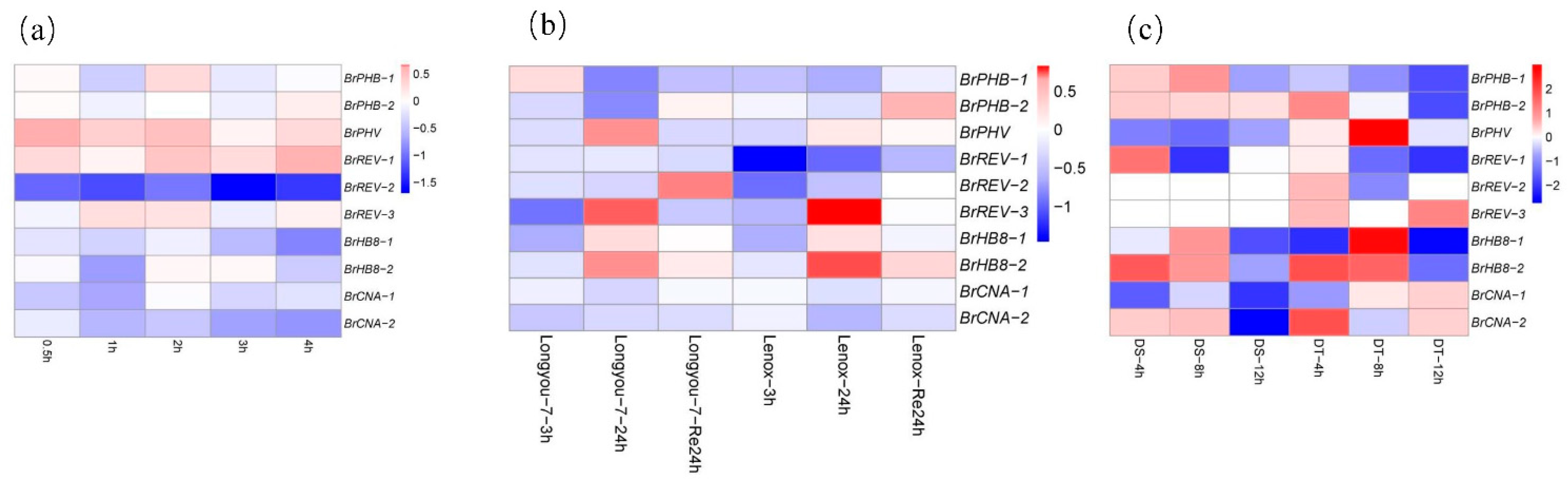
To gain information about the expression pattern of HD-ZIP III genes in response to abiotic stresses, we utilized the public data on transcriptional profiling from B. rapa under heat, cold and drought stress conditions. For the heat treatment, Chinese cabbage line “Chiifu” seedlings were subjected to 45 °C and the materials were collected at 0, 0.5, 1, 2, 3 and 4 h, respectively [52] (Figure 6a, Table S6). Only BrREV-2 was downregulated two times under heat stress, with the lowest expression at 3 h. The expression level of the other genes did not change significantly.
Upon cold stress treatment, the expression levels of 10 HD-ZIP III genes were compared in transcriptome data from winter B. rapa [53]. A cold-tolerant cultivar (Longyou-7) and a cold-sensitive cultivar (Lenox) were subjected to 4 °C for 3 h, 24 h and recovery at 22 °C for 24 h (Figure 6b, Table S7). Analysis showed that BrPHB-1 and BrPHB-2 were downregulated at 24 h in Longyou-7 and then restored. There was a decreased expression for BrREV-1 and BrREV-2 at 3 h in Lenox, and BrREV-1 was also decreased at 24 h in Lenox. The transcript accumulation of BrREV-3 was decreased at 3 h and then increased in Longyou-7 and Lenox. It was obvious that different copies from PHB and REV responded to cold stress at early or late stages.
Transcriptome data from Chinese cabbage root were used to analyze the expression pattern of BrHD-ZIP III genes under drought stress [54]. A drought-sensitive (DS) and a drought-tolerant (DT) genotype were treated with 2.5% PEG 6000 in seedlings and they were collected at 4 h, 8 h and 12 h (Figure 6c, Table S8). In the drought-sensitive line, BrPHV showed downregulation up to 4 and 8 h. BrHB8-1, BrCNA-1 and BrCNA-2 decreased at 12 h. Upregulation in the expression levels of BrREV-1 and BrHB8-2 were seen at 4 h with a further decline of BrREV-1 at 8 h. However, the genes showed different expression profiles in the drought-tolerant line. Most of the genes responded to drought stress at at least one time point. Some genes showed the same expression pattern at different time points. The transcript of BrREV-1 was decreased at 8 h and 12 h. Some genes showed the reverse expression pattern. For example, BrHB8-2 exhibited upregulated expression at 4 h and 8 h, with a decreased expression at 12 h. The diverse expression profiling of these HD-ZIP III genes in two plant materials’ exposure to drought stress implies they may play different roles in the response of drought tolerance in B. rapa. The expression analyses will provide useful information for further characterization HD-ZIP III functions in B. rapa.
3.8. Analysis of miR165/166 Target Sequences Variation in HD-ZIP III Genes
MicroRNAs (miRNAs) are small non-coding RNAs that bind to complementary mRNA and negatively regulate the target gene expression [57]. Previous studies revealed that HD-ZIP III genes were post-transcriptionally regulated by miR165/166 [8,11]. To check the genetic variation at the miRNA165/166 binding sites of HD-ZIP III genes in these three Brassica species, we divided all the identified HD-ZIP III genes into five groups according to the phylogenetic relationship and compared the miRNA165/166 complementary sites in each group. All the homologous genes of PHV shared identical targeting sequences (Figure 7b), where groups of HB8 homologous genes and CNA groups also harbored the same sequences (Figure 7d,e). Members from the PHB group exhibited a single nucleotide difference. There was a “G” at the fifth miRNA165/166 complementary site for each of BrPHB-2, BoPHB-2 and BnPHB-3, which was identical with AtPHB in Arabidopsis, while the rest of the member genes contained a “U” at the same position (Figure 7a). BrREV-2 contained a “C” at the second position, and the remaining genes contained a “U”. Five genes had a “G” at the fifth position, which was in accordance with the Arabidopsis sequences. However, the other four genes BrREV-3, BoREV-2, BnREV-3 and BnREV-4 were mutated to “U” (Figure 7c).
In order to confirm the existence of different nucleotides of HD-ZIP III genes at the miR165/166 complementary sites, the REV gene of B. rapa was selected and three cultivars, namely, one heading Chinese cabbage (Bre) and two non-heading Chinese cabbages (HQ and ZCT), were used for amplifying the fragments of BrREV-2 and BrREV-3. The sequencing results were compared with the identified miR165/166 in B. rapa (Figure S2) [55]. This showed that all the amplified nucleotides at the miRNA165/166 target sites were the same as those in the reference genome (Figure 8a–c), indicating the existence of polymorphisms in the REV members.
4. Discussion
HD-ZIP transcription factors are widely disseminated in plants and play important roles in plant growth and stress responses. The HD-ZIP family consists of four well-defined subfamilies I to IV [1,2,3]. The HD-ZIP III subfamily was the smallest one of the HD-ZIP family that has been connected with many developmental events, such as embryo patterning, apical meristem formation, organ polarity establishment and vascular development [6,7,13,14,15]. Recent studies showed the potential of this subfamily of genes in enhancing tolerance to stresses [25,26,27,28]. There is no comprehensive study of the HD-ZIP III genes in Brassica crops. In this study, a total of 35 members were identified and characterized from B. rapa, B. oleracea and B. napus. Their expression profiling in response to abiotic stress was also investigated.
Previous studies revealed five HD-ZIP III genes in Arabidopsis, rice, maize, grape and pepper [16,37,38,41,44]; six in M. truncatula and tomato [33,40]; eight in populus [39] and nine in cassava and sesame [30,42]. The number of HD-ZIP III genes identified in B. napus was 16 and it was larger than that in other plant species. BnaA02g06170D was a shortened copy of REV in which the homeobox domain was missing, and therefore, it was excluded from this study. The Brassicaceae species experienced a whole-genome triplication (WGT) event [58,59]. A single AtHD-ZIP III gene had multiple orthologues in the Brassica species. B. rapa and B. oleracea would be expected to contain three copies and B. napus contains six copies of each Arabidopsis gene. However, the number of the most HD-ZIP III genes in the B. rapa and B. oleracea genomes was less than three times higher compared with Arabidopsis, and only 2–4 copies were identified in B. napus. It was reported that 35% of the genes were lost via a deletion mechanism when genome triplication occurred in the Brassica lineage [60]. The results suggested that some HD-ZIP III genes might have been lost during evolution. In Arabidopsis, single loss-of-function mutants of HD-ZIP III genes did not show any obvious defect during embryogenesis, whereas the rev mutant showed identifiable defects post-embryonically [16]. We found that only BrREV retained three copies in B. rapa, while the other members retained one or two copies, implying that REV might have irreplaceable functions. PHB and PHV had a closer relationship and there was only one copy of PHV and two copies of PHB in B. rapa and B. oleracea. It is possible that functionally redundant copies were lost while the important copies remained, where these copies play a vital function.
The phylogenetic tree of HD-ZIP III proteins from B. rapa, B. oleracea, B. napus and Arabidopsis revealed that they could be classified into three clades. Homologous genes from different species were closer and gathered in a small clade. Most of one single chromosome contained one HD-ZIP III, while a few contained two or three genes. Gene structure analysis showed that the number of introns ranged from 13–18. Among the 15 conserved motifs, these HD-ZIP III proteins shared at least 11 common motifs. The similarities in gene structure and conserved motifs of the HD-ZIP III genes inferred their evolutionary relationships. The majority of the HD-ZIP III proteins contained four conserved domains, while four of them lacked the MEHKHA domain and two proteins had a short MEHKHA domain. The deduced protein sequences were shorter than the other ones, which may have been due to the incomplete genome sequencing or the loss of the N terminal during evolution.
Cis-elements located in the promoter regions play crucial roles during growth and environmental changes through regulating gene expression. Cis-elements analysis revealed that HD-ZIP III genes contained various types of cis-elements. Light is a major source of energy and an indispensable environmental signal regulating developmental processes. OsHox32, a member of HD-ZIP III genes in rice, is highly expressed during the light stage and lower expressed during the dark stage [61]. The light-responsive elements were most abundant in the HD-ZIP III promoters of the three Brassica species, which indicated they may play important roles in this process. Phytohormones-related elements, including those responses to MeJA, ethylene, ABA, SA and auxin, were present in the promoters of the HD-ZIP III genes. Furthermore, cis-acting elements associated with abiotic responses were located in the HD-ZIP III promoters, such as responsive to low-temperature and responsive to drought. Phytohormones play significant roles in maintaining the developmental processes and regulating stress responses in plants [62]. ABA is involved in the responses to a variety of abiotic stresses, such as drought, cold, heat stress and salinity [62]. It was reported that the expression levels of REV, PHB and PHV were decreased in response to ABA application in Arabidopsis [63]. REV positively regulates the transcription of PYL6 (ORK1/PYRABACTIN RESISTANCE1 LIKE6), a gene that encodes the ABA receptor protein [64,65]. Among the identified cis-elements, 22 out of 35 HD-ZIP III genes contained ABA-responsive elements, where the number of elements ranged from 1 to 9, indicating the involvement of HD-ZIP III genes in abiotic stresses and ABA response. SA and JA are known to play significant roles in regulating plant defense responses [66]. Interaction between auxin and ethylene regulates root development and architecture, which is a key aspect that is relevant to drought and salinity tolerance [67]. The analysis indicated that HD-ZIP III genes might respond to different stress involving phytohormones. MYB and MYC are involved in drought, cold and salt stress, as well as ABA [68,69], and they were distributed in the promoters of the HD-ZIP III genes. Defense and stress, wounding and pathogen response elements (W box and WUN motif) were also predicted, suggesting the involvement of HD-ZIP III genes in response to biotic and abiotic stress.
The expression profiling showed that HD-ZIP III genes had a broad expression in all tissues detected in the three Brassica species, implying that they participate in many development stages. It was reported that all genes in the HD-ZIP III subfamily were highly expressed in vascular bundle tissues in Eucalyptus grandis [43]. In our study, genes were highly expressed in the root and stem, especially for CNA and HB8, indicating their involvement in vascular development.
Numerous studies revealed the functions of HD-ZIP I and HD-ZIP II subfamily genes in response to a variety of abiotic stresses in different plants. Recent studies indicate that HD-ZIP III subfamily genes are also involved in the stress pathway [70]. For example, three tested members of MtHDZ III genes were significantly induced in response to cold and salt [33]. In soybean, Gmhdz59 was upregulated by both drought and salt [29]. Using the public data, we found that BrREV-2 responded to heat stress in particular. Members of BrPHB and BrREV reduced their expression levels under cold stress, and a subset of genes responded to drought stress. In Arabidopsis, reduced miR165/166 expression levels are more resistant to drought and cold stress [26,27]. PHB can directly bind to the BG1 promoter, which, in turn, modulates ABA homeostasis and improved drought resistance [26]. Leaf rolling reduces water loss, especially under drought stress conditions. In rice, overexpression of OsHox32 produces narrow and rolled leaves, and water use efficiency is significantly improved [61]. miR166 knockdown plants (STTM166), as well as overexpressing a miR166-resistant target OsHB4, display a rolled-leaf phenotype and improve drought resistance in rice [28]. Moreover, STTM166 plants alter stem vasculature and reduce water conductance. From our expression data analysis, it was clear that BrHD-ZIP III genes were responsive to different stresses. Considering the predicted cis-elements of promoters and the expression pattern of HD-ZIP III genes under stress conditions, it can be concluded that the regulation of HD-ZIP III activity might help to coordinate the development process with environmental stresses in Brassica crops.
miR165/166 was detected in several plant species [71]. By comparing the sequences, we found that each group of the PHV, HB8 and CNA genes shared the same miR165/166 target sites, indicating their conservation during evolution. There existed a single nucleotide difference among the PHB members and three kinds of REV sequences that differed by only one nucleotide from each other. PCR and sequencing results confirmed the polymorphisms of the REV genes in B. rapa. Mutations at the miR165/166 targeting site changed the mRNA expression level, which may cause morphological phenotypes, such as leaf shape with an abnormal adaxial–abaxial polarity [8,11,12,22]. B. rapa contains vegetable crops with different leaf morphologies [72,73]. Overexpression of Brp-MIR166g results in the downregulation of REV and PHB target genes and changes the direction and degree of leaf curvature in Chinese cabbage [73]. Leaf shape has a significant impact on yield, quality and stress responses. For example, disruption in the establishment of plant adaxial–abaxial polarity can give rise to rolling leaves and appropriate leaf rolling is thought to be an adaptive mechanism that can conserve water under drought stress [28,61]. The regulations of the differences in target sites, the expression level of the corresponding genes and the morphological variations and their responses to stress need to be further explored.
5. Conclusions
In this study, we performed a comprehensive study of HD-ZIP III genes in three Brassica species and identified 10, 9 and 16 HD-ZIP III genes from B. rapa, B. oleracea and B. napus, respectively, via genome-wide analysis. These genes were unevenly distributed on the chromosomes. Members that were in the same group had similar exon–intron structures. Cis-element analysis from the promoter of HD-ZIP III genes revealed that they might respond to diverse environmental stresses. HD-ZIP III genes were constitutively expressed in all tested tissues. A subset of genes responded to heat, cold and drought stress in a specific manner in B. rapa through analyzing the public data. The HD-ZIP III genes shared conserved miR165/166 complementary sites, whereas one and two polymorphisms existed in PHB and REV groups. This study provides clues for functional characterization and the response of the HD-ZIP III genes during abiotic stress.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/d13120684/s1, Figure S1: Logo of the conserved motifs of the HD-ZIP III proteins. Figure S2: Prediction of miR165/166 stem-loop structures that produced miR165/166 in B. rapa using an RNAfold web server. Predicted secondary stem-loop structures of miR165a (a) and miR166a-1 (b) using the miRNA precursor sequences from B. rapa. Red lines indicate the mature miRNA sequences. Table S1: Primer sequences used in this study. Table S2: The detailed information of cis-elements in the promoter regions of HD-ZIP III genes in B. rapa, B. oleracea and B. napus. Table S3: The FPKM values of HD-ZIP III genes in B. rapa. Table S4: The FPKM values of the HD-ZIP III genes in B. oleracea. Table S5: The FPKM values of the HD-ZIP III genes in B. napus. Table S6: Expression values of the HD-ZIP III genes under 45 °C heat treatment in B. rapa. Table S7: Expression levels of the HD-ZIP III genes under cold treatment in B. rapa. Table S8: The log2-transformed FPKM values of the HD-ZIP III genes under drought treatment in B. rapa.
Author Contributions
H.W. designed the experiments and wrote the paper; H.W., W.S. and M.Y. analyzed the data; Y.X. and S.L. performed the sampling and experiments; H.W. and R.W. reviewed the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Natural Science Foundation of Jiangsu Province (grant no. BK20170756), the National Natural Science Foundation of China (31700280), and the Natural Science Foundation of the Jiangsu Higher Education Institutions of China (17KJB180002).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All of the analyzed data are available in the manuscript and Supplementary Materials.
Acknowledgments
We thank Yuke He from Shanghai Institute of Plant Physiology and Ecology, Chinese Academy of Sciences, for providing the B. rapa seeds.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ariel, F.D.; Manavella, P.A.; Dezar, C.A.; Chan, R.L. The true story of the HD-Zip family. Trends Plant Sci. 2007, 12, 419–426. [Google Scholar] [CrossRef]
- Elhiti, M.; Stasolla, C. Structure and function of homodomain-leucine zipper (HD-Zip) proteins. Plant Signal. Behav. 2009, 4, 86–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, K.; Brocchieri, L.; Bürglin, T.R. A Comprehensive Classification and Evolutionary Analysis of Plant Homeobox Genes. Mol. Biol. Evol. 2009, 26, 2775–2794. [Google Scholar] [CrossRef] [Green Version]
- Schrick, K.; Nguyen, D.; Karlowski, W.M.; Mayer, K.F. START lipid/sterol-binding domains are amplified in plants and are predominantly associated with homeodomain transcription factors. Genome Biol. 2004, 5, R41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, K.; Bürglin, T.R. Mekhla, a Novel Domain with Similarity to PAS Domains, Is Fused to Plant Homeodomain-Leucine Zipper III Proteins. Plant Physiol. 2006, 140, 1142–1150. [Google Scholar] [CrossRef] [Green Version]
- Floyd, S.K.; Zalewski, C.S.; Bowman, J.L. Evolution of Class III Homeodomain–Leucine Zipper Genes in Streptophytes. Genetics 2006, 173, 373–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prigge, M.; Clark, S.E. Evolution of the class III HD-Zip gene family in land plants. Evol. Dev. 2006, 8, 350–361. [Google Scholar] [CrossRef] [PubMed]
- Emery, J.F.; Floyd, S.K.; Alvarez, J.; Eshed, Y.; Hawker, N.P.; Izhaki, A.; Baum, S.F.; Bowman, J. Radial Patterning of Arabidopsis Shoots by Class III HD-ZIP and KANADI Genes. Curr. Biol. 2003, 13, 1768–1774. [Google Scholar] [CrossRef] [Green Version]
- Carlsbecker, A.; Lee, J.Y.; Roberts, C.J.; Dettmer, J.; Lehesranta, S.; Zhou, J.; Lindgren, O.; Moreno-Risueno, M.A.; Vaten, A.; Thitamadee, S.; et al. Cell signalling by microRNA165/6 directs gene dose-dependent root cell fate. Nature 2010, 465, 316–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Q.; Wang, H. The role of HD-ZIP III transcription factors and miR165/166 in vascular development and secondary cell wall formation. Plant Signal. Behav. 2015, 10, e1078955. [Google Scholar] [CrossRef] [Green Version]
- Mallory, A.C.; Reinhart, B.J.; Jones-Rhoades, M.W.; Tang, G.L.; Zamore, P.D.; Barton, M.K.; Bartel, D.P. MicroRNA control of PHABULOSA in leaf development: Importance of pairing to the microRNA 5′ region. EMBO J. 2004, 23, 3356–3364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Jung, J.H.; Reyes, J.L.; Kim, Y.S.; Kim, S.Y.; Chung, K.S.; Kim, J.A.; Lee, M.; Lee, Y.; Narry Kim, V.; et al. mi-croRNA-directed cleavage of ATHB15 mRNA regulates vascular development in Arabidopsis inflorescence stems. Plant J. 2005, 42, 84–94. [Google Scholar] [CrossRef] [Green Version]
- Ramachandran, P.; Carlsbecker, A.; Etchells, J.P. Class III HD-ZIPs govern vascular cell fate: An HD view on patterning and differentiation. J. Exp. Bot. 2017, 68, 55–69. [Google Scholar] [CrossRef] [Green Version]
- Sessa, G.; Carabelli, M.; Possenti, M.; Morelli, G.; Ruberti, I. Multiple Links between HD-Zip Proteins and Hormone Networks. Int. J. Mol. Sci. 2018, 19, 4047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manuela, D.; Xu, M. Patterning a Leaf by Establishing Polarities. Front. Plant Sci. 2020, 11, 568730. [Google Scholar] [CrossRef] [PubMed]
- Prigge, M.J.; Otsuga, D.; Alonso, J.M.; Ecker, J.R.; Drews, G.N.; Clark, S.E. Class III Homeodomain-Leucine Zipper Gene Family Members Have Overlapping, Antagonistic, and Distinct Roles in Arabidopsis Development. Plant Cell 2005, 17, 61–76. [Google Scholar] [CrossRef] [Green Version]
- McConnell, J.R.; Emery, J.; Eshed, Y.; Bao, N.; Bowman, J.; Barton, M.K. Role of PHABULOSA and PHAVOLUTA in deter-mining radial patterning in shoots. Nature 2001, 411, 709–713. [Google Scholar] [CrossRef] [PubMed]
- Baima, S.; Possenti, M.; Matteucci, A.; Wisman, E.; Altamura, M.M.; Ruberti, I.; Morelli, G. The Arabidopsis ATHB-8 HD-Zip Protein Acts as a Differentiation-Promoting Transcription Factor of the Vascular Meristems. Plant Physiol. 2001, 126, 643–655. [Google Scholar] [CrossRef] [Green Version]
- Green, K.A.; Prigge, M.; Katzman, R.B.; Clark, S.E. CORONA, a Member of the Class III Homeodomain Leucine Zipper Gene Family in Arabidopsis, Regulates Stem Cell Specification and Organogenesis. Plant Cell 2005, 17, 691–704. [Google Scholar] [CrossRef] [Green Version]
- Juarez, M.; Twigg, R.W.; Timmermans, M.C.P. Specification of adaxial cell fate during maize leaf development. Development 2004, 131, 4533–4544. [Google Scholar] [CrossRef] [Green Version]
- Itoh, J.-I.; Hibara, K.-I.; Sato, Y.; Nagato, Y. Developmental Role and Auxin Responsiveness of Class III Homeodomain Leucine Zipper Gene Family Members in Rice. Plant Physiol. 2008, 147, 1960–1975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rong, F.; Chen, F.; Huang, L.; Zhang, J.; Zhang, C.; Hou, D.; Cheng, Z.; Weng, Y.; Chen, P.; Li, Y. A mutation in class III homeodomain-leucine zipper (HD-ZIP III) transcription factor results in curly leaf (cul) in cucumber (Cucumis sativus L.). Theor. Appl. Genet. 2018, 132, 113–123. [Google Scholar] [CrossRef]
- Du, J.; Miura, E.; Robischon, M.; Martinez, C.; Groover, A. The Populus Class III HD ZIP Transcription Factor POPCORONA Affects Cell Differentiation during Secondary Growth of Woody Stems. PLoS ONE 2011, 6, e17458. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Song, N.; Xu, P.; Sun, J.; Li, L. A HD-ZIP III gene, PtrHB4, is required for interfascicular cambium development in Populus. Plant Biotechnol. J. 2017, 16, 808–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandt, R.; Salla-Martret, M.; Bou-Torrent, J.; Musielak, T.; Stahl, M.; Lanz, C.; Ott, F.; Schmid, M.; Greb, T.; Schwarz, M.; et al. Genome-wide binding-site analysis of REVOLUTA reveals a link between leaf patterning and light-mediated growth responses. Plant J. 2012, 72, 31–42. [Google Scholar] [CrossRef]
- Yan, J.; Zhao, C.; Zhou, J.; Yang, Y.; Wang, P.; Zhu, X.; Tang, G.; Bressan, R.A.; Zhu, J.K. The miR165/166 Mediated Regulatory Module Plays Critical Roles in ABA Homeostasis and Response in Arabidopsis thaliana. PLoS Genet. 2016, 12, e1006416. [Google Scholar] [CrossRef]
- Yang, T.; Wang, Y.; Teotia, S.; Wang, Z.; Shi, C.; Sun, H.; Gu, Y.; Zhang, Z.; Tang, G. The interaction between miR160 and miR165/166 in the control of leaf development and drought tolerance in Arabidopsis. Sci. Rep. 2019, 9, 2832. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, H.; Srivastava, A.K.; Pan, Y.; Bai, J.; Fang, J.; Shi, H.; Zhu, J.K. Knockdown of Rice MicroRNA166 Confers Drought Resistance by Causing Leaf Rolling and Altering Stem Xylem Development. Plant Physiol. 2018, 176, 2082–2094. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Chen, Z.; Zhao, H.; Zhao, Y.; Cheng, B.; Xiang, Y. Genome-wide analysis of soybean HD-Zip gene family and ex-pression profiling under salinity and drought treatments. PLoS ONE 2014, 9, e87156. [Google Scholar]
- Ding, Z.; Fu, L.; Yan, Y.; Tie, W.; Xia, Z.; Wang, W.; Peng, M.; Hu, W.; Zhang, J. Genome-wide characterization and expression profiling of HD-Zip gene family related to abiotic stress in cassava. PLoS ONE 2017, 12, e0173043. [Google Scholar] [CrossRef] [Green Version]
- Yue, H.; Shu, D.; Wang, M.; Xing, G.; Zhan, H.; Du, X.; Song, W.; Nie, X. Genome-Wide Identification and Expression Analysis of the HD-Zip Gene Family in Wheat (Triticum aestivum L.). Genes 2018, 9, 70. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Dong, J.; Cao, M.; Gao, X.; Wang, D.; Liu, B.; Chen, Q. Genome-wide identification and characterization of HD-ZIP genes in potato. Gene 2019, 697, 103–117. [Google Scholar] [CrossRef]
- Li, Z.; Gao, Z.; Li, R.; Xu, Y.; Kong, Y.; Zhou, G.; Meng, C.; Hu, R. Genome-wide identification and expression profiling of HD-ZIP gene family in Medicago truncatula. Genomics 2020, 112, 3624–3635. [Google Scholar] [CrossRef]
- Sharif, R.; Xie, C.; Wang, J.; Cao, Z.; Zhang, H.; Chen, P.; Yuhong, L. Genome wide identification, characterization and ex-pression analysis of HD-ZIP gene family in Cucumis sativus L. under biotic and various abiotic stresses. Int. J. Biol. Macromolecules 2020, 158, 502–520. [Google Scholar] [CrossRef]
- Talalay, P.; Fahey, J.W. Phytochemicals from Cruciferous Plants Protect against Cancer by Modulating Carcinogen Metabolism. J. Nutr. 2001, 131, 3027S–3033S. [Google Scholar] [CrossRef] [PubMed]
- Chalhoub, B.; Denoeud, F.; Liu, S.; Parkin, I.A.P.; Tang, H.; Wang, X.; Chiquet, J.; Belcram, H.; Tong, C.; Samans, B.; et al. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science 2014, 345, 950–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agalou, A.; Purwantomo, S.; Övernäs, E.; Johannesson, H.; Zhu, X.; Estiati, A.; De Kam, R.J.; Engström, P.; Slamet-Loedin, I.; Zhu, Z.; et al. A genome-wide survey of HD-Zip genes in rice and analysis of drought-responsive family members. Plant Mol. Biol. 2007, 66, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhou, Y.; Jiang, H.; Li, X.; Gan, D.; Peng, X.; Zhu, S.; Cheng, B. Systematic Analysis of Sequences and Expression Patterns of Drought-Responsive Members of the HD-Zip Gene Family in Maize. PLoS ONE 2011, 6, e28488. [Google Scholar] [CrossRef] [Green Version]
- Hu, R.; Chi, X.; Chai, G.; Kong, Y.; He, G.; Wang, X.; Shi, D.; Zhang, D.; Zhou, G. Genome-Wide Identification, Evolutionary Expansion, and Expression Profile of Homeodomain-Leucine Zipper Gene Family in Poplar (Populus trichocarpa). PLoS ONE 2012, 7, e31149. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Chen, X.; Guan, X.; Liu, Y.; Chen, H.; Wang, T.; Mouekouba, L.D.O.; Li, J.; Wang, A. A genome-wide survey of homeodomain-leucine zipper genes and analysis of cold-responsive HD-Zip I members’ expression in tomato. Biosci. Biotechnol. Biochem. 2014, 78, 1337–1349. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, C.; Guo, Y.; Niu, W.; Wang, Y.; Xu, Y. Evolution and expression analysis reveal the potential role of the HD-Zip gene family in regulation of embryo abortion in grapes (Vitis vinifera L.). BMC Genom. 2017, 18, 744. [Google Scholar] [CrossRef] [Green Version]
- Wei, M.; Liu, A.; Zhang, Y.; Zhou, Y.; Li, D.; Dossa, K.; Zhou, R.; Zhang, X.; You, J. Genome-wide characterization and ex-pression analysis of the HD-Zip gene family in response to drought and salinity stresses in sesame. BMC Genom. 2019, 20, 748. [Google Scholar] [CrossRef]
- Zhang, J.; Wu, J.; Guo, M.; Aslam, M.; Wang, Q.; Ma, H.; Li, S.; Zhang, X.; Cao, S. Genome-wide characterization and ex-pression profiling of Eucalyptus grandis HD-Zip gene family in response to salt and temperature stress. BMC Plant Biol. 2020, 20, 451. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhu, R.; Ji, X.; Li, H.J.; Lv, H.; Zhang, H.Y. Genome-Wide Characterization and Expression Analysis of the HD-ZIP Gene Family in Response to Salt Stress in Pepper. Int. J. Genom. 2021, 2021, 1–14. [Google Scholar] [CrossRef]
- Wang, X.; Wu, J.; Liang, J.; Cheng, F.; Wang, X. Brassica database (BRAD) version 2.0: Integrating and mining Brassicaceae species genomic resources. Database 2015, 2015, bav093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1897. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Tong, C.; Wang, X.; Yu, J.; Wu, J.; Li, W.; Huang, J.; Dong, C.; Hua, W.; Liu, S. Comprehensive analysis of RNA-seq data reveals the complexity of the transcriptome in Brassica rapa. BMC Genom. 2013, 14, 689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Liu, Y.; Yang, X.; Tong, C.; Edwards, D.; Parkin, I.A.P.; Zhao, M.; Ma, J.; Yu, J.; Huang, S.; et al. The Brassica oleracea genome reveals the asymmetrical evolution of polyploid genomes. Nat. Commun. 2014, 5, 3930. [Google Scholar] [CrossRef]
- Chao, H.; Li, T.; Luo, C.; Huang, H.; Ruan, Y.; Li, X.; Niu, Y.; Fan, Y.; Sun, W.; Zhang, K.; et al. BrassicaEDB: A Gene Expression Database for Brassica Crops. Int. J. Mol. Sci. 2020, 21, 5831. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Yi, H.; Lee, J.; Nou, I.-S.; Han, C.-T.; Hur, Y. Global Gene-Expression Analysis to Identify Differentially Expressed Genes Critical for the Heat Stress Response in Brassica rapa. PLoS ONE 2015, 10, e0130451. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Coulter, J.A.; Liu, L.; Zhao, Y.; Chang, Y.; Pu, Y.; Zeng, X.; Xu, Y.; Wu, J.; Fang, Y.; et al. Transcriptome Analysis Reveals Key Cold-Stress-Responsive Genes in Winter Rapeseed (Brassica rapa L.). Int. J. Mol. Sci. 2019, 20, 1071. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.M.; Samans, B.; Chen, S.; Kibret, K.B.; Hatzig, S.; Turner, N.C.; Nelson, M.N.; Cowling, W.A.; Snowdon, R.J. Drought-Tolerant Brassica rapa Shows Rapid Expression of Gene Networks for General Stress Responses and Programmed Cell Death Under Simulated Drought Stress. Plant Mol. Biol. Rep. 2017, 35, 416–430. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Wang, H.; Lu, Y.; De Ruiter, M.; Cariaso, M.; Prins, M.; Van Tunen, A.; He, Y. Identification of conserved and novel microRNAs that are responsive to heat stress in Brassica rapa. J. Exp. Bot. 2011, 63, 1025–1038. [Google Scholar] [CrossRef]
- Ibraheem, O.; Botha, C.E.; Bradley, G. In silico analysis of cis-acting regulatory elements in 5′ regulatory regions of sucrose transporter gene families in rice (Oryza sativa Japonica) and Arabidopsis thaliana. Comput. Biol. Chem. 2010, 34, 268–283. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Lysak, M.A.; Koch, M.A.; Pecinka, A.; Schubert, I. Chromosome triplication found across the tribe Brassiceae. Genome Res. 2005, 15, 516–525. [Google Scholar] [CrossRef] [Green Version]
- Cheng, F.; Wu, J.; Wang, X. Genome triplication drove the diversification of Brassica plants. Hortic. Res. 2014, 1, 14024. [Google Scholar] [CrossRef] [Green Version]
- Town, C.D.; Cheung, F.; Maiti, R.; Crabtree, J.; Haas, B.J.; Wortman, J.R.; Hine, E.E.; Althoff, R.; Arbogast, T.S.; Tallon, L.J.; et al. Comparative genomics of Brassica oleracea and Arabidopsis thaliana reveal gene loss, fragmentation, and dispersal after polyploidy. Plant Cell 2006, 18, 1348–1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.Y.; Shen, A.; Xiong, W.; Sun, Q.L.; Luo, Q.; Song, T.; Li, Z.L.; Luan, W.J. Overexpression of OsHox32 Results in Plei-otropic Effects on Plant Type Architecture and Leaf Development in Rice. Rice 2016, 9, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandt, R.; Cabedo, M.; Xie, Y.; Wenkel, S. Homeodomain leucine-zipper proteins and their role in synchronizing growth and development with the environment. J. Integr. Plant Biol. 2014, 56, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Reinhart, B.J.; Magnani, E.; Huang, T.; Kerstetter, R.; Barton, M.K. Of blades and branches: Understanding and ex-panding the Arabidopsis ad/abaxial regulatory network through target gene identification. Cold Spring Harb. Symp. Quant. Biol. 2012, 77, 31–45. [Google Scholar] [CrossRef] [Green Version]
- Reinhart, B.J.; Liu, T.; Newell, N.R.; Magnani, E.; Huang, T.; Kerstetter, R.; Michaels, S.; Barton, M.K. Establishing a Framework for the Ad/Abaxial Regulatory Network of Arabidopsis: Ascertaining Targets of Class III Homeodomain Leucine Zipper and Kanadi Regulation. Plant Cell 2013, 25, 3228–3249. [Google Scholar] [CrossRef] [Green Version]
- Bari, R.; Jones, J.D.G. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef]
- Kohli, A.; Sreenivasulu, N.; Lakshmanan, P.; Kumar, P.P. The phytohormone crosstalk paradigm takes center stage in un-derstanding how plants respond to abiotic stresses. Plant Cell Rep. 2013, 32, 945–957. [Google Scholar] [CrossRef]
- Abe, H.; Yamaguchi-Shinozaki, K.; Urao, T.; Iwasaki, T.; Hosokawa, D.; Shinozaki, K. Role of Arabidopsis MYC and MYB homologs in drought- and abscisic acid-regulated gene expression. Plant Cell 1997, 9, 1859–1868. [Google Scholar]
- Abe, H.; Urao, T.; Ito, T.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) Function as Transcriptional Activators in Abscisic Acid Signaling. Plant Cell 2003, 15, 63–78. [Google Scholar] [CrossRef] [Green Version]
- Sharif, R.; Raza, A.; Chen, P.; Li, Y.; El-Ballat, E.M.; Rauf, A.; Hano, C.; El-Esawi, M.A. HD-ZIP Gene Family: Potential Roles in Improving Plant Growth and Regulating Stress-Responsive Mechanisms in Plants. Genes 2021, 12, 1256. [Google Scholar] [CrossRef]
- Sunkar, R.; Jagadeeswaran, G. In silico identification of conserved microRNAs in large number of diverse plant species. BMC Plant Biol. 2008, 8, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, D.; Wang, H.; Basnet, R.K.; Zhao, J.; Lin, K.; Hou, X.; Bonnema, G. Genetic Dissection of Leaf Development in Brassica rapa Using a Genetical Genomics Approach. Plant Physiol. 2014, 164, 1309–1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, W.; Wang, H.; Bai, J.; Wu, F.; He, Y. Association of microRNAs with Types of Leaf Curvature in Brassica rapa. Front. Plant Sci. 2018, 9, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
The chromosomal locations of HD-ZIP III genes in B. rapa (a), B. oleracea (b) and B. napus (c).
Figure 1.
The chromosomal locations of HD-ZIP III genes in B. rapa (a), B. oleracea (b) and B. napus (c).

Figure 2.
Phylogenetic relationships of HD-ZIP III genes from B. rapa, B. oleracea, B. napus and A. thaliana. The phylogenetic tree was constructed with MEGA7.0 software using the NJ method with 1000 bootstrap replicates. The blue triangles, red circles, orange diamonds and green squares represent the HD-ZIP III proteins in B. rapa, B. oleracea, B. napus and A. thaliana, respectively.
Figure 2.
Phylogenetic relationships of HD-ZIP III genes from B. rapa, B. oleracea, B. napus and A. thaliana. The phylogenetic tree was constructed with MEGA7.0 software using the NJ method with 1000 bootstrap replicates. The blue triangles, red circles, orange diamonds and green squares represent the HD-ZIP III proteins in B. rapa, B. oleracea, B. napus and A. thaliana, respectively.

Figure 3.
Gene structures, conserved motifs and conserved domains of the HD-ZIP III proteins in B. rapa, B. oleracea and B. napus. (a) Intron/exon construction of the HD-ZIP III genes. Green boxes, yellow boxes and black lines indicate exons, untranslated regions and introns, respectively. (b) Distribution of the 15 conserved motifs of HD-ZIP III proteins. (c) Distribution of the four conserved domains of the HD-ZIP III proteins.
Figure 3.
Gene structures, conserved motifs and conserved domains of the HD-ZIP III proteins in B. rapa, B. oleracea and B. napus. (a) Intron/exon construction of the HD-ZIP III genes. Green boxes, yellow boxes and black lines indicate exons, untranslated regions and introns, respectively. (b) Distribution of the 15 conserved motifs of HD-ZIP III proteins. (c) Distribution of the four conserved domains of the HD-ZIP III proteins.

Figure 4.
Cis-element analysis of the HD-ZIP III promoters in B. rapa, B. oleracea and B. napus.

Figure 5.
Expression profiling of the HD-ZIP III genes in different tissues. Expression profiling of the HD-ZIP III genes in B. rapa (a), B. oleracea (b) and B. napus (c). The heatmaps were built using log2-transformed FPKM values.
Figure 5.
Expression profiling of the HD-ZIP III genes in different tissues. Expression profiling of the HD-ZIP III genes in B. rapa (a), B. oleracea (b) and B. napus (c). The heatmaps were built using log2-transformed FPKM values.

Figure 6.
Expression profiling of HD-ZIP III genes under abiotic stresses in B. rapa. (a) Differential expression profiling of HD-ZIP III genes under 45 °C heat stress for 0.5, 1, 2, 3 and 4 h in Chinese cabbage. (b) Differential expression profiling of HD-ZIP III genes under low-temperature stress at 4 °C for 3 h, 24 h and recovery at 22 °C for 24 h in a cold-tolerant cultivar (Longyou-7) and a cold-sensitive cultivar (Lenox). (c) Differential expression profiling of HD-ZIP III genes after a drought treatment for 4, 8 and 12 h in drought-sensitive (DS) and drought-tolerant (DT) genotypes. The heatmaps were built using log2-based fold change values of the treatment/control.
Figure 6.
Expression profiling of HD-ZIP III genes under abiotic stresses in B. rapa. (a) Differential expression profiling of HD-ZIP III genes under 45 °C heat stress for 0.5, 1, 2, 3 and 4 h in Chinese cabbage. (b) Differential expression profiling of HD-ZIP III genes under low-temperature stress at 4 °C for 3 h, 24 h and recovery at 22 °C for 24 h in a cold-tolerant cultivar (Longyou-7) and a cold-sensitive cultivar (Lenox). (c) Differential expression profiling of HD-ZIP III genes after a drought treatment for 4, 8 and 12 h in drought-sensitive (DS) and drought-tolerant (DT) genotypes. The heatmaps were built using log2-based fold change values of the treatment/control.

Figure 7.
Overview of the miR165/166 target sequences variation in the HD-ZIP III genes. Alignment of the miR165/166 putative target areas of PHB (a), PHV (b), REV (c), HB8 (d) and CNA genes (e). Red indicates the genetic variation of the miR165/166 target sites.
Figure 7.
Overview of the miR165/166 target sequences variation in the HD-ZIP III genes. Alignment of the miR165/166 putative target areas of PHB (a), PHV (b), REV (c), HB8 (d) and CNA genes (e). Red indicates the genetic variation of the miR165/166 target sites.

Figure 8.
Validation of genetic variation in the BrREV genes targeted by miR165/166. (a) Alignment of the PCR sequencing results from B. rapa (Bre, HQ and ZCT) plants and the BrREV reference genes. (b) Sequencing signal of BrREV-2 at the miR165/166 target area in the B. rapa (HQ) plant. (c) Sequencing signal of BrREV-3 at the miR165/166 target area in the B. rapa (HQ) plant. Red arrows indicate the polymorphisms that differed from other BrREV and AtREV genes.
Figure 8.
Validation of genetic variation in the BrREV genes targeted by miR165/166. (a) Alignment of the PCR sequencing results from B. rapa (Bre, HQ and ZCT) plants and the BrREV reference genes. (b) Sequencing signal of BrREV-2 at the miR165/166 target area in the B. rapa (HQ) plant. (c) Sequencing signal of BrREV-3 at the miR165/166 target area in the B. rapa (HQ) plant. Red arrows indicate the polymorphisms that differed from other BrREV and AtREV genes.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Identification and characterization of HD-ZIP III genes in B. rapa, B. oleracea and B. napus.
Table 1.
Identification and characterization of HD-ZIP III genes in B. rapa, B. oleracea and B. napus.
| Gene Name | Gene ID | Chr | Start | Stop | Amino Acid Length | Molecular Weight (kDa) | pI | Instability Index | GRAVY |
|---|---|---|---|---|---|---|---|---|---|
| BrPHB-1 | BraA05g010360.3C | A05 | 5534983 | 5539344 | 849 | 92.97 | 6.03 | 44.78 | −0.210 |
| BrPHB-2 | BraA04g024760.3C | A04 | 17918117 | 17922205 | 865 | 94.48 | 5.90 | 43.86 | −0.148 |
| BrPHV | BraA09g034560.3C | A09 | 27178071 | 27182489 | 840 | 92.16 | 6.06 | 42.97 | −0.199 |
| BrREV-1 | BraA10g018460.3C | A10 | 13671299 | 13675382 | 850 | 93.04 | 5.65 | 51.36 | −0.127 |
| BrREV-2 | BraA02g010200.3C | A02 | 4904176 | 4908251 | 827 | 91.23 | 5.58 | 51.43 | −0.124 |
| BrREV-3 | BraA06g021080.3C | A06 | 12282999 | 12287254 | 839 | 92.31 | 5.67 | 49.23 | −0.132 |
| BrHB8-1 | BraA01g005160.3C | A01 | 2462816 | 2467248 | 833 | 91.94 | 5.84 | 49.61 | −0.182 |
| BrHB8-2 | BraA08g016820.3C | A08 | 13619843 | 13624628 | 832 | 91.95 | 5.87 | 49.71 | −0.177 |
| BrCNA-1 | BraA06g002210.3C | A06 | 1326896 | 1330911 | 862 | 94.65 | 6.11 | 47.79 | −0.068 |
| BrCNA-2 | BraA08g002260.3C | A08 | 1609153 | 1612837 | 834 | 91.61 | 5.94 | 50.73 | −0.115 |
| BoPHB-1 | Bol027381 | C04 | 21283879 | 21288277 | 849 | 92.96 | 6.00 | 45.41 | −0.230 |
| BoPHB-2 | Bol037848 | C04 | 35825190 | 35829211 | 815 | 89.12 | 5.77 | 44.40 | −0.150 |
| BoPHV | Bol022567 | C05 | 10827887 | 10832270 | 840 | 92.17 | 6.06 | 42.10 | −0.210 |
| BoREV-1 | Bol035990 | C02 | 6843719 | 6847836 | 846 | 92.93 | 5.73 | 51.34 | −0.140 |
| BoREV-2 | Bol041327 | C03 | 34587260 | 34590833 | 704 | 77.17 | 5.79 | 47.09 | −0.132 |
| BoHB8-1 | Bol017514 | C03 | 53356119 | 53360922 | 832 | 91.87 | 5.84 | 48.94 | −0.173 |
| BoHB8-2 | Bol017856 | C01 | 3015634 | 3020035 | 833 | 91.94 | 5.84 | 49.12 | −0.184 |
| BoCNA-1 | Bol019484 | C06 | 12279219 | 12283287 | 838 | 91.90 | 6.04 | 48.98 | −0.102 |
| BoCNA-2 | Bol035052 | C03 | 55788767 | 55792479 | 835 | 91.58 | 6.04 | 48.81 | −0.108 |
| BnPHB-1 | BnaA05g09120D | chrA05 | 5027162 | 5031704 | 849 | 92.96 | 6.03 | 44.86 | −0.208 |
| BnPHB-2 | BnaC04g10480D | chrC04 | 8071926 | 8076487 | 849 | 92.94 | 6.00 | 45.13 | −0.227 |
| BnPHB-3 | BnaA04g20300D | chrA04 | 15670968 | 15675279 | 813 | 89.18 | 6.12 | 44.70 | −0.199 |
| BnPHV-1 | BnaA09g26050D | chrA09 | 19171300 | 19175944 | 840 | 92.13 | 6.06 | 42.97 | −0.195 |
| BnPHV-2 | BnaC05g23470D | chrC05 | 17572083 | 17576637 | 840 | 92.17 | 6.06 | 42.10 | −0.210 |
| BnREV-1 | BnaA10g13520D | chrA10 | 10868507 | 10873099 | 850 | 93.05 | 5.65 | 51.59 | −0.130 |
| BnREV-2 | BnaC09g54340D | chrC09_random | 4170497 | 4174807 | 847 | 92.75 | 5.65 | 51.51 | −0.124 |
| BnREV-3 | BnaA06g18550D | chrA06 | 10716894 | 10721117 | 839 | 92.23 | 5.62 | 48.90 | −0.102 |
| BnREV-4 | BnaC03g55030D | chrC03 | 43566142 | 43570110 | 766 | 84.21 | 6.02 | 46.67 | −0.117 |
| BnHB8-1 | BnaC03g66860D | chrC03 | 56525421 | 56529523 | 738 | 81.42 | 6.15 | 50.82 | −0.212 |
| BnHB8-2 | BnaA08g11980D | chrA08 | 10718251 | 10723616 | 738 | 81.35 | 6.15 | 51.38 | −0.209 |
| BnHB8-3 | BnaAnng30670D | chrAnn_random | 35003553 | 35006954 | 630 | 69.27 | 5.72 | 52.43 | −0.276 |
| BnCNA-1 | BnaA06g01940D | chrA06 | 1129905 | 1134711 | 838 | 91.87 | 6.04 | 48.79 | −0.117 |
| BnCNA-2 | BnaC06g05240D | chrC06 | 5886331 | 5891156 | 838 | 91.90 | 6.04 | 48.98 | −0.102 |
| BnCNA-3 | BnaA08g01600D | chrA08 | 1331344 | 1335319 | 835 | 91.65 | 6.04 | 50.10 | −0.117 |
| BnCNA-4 | BnaC03g69040D | chrC03 | 58725926 | 58729579 | 721 | 79.19 | 6.46 | 51.47 | −0.128 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, H.; Shao, W.; Yan, M.; Xu, Y.; Liu, S.; Wang, R. Genome-Wide Analysis and Expression Profiling of HD-ZIP III Genes in Three Brassica Species. Diversity 2021, 13, 684. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120684
AMA Style
Wang H, Shao W, Yan M, Xu Y, Liu S, Wang R. Genome-Wide Analysis and Expression Profiling of HD-ZIP III Genes in Three Brassica Species. Diversity. 2021; 13(12):684. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120684
Chicago/Turabian StyleWang, Han, Wenna Shao, Min Yan, Ye Xu, Shaohua Liu, and Renlei Wang. 2021. "Genome-Wide Analysis and Expression Profiling of HD-ZIP III Genes in Three Brassica Species" Diversity 13, no. 12: 684. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120684
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.