
The Current State of Eunicida (Annelida) Systematics and Biodiversity

 , and
, and 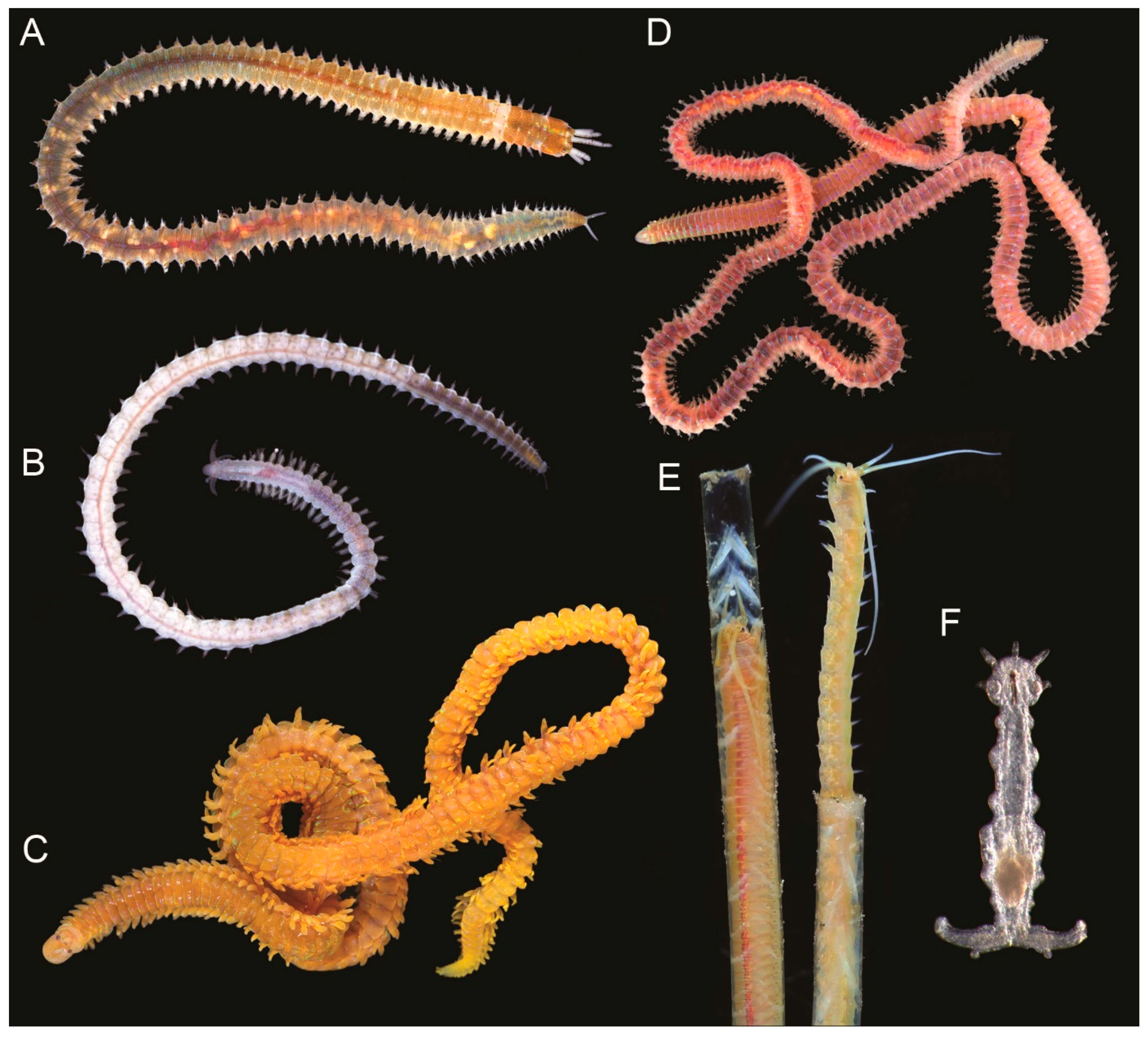{kind=link}
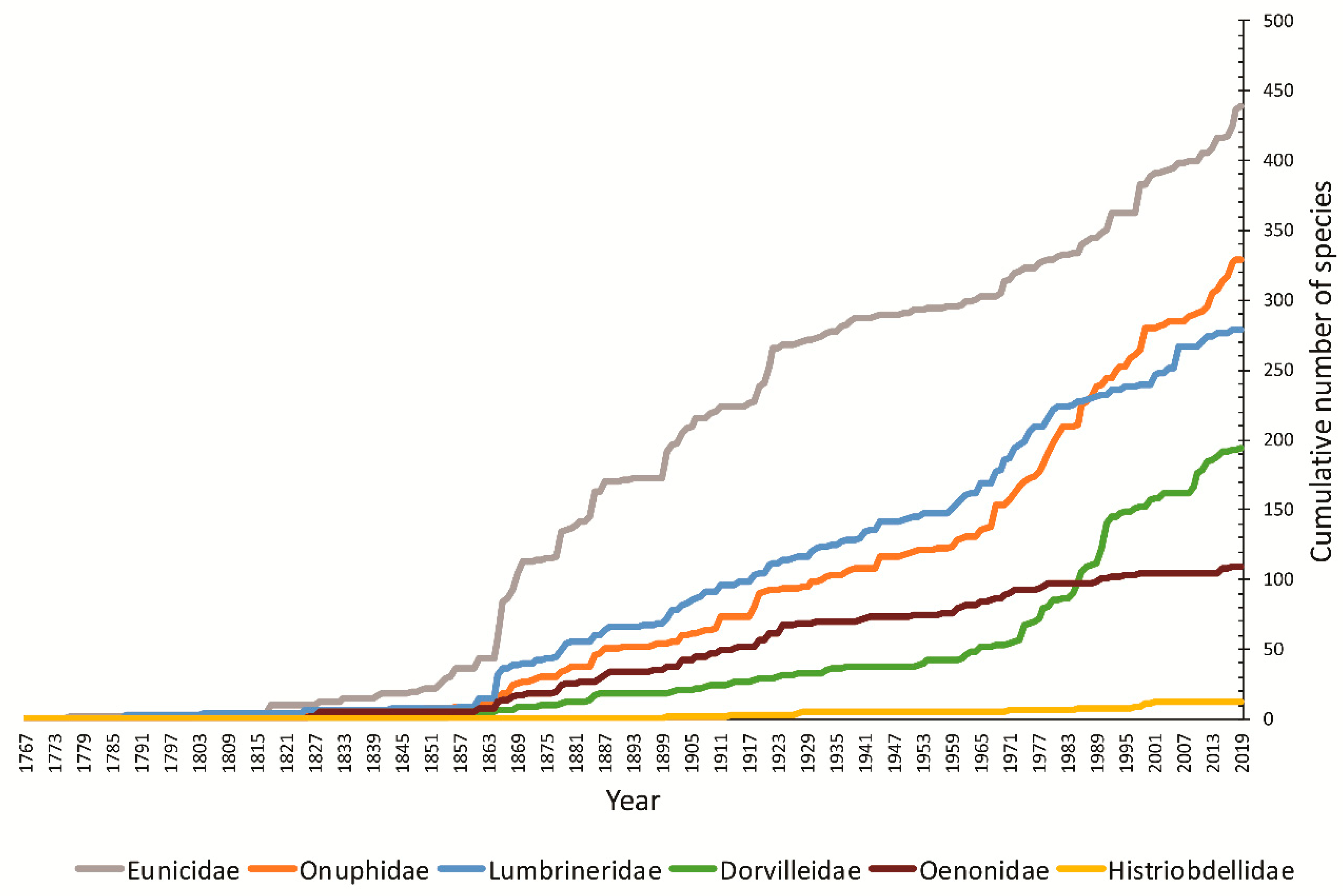{kind=link}
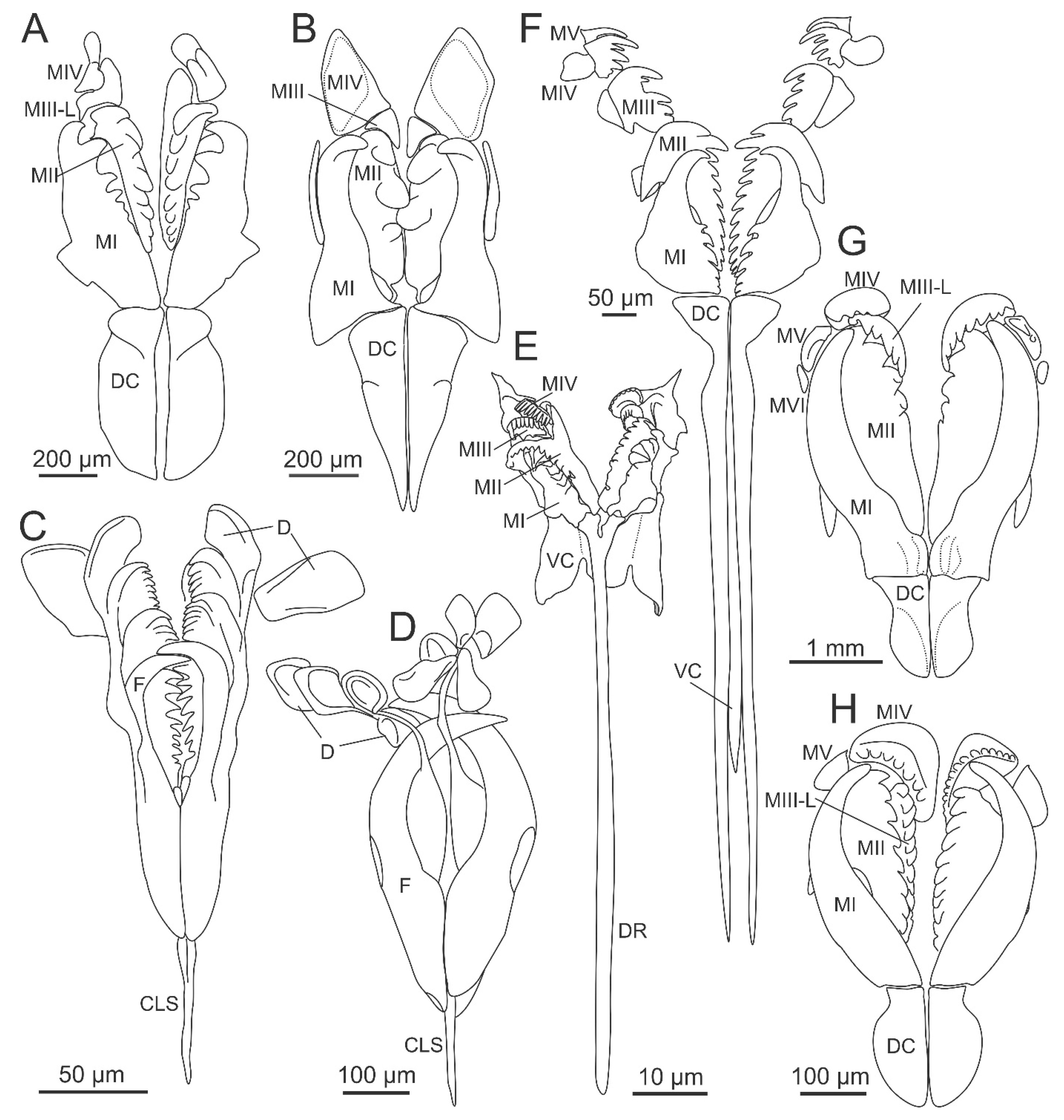{kind=link}
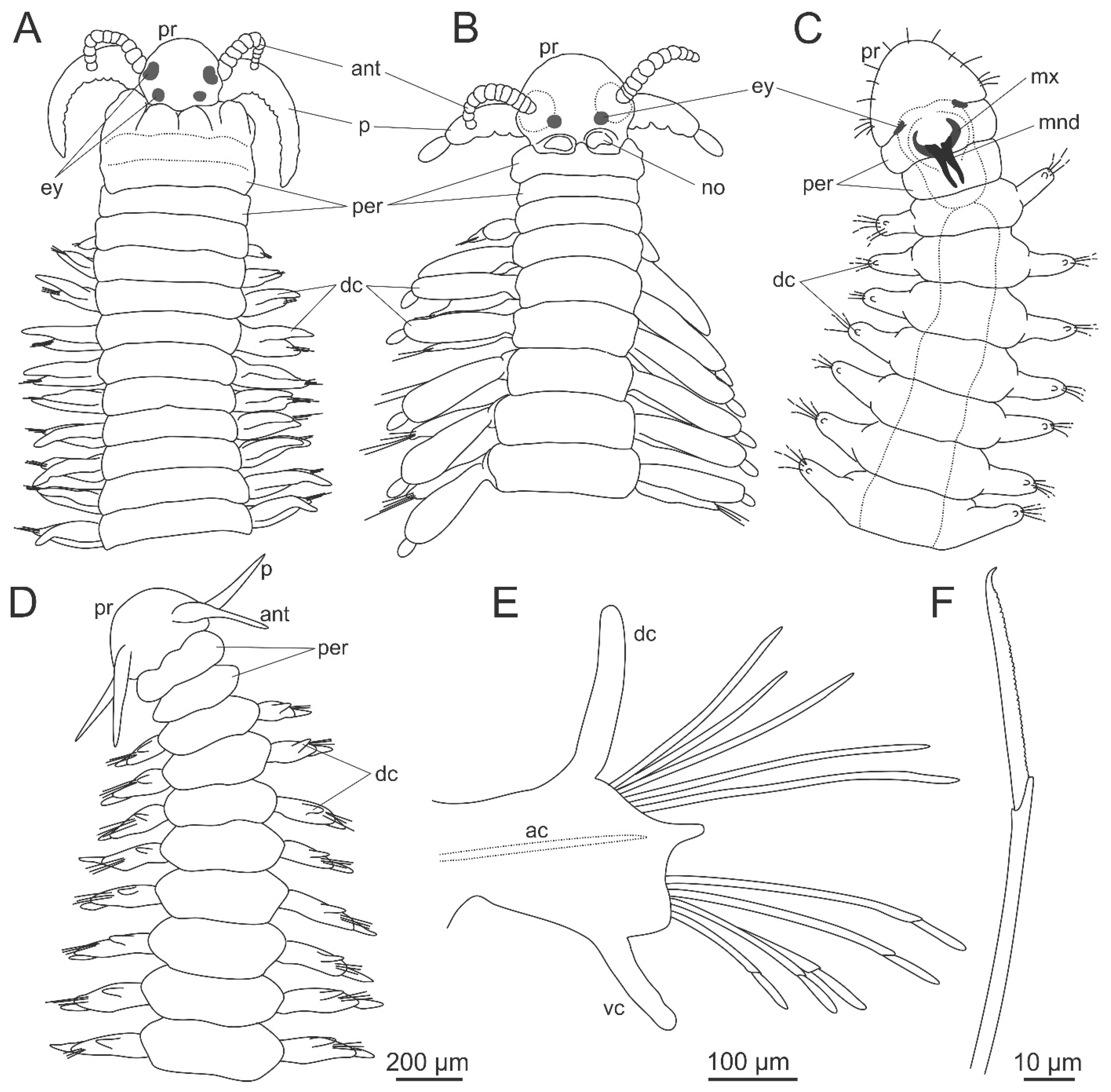{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
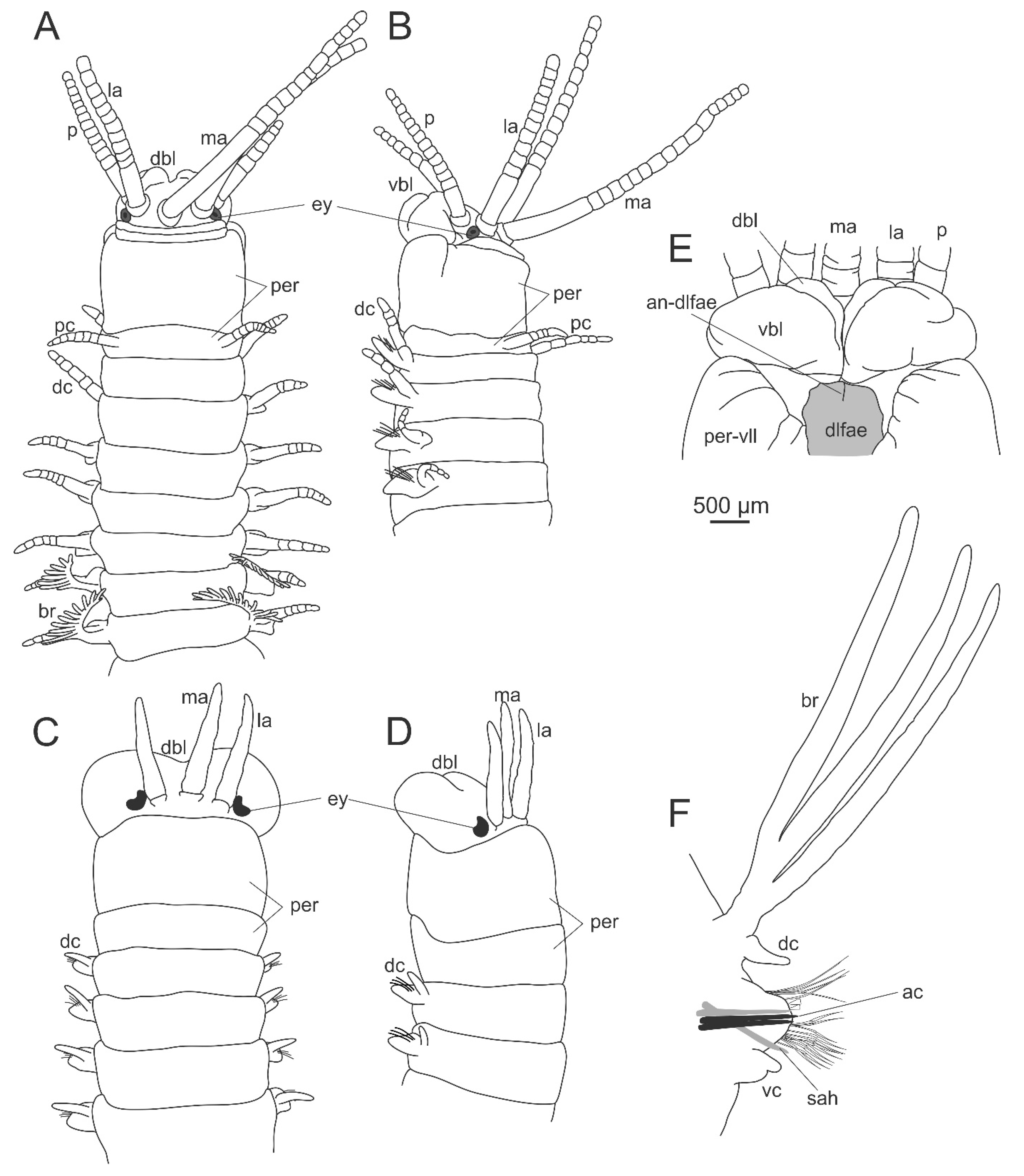
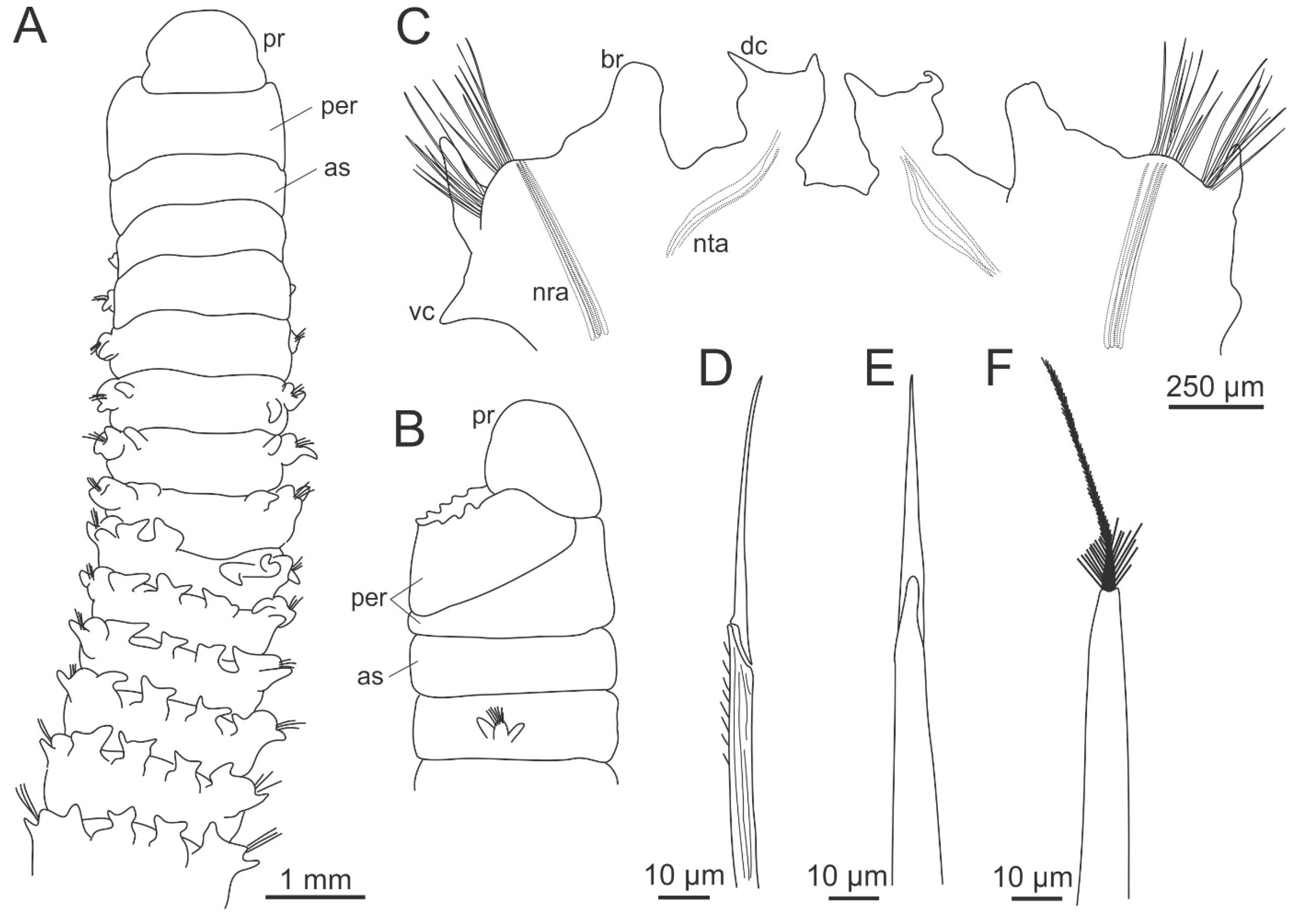
2. Dorvilleidae Chamberlin, 1919
2.1. Systematics and Phylogeny
2.2. Morphology
2.3. Feeding and Life History
2.4. Habitats and Ecology
2.5. Distribution Patterns and Diversity
2.6. Exploitation
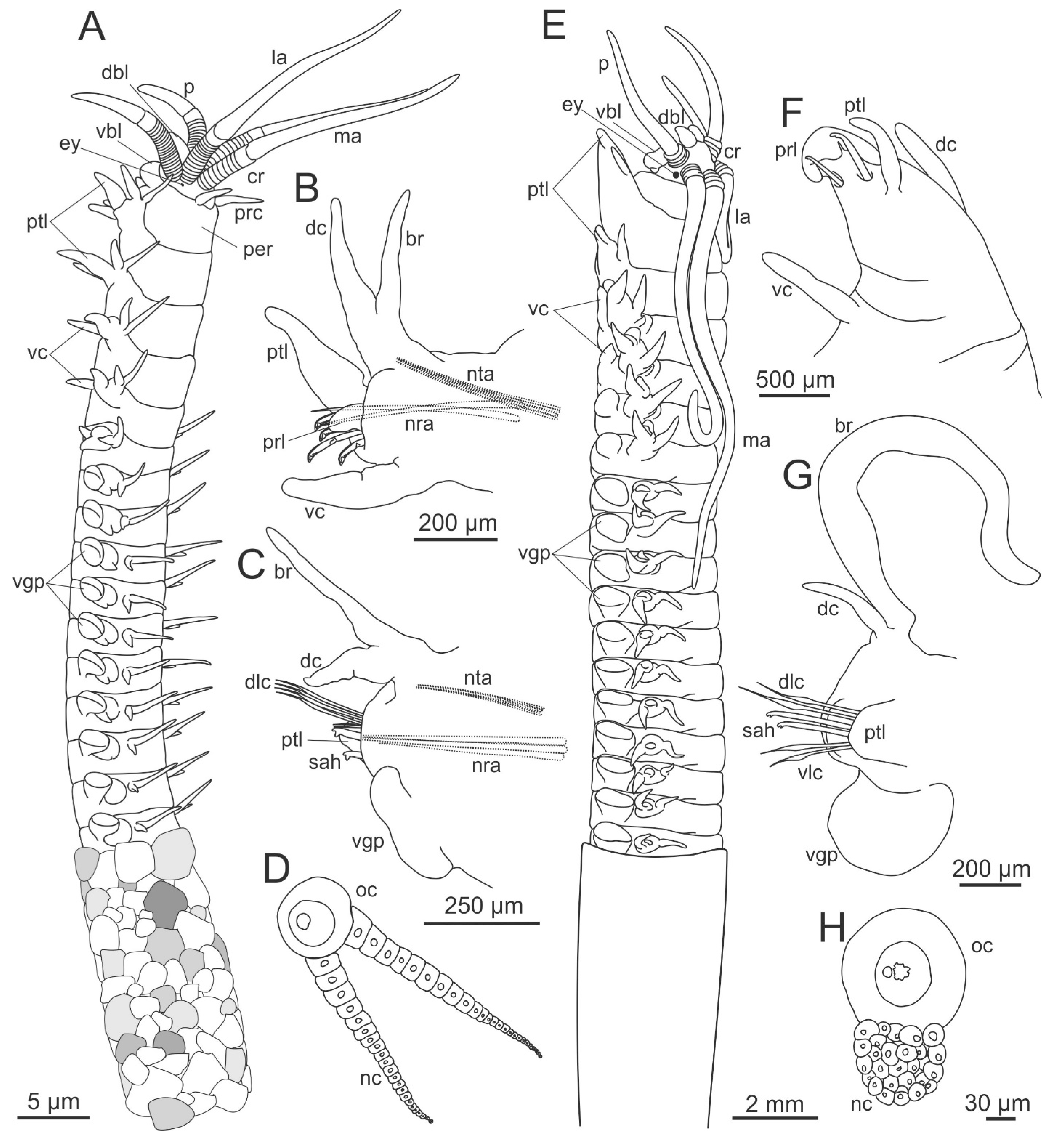
3. Eunicidae Berthold, 1827
3.1. Systematics and Phylogeny
3.2. Morphology
3.3. Feeding and Life History
3.4. Habitats and Ecology
3.5. Distribution Patterns and Diversity
3.6. Exploitation
4. Hartmaniellidae Imajima 1977
4.1. Systematics and Phylogeny
4.2. Morphology
4.3. Feeding and Life History
4.4. Habitat, Distribution and Diversity
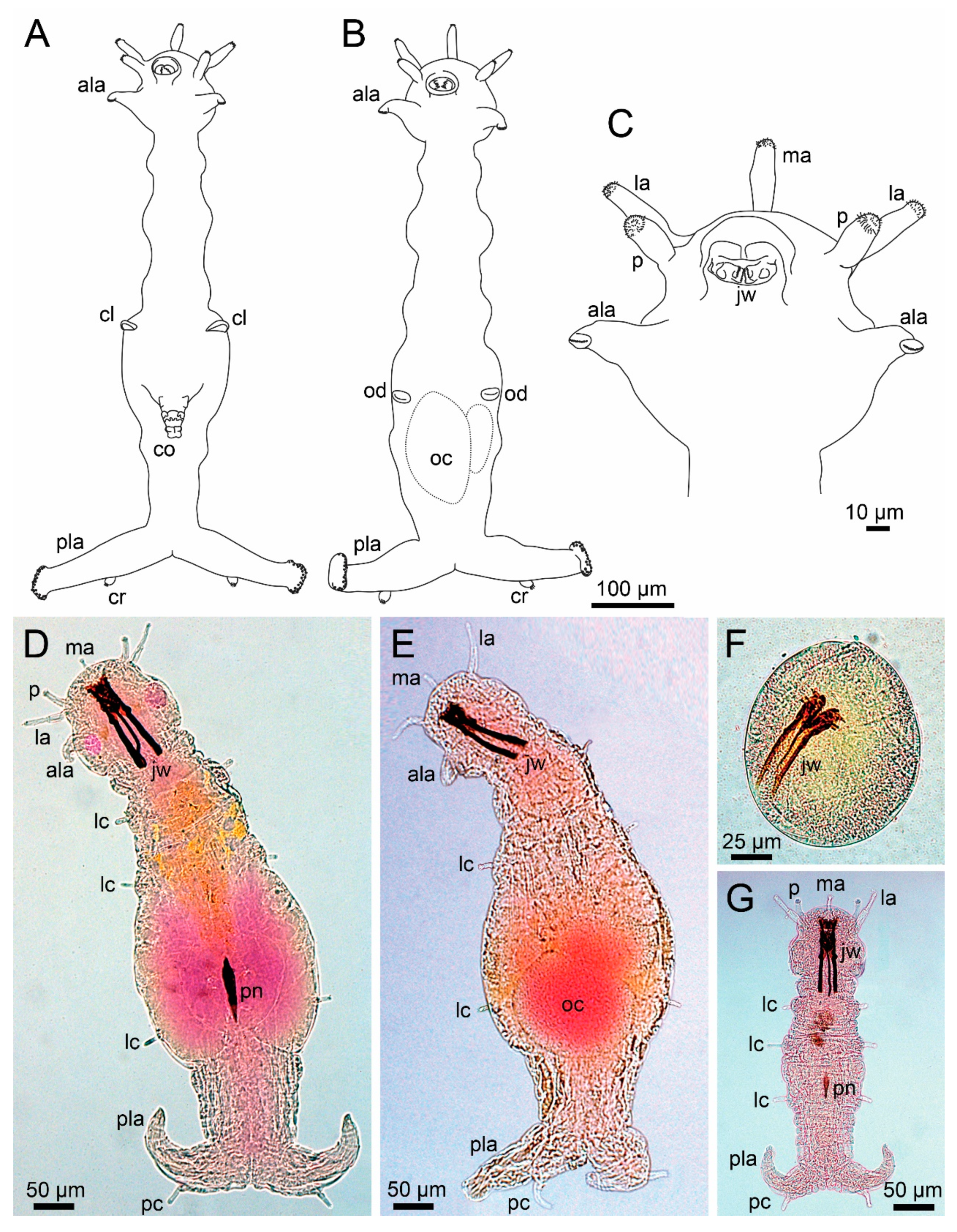
5. Histriobdellidae Claus and Moquin-Tandon, 1884
5.1. Systematics and Phylogeny
5.2. Morphology
5.3. Feeding and Life History
5.4. Habitats and Ecology
5.5. Distribution Patterns and Diversity
6. Lumbrineridae Schmarda, 1861
6.1. Systematics and Phylogeny
6.2. Morphology
6.3. Feeding and Life History
6.4. Habitats and Ecology
6.5. Distribution Patterns and Diversity
6.6. Exploitation
7. Oenonidae Kinberg, 1865
7.1. Systematics and Phylogeny
7.2. Morphology
7.3. Feeding and Life History
7.4. Habitats and Ecology
7.5. Distribution Patterns and Diversity
7.6. Exploitation
8. Onuphidae Kinberg, 1865
8.1. Systematics and Phylogeny
8.2. Morphology
8.3. Feeding and Life History
8.4. Habitats and Ecology
8.5. Distribution Patterns and Diversity
8.6. Exploitation
9. Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Uschakov, P.V. Phylum Annelida, Class Polychaeta. In Atlas of the Invertebrates of the Far Eastern Seas of the USSR; Pavlovskii, E.N., Ed.; Stony Brook University: Stony Brook, NY, USA, 1955; pp. 109–128. [Google Scholar]
- Dales, R.P. The polychaete stomodeum and the inter-relationships of the families of Polychaeta. Proc. Zool. Soc. Lond. 1962, 139, 389–428. [Google Scholar] [CrossRef]
- Ehlers, E. Die Borstenwürmer (Annelida Chaetopoda) Nach Systematischen und Anatomischen Untersuchungen Dargestellt. Volume II; Wilhelm Engelmann: Leipzig, Germany, 1868; p. 748. [Google Scholar]
- Fauvel, P. Polychètes Errantes. Faune Fr. 1923, 5, 1–488. [Google Scholar]
- Budaeva, N.; Zanol, J. Eunicida Dales, 1962. In Handbook of Zoology Annelida, Volume 3: Sedentaria III, Errantia I; Purschke, G., Westheide, W., Böggemann, M., Eds.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2021; pp. 353–360. [Google Scholar]
- Struck, T.H.; Golombek, A.; Weigert, A.; Franke, F.A.; Westheide, W.; Purschke, G.; Bleidorn, C.; Halanych, K.M. The Evolution of Annelids Reveals Two Adaptive Routes to the Interstitial Realm. Curr. Biol. 2015, 25, 1993–1999. [Google Scholar] [CrossRef] [Green Version]
- Weigert, A.; Helm, C.; Meyer, M.; Nickel, B.; Arendt, D.; Hausdorf, B.; Santos, S.R.; Halanych, K.M.; Purschke, G.; Bleidorn, C.; et al. Illuminating the base of the Annelid tree using transcriptomics. Mol. Biol. Evol. 2014, 31, 1391–1401. [Google Scholar] [CrossRef] [PubMed]
- Struck, T.H.; Purschke, G.; Halanych, K.M. Phylogeny of Eunicida (Annelida) and exploring data congruence using a partition addition bootstrap alteration (PABA) approach. Syst. Biol. 2006, 55, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Parry, L.A.; Edgecombe, G.D.; Eibye-Jacobsen, D.; Vinther, J. The impact of fossil data on annelid phylogeny inferred from discrete morphological characters. Proc. R. Soc. B Biol. Sci. 2016, 283, 20161378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanol, J.; Halanych, K.M.; Struck, T.H.; Fauchald, K. Phylogeny of the bristle worm family Eunicidae (Eunicida, Annelida) and the phylogenetic utility of noncongruent 16S, COI and 18S in combined analyses. Mol. Phylogenet. Evol. 2010, 55, 660–676. [Google Scholar] [CrossRef]
- Zanol, J.; Halanych, K.M.; Fauchald, K. Reconciling taxonomy and phylogeny in the bristleworm family Eunicidae (polychaete, Annelida). Zool. Scr. 2014, 43, 79–100. [Google Scholar] [CrossRef]
- Budaeva, N.; Schepetov, D.; Zanol, J.; Neretina, T.; Willassen, E. When molecules support morphology: Phylogenetic reconstruction of the family Onuphidae (Eunicida, Annelida) based on 16S rDNA and 18S rDNA. Mol. Phylogenet. Evol. 2016, 94, 791–801. [Google Scholar] [CrossRef]
- Orensanz, J.M. The Eunicemorph polychaete annelids from Antarctic and Subantarctic Seas. With addenda to the Eunicemorpha of Argentina, Chile, New Zealand, Australia, and the Southern Indian Ocean. Antarct. Res. Ser. 1990, 52, 1–183. [Google Scholar]
- Paxton, H. Phylogeny of Eunicida (Annelida) based on morphology of jaws. Zoosymposia 2009, 264, 241–264. [Google Scholar] [CrossRef] [Green Version]
- Read, G.; Fauchald, K. World Polychaeta Database. Eunicida. Available online: http://www.marinespecies.org/polychaeta/aphia.php?p=taxdetails&id=895 (accessed on 2 December 2020).
- Wiklund, H.; Glover, A.G.; Dahlgren, T.G. Three new species of Ophryotrocha (Annelida: Dorvilleidae) from a whale-fall in the North-East Atlantic. Zootaxa 2009, 2228, 43–56. [Google Scholar] [CrossRef] [Green Version]
- Budaeva, N. Leptoecia midatlantica, a new species of the deep-sea quill-worms (Polychaeta: Onuphidae: Hyalinoeciinae) from the Mid-Atlantic Ridge. Zootaxa 2012, 3176, 45–60. [Google Scholar]
- Yen, N.K.; Rouse, G.W. Phylogeny, biogeography and systematics of Pacific vent, methane seep, and whale-fall Parougia (Dorvilleidae: Annelida), with eight new species. Invertebr. Syst. 2020, 34, 200–233. [Google Scholar]
- Hilbig, B.; Blake, J.A. Dorvilleidae (Annelida: Polychaeta) from the U.S. Atlantic slope and rise. Description of two new genera and 14 new species, with generic revision of Ophryotrocha. Zool. Scr. 1991, 20, 147–183. [Google Scholar] [CrossRef]
- Westheide, W.; Riser, N.W. Morphology and phylogenetic relationships of the neotenic interstitial polychaete Apodotrocha progenerans n.gen., n.sp. (Annelida). Zoomorphology 1983, 103, 67–87. [Google Scholar] [CrossRef]
- Westheide, W.; von Nordheim, H. Interstitial Dorvilleidae (Annelida, Polychaeta) from Europe Australia and New Zealand. Zool. Scr. 1985, 14, 183–199. [Google Scholar] [CrossRef]
- Westheide, W. Ikosipodus carolensis gen. et sp. n., an interstitial neotenic polychaete from North Carolina, U.S.A., and its phylogenetic relationships within Dorvilleidae. Zool. Scr. 1982, 11, 117–126. [Google Scholar] [CrossRef]
- Núñez, J. Marycarmenia lysandrae, a new genus and interstitial species (Polychaeta: Dorvilleidae) from Madeira. Bull. Mar. Sci. 1998, 62, 115–119. [Google Scholar]
- Zanol, J.; da Silva, T.D.S.C.; Hutchings, P. Marphysa (Eunicidae, polychaete, Annelida) species of the Sanguinea group from Australia, with comments on pseudo-cryptic species. Invertebr. Biol. 2016, 135, 328–344. [Google Scholar] [CrossRef]
- Molina-Acevedo, I.C.; Carrera-Parra, L.F. Reinstatement of three species of the Marphysa sanguinea complex (Polychaeta: Eunicidae) from the Grand Caribbean Region. Zootaxa 2015, 3925, 37–55. [Google Scholar] [CrossRef] [PubMed]
- Borisova, P.B.; Schepetov, D.M.; Budaeva, N.E. Aponuphis Kucheruk, 1978 (Annelida: Onuphidae) from western African waters. Invertebr. Zool. 2018, 15, 19–41. [Google Scholar] [CrossRef]
- Hints, O.; Eriksson, M.E. Diversification and biogeography of scolecodont-bearing polychaetes in the Ordovician. Palaeogeogr. Palaeoclim. Palaeoecol. 2007, 245, 95–114. [Google Scholar] [CrossRef]
- Hartman, O. Polychaetous Annelids. Part V. Eunicea. Allan Hancock Pac. Exped. 1944, 10, 1–237. [Google Scholar]
- Kielan-Jaworowska, Z. Polychaete jaw apparatuses from the Ordovician and Silurian of Poland and a comparison with modern forms. Palaeontol. Pol. 1966, 16, 9–152. [Google Scholar]
- Eibye-Jacobsen, D.; Kristensen, R.M. A new genus and species of Dorvilleidae (Annelida, Polychaeta) from Bermuda, with a phylogenetic analysis of Dorvilleidae, Iphitimidae and Dinophilidae. Zool. Scr. 1994, 23, 107–131. [Google Scholar] [CrossRef]
- Ravara, A.; Marçal, A.R.; Wiklund, H.; Hilário, A. First account on the diversity of Ophryotrocha (Annelida, Dorvilleidae) from a mammal-fall in the deep-Atlantic Ocean with the description of three new species. Syst. Biodivers. 2015, 13, 875–890. [Google Scholar] [CrossRef]
- Budaeva, N.; Fauchald, K. Phylogeny of the Diopatra generic complex with a revision of Paradiopatra Ehlres, 1887 (Polychaeta: Onuphidae). Zool. J. Linn. Soc. 2011, 163, 319–436. [Google Scholar] [CrossRef] [Green Version]
- Clemo, W.C.; Dorgan, K.M. Functional morphology of eunicidan (Polychaeta) jaws. Biol. Bull. 2017, 233, 227–241. [Google Scholar] [CrossRef]
- Jumars, P.A.; Dorgan, K.M.; Lindsay, S.M. Diet of worms emended: An update of polychaete feeding guilds. Annu. Rev. Mar. Sci. 2015, 7, 497–520. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, M.E.; Parry, L.A.; Rudkin, D.M. Earth’s oldest ‘Bobbit worm’—Gigantism in a Devonian eunicidan polychaete. Sci. Rep. 2017, 7, 43061. [Google Scholar] [CrossRef] [PubMed]
- Fauchald, K. A review of the genus Eunice (Eunicidae: Polychaeta) based upon type material. Smithson. Contrib. Zool. 1992, 523, 1–422. [Google Scholar] [CrossRef] [Green Version]
- Zanol, J. Homology of prostomial and pharyngeal structures in Eunicida (Annelida) based on innervation and morphological similarities. J. Morphol. 2010, 271, 1023–1043. [Google Scholar] [CrossRef] [PubMed]
- Tzetlin, A.; Purschke, G. Pharynx and intestine. Hydrobiologia 2005, 535, 199–225. [Google Scholar]
- Tilic, E.; Bartolomaeus, T.; Rouse, G.W. Chaetal type diversity increases during evolution of Eunicida (Annelida). Org. Divers. Evol. 2016, 16, 105–119. [Google Scholar] [CrossRef]
- Starunov, V. Musculature and innervation of the pygidium in Eunicida (Annelida: Errantia). Biol. Commun. 2019, 64, 31–40. [Google Scholar] [CrossRef]
- Fernández, P.V.; Bone, D.; Rodríguez, C. Cryptofaunal polychaetes from oceanic and continental reefs in Venezuela. Bull. Mar. Sci. 2012, 88, 339–352. [Google Scholar] [CrossRef]
- Hernández-Alcántara, P.; Cruz-Pérez, I.N.; Solís-Weiss, V. Composition and diversity patterns of Eunicida and Amphinomida (Annelida) associated to dead coral in the Chinchorro Bank Biosphere Reserve, Caribbean Sea. J. Mar. Biol. Assoc. UK 2019, 99, 1547–1555. [Google Scholar] [CrossRef]
- Seixas, V.C.; Steiner, T.M.; Solé-Cava, A.M.; Amaral, A.C.Z.; Paiva, P.C. Hidden diversity within the Diopatra cuprea complex (Annelida: Onuphidae): Morphological and genetics analyses reveal four new species in the south-west Atlantic. Zool. J. Linn. Soc. 2020, 1840, 1–35. [Google Scholar] [CrossRef]
- Taboada, S.; Leiva, C.; Bas, M.; Schult, N.; McHugh, D. Cryptic species and colonization processes in Ophryotrocha (Annelida, Dorvilleidae) inhabiting vertebrate remains in the shallow-water Mediterranean. Zool. Scr. 2017, 46, 611–624. [Google Scholar] [CrossRef]
- Hutchings, P.A.; Karageorgopoulos, P. Designation of a neotype of Marphysa sanguinea (Montagu, 1813) and a description of a new species of Marphysa from eastern Australia. Hydrobiologia 2003, 496, 87–94. [Google Scholar] [CrossRef]
- van Rensburg, H.; Matthee, C.A.; Simon, C.A. Moonshine worms Diopatra aciculata (Onuphidae, Annelida) in the Knysna Estuary, South Africa; taxonomy and distribution. J. Mar. Biol. Assoc. UK 2020, 100, 897–907. [Google Scholar] [CrossRef]
- Lavesque, N.; Hutchings, P.; Abe, H.; Daffe, G.; Gunton, L.M.; Glasby, C.J. Confirmation of the exotic status of Marphysa victori Lavesque, Daffe, Bonifácio & Hutchings, 2017 (Annelida) in French waters and synonymy of Marphysa bulla Liu, Hutchings & Kupriyanova, 2018. Aquat. Invasions 2020, 15, 355–366. [Google Scholar] [CrossRef]
- Paxton, H.; Wiklund, H.; Alexander, F.; Taboada, S. Is the Antarctic Ophryotrocha orensanzi (Annelida: Dorvilleidae) a circumpolar non-specialized opportunist? Syst. Biodivers. 2017, 15, 105–114. [Google Scholar] [CrossRef]
- Schulze, A. Phylogeny and genetic diversity of Palolo worms (Palola, Eunicidae) from the tropical North Pacific. Biol. Bull. 2006, 210, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Rouse, G.W. The epitome of hand waving? Larval feeding and hypotheses of metazoan phylogeny. Evol. Dev. 2000, 2, 222–233. [Google Scholar] [CrossRef]
- Steiner, T.M.; Amaral, A.C.Z. Arabella aracaensis, a new species with growth rings on its mandibles, and some remarks on the endoparasitic Labrorostratus prolificus (Polychaeta: Oenonidae) from southeast Brazil. J. Nat. Hist. 2009, 43, 2537–2551. [Google Scholar] [CrossRef]
- Åkesson, B.; Rice, S.A. Two new Dorvillea species (Polychaeta, Dorvilleidae) with obligate asexual reproduction. Zool. Scr. 1992, 21, 351–362. [Google Scholar] [CrossRef]
- Paxton, H. Taxonomy and aspects of the life history of Australian beachworms (Polychaeta: Onuphidae). Aust. J. Mar. Freshw. Res. 1979, 30, 265–294. [Google Scholar] [CrossRef]
- Gambi, M.C. Polychaetes of commercial and applied interest in Italy: An overview. Mémoires Muséum Natl. D’histoire Nat. 1994, 162, 593–601. [Google Scholar]
- Bettoso, N.; Faverio, G.; Orel, G. Note sulla biologia e sulla pesca del verme de rimini (Eunice aphroditois Pallas) in alto Adriatico. Hydrores 1998, 16, 24–34. [Google Scholar]
- Cunha, T.; Hall, A.; Queiroga, H. Estimation of the Diopatra neapolitana annual harvest resulting from digging activity in Canal de Mira, Ria de Aveiro. Fish. Res. 2005, 76, 56–66. [Google Scholar] [CrossRef]
- Osman, I.; Gabr, H.; Saito, H.; El-Etreby, S. Reproductive biology of the highly commercial polychaetes in the Suez Canal. J. Mar. Biol. Assoc. UK 2010, 90, 281–290. [Google Scholar] [CrossRef]
- Osman, I.H.; Gabr, H.R.; El-etreby, S.G. Rearing trials of Halla parthenopeia under laboratory conditions (Polychaeta: Oenonidae). J. Exp. Mar. Biol. Ecol. 2010, 383, 1–7. [Google Scholar] [CrossRef]
- Safarik, M.; Redden, A.M.; Schreider, M.J. Density-dependent growth of the polychaete Diopatra aciculata. Sci. Mar. 2006, 70, 337–341. [Google Scholar]
- Mondragón, C. Of Winds, Worms and Mana: The Traditional Calendar of the Torres Islands, Vanuatu. Oceania 2004, 74, 289–308. [Google Scholar] [CrossRef]
- Chamberlin, R.V. The Annelida Polychaeta. Mem. Mus. Comp. Zool. Harv. Coll. 1919, 48, 1–514. [Google Scholar]
- Núñez, J. Annelida Polychaeta: Staurocephalidae Verrill, 1900. In Annelida Polychaeta V, Fauna Ibérica, Vol. 45; Etal, P.J., Ed.; Museo Nacional de Ciencias Naturales. Consejo Superior de Investigaciones Cientificas: Madrid, Spain, 2018. [Google Scholar]
- Salazar-Vallejo, S.; Carrera-Parra, L.F. Reversal of precedence for Dorvillea Parfitt, 1866 (Annelida, Errantia) over its senior homonym Dorvillea Leach, 1852 (Mollusca, Bivalvia): Maintenance of prevailing usage for Dorvillea Parfitt, 1866 and Dorvilleidae Chamberlin, 1919. Bull. Zool. Nomencl. 2020, 77, 105–111. [Google Scholar] [CrossRef]
- delle Chiaje, S. Memorie Sulla Storia e Notomia Degli Animali Senza Vertebre del Regno di Napoli. Volume 3; Stamperia della Societá Tipografica: Naples, Italy, 1828; p. 232. [Google Scholar]
- Pettibone, M.H. New species of polychaete worms from the Atlantic Ocean, with a revision of the Dorvilleidae. Proc. Biol. Soc. Wash. 1961, 74, 167–186. [Google Scholar]
- Orensanz, J.M. Los anelidos poliquetos de la provincia biogeografica Argentina. III. Dorvilleidae. Physis 1973, 32, 325–342. [Google Scholar]
- Jumars, P.A. A generic revision of the Dorvilleidae (Polychaeta), with six new species from the deep North Pacific. Zool. J. Linn. Soc. 1974, 54, 101–135. [Google Scholar] [CrossRef]
- Wolf, P.S. Four new genera of Dorvilleidae (Annelida: Polychaeta) from the Gulf of Mexico. Proc. Biol. Soc. Wash. 1986, 99, 616–626. [Google Scholar]
- Wiklund, H.; Purschke, G.; Ravara, A. Dorvilleidae Chamberlin, 1919. In Handbook of Zoology Annelida, Volume 3: Sedentaria III, Errantia I; Purschke, G., Böggemann, M., Westheide, W., Eds.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2021; pp. 361–382. ISBN 978-3-11-029148-3. [Google Scholar]
- Dahlgren, T.G.; Åkesson, B.; Schander, C.; Halanych, K.M.; Sundberg, P. Molecular Phylogeny of the Model Annelid Ophryotrocha. Biol. Bull. 2001, 201, 193–203. [Google Scholar] [CrossRef] [Green Version]
- Heggoy, K.K.; Schander, C.; Åkesson, B. The phylogeny of the annelid genus Ophryotrocha (Dorvilleidae). Mar. Biol. Res. 2007, 3, 412–420. [Google Scholar] [CrossRef]
- Ravara, A.; Wiklund, H.; Cunha, M.R. New species and records of Dorvilleidae (Annelida, Polychaeta) from deep-sea organic substrata, NE Atlantic. Eur. J. Taxon. 2020, in press. [Google Scholar]
- Taboada, S.; Wiklund, H.; Glover, A.G.; Dahlgren, T.G.; Cristobo, J.; Ávila, C. Two new Antarctic Ophryotrocha (Annelida: Dorvilleidae) described from shallow-water whale bones. Polar Biol. 2013, 36, 1031–1045. [Google Scholar] [CrossRef]
- Wiklund, H.; Iris, V.; Glover, A.G.; Smith, C.R.; Baco, A.M.Y.R.; Dahlgren, T.G. Systematics and biodiversity of Ophryotrocha (Annelida, Dorvilleidae) with descriptions of six new species from deep-sea whale-fall and wood-fall habitats in the north-east Pacific. Syst. Biodivers. 2012, 10, 243–259. [Google Scholar] [CrossRef]
- Struck, T.H.; Halanych, K.M.; Purschke, G. Dinophilidae (Annelida) is most likely not a progenetic Eunicida: Evidence from 18S and 28S rDNA. Mol. Phylogenet. Evol. 2005, 37, 619–623. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, R.M.; Eibye-Jacobsen, D. Ultrastructure of spermiogenesis and spermatozoa in Diurodrilus subterraneus (Polychaeta, Diurodrilidae). Zoomorphology 1995, 115, 117–132. [Google Scholar] [CrossRef]
- Laumer, C.E.; Bekkouche, N.; Kerbl, A.; Goetz, F.; Neves, R.C.; Sørensen, M.V.; Kristensen, R.M.; Hejnol, A.; Dunn, C.W.; Giribet, G.; et al. Spiralian Phylogeny Informs the Evolution of Microscopic Lineages. Curr. Biol. 2015, 25, 2000–2006. [Google Scholar] [CrossRef] [Green Version]
- Simonini, R.; Massamba-N’Siala, G.; Grandi, V.; Prevedelli, D. Distribution of the genus Ophryotrocha (Polychaeta) in Italy: New records and comments on the biogeography of Mediterranean species. Vie Milieu-Life Environ. 2009, 59, 79–88. [Google Scholar]
- Åkesson, B. Reproduction and larval morphology of five Ophryotrocha species (Polychaeta, Dorvilleidae). Zool. Scr. 1973, 2, 145–155. [Google Scholar] [CrossRef]
- Åkesson, B. Reproduction in the genus Ophryotrocha (Polychaeta, Dorvilleidae). Pubbl. della Stn. Zool. di Napoli 1975, 39, 377–398. [Google Scholar]
- Oug, E. New and lesser known Dorvilleidae (Annelida, Polychaeta) from Scandinavian and northeast American waters. Sarsia 1978, 63, 285–303. [Google Scholar]
- Purschke, G.; Nowak, K. Ultrastructure of pigmented eyes in Dorvilleidae (Annelida, Errantia, Eunicida) and their importance for understanding the evolution of eyes in polychaetes. Acta Zool. 2015, 96, 67–81. [Google Scholar] [CrossRef]
- Richards, T.L. Reproduction and development of the polychaete Stauronereis rudolphi, including a summary of development in the superfamily Eunicea. Mar. Biol. 1967, 1, 124–133. [Google Scholar] [CrossRef]
- Jamieson, B.G.M.; Rouse, G.W. The spermatozoa of the Polychaeta (Annelida): An ultrastructural review. Biol. Rev. 1989, 64, 93–157. [Google Scholar] [CrossRef]
- Åkesson, B. A new Ophryotrocha species of the Labronica group (Polychaeta, Dorvilleidae) revealed in crossbreeding experiments. In Proceedings of the NATO Conference Series (Marine Science). 2 Marine Organisms, Venice, Italy, 24 March–4 April 1977; Battaglia, B., Beardmore, J., Eds.; Plenum Publishing: New York, NY, USA, 1978; pp. 573–590. [Google Scholar]
- Prevedelli, D.; Simonini, R. Effect of temperature on demography of Ophryotrocha labronica (Polychaeta: Dorvilleidae). Vie Milieu-Life Environ. 2001, 51, 173–180. [Google Scholar]
- Åkesson, B. Ophryotrocha labronica as test animal for the study of marine pollution. Helgoländer Wiss. Meeresunters. 1970, 20, 293–303. [Google Scholar] [CrossRef] [Green Version]
- Simonini, R. Distribution and ecology of the genus Ophryotrocha (Polychaeta: Dorvilleidae) in Italian harbors and lagoons. Vie Milieu-Life Environ. 2002, 2, 59–65. [Google Scholar]
- Paxton, H.; Davey, A. A new species of Ophryotrocha (Annelida: Dorvilleidae) associated with fish farming at Macquarie Harbour, Tasmania, Australia. Zootaxa 2010, 2509, 53–61. [Google Scholar] [CrossRef] [Green Version]
- Salvo, F.; Wiklund, H.; Dufour, S.D.; Hamoutene, D.; Pohle, G.; Worsaae, K. A new annelid species from whalebones in Greenland and aquaculture sites in Newfoundland: Ophryotrocha cyclops, sp. nov. (Eunicida: Dorvilleidae). Zootaxa 2014, 3887, 555–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonini, R.; Grandi, V.; Massamba-N’Siala, G.; Pia Martino, M.; Castelli, A.; Prevedelli, D. Diversity, habitat affinities and diet of Ophryotrocha species (Polychaeta, Dorvilleidae) living in Mediterranean harbour habitats. Vie Milieu-Life Environ. 2010, 60, 27–38. [Google Scholar]
- Taboada, S.; Bas, M.; Ávila, C. A new Parougia species (Annelida, Dorvilleidae) associated with eutrophic marine habitats in Antarctica. Polar Biol. 2015, 38, 517–527. [Google Scholar] [CrossRef]
- Detinova, N.N. Polychaetous worms from the Reykjanes Ridge (the North Atlantic) Bottom Fauna from Mid-Ocean Rises in the North Atlantic Donnaya fauna. Otkryto-Okeanicheskikh Pod. Sev. Atl. Tr. Institua Okeanol. 1985, 120, 96–136. [Google Scholar]
- Fauchald, K. Benthic polychaetous annelids from deep water off western Mexico and adjacent areas in the Eastern Pacific Ocean. Allan Hancock Monogr. Mar. Biol. 1972, 7, 1–575. [Google Scholar]
- Hilbig, B. Reconsideration of the genus Exallopus Jumars, 1974 (Polychaeta: Dorvilleidae) with descriptions of two new species from the western North Atlantic. Ophelia 1991, S5, 101–110. [Google Scholar]
- Thornhill, D.J.; Struck, T.H.; Ebbe, B.; Lee, R.W.; Mendoza, G.F.; Levin, L.A.; Halanych, K.M. Adaptive radiation in extremophilic Dorvilleidae (Annelida): Diversification of a single colonizer or multiple independent lineages? Ecol. Evol. 2012, 2, 1958–1970. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.-S.; Zhou, Y.-D.; Wang, C.-S.; Rouse, G. A new species of Ophryotrocha (Annelida, Eunicida, Dorvilleidae) from hydrothermal vents on the Southwest Indian Ridge. Zookeys 2017, 687, 1–9. [Google Scholar]
- Blake, J.A. Polychaeta from the vicinity of deep-sea geothermal vents in the eastern Pacific. I: Euphrosinidae, Phyllodocidae, Hesionidae, Nereididae, Glyceridae, Dorvilleidae, Orbiniidae and Maldanidae. Bull. Biol. Soc. Wash. 1985, 6, 67–101. [Google Scholar]
- Blake, J.A.; Hilbig, B. Polychaeta from the vicinity of deep-sea hydrothermal vents in the Eastern Pacific Ocean. II. New species and records from the Juan de Fuca and Explorer Ridge systems. Pac. Sci. 1990, 44, 219–253. [Google Scholar]
- Hilbig, B.; Fiege, D. A new species of Dorvilleidae (Annelida: Polychaeta) from a cold seep site in the northeast Pacific. Proc. Biol. Soc. Wash. 2001, 114, 396–402. [Google Scholar]
- Paxton, H.; Morineaux, M. Three species of Dorvilleidae (Annelida: Polychaeta) associated with Atlantic deep-sea reducing habitats, with the description of Ophryotrocha fabriae, new species. Proc. Biol. Soc. Wash. 2009, 122, 14–25. [Google Scholar] [CrossRef]
- Solís-Weiss, V.; Hilbig, B. Redescription of Ophryotrocha platykephale Blake (Polychaeta, Dorvilleidae) from the Guaymas Basin hydrothermal vents. Bull. South. Calif. Acad. Sci. 1992, 91, 92–96. [Google Scholar]
- Fauchald, K. Polychaetous annelids of the families Eunicidae, Lumbrineridae, Iphitimidae, Arabellidae, Lysaretidae and Dorvilleidae from western Mexico. Allan Hancock Monogr. Mar. Biol. 1970, 5, 1–335. [Google Scholar]
- Wolf, P.S. Family Dorvilleidae Chamberlin, 1919. In Taxonomic Guide to the Polychaetes of the Northern Gulf of Mexico. Final Report to the Minerals Management Service, Contract 14-12-001-29091. Volume VI; Uebelacker, J.M., Johnson, P.G., Eds.; Barry A. Vittor & Associates: Mobile, AL, USA, 1984; pp. 44.1–44.37. [Google Scholar]
- Wolf, P.S. Three new species of Dorvilleidae (Annelida: Polychaeta) from Puerto Rico and Florida and a new genus for dorvillleids from Scandinavia and North America. Proc. Biol. Soc. Wash. 1986, 99, 627–638. [Google Scholar]
- Hilbig, B. Family Dorvilleidae Chamberlin, 1919. In Taxonomic Atlas of the Benthic Fauna of the Santa Maria Basin and the Western Santa Barbara Channel. Vol. 5. The Annelida Part 2, Polychaeta: Phyllodocida (Syllidae and Scale-Bearing Families), Amphinomida, and Eunicida; Lake, J.A., Hilbig, B., Scott, P.H., Eds.; Santa Barbara Museum of Natural History Publications: Santa Barbara, CA, USA, 1995; pp. 341–364. [Google Scholar]
- Paiva, P.C.; Nonato, E.F. On the genus Iphitime (Polychaeta: Iphitimidae) and description of Iphitime sartorae sp. nov. a commensal of brachyuran crabs. Ophelia 1991, 34, 209–215. [Google Scholar] [CrossRef]
- Imajima, M. Dorvilleidae (Annelida, Polychaeta) from Japan. I. The genus Dorvillea (Dorvillea). Bull. Natl. Sci. Mus. Tokyo 1992, 18, 131–147. [Google Scholar]
- Núñez, J.; Pascual, M.; Brito, M.C. Contribución al estudio de los Dorvilleidos (Annelida, Polychaeta) de Canarias. Rev. Acad. Canar. Cienc. 1996, 8, 139–151. [Google Scholar]
- Hartmann-Schröder, G.; Rosenfeldt, P. Die Polychaeten der ’Polarstern’-Reise ANT V/1 in die Antarktis 1986. Teil 1: Euphrosinidae bis Iphitimidae. Mitt. Hambg. Zool. Mus. Inst. 1992, 89, 85–124. [Google Scholar]
- Cunha, M.R.; Matos, F.L.; Génio, L.; Hilário, A.; Moura, C.J.; Ravara, A.; Rodrigues, C.F. Are organic falls bridging reduced environments in the deep sea?—Results from colonization experiments in the Gulf of Cádiz. PLoS ONE 2013, 8, e76688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levin, L.A.; Ziebis, W.; Mendoza, G.F.; Growney, V.A.; Tryon, M.D.; Brown, K.M.; Mahn, C.; Gieskes, J.M.; Rathburn, A.E. Spatial heterogeneity of macrofauna at northern California methane seeps: Influence of sulfide concentration and fluid flow. Mar. Ecol. Prog. Ser. 2003, 265, 123–139. [Google Scholar] [CrossRef]
- Prevedelli, D.; Massamba-N’Siala, G.; Simonini, R. The seasonal dynamics of six species of Dorvilleidae (Polychaeta) in the harbour of La Spezia (Italy). Mar. Ecol. Evol. Perspect. 2005, 26, 286–293. [Google Scholar] [CrossRef]
- Fauchald, K.; Hancock, D.R. Deep-water polychaetes from a transect off central Oregon. Allan Hancock Monogr. Mar. Biol. 1981, 11, 1–73. [Google Scholar]
- Hartmann-Schröder, G. Die polychaeten der Fahrten 11, 19, 21 und 26 (1967-1972) von F.S. “Meteor” in das Gebiet zwischen dem Golf von Biscaya und dem Auftriebsgebiet vor Westafrika. Meteor Forsch.-Ergeb. 1981, 33, 23–36. [Google Scholar]
- Schulze, A.; Timm, L.E. Palolo and un: Distinct clades in the genus Palola (Eunicidae: Polychaeta). Mar. Biodivers. 2011, 42, 161–171. [Google Scholar] [CrossRef]
- Nederlof, M.A.J.; Jansen, H.M.; Dahlgren, T.G.; Fang, J.; Meier, S.; Strand, Ø.; Sveier, H.; Verdegem, M.C.J.; Smaal, A.C. Application of polychaetes in (de)coupled integrated aquaculture: Production of a high-quality marine resource. Aquac. Environ. Interact. 2019, 11, 221–237. [Google Scholar] [CrossRef] [Green Version]
- Åkesson, B. The use of certain polychaetes in bioassay studies. Rapp. Proces Verbaux Reun. 1980, 179, 315–321. [Google Scholar]
- Lamarck, J.B. Histoire Naturelle des Animaux Sans Vertèbres, Préséntant les Caractères Généraux et Particuliers de ces Animaux, Leur Distribution, Leurs Classes, Leurs Familles, Leurs Genres, et la Citation des Principales Espèces qui s’y Rapportent; Precedes d’une Introduction Offrant la Determination des Caracteres Essentiels de L’animal, sa Distinction du Vegetal et Desautres Corps Naturels, Enfin, l’Exposition des Principes Fondamentaux de la Zoologie. Vol. 5; Deterville Libraire & Verdiere Libraire: Paris, France, 1818; p. 612. [Google Scholar]
- Hartmann-Schröder, G.; Zibrowius, H. Polychaeta associated with Antipatharia (Cnidaria: Anthozoa): Description of Polynoidae and Eunicidae. Mitt. Hambg. Zool. Mus. Inst. 1998, 95, 29–44. [Google Scholar]
- Zanol, J.; Fauchald, K.; Paiva, P.C. A phylogenetic analysis of the genus Eunice (Eunicidae, polychaete, Annelida). Zool. J. Linn. Soc. 2007, 150, 413–434. [Google Scholar] [CrossRef] [Green Version]
- Molina-Acevedo, I.C.; Carrera-Parra, L.F. Revision of Marphysa de Quatrefages, 1865 and some species of Nicidion Kinberg, 1865 with the erection of a new genus (Polychaeta: Eunicidae) from the Grand Caribbean. Zootaxa 2017, 4241, 1–62. [Google Scholar] [CrossRef]
- Lu, H.; Fauchald, K. A phylogenetic and biogeographic study of Euniphysa (Eunicidae, Polychaeta). J. Nat. Hist. 2000, 34, 997–1044. [Google Scholar] [CrossRef]
- Carrera-Parra, L.F.; Fauchald, K.; Gambi, M.C. Revision of the taxonomic status of Lysidice (Polychaeta, Eunicidae) in the Western Caribbean Sea with observation on species reproductive features and habitat preference. Ital. J. Zool. 2011, 78, 27–40. [Google Scholar] [CrossRef] [Green Version]
- Molina-Acevedo, I.C.; Idris, I. Reinstatement of species belonging Marphysa sanguinea complex (Annelida: Eunicidae) and description of new species from the mid-Pacific Ocean and the Adriatic Sea. Zootaxa 2020, 4816, 1–48. [Google Scholar] [CrossRef] [PubMed]
- Kara, J.; Molina-Acevedo, I.C.; Zanol, J.; Simon, C.; Idris, I. Morphological and molecular systematic review of Marphysa Quatrefages, 1865 (Annelida: Eunicidae) species from South Africa. PeerJ 2020, 8, e10076. [Google Scholar] [CrossRef] [PubMed]
- Fauchald, K. Review of the types of Palola (Eunicidae: Polychaeta). J. Nat. Hist. 1992, 26, 1177–1225. [Google Scholar] [CrossRef]
- Molina-Acevedo, I.C. Morphological revision of the Subgroup 1 Fauchald, 1970 of Marphysa de Quatrefages, 1865 (Eunicidae: Polychaeta). Zootaxa 2018, 4480, 1–125. [Google Scholar] [CrossRef] [PubMed]
- Molina-Acevedo, I.C. Redescription of two species previously regarded as Marphysa de Quatrefages, 1865, with the description of new species of Treadwellphysa Molina-Acevedo and Carrera-Parra, 2017. J. Nat. Hist. 2019, 53, 517–540. [Google Scholar] [CrossRef]
- Carrera-Parra, L.F.; Salazar-Vallejo, S.I. A new genus and 12 new species of Eunicidae (Polychaeta) from the Caribbean Sea. J. Mar. Biol. Assoc. UK 1998, 78, 145–182. [Google Scholar] [CrossRef]
- Glasby, C.J.; Hutchings, P.A. A new species of Marphysa Quatrefages, 1865 (Polychaeta: Eunicida: Eunicidae) including the genus Nauphanta Kinberg, 1865. Zootaxa 2010, 45, 29–45. [Google Scholar] [CrossRef]
- Martin, D.; Gil, J.; Zanol, J.; Meca, M.A.; Portela, R.P. Digging the diversity of Iberian bait worms Marphysa (Annelida, Eunicidae). PLoS ONE 2020, 15, e0226749. [Google Scholar] [CrossRef] [PubMed]
- Steiner, T.M.; Nogueira, J.M.D.M.; Amaral, A.C.Z. Eunice rubra Grube (Annelida, Polychaeta, Eunicidae), a redescription based on the holotype and Brazilian specimens. Rev. Bras. Zool. 2002, 19, 243–259. [Google Scholar] [CrossRef] [Green Version]
- Parapar, J.; Freire, J.; Urgorri, V.; Besteiro, C. Morphological variability in Eunice vittata (Chiaje, 1828) (Polychaeta, Eunicidae) in the Ría de Ferrol (Galicia, NW Spain). Ophelia 1993, 37, 117–125. [Google Scholar] [CrossRef]
- Miura, T. Japanese polychaetes of the genera Eunice and Euniphysa: Taxonomy and branchial distribution patterns. Publ. Seto Mar. Biol. Lab. 1986, 31, 269–325. [Google Scholar] [CrossRef] [Green Version]
- Salazar-Vallejo, S.I.; Carrera-Parra, L.F. Eunícidos (Polychaeta) del caribe mexicano con claves para las especies del Gran Caribe: Fauchaldius, Lysidice, Marphysa, Nematonereis y Palola. Rev. Biol. Trop. 1998, 45, 1481–1498. [Google Scholar]
- Parapar, J.; Harto, I. On the presence in the Iberian waters and morphological variability in Eunice roussaei Quatrefages, 1866 (Polychaeta, Eunicidae). Bol. Real Soc. Esp. Hist. Nat. Secc. Biol. 2001, 96, 165–174. [Google Scholar]
- Costa-Paiva, E.M.; Paiva, P.C. A morphometric analysis of Eunice Cuvier (Annelida, Polychaeta) species. Rev. Bras. Zool. 2007, 24, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Treadwell, A.L. Leodicidae of the West Indian region. Carnegie Inst. Wash. Publ. 1921, 293, 1–131. [Google Scholar] [CrossRef]
- Carrera-Parra, L.F. Eunicidae Berthold, 1827. In Poliquetos (Annelida: Polychaeta) de México y América Tropical; de León-González, J.Á., Bastida-Zavala, J.R., Parra, L.F.C., García-Garza, M.E., Peña-Rivera, A., Salazar-Vallejo, S.I., Solís-Weiss, V., Eds.; Universidad Autónoma de Nuevo León: Monterrey, Mexico, 2009; pp. 165–181. [Google Scholar]
- Zanol, J.; Budaeva, N. Eunicidae. In Handbook of Zoology Annelida, Volume 3: Sedentaria III, Errantia I; Purschke, G., Westheide, W., Böggemann, M., Eds.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2021; pp. 414–452. [Google Scholar]
- Lachat, J.; Haag-Wackernagel, D. Novel mobbing strategies of a fish population against a sessile annelid predator. Sci. Rep. 2016, 6, 33187. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, D.K. Nachweise der Epitokie als Fortpflanzungsmodus des Polychaeten Eunice siciliensis (Polychaeta: Annelida). Mar. Biol. 1972, 11, 341–344. [Google Scholar] [CrossRef]
- Pamungkas, J. Species richness and macronutrient content of wawo worms (Polychaeta, Annelida) from Ambonese waters, Maluku, Indonesia. Biodiveristy Data J. 2015, 3, e4251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Southern, R. Polychaeta of the Chilka Lake and also of fresh and brackish water in other parts of India. Mem. Indian Mus. 1921, 4, 563–659. [Google Scholar]
- Aiyar, R.G. An account of the development and breeding habits of a brackish water Polychaete worm of the genus Marphysa. J. Linn. Soc. Lond. 1931, 37, 387–403. [Google Scholar] [CrossRef]
- Pillai, T.G. Studies on a brackish-water polychaetous annelid, Marphysa borradailei, sp. n. from Ceylon. Ceylon J. Sci. (Biol. Sci.) 1958, 1, 94–106. [Google Scholar]
- Pettibone, M.H. Marine polychate worms of the New England region Part 1. Aphroditidae through Trochochetidae. United States Natl. Mus. Bull. 1963, 227, 1–356. [Google Scholar]
- Åkesson, B. The embryology of the Polychaete Eunice kobiensis. Acta Zool. 1967, 48, 141–192. [Google Scholar] [CrossRef]
- Prevedelli, D.; N’siala, G.M.; Ansaloni, I.; Simonini, R. Life cycle of Marphysa sanguinea (Polychaeta: Eunicidae) in the Venice Lagoon (Italy). Mar. Ecol. 2007, 28, 384–393. [Google Scholar] [CrossRef]
- Malathi, E.; Sunder Raj, S.K.; Mercy Bai, P. Larval development of Marphysa gravelyi (Polychaeta: Eunicidae) from Pulicat Lake, India. Ital. J. Zool. 2011, 78, 249–254. [Google Scholar] [CrossRef] [Green Version]
- Herpin, R. Recherches biologiques sur la reproduction et le développement de quelques Annélides polychètes. Bull. Société Sci. Nat. L’ouest Fr. 1925, 5, 1–250. [Google Scholar]
- Goldstein, M.C.; Carson, H.S.; Eriksen, M. Relationship of diversity and habitat area in North Pacific plastic-associated rafting communities. Mar. Biol. 2014, 161, 1441–1453. [Google Scholar] [CrossRef]
- Jang, M.; Shim, W.J.; Han, G.M.; Song, Y.K.; Hong, S.H. Formation of microplastics by polychaetes (Marphysa sanguinea) inhabiting expanded polystyrene marine debris. Mar. Pollut. Bull. 2018, 131, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Díaz, O.; Liñero-Arana, I. Epifaunal polychaetes on Pinctada imbricata Roding, 1798 (Bivalvia: Pteriidae) from the Gulf of Cariaco Venezuela. Interciencia 2003, 28, 298–301. [Google Scholar]
- Hartman, O. Descriptions of new species and new generic records of polychaetous annelids from California of the families Glyceridae, Eunicidae, Stauronereidae, and Opheliidae. Univ. Calif. Publ. Zool. 1938, 43, 93–111. [Google Scholar]
- Gambi, M.C. Spatio-temporal distribution and ecological role of polychaete borers of Posidonia oceanica (L.) Delile scales. Bull. Mar. Sci. 2002, 71, 1323–1331. [Google Scholar]
- Molodtsova, T.; Budaeva, N. Modifications of corallum morphology in black corals as an effect of associated fauna. Bull. Mar. Sci. 2007, 81, 469–479. [Google Scholar]
- Quiroga, E.; Levin, L.A. Eunice pennata (Polychaeta: Eunicidae) From Active and Passive Cold Seep Sites in Central and Southern Chile (36–46° S). An. Inst. Patagon. 2010, 38, 31–37. [Google Scholar] [CrossRef]
- Mueller, C.E.; Lundälv, T.; Middelburg, J.J.; van Oevelen, D. The Symbiosis between Lophelia pertusa and Eunice norvegica stimulates coral calcification and worm assimilation. PLoS ONE 2013, 8, e58660. [Google Scholar] [CrossRef] [Green Version]
- Kise, H.; Reimer, J.D. Unexpected diversity and a new species of Epizoanthus (Anthozoa, hexacorallia) attached to eunicid worm tubes from the Pacific Ocean. Zookeys 2016, 2016, 49–71. [Google Scholar] [CrossRef] [Green Version]
- Roberts, J.M. Reef-aggregating behaviour by symbiotic eunicid polychaetes from cold-water corals: Do worms assemble reefs? J. Mar. Biol. Assoc. UK 2005, 85, 813–819. [Google Scholar] [CrossRef]
- Hutchings, P.A. Biological destruction of coral reefs. Coral Reefs 1986, 4, 239–252. [Google Scholar] [CrossRef]
- Chisholm, J.R.M.; Kelley, R. Worms start the reef-building process. Nature 2001, 409, 152. [Google Scholar] [CrossRef]
- Ochoa-Rivera, V.; Granados-Barba, A.; Solís-Weiss, V. The polychaete cryptofauna from Cozumel Island, Mexican Caribbean. Bull. Mar. Sci. 2000, 67, 137–146. [Google Scholar]
- Hartman, O. Polychaeta Errantia of Antarctica. Antarct. Res. Ser. 1964, 3, 1–131. [Google Scholar]
- Winsnes, I.M. Eunicid polychaetes (Annelida) from Scandinavian and adjacent waters. Family Eunicidae. Zool. Scr. 1989, 18, 483–500. [Google Scholar] [CrossRef]
- Amoureux, L. Annélides Polychètes profondes de Madagascar Description de deux nouvelles espèces (Collections Crosnier et Jouannie). Bull. Muséum Natl. d’Histoire Nat. 1978, 344, 1093–1108. [Google Scholar]
- Nonato, E.F. Contribuição ao Conhecimento dos Anelídeos Poliquetas Bentônicos da Plataforma Continental Brasileira, Entre Cabo Frio e o Arroio Chuí. Livre Docência Thesis, Universidade de São Paulo, São Paulo, Brazil, 1981. [Google Scholar]
- Orensanz, J.M. Los anelidos poliquetos de la provincia biogeografica Argentina. VII. Eunicidae y Lysaretidae. Physis 1975, 34, 85–111. [Google Scholar]
- Fauchald, K. Review of the types and key to the species of Eunice (Eunicidae: Polychaeta) from the Australian region. Rec. Aust. Mus. 1986, 38, 241–262. [Google Scholar] [CrossRef] [Green Version]
- Zanol, J.; da Silva, T.S.C.; Hutchings, P. One new species and two redescriptions of Marphysa (Eunicidae, Annelida) species of the Aenea-group from Australia. Zootaxa 2017, 4268, 411–426. [Google Scholar] [CrossRef] [PubMed]
- Zanol, J.; Hutchings, P.A.; Fauchald, K. Eunice sensu latu (Annelida: Eunicidae) from Australia: Description of seven new species and comments on previously reported species of the genera Eunice, Leodice and Nicidion. Zootaxa 2020, 4748, 1–43. [Google Scholar] [CrossRef]
- Schulze, A. Six genetically distinct clades of Palola (Eunicidae, Annelida) from Lizard Island, Great Barrier Reef, Australia. Zootaxa 2015, 4019, 695–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kott, P. Nereidae and Eunicidae of south Western Australia; also notes on the ecology of Western Australian limestone reefs. J. R. Soc. West. Aust. 1951, 35, 85–130. [Google Scholar]
- Morgado, E.H.; Amaral, A.C.Z. Anelidos poliquetos associados a um briozoario I. Eunicidae, Lumbrineridae, Lysaretidae e Dorvilleidae. Iheringia 1981, 60, 33–54. [Google Scholar]
- Steiner, T.; Amaral, A.C.Z. Two New Species of Marphysa Quatrefages, 1865 (Eunicidae, Polychaeta) From Intertidal Sandy Beaches of the São Sebastião Channel, State of São Paulo (Brazil). Bull. Mar. Sci. 2000, 67, 479–489. [Google Scholar]
- Zanol, J.; Paiva, P.C.; Attolini, F.S. Eunice and Palola (Eunicidae: Polychaeta) from the eastern Brazilian Coast (13°00′–22°30′ S). Bull. Mar. Sci. 2000, 67, 449–463. [Google Scholar]
- Nogueira, J.M.M.; Steiner, T.M.; Amaral, A.C.Z. Descriptions of two new species of Eunice Cuvier, 1817 (Polychaeta: Eunicidae) from coastal islands of the State of São Paulo, Brazil. Sci. Mar. 2001, 65, 47–57. [Google Scholar] [CrossRef]
- Bergamo, G.; Carrerette, O.; Zanol, J.; Nogueira, J.M.M. Species of Eunicidae (Annelida, Errantia, Eunicida) from rhodolith beds off the states of Paraíba and Pernambuco, northeastern Brazilian coast. Zootaxa 2018, 4521, 376–390. [Google Scholar] [CrossRef] [PubMed]
- George, J.D.; Hartmann-Schröder, G. Polychaetes: British Amphinomida, Spintherida and Eunicida. Keys and notes for the identification of the species. Synop. Br. Fauna 1985, 32, 1–221. [Google Scholar]
- Núñez, J.; Pascual, M.; Delgado, J.D.; Brito, M.C. Anélidos poliquetos de Canarias: Familia Eunicidae. Vieraea 1997, 26, 47–75. [Google Scholar]
- Carrera-Parra, L.F.; Salazar-Vallejo, S.I. Eunícidos (Polychaeta) del Caribe mexicano con claves para las especies del Gran Caribe: Eunice. Rev. Biol. Trop. 1998, 45, 1499–1521. [Google Scholar]
- Salazar-Vallejo, S.I.; Carrera-Parra, L.F.; de León-González, J.A. Giant Eunicid Polychaetes (Annelida) in shallow tropical and temperate seas. Rev. Biol. Trop. 2011, 59, 1463–1474. [Google Scholar] [CrossRef] [Green Version]
- Carrera-Parra, L.F.; Salazar-Vallejo, S.I. Redescriptions of Eunice filamentosa and E. denticulata and description of E. tovarae n. sp. (Polychaeta: Eunicidae), highlighted with morphological and molecular data. Zootaxa 2011, 2880, 51–64. [Google Scholar] [CrossRef]
- de Léon-González, J.A. A new eunicid polychaete from the Caribbean Coast of Mexico. Rev. Biol. Trop. 1988, 36, 75–79. [Google Scholar]
- Liu, Y.; Hutchings, P.; Sun, S. Three new species of Marphysa Quatrefages, 1865 (Polychaeta: Eunicida: Eunicidae) from the south coast of China and redescription of Marphysa sinensis Monro, 1934. Zootaxa 2017, 4263, 228–250. [Google Scholar] [PubMed]
- Liu, Y.; Hutchings, P.; Kupriyanova, E. Two new species of Marphysa Quatrefages, 1865 (Polychaeta: Eunicida: Eunicidae) from northern coast of China and redescription for Marphysa orientalis Treadwell, 1936. Zootaxa 2018, 4377, 191. [Google Scholar] [CrossRef] [PubMed]
- de León-González, J.A.; Rivera, C.G.; Romero, M.Y. Sublittoral Eunicidae and Onuphidae (Polychaeta) from softbottom of El Salvador, eastern Pacific. J. Mar. Biol. Assoc. UK 2004, 84, 93–101. [Google Scholar]
- Treadwell, A.L. Leodicidae from Fiji and Samoa. Carnegie Inst. Washingt. Publ. 1922, 312, 127–170. [Google Scholar]
- Lavesque, N.; Daffe, G.; Bonifácio, P.; Hutchings, P. A new species of the Marphysa sanguinea complex from French waters (Bay of Biscay, NE Atlantic) (Annelida, Eunicidae). Zookeys 2017, 716, 1–17. [Google Scholar] [CrossRef]
- Gathof, J.M. Family Eunicidae Savigny, 1818. In Taxonomic Guide to the Polychaetes of the Northern Gulf of Mexico. Final Report to the Minerals Management Service, Contract 14-12-001-29091. Volume VI; Uebelacker, J.M., Johnson, P.G., Eds.; U.S. Department of the Interior Mineral Management Service Gulf of Mexico OCS.: Mobile, Italy, 1984; pp. 40.1–40.31. [Google Scholar]
- de León-González, J.Á.; Díaz-Castañeda, V. Eunicidae (Annelida: Polychaeta) associated with Phragmathopoma caudata Morch, 1863 and some coral reefs from Veracruz, Gulf of Mexico. Sci. Mar. 2006, 70, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Zhang, Y.; Qiu, J. A New Species in the Marphysa sanguinea Complex (Annelida, Eunicidae) from Hong Kong. Zool. Stud. 2018, 48, 1–13. [Google Scholar] [CrossRef]
- Arias, A.; Fernández-Álvarez, F.Á.; Martins, R.; Anadón, N. Rediscovery and redescription of Leodice laurillardi (Quatrefages, 1866) comb. nov. (Annelida: Eunicidae)—A rare European polychaete or just an overlooked species? Zootaxa 2015, 3964, 475–481. [Google Scholar] [CrossRef] [Green Version]
- Hutchings, P.; Lavesque, N.; Priscilla, L.; Daffe, G.; Malathi, E.; Glasby, C.J. A new species of Marphysa (Annelida: Eunicida: Eunicidae) from India, with notes on previously described or reported species from the region. Zootaxa 2020, 4852, 285–308. [Google Scholar] [CrossRef] [PubMed]
- Veeramuthu, S.; Ramadoss, R.; Subramaniyan, B.; Jeyaram, S.; Fernando, O.J. Abundance of the Boring Polychaetes of Eunicidae (Annelida) in Great Nicobar Islands. Our Nat. 2013, 10, 76–88. [Google Scholar] [CrossRef] [Green Version]
- Miura, T. Eunicid polychaetous annelids from Japan—I. La Mer (Bulletin la Société Fr. D’océanographie) 1977, 15, 1–20. [Google Scholar]
- Miura, T. Eunicid polychaetous annelids from Japan—II. La Mer (Bulletin la Société Fr. D’océanographie) 1977, 15, 61–81. [Google Scholar]
- Miura, T. Eunicid polychaetous annelids from Japan—III. La Mer (Bulletin la Société Fr. D’océanographie) 1979, 17, 33–42. [Google Scholar]
- Miura, T. New or little known species of the Family Eunicidae (Annelida, Polychaeta) from Japan. Proc. Jpn. Soc. Syst. Zool. 1987, 36, 1–9. [Google Scholar]
- Abe, H.; Tanaka, M.; Taru, M.; Abe, S.; Nishigaki, A. Molecular evidence for the existence of five cryptic species within the Japanese species of Marphysa (Annelida: Eunicidae) known as “iwa-mushi”. Plankt. Benthos Res. 2019, 14, 303–314. [Google Scholar] [CrossRef]
- Choi, H.K.; Kim, J.G.; Kang, D.W.; Yoon, S.M. A new species of Leodice from Korean waters (Annelida, Polychaeta, Eunicidae). Zookeys 2017, 2017, 53–67. [Google Scholar] [CrossRef] [Green Version]
- Idris, I.; Hutchings, P.; Arshad, A. Description of a new species of Marphysa Quatrefages, 1865 (Polychaeta: Eunicidae) from the west coast of Peninsular Malaysia and comparisons with species from Marphysa Group A from the Indo-West Pacific and Indian Ocean. Mem. Mus. Vic. 2014, 71, 109–121. [Google Scholar] [CrossRef] [Green Version]
- Katsiaras, N.; Simboura, N.; Koutsoubas, D. The rare subgroup C1 of Marphysa (Polychaeta, Eunicidae): Re-description of species and first records in the Mediterranean Sea. Zootaxa 2014, 3873, 201–217. [Google Scholar] [CrossRef] [PubMed]
- Kurt-Sahin, G.; Çinar, M.E. Eunicidae (Polychaeta) species in and around Iskenderun Bay (Levantine Sea, Eastern Mediterranean) with a new alien species for the Mediterranean Sea and a re-description of Lysidice collaris. Turk. J. Zool. 2009, 33, 331–347. [Google Scholar] [CrossRef]
- Kurt-Sahin, G.; Çinar, M.E. Presence of Marphysa disjuncta (Polychaeta: Eunicidae) in the Mediterranean Sea. Mediterr. Mar. Sci. 2009, 10, 145–150. [Google Scholar] [CrossRef] [Green Version]
- Kurt-Sahin, G. Marphysa cinari, a new species of Eunicidae (Polychaeta) from the coasts of Turkey (eastern Mediterranean) and re-descriptions of Marphysa kinbergi McIntosh, 1910 and Marphysa disjuncta Hartman, 1961. J. Nat. Hist. 2014, 48, 1989–2006. [Google Scholar] [CrossRef]
- Kurt-Sahin, G.; Çinar, M.E. Distribution of Eunicidae (Annelida: Polychaeta) along the Levantine coast of Turkey, with special emphasis on alien species. Mar. Biodivers. 2017, 47, 421–431. [Google Scholar] [CrossRef]
- Cantone, G. Censimento dei policheti dei mari italiani: Eunicidae Berthold, 1827. Atti Soc. Toscana Sci. Nat. Resid. Pisa Mem. Ser. B 1993, 100, 229–243. [Google Scholar]
- de León-González, J.A. Eunice orensanzi n.sp. from the western coast of Baja California Sur, México and key to the Mexican Eunice (Polychaeta: Eunicidae). Rev. Biol. Trop. 1990, 38, 259–266. [Google Scholar]
- Ruiz-Cancino, G.; Carrera-Parra, L.F.; Bastida-Zavala, R. Eunícidos (Polychaeta: Eunicidae) del Pacífico sur de México. Ciencia y Mar 2010, 14, 27–60. [Google Scholar]
- Knox, G.A.; Green, K.M. The polychaetous annelids of New Zealand Part 4. Eunicidae. J. R. Soc. N. Z. 1972, 2, 459–470. [Google Scholar] [CrossRef] [Green Version]
- Glasby, C.J.; Mandario, M.A.E.; Burghardt, I.; Kupriyanova, E.; Gunton, L.M.; Hutchings, P.A. A new species of the sanguinea-group Quatrefages, 1866 (Annelida: Eunicidae: Marphysa) from the Philippines. Zootaxa 2019, 4674, 264–282. [Google Scholar] [CrossRef]
- Díaz, A.; López, E. Four New Records of Eunicidae (Annelida: Polychaeta) from the Western Philippines Islands. Thalassas 2020, 36, 321–332. [Google Scholar] [CrossRef]
- Day, J.H. A Monograph on the Polychaeta of Southern Africa. Part I. Errantia; Trustees of the British Museum (Natural History): London, UK, 1967; p. 458. [Google Scholar]
- Lewis, C.; Karageorgopoulos, P. A new species of Marphysa (Eunicidae) from the western Cape of South Africa. J. Mar. Biol. Assoc. UK 2008, 88, 277–287. [Google Scholar] [CrossRef]
- Wu, Q.Q.; He, M.H. A new genus and new species of Eunicidae from Taiwan Strait. Acta Zootaxon. Sin. 1988, 13, 123–126. [Google Scholar]
- Hsueh, P.-W.; Li, Y.-H. New species and new records of eunicids (Polychaeta, Eunicidae) from Taiwan. Zootaxa 2014, 3802, 151–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehlers, E. Reports on the results of dredging, under the direction of L. F. Pourtalès, during the years 1868-1870, and of Alexander Agassiz, in the Gulf of Mexico (1877-78), and in the Caribbean Sea (1878-79), in the U.S. Coast Survey steamer “Blake”, Lieut-Com. C. D. Sigsbee, U.S.N. and Commander J. R. Bartlett, U.S.N., commanding. XXXI. Report on the Annelids. Mem. Mus. Comp. Zool. Harv. Coll. 1887, 15, 1–335. [Google Scholar]
- Ebbs, N.K. The coral-inhabiting polychaetes of the northern Florida reef tract. Part I. Aphroditidae, Polynoidae, Amphinomidae, Eunicidae and Lysaretidae. Bull. Mar. Sci. 1966, 16, 485–555. [Google Scholar]
- Hartman, O. Polychaetous annelids from California, including the descriptions of two new genera and nine new species. Allan Hancock Pac. Exped. 1944, 10, 239–307. [Google Scholar]
- Liñero-Arana, I. Poliquetos errantes bentónicos de la platforma continental nor-oriental de Venezuela. II. Eunicidae. Bol. Inst. Oceanogr. Venez. Univ. Oriente 1990, 24, 91–103. [Google Scholar]
- Díaz-Díaz, O.F.; Bone, D.; Rodríguez, C.T.; López, A. Polychaetes associated to calcareous sediments, Venezuela: Eunicidae. Panam. J. Aquat. Sci. 2016, 11, 18–25. [Google Scholar]
- Iannotta, M.A.; Gambi, M.C.; Patti, F.P. Molecular evidence of intraspecific variability in Lysidice ninetta (Polychaeta: Eunicidae) in the Mediterranean Sea. Aquat. Biol. 2009, 6, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Schulze, A. The Bobbit worm dilemma: A case for DNA. Rev. Biol. Trop. 2011, 59, 1475–1477. [Google Scholar]
- Hutchings, P.; Kupriyanova, E. Cosmopolitan polychaetes—Fact or fiction? Personal and historical perspectives. Invertebr. Syst. 2017, 32, 1–9. [Google Scholar] [CrossRef]
- Iannotta, M.A.; Patti, F.P.; Ambrosino, M.; Procaccini, G.; Gambi, M. Phylogeography of two species of Lysidice (Polychaeta, Eunicidae) associated to the seagrass Posidonia oceanica in the Mediterranean Sea. Mar. Biol. 2007, 150, 1115–1126. [Google Scholar] [CrossRef]
- Martin, D. Anélidos poliquetos asociados a las concreciones de algas calcáreas del litoral Catalán. Miscellània Zoològica Barc. 1987, 11, 61–75. [Google Scholar]
- Lavesque, N.; Daffe, G.; Grall, J.; Zanol, J.; Gouillieux, B.; Hutchings, P. Guess who? On the importance of using appropriate name: Case study of Marphysa sanguinea (Montagu, 1813). Zookeys 2019, 859, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Pettibone, M.H. Annelida. In Synopsis and Classification of Living Organisms. 2; Parker, S.P., Ed.; McGraw-Hill Book Co.: New York, NY, USA, 1982; pp. 1–43. [Google Scholar]
- Carrera-Parra, L.F. Redescription of Hartmaniella tulearensis n. comb. (Amoureux, 1978) with comments on Hartmaniella sp and affinities of the family (Polychaeta: Hartmaniellidae). J. Nat. Hist. 2003, 37, 49–55. [Google Scholar] [CrossRef]
- Gathof, J.M. Family Hartmaniellidae Imajima, 1977. In Taxonomic Guide to the Polychaetes of the Northern Gulf of Mexico. Final Report to the Minerals Management Service, Contract 14-12-001-29091. Volume VII; Uebelacker, J.M., Johnson, P.G., Eds.; Barry A. Vittor & Associates Inc.: Mobile, AL, USA, 1984; pp. 57.1–57.3. [Google Scholar]
- Eibye-Jacobsen, D.; Oug, E. A report of Hartmaniellidae (Annelida: Polychaeta) from the Andaman Sea, Indian Ocean. Phuket Mar. Biol. Cent. Spec. Publ. 2002, 24, 101–114. [Google Scholar]
- Al-Hakim, I.; Glasby, C.J. Polychaeta (Annelida) of the Natuna Islands, South China Sea. Raffles Bull. Zool. 2004, 11, 25–45. [Google Scholar]
- Imajima, M. A new polychaete family, Hartmaniellidae, from Japan. In Essays on Polychaetous Annelids in Memory of Dr. Olga Hartman; Reish, D.J., Fauchald, K., Eds.; The Allan Hancock Foundation, University of Southern California: Los Angeles, CA, USA, 1977; pp. 211–216. [Google Scholar]
- Fauchald, K.; Rouse, G.W. Polychaete systematics: Past and present. Zool. Scr. 1997, 26, 71–138. [Google Scholar] [CrossRef]
- Szaniawski, H.; Imajima, M. Hartmaniellidae—Living fossils among polychaetes. Acta Palaeontol. Pol. 1996, 41, 111–125. [Google Scholar]
- Lu, H. Systematics of Eunicida (Polychaeta: Annelida) Homology, Ontogeny, Morphometrics, Fossils and Phylogeny. Ph.D. Thesis, George Washington University, Washington, DC, USA, 2006. [Google Scholar]
- Van Beneden, P.J. Histoire naturelle d’un animal nouveau designe sous le nom d’Histriobdella. Bull. l’Académie R. Belg. Brux. 1858, 2, 270–303. [Google Scholar]
- Foettinger, A. Recherches sur l’organisation de Histriobdella homari, P.-J. Van Beneden rapportée aux Archiannélides. Arch. Biol. (Liege) 1884, 5, 435–516. [Google Scholar]
- Shearer, C. On the anatomy of Histriobdella homari. Q. J. Microsc. Sci. 1910, 55, 17–20. [Google Scholar]
- Cordero, E.H. Un nuevo Arquianélido, Stratiodrilus platensis, n. sp., que habita sobre Aegla laevis (Latr.). Nota preliminar. Physis 1927, 8, 574–578. [Google Scholar]
- Roubaud, G. Recherches sur les Stratiodrilus platensis Cordero, Archiannelides parasites des Aegla des lacs de Patagonie. In Biologie de l’Amerique Australe II. Études sur les Faune du sol; Deboutteville, C.D., Rapport, E., Eds.; Éditions du Centre National de la Recherche Scientifique: Paris, France, 1962; pp. 31–54. [Google Scholar]
- Mesnil, F.; Caullery, M. L’appareil maxillaire d’Histriobdella homari; affinités des Hitriobdellides avec les Euniciens. Comptes Rendus l’Académie des Sci. 1922, 174, 913–917. [Google Scholar]
- Tzetlin, A.B. Ophryotrocha schubravyi sp.n. and the problem of evolution of the mouthparts in the Eunicemorpha (Polychaeta). Zool. Zhurnal 1980, 59, 666–676. (In Russian) [Google Scholar]
- Tzetlin, A.; Budaeva, N.; Vortsepneva, E.; Helm, C. New insights into the morphology and evolution of the ventral pharynx and jaws in Histriobdellidae (Eunicida, Annelida). Zool. Lett. 2020, 6, 1–19. [Google Scholar] [CrossRef]
- Gelder, S.R.; Jennings, J.B. The nervous system of the aberrant symbiotic polychaete Histriobdella homari and its implications for the taxonomic position of the Histriobdellidae. Zool. Anz. 1975, 194, 293–304. [Google Scholar]
- Jennings, J.B.; Gelder, S.R. Observations on the Feeding Mechanism, Diet and Digestive Physiology of Histriobdella homari Van Beneden 1858: An Aberrant Polychaete Symbiotic with North American and European Lobsters. Biol. Bull. 1976, 151, 489–517. [Google Scholar] [CrossRef]
- Fauchald, K. The polychaete worms. Definitions and keys to the orders, families and genera. Nat. Hist. Mus. Los Angeles Cty. Sci. Ser. 1977, 28, 1–188. [Google Scholar]
- Harrison, L. On the genus Stratiodrilus (Archiannelida: Histriobdella) with a description of a new species from Madagascar. Rec. Aust. Mus. 1928, 16, 116–121. [Google Scholar] [CrossRef] [Green Version]
- Dioni, W. Didymorchis, Temnocephala (Platyhelminthes) y Stratiodrilus (Annelida) vermes epizoicos sobre Aegla y Parastacus (Crustacea, Decapoda) de lagos Andino-Patagonicos: Notas taxonomicas y biogeograficas. Acta Zool. Lilloana 1972, 29, 167–179. [Google Scholar]
- Rouse, G.W.; Pleijel, F. Polychaetes; Oxford University Press: New York, NY, USA, 2001; p. 354. [Google Scholar]
- Glasby, C.J.; Timm, T. Global diversity of polychaetes (Polychaeta; Annelida) in freshwater. Hydrobiologia 2008, 595, 107–115. [Google Scholar] [CrossRef]
- Rosa, J.J.D.S.; Marçal, I.C.; Teixeira, G.M.; Aguiar, A. Checklist of species of Stratiodrilus Haswell, 1900 (Annelida: Histriobdellidae), and new host records from Southern Brazil. Zootaxa 2018, 4399, 412–422. [Google Scholar] [CrossRef] [PubMed]
- Steiner, T.M.; Amaral, A.C.Z. The family Histriobdellidae (Annelida, Polychaet) including descriptions of two new species from Brazil and a new genus. Contrib. Zool. 1999, 68, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.T. New substitute name for the genus Dayus Steiner & Amaral, 1999 (Annelida: Polychaeta: Histriobdellidae). Zootaxa 2014, 3802, 400. [Google Scholar]
- Amato, J.F.R.; Daudt, L.C.C.; Amato, S.B. New species of Stratiodrilus (Polychaeta, Histriobdellidae) from freshwater anomuran crustaceans of Southern Brazil. Biociências 2004, 12, 121–127. [Google Scholar]
- Lang, K. A contribution to the morphology of Stratiodrilus platensis Cordero (Histriobdellidae). Ark. Zool. 1949, 42, 1–30. [Google Scholar]
- Rouse, G.W. Histriobdellidae Vaillant, 1890. In Polychaetes; Rouse, G., Pleijel, F., Eds.; Oxford University Press: New York, NY, USA, 2001; pp. 160–163. [Google Scholar]
- Gelder, S.R.; Tyler, S. Anatomical and cytochemical studies on the adhesive organs of the ectosymbiont Histriobdella homari (Annelida: Polychaeta). Trans. Am. Microsc. Soc. 1986, 105, 348–356. [Google Scholar] [CrossRef]
- Haswell, W.A. On a new histriobdellid. Q. J. Microsc. Sci. 1900, 43, 299–335. [Google Scholar]
- Haswell, W.A. Notes on the Histriobdellidae. Q. J. Microsc. Sci. 1913, 59, 197–226. [Google Scholar]
- Daudt, L.C.C.; Amato, J.F.R. Morphological variation of Stratiodrius circensis (Polychaeta, Histriobdellidae) from a new host, Aegla leptodactyla (Crustacea, Anomura, Aeglidae) with identification of its type host species. Zootaxa 2007, 1450, 57–62. [Google Scholar] [CrossRef]
- Jamieson, B.G.M.; Afzelius, B.A.; Franzén, Å. Ultrastructure of the acentriolar, aflagellate spermatozoa and the eggs of Histriobdella homari and Stratiodrilus novaehollandiae (Histriobdellidae, Polychaeta). J. Submicrosc. Cytol. 1985, 17, 363–372. [Google Scholar]
- Simon, J.L. Incidence and behavior of Histriobdella homari (Annelida: Polychaeta) a commensal of the American lobster. Bioscience 1968, 18, 35–36. [Google Scholar]
- Boghen, A.D. A parasitological survey of the American lobster Homarus americanus from the Northumberland Strait, southern Gulf of St. Lawrence. Can. J. Zool. 1978, 56, 2460–2462. [Google Scholar] [CrossRef]
- Rouse, G.W. Polychaeta (Bristle Worms)—Chapter 5: Minor groups and fossils. In Marine Parasitology; Rohde, K., Ed.; CSIRO Publishing: Victoria, Australia, 2005; pp. 193–196. [Google Scholar]
- Martin, D.; Britayev, T.A. Symbiotic polychaetes: Review of known species. Oceanogr. Mar. Biol. Annu. Rev. 1998, 36, 217–340. [Google Scholar]
- Brattey, J.; Campbell, A. Occurrence of Histriobdella homari (Annelida: Polychaeta) on the American lobster in the Canadian Maritimes. Can. J. Zool. 1985, 63, 392–395. [Google Scholar] [CrossRef]
- Lerch, F.; Uglem, I. High density of Histriobdella homari van Beneden, 1858 (Annelida, Polychaeta) on ovigerous female european lobsters (Decapoda, Nephropidae). Crustaceana 1996, 69, 916–921. [Google Scholar] [CrossRef]
- Führ, I.M. A new histriobdellid on a marine isopod from South Africa. S. Afr. J. Sci. 1971, 67, 325–326. [Google Scholar]
- Bond-Buckup, G.; Buckup, L. A Família Aeglidae (Crustacea, Decapoda, Anomura). Arq. Zool. 1994, 32, 159–346. [Google Scholar]
- Briggs, R.P.; Atkinson, R.J.A.; McAliskey, M.; Rogerson, A. Histriobdella homari on Nephrops norvegicus from the Irish Sea and Clyde Sea area. J. Mar. Biol. Assoc. UK 1997, 77, 557–559. [Google Scholar] [CrossRef]
- Haswell, W.A. On the embryology of Stratiodrilus (Histriobdellidae). Q. J. Microsc. Sci. 1916, 61, 301–311. [Google Scholar]
- Amato, J.F.R.; Amato, S.B.; Daudt, L.C.C. The eggs of Stratiodrilus sp. (Polychaeta, Histriobdellidae). Biociências 2002, 10, 49–54. [Google Scholar]
- Vila, I.; Véliz, D. Life cycle of a freshwater Gondwanic remnant polychaete Stratiodrilus aeglaphilus (Annelida: Eunicida: Histriobdellidae), commensal with Aegla laevis (Crustacea: Anomura). Guyana 2014, 78, 120–126. [Google Scholar] [CrossRef] [Green Version]
- Cannon, L.R.G.; Jennings, J.B. Occurrence and nutritional relationships of four ectosymbiotes of the freshwater crayfishes Cherax dispar Riek and Cherax punctatus Clark (Crustacea: Decapoda) in Queensland. Aust. J. Mar. Freshw. Res. 1987, 38, 419–427. [Google Scholar] [CrossRef]
- Pardo, R.; Vila, I.; Bustamante, R. Demographic response of Stratiodrilus aeglaphilus (Annelida, Histriobdellidae) to organic enrichment: Experimental assessement. Hydrobiologia 2008, 598, 11–16. [Google Scholar] [CrossRef]
- Uzmann, J.R. Histriobdella homari (Annelida: Polychaeta) in the American lobster, Homarus americanus. J. Parasitol. 1967, 53, 210–211. [Google Scholar] [CrossRef]
- Moyano, H.I.G.; Carrasco, F.D.; Gacitua, S. Sobre las especies chilenas de Stratiodrilus Haswell, 1900, (Polychaeta, Histriobdellidae). Bol. Soc. Biol. Concepc. 1993, 64, 147–157. [Google Scholar]
- Carrera-Parra, L.F. Lumbrineridae Schmarda, 1861. In Poliquetos (Annelida: Polychaeta) de México y América Tropical; de León-González, J.Á., Bastida-Zavala, J.R., Carrera-Parra, L.F., García-Garza, M.E., Peña-Rivera, A., Salazar-Vallejo, S.I., Solís-Weiss, V., Eds.; Universidad Autónoma de Nuevo León: Monterrey, Mexico, 2009; pp. 263–275. [Google Scholar]
- Carrera-Parra, L.F. Recognition of Cenogenus Chamberlin, 1919 (Polychaeta: Lumbrineridae) based on type material. Proc. Biol. Soc. Wash. 2001, 114, 720–724. [Google Scholar]
- Orensanz, J.M. Los anelidos poliquetos de la provincia biogeografica Argentina. IV. Lumbrineridae. Physis 1973, 32, 343–393. [Google Scholar]
- Frame, A.B. The lumbrinerids (Annelida: Polychaeta) collected in two northwestern Atlantic surveys with descriptions of a new genus and two new species. Proc. Biol. Soc. Wash. 1992, 105, 185–218. [Google Scholar]
- Carrera-Parra, L.F. Phylogenetic analysis of Lumbrineridae Schmarda, 1861 (Annelida: Polychaeta). Zootaxa 2006, 1332, 1–36. [Google Scholar] [CrossRef]
- Carrera-Parra, L.F. Phylogenetic relationship of Lumbricalus Frame, 1992 (Polychaeta: Lumbrineridae), with ecological and distribution data. Mar. Ecol. 2005, 26, 166–170. [Google Scholar] [CrossRef]
- Carrera-Parra, L.F.; Orensanz, J.M. Revision of Kuwaita Mohammad, 1973 (Annelida, Polychaeta, Lumbrineridae). Zoosystema 2002, 24, 273–281. [Google Scholar]
- Carrera-Parra, L.F. Revision of Lumbricalus (Polychaeta: Lumbrineridae). J. Mar. Biol. Assoc. UK 2004, 84, 81–91. [Google Scholar] [CrossRef]
- Carrera-Parra, L.F. Revision of Lumbrineris de Blainville, 1828 (Polychaeta: Lumbrineridae). Zootaxa 2006, 1336, 1–64. [Google Scholar] [CrossRef]
- Hilbig, B. Family Lumbrineridae Malmgren, 1867, emended Orensanz, 1990. In Taxonomic Atlas of the Benthic Fauna of the Santa Maria Basin and Western Santa Barbara Channel. 5—The Annelida Part 2; Blake, J.A., Hilbig, B., Scott, P.H., Eds.; Santa Barbara Museum of Natural History: Santa Barbara, CA, USA, 1995; pp. 279–313. [Google Scholar]
- Pleijel, F. Lumbrineridae Schmarda, 1861. In Polychaetes; Rouse, G., Pleijel, F., Eds.; Oxford University Press: New York, NY, USA, 2001; pp. 164–165. [Google Scholar]
- Pruvot, G. Recherches anatomiques et morphologiques sur le système nerveux des Annélides polychètes. Arch. Zool. Expérimentale Générale 1885, 3, 211–326. [Google Scholar]
- Fauchald, K.; Jumars, P. The diet of worms: A study of polychaete feeding guilds. Oceanogr. Mar. Biol. Annu. Rev. 1979, 17, 193–284. [Google Scholar] [CrossRef]
- Valderhaug, V.A. Population structure and production of Lumbrineris fragilis (Polychaeta, Lumbrineridae) in the Oslofjord (Norway) with a note on metal content of jaws. Mar. Biol. 1985, 86, 203–211. [Google Scholar] [CrossRef]
- Carrasco, F.D.; Oyarzún, C. Diet of the polychaete Lumbrineris tetraura (Schmarda) (Lumbrineridae) in a polluted soft-bottom environmen. Bull. Mar. Sci. 1988, 42, 358–365. [Google Scholar]
- Petch, D.A. Selective Deposit-Feeding by Lumbrineris cf. latreilli (Polychaeta, Lumbrineridae), with a New Method for Assessing Selectivity by Deposit-Feeding Organisms. Mar. Biol. 1986, 93, 443–448. [Google Scholar]
- Gravier, C.; Dantan, J.L. Péchés nocturnes à la lumière dans la Baie d’Alger. Annélides polychètes. Ann. l’Institut Océanographique 1928, 5, 1–187. [Google Scholar]
- Rouse, G.W. An ultrastructural study of the spermatozoa of Eulalia sp.(Phyllodocidae), Lepidonotus sp. (Polynoidae), Lumbrineris sp.(Lumbrineridae) and Owenia fusiformis (Oweniidae). Helgoländer Wiss. Meeresunters. 1988, 42, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Messina, P.; Di Filippo, M.; Gambi, M.C.; Zupo, V. In vitro fertilisation and larval development of a population of Lumbrineris (Scoletoma) impatiens (Claparede) (Polychaeta, Lumbrineridae) of the Gulf of Naples (Italy) in relation to aquaculture. Invertebr. Reprod. Dev. 2005, 48, 31–40. [Google Scholar] [CrossRef]
- Conti, G.; Loy, F.; Diana, M.; Messina, P. The spermatozoon of Lumbrineris (Scoletoma) impatiens (Claparède, 1868) (Polychaeta, Lumbrineridae). Ital. J. Zool. 2011, 78, 215–218. [Google Scholar] [CrossRef]
- Rouse, G.W.; Jamieson, B.G.M. An ultrastructural study of the sprematozoa of the polychaetes Eurythoe complanata (Amphinomidae), Clymenella sp. and Micromaldane sp. (Maldanidae), with definition of sperm types in relation to reproductive biology. J. Submicrosc. Cytol. 1987, 19, 573–584. [Google Scholar]
- Martins, R.; Carrera-Parra, L.F.; Quintino, V.; Rodrigues, A.M. Lumbrineridae (Polychaeta) from the Portuguese continental shelf (NE Atlantic) with the description of four new species. Zootaxa 2012, 3416, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Hartman, O. New species of polychaetous annelids from southern California. With a discussion on the host of one of them. Allan Hancock Pac. Exped. 1939, 7, 157–172. [Google Scholar]
- Nishihira, M.; Tsuchiya, M.; and Sato, M. Ecological aspect of the breeding of the polychaete, Lumbrineris latreilli (Audouin et Milne-Edwards) at Asamushi, northern Japan. Bull. Mar. Biol. Stn. Asamushi 1980, 16, 201–212. [Google Scholar]
- Nishihira, M.; Tsuchiya, M.; Sato, M. Distribution and spawning site selection of the polychaete Lumbrineris latreilli (Audouin et Milne-Edwards) at Asamushi, northern Japan. Bull. Mar. Biol. Stn. Asamushi 1981, 17, 17–28. [Google Scholar]
- Nishihira, M.; Tsuchiya, M.; Sato, M. Dispersal and recruitment of juveniles of the polychaete Lumbrineris latreilli (Audouin et Milne-Edwards). Bull. Mar. Biol. Stn. Asamushi 1984, 17, 191–203. [Google Scholar]
- Carrera-Parra, L.F. Lumbrineridae (Annelida: Polychaeta) from the Grand Caribbean region with the description of six new species. J. Mar. Biol. Assoc. UK 2001, 81, 599–621. [Google Scholar] [CrossRef]
- Ayyagari, A.; Kondamudi, R. Ecological significance of the association between Stomopneustes variolaris (Echinoidea) and Lumbrineris latreilli (Polychaeta) from Visakhapatnam Coast, India. J. Mar. Biol. 2014, 2014, 640784. [Google Scholar] [CrossRef] [Green Version]
- Zibrowius, H.; Southward, E.C.; Day, J.H. New observations on a little known species of Lumbrineris (Polychaeta) living on various cnidarians, with notes on its recent and fossil scleractinian hosts. J. Mar. Biol. Assoc. UK 1975, 55, 83–108. [Google Scholar] [CrossRef]
- García-Gómez, S.C.; Carrera-Parra, L.F.; Mas, F.A.; Freitas, R.; Martins, R. Novel insights on the diversity and ecology of the family Lumbrineridae (Polychaeta) along the Iberian Peninsula coasts. J. Mar. Biol. Assoc. UK 2016, 96, 1427–1435. [Google Scholar] [CrossRef]
- Katsiaras, N.; Rousou, M.; Carrera-Parra, L.F.; García-Gómez, S.C.; Simboua, N.; Louizidou, P.; Chintiroglou, C.C.; Martins, R. Taxonomy, ecology and geographic distribution of Gallardoneris iberica (Polychaeta, Lumbrineridae) in southern Europe. J. Mar. Biol. Assoc. UK 2018, 98, 1609–1618. [Google Scholar] [CrossRef]
- Arias, A.; Carrera-Parra, L.F. First record of the genus Kuwaita (Annelida: Lumbrineridae) in Europe with the description of a new species and new ultramorphological data for the genus. Zootaxa 2014, 3887, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Levenstein, R.Y. A new genus and species of Polychaeta (Family Lumbrineridae) from the deep-water trenches the the North Pacific. In Essays on Polychaetous Annelids in Memory of Dr. Olga Hartman; Reish, D.J., Fauchald, K., Eds.; The Allan Hancock Foundation, University of Southern California: Los Angeles, CA, USA, 1977; pp. 189–198. [Google Scholar]
- Oug, E. A new small species of Lumbrineris from northern Norway and arctic waters, with comments on L. minuta (Theel, 1879) and L. vanhoeffeni (Michaelsen, 1898)—(Polychaeta: Lumbrineridae). Ophelia 1998, 49, 147–162. [Google Scholar] [CrossRef]
- Hartmann-Schröder, G. Zur Kenntnis des Eulitorals der australischen Küsten unter besonderer Berücksichtigung der Polychaeten und Ostracoden, 13. Die Polychaeten der antiborealen Küste von Victoria (Australien) (Zwischen Warrnambool im Wasten und Port Welshpool im Osten). Mitt. Hambg. Zool. Mus. Inst. 1987, 84, 27–66. [Google Scholar]
- Camargo, M.G.; da Lana, P.C. Lumbrineridae (Polychaeta: Eunicemorpha) da costa sul e sudeste do Brasil. I. Iheringia Série Zool. 1995, 79, 77–91. [Google Scholar]
- Camargo, M.G.; Lana, P. da C. Lumbrineridae (Polychaeta: Eunicemorpha) da costa sul e sudeste do Brasil. II. Lumbrineris. Iheringia 1995, 79, 93–120. [Google Scholar]
- Núñez, J.; Talavera, J.A.; Ocaña, O. Anélidos Poliquetos de Canarias: Familia Lumbrineridae. Vieraea 1991, 20, 89–100. [Google Scholar]
- Paxton, H.; Chou, L.M. Polychaetous annelids from the South China Sea. Raffles Bull. Zool. 2000, 48, 209–232. [Google Scholar]
- Cai, W.; Li, X. Eranno carrerai sp. nov. a new polychaete worm species (Polychaeta: Lumbrineridae) from coastal waters of China. Chin. J. Oceanol. Limnol. 2011, 29, 217–231. [Google Scholar] [CrossRef]
- Cai, W.; Li, X. A new species and new recorded species of Lumbrineridae Schmarda, 1861 (Anellida: Polychaeta) from China. Chin. J. Oceanol. Limnol. 2011, 29, 356–365. [Google Scholar] [CrossRef]
- Dean, K.D. Marine biodiversity of Costa Rica: Class Polychaeta (Annelida). Rev. Biol. Trop. 2004, 52, 131–181. [Google Scholar] [PubMed]
- Crossland, C. Polychaeta of tropical East Africa, the Red Sea, and Cape Verde Islands collected by Cyril Crossland, and of the Maldive Archipelago collected by Professor Stanley Gardiner, M.A., F.R.S. The Lumbriconereidae and Staurocephalidae. Proc. Zool. Soc. Lond. 1924, 1, 1–106. [Google Scholar] [CrossRef]
- Hartmann-Schröder, G. Zur Ökologie der Polychaeten des Mangrove-Estero-Gebietes von El Salvador. Beiträge zur Neotropischen Fauna 1959, 1, 69–183. [Google Scholar] [CrossRef]
- Perkins, T.H. Lumbrineridae, Arabellidae, and Dorvilleidae (Polychaeta) from Florida, with descriptions of six new species. Proc. Biol. Soc. Wash. 1979, 92, 415–465. [Google Scholar]
- Uebelacker, J.M. Family Lumbrineridae Malmgren, 1867. In Taxonomic Guide to the Polychaetes of the Northern Gulf of Mexico. Final Report to the Minerals Management Service, Contract 14-12-001-29091. Volume VI; Uebelacker, J.M., Johnson, P.G., Eds.; U.S. Department of the Interior Mineral Management Service Gulf of Mexico OCS.: Mobile, Italy, 1984; p. 41. [Google Scholar]
- Granados-Barba, A.; Solís-Weiss, V. New records of polychaetous annelids (Order: Eunicida) from the southeastern Gulf of Mexico. Bull. Mar. Sci. 1994, 52, 420–427. [Google Scholar]
- Solís-Weiss, V.; Granados-Barba, A.; Rodríguez-Villanueva, L.V.; Miranda-Vázquez, L.A.; Ochoa-Rivera, V.; Hernández-Alcántara, P. The Lumbrineridae of the continental shelf in mexican portion of the Gulf of Mexico. Mitt. Hambg. Zool. Mus. Inst. 1995, 92, 61–75. [Google Scholar]
- Fauvel, P. Annelida Polychaeta of the Indian Museum, Calcutta. Mem. Indian Mus. 1932, 12, 1–262. [Google Scholar]
- Imajima, M.; Higuchi, M. Lumbrineridae of polychaetous annelids from Japan, with descriptions of six new species. Bull. Natl. Sci. Mus. 1975, 1, 5–37. [Google Scholar]
- Imajima, M. Six species of Lumbrinerides (Polychaeta, Lumbrineridae) from Japan. Bull. Natl. Sci. Mus. 1985, 11, 171–184. [Google Scholar]
- Miura, T. Classification and morphological variations of the Japanese species of Lumbrinerides (Annelida: Lumbrineridae). Species Divers. 2017, 22, 7–27. [Google Scholar] [CrossRef] [Green Version]
- Ramos, J.M. Lumbrineridae (Polychètes errantes) de Méditterranée. Ann. l’Institut Océanograph. 1976, 52, 103–137. [Google Scholar]
- Miura, T. Lumbrineridae (Annélides Polychètes) abyssaux récoltés au cours de campagnes du Centre Océanologique de Bretagne dans l’Atlantique et la Mediterranée. Bull. Muséum d’Histoire Nat. 1980, 2, 1019–1057. [Google Scholar]
- Carrera-Parra, L.F.; Çinar, M.E.; Dagli, E. Description of a new species of Lumbrineris (Polychaeta: Lumbrineridae) from the coasts of Turkey (eastern Mediterranean). Mar. Biodivers. 2011, 41, 343–347. [Google Scholar] [CrossRef]
- D’Alessandro, M.; Cosentino, A.; Giacobbe, S.; Andaloro, F.; Romeo, T. Description of a new species of Abyssoninoe (Polychaeta: Lumbrineridae) from the NE Sicily (central Mediterranean Sea). J. Mar. Biol. Assoc. UK 2014, 94, 747–752. [Google Scholar] [CrossRef]
- Knox, G.A.; Green, K.M. The Polychaetous Annelids of New Zealand Part 2: Lumbrineridae. J. R. Soc. N. Z. 1972, 2, 69–81. [Google Scholar]
- Oug, E. Lumbrineridae from the Andaman Sea, Thailand, with notes on Oenonidae and Dorvilleidae (Annelida: Polychaeta). Phuket Mar. Biol. Cent. Spec. Publ. 2002, 24, 117–138. [Google Scholar]
- Parapar, J.; Oconnor, B.; Besteiro, C.; Urgorri, V. Abyssoninoe hibernica (McIntosh) (Polychaeta: Lumbrineridae) a valid species from the Northeast Atlantic. Sarsia 1994, 79, 157–162. [Google Scholar] [CrossRef]
- Aguirrezabalaga, F.; Carrera-Parra, L.F. Lumbrineridae (Polychaeta) from the Capbreton Canyon (Bay of Biscay, NE Atlantic) with the description of two new species. Sci. Mar. 2006, 70, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Alcántara, P.; Pérez-Mendoza, A.Y.; Solís-Weiss, V. Description of three new species of Ninoe and Cenogenus (Polychaeta: Lumbrineridae) from the Mexican Pacific. Sci. Mar. 2006, 70, 81–90. [Google Scholar]
- Fauchald, K. Deep-water errant polychaetes from Hardangerfjorden, western Norway. Sarsia 1974, 57, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Winsnes, I.M. A new species of Lumbrineris (Polychaeta) and a new subspecies of Lumbrineris scopa Fauchald from the coast of Norway. Zool. Scr. 1981, 10, 91–94. [Google Scholar] [CrossRef]
- Winsnes, I.M. Augeneria algida (Wiren) comb. n., a deep-sea lumbrinerid from Spitzbergen with aberrant setae (Annelida, Polychaeta): Redescription of holotype. Zool. Scr. 1987, 16, 39–45. [Google Scholar] [CrossRef]
- Oug, E. Guide to identification of Lumbrineridae (Polychaeta) in Norwegian and adjacent waters. Nor. Polych. Forum Guid. 2011, 1–22. [Google Scholar]
- Hartmann-Schröder, G. Zur polychaeten-fauna von Peru. Beiträge zur Neotropischen Fauna 1960, 2, 1–44. [Google Scholar] [CrossRef]
- Hartmann-Schröder, G. Die Polychaeten des Eulitorals. Zur Kenntnis des Eulitorals der chilenischen Pazifikküste und der argentinischen Küste Südpatagoniens unter besonderer Berücksichtigung der Polychaeten und Ostracoden. Mitt. Hambg. Zool. Mus. Inst. 1962, 60, 57–167. [Google Scholar]
- Rozbaczylo, N.; Vásquez-Yáñez, P.; Moreno, R.A.; Díaz-Díaz, O. Poliquetos bentónicos Amphinomida, Phyllodocida y Eunicida (Annelida: Polychaeta) de la región de fiordos y canales australes de Chile recolectados durante los cruceros CIMAR 13 al 20 fiordos. An. Inst. Patagon. 2018, 45, 51–91. [Google Scholar] [CrossRef] [Green Version]
- Rozbaczylo, N.; Moreno, R.A.; Díaz-Díaz, O. Poliquetos bentónicos submareales de fondos blandos de la región de Aysén, Chile: Clados Amphinomida, Eunicida, Spionida, Sabellida y Scolecida (Annelida, Polychaeta). Investig. Mar. 2006, 34, 43–62. [Google Scholar] [CrossRef]
- Gallardo, V.A. Polychaeta from the bay of Nha Trang, South Viet Nam. NAGA Rep. 1968, 4, 35–279. [Google Scholar]
- Fauvel, P.; Rullier, F. Annélides Polychètes. Resultats Scientifiques des Campagnes de Calypso IV. Ann. l’Institut Océanograph. 1959, 37, 143–205. [Google Scholar]
- Amoureux, L. Quelques annélides polychètes de l’Afrique occidentale et équatoriale. Cah. ORSTOM sér Océnnogr 1973, 11, 41–65. [Google Scholar]
- Intes, A.; Le Loeuff, P. Les annélides polychètes de Côte d’Ivoire. 1. Polychètes Errantes—Compte rendu systématique. Cah. ORSTOM sér Océnnogr 1975, 13, 267–321. [Google Scholar]
- Brasier, M.J.; Wiklund, H.; Neal, L.; Jeffreys, R.; Linse, K.; Ruhl, H.; Glover, A.G. DNA barcoding uncovers cryptic diversity in 50% of deep-sea antarctic polychaetes. R. Soc. Open Sci. 2016, 3, 160432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Font, T.; Gil, J.; Lloret, J. The commercialization and use of exotic baits in recreational fisheries in the north-western Mediterranean: Environmental and management implications. Aquat. Conserv. Mar. Freshw. Ecosyst. 2018, 28, 651–661. [Google Scholar] [CrossRef] [Green Version]
- Núñez, J.; Núñez, L.; Maggio, Y. Invertebrados que se Comercializan en Canarias Como cebo vivo para la Pesca Deportiva; Gobierno de Canarias: La Laguna, Spain, 2011. [Google Scholar]
- Usamentiaga, P.; Rodríguez, F.; Martin-Gil, D.; Saiz, N.; Morchon, E.; Alonso, S.; Jerez, J.; Bartolomé, B. Protein contact dermatitis by fishing bait (Lumbrinereis latreilli). Contact Dematitis 2005, 53, 236–237. [Google Scholar] [CrossRef]
- Ruggeri, G.D. Drugs from the Sea. Marine organisms with novel chemical constituents are excellent sources of new drugs. Science 1976, 194, 491–497. [Google Scholar]
- Montagu, G. Description of several marine animals found on the south coast of Devonshire. Trans. Linn. Soc. Lond. 1804, 7, 80–84. [Google Scholar] [CrossRef]
- Kinberg, J.G.H. Annulata nova. Öfversigt af Königlich Vetensk. Förhandlingar Stock 1865, 21, 559–574. [Google Scholar]
- Colbath, G.K. Revision of the family Lysaretidae, and recognition of the family Oenonidae Kinberg, 1865 (Eunicida: Polychaeta). Proc. Biol. Soc. Wash. 1989, 102, 116–123. [Google Scholar]
- Dean, H.K. A new arabellid polychaete living in the mantle cavity of deep-sea wood boring bivalves (family Pholadidae). Proc. Biol. Soc. Wash. 1992, 105, 224–232. [Google Scholar]
- Carrera-Parra, L.F. Oenonidae Kinberg, 1865. In Poliquetos (Annelida: Polychaeta) de México y América Tropical; de León-González, J.Á., Bastida-Zavala, J.R., Carrera-Parra, L.F., García-Garza, M.E., Peña-Rivera, A., Salazar-Vallejo, S.I., Solís-Weiss, V., Eds.; Universidad Autónoma de Nuevo León: Monterrey, Mexico, 2009; pp. 355–362. [Google Scholar]
- Zanol, J.; Ruta, C. New and previously known species of Oenonidae (Polychaeta: Annelida) from Lizard Island, Great Barrier Reef, Australia. Zootaxa 2015, 4019, 745–772. [Google Scholar] [CrossRef] [Green Version]
- Saito, H.; Imabayashi, H.; Kawai, K. Growth of the bivalve-feeder Halla okudai (Polychaete: Lysaretidae) under wild and rearing conditions, in relation to species and abundance of prey organisms. Fish. Sci. 1999, 65, 230–234. [Google Scholar] [CrossRef] [Green Version]
- Kawai, K.; Kunitake, H.; Saito, H.; Imabayashi, H. Paralytic and digestive activities of jelly-like substances secreted by a lysaretid polychaete, Halla okudai. Benthos Res. 1999, 54, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Iori, D.; Forti, L.; Massamba-N’Siala, G.; Prevedelli, D.; Simonini, R. Toxicity of the purple mucus of the polychaete Halla parthenopeia (Oenonidae) revealed by a battery of ecotoxicological bioassays. Sci. Mar. 2014, 78, 589–595. [Google Scholar]
- Bielig, H.J.; Möllinger, H. Über hallochrom, den epithelfarbstoff von Halla parthenopeia Costa. Physiol. Chem. 1960, 321, 276–289. [Google Scholar] [CrossRef]
- Carrera-Parra, L.F. Oenonidae Kinberg, 1865. In Poliquetos (Annelida: Polychaeta) de México y América Tropical; de León-González, J.A., Bastida-Zavala, J.R., Carrera-Parra, L.F., García-Garza, M.E., Salazar-Vallejo, S.I., Solís-Weiss, V., Tovar-Hernández, M.A., Eds.; unpublished.
- Amaral, A.C. Um poliqueto endoparasita, Labrorostratus prolificus sp. nov. em nereídeo. Bol. Inst. Oceanogr. 1977, 26, 285–292. [Google Scholar] [CrossRef]
- Abd-Elnaby, F.A. Observations on Spermatogenesis in Polychaete. Species, Halla parthenopeia (Family: Oenonidae, Polychaeta). World Appl. Sci. J. 2009, 6, 144–153. [Google Scholar]
- Pettibone, M.H. Endoparasitic polychaetous annelids of the family Arabellidae with descriptions of new species. Biol. Bull. Biol. Lab. 1957, 113, 170–187. [Google Scholar] [CrossRef]
- Pleijel, F. Oenonidae Kinberg, 1865. In Polychaetes; Rouse, G., Pleijel, F., Eds.; Oxford University Press: New York, NY, USA, 2001; pp. 166–168. [Google Scholar]
- Hilbig, B. Family Oenonidae Kinberg, 1865, emended Orensanz, 1990. In Taxonomic Atlas of the Benthic Fauna of the Santa Maria Basin and Western Santa Barbara Channel. 5—The Annelida Part 2; Blake, J.A., Hilbig, B., Scott, P.H., Eds.; Santa Barbara Museum of Natural History: Santa Barbara, CA, USA, 1995; pp. 315–339. [Google Scholar]
- Hernández-Alcántara, P.; Sólis-Weiss, V. Parasitism among polychaetes: A rare case illustrated by a new species: Labrorostratus zaragozensis, n. sp. (Oenonidae) found in the Gulf of California, Mexico. J. Parasitol. 1998, 84, 978–982. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Alcántara, P.; Cruz-Pérez, I.N.; Sólis-Weiss, V. Labrorostratus caribensis, a new oenonid polychaete from the Grand Caribbean living in the body cavity of a nereidid, with emendation of the genus. Zootaxa 2015, 4048, 127–139. [Google Scholar] [CrossRef]
- Carr, C.M. Polychaete diversity and distribution patterns in Canadian marine waters. Mar. Biodivers. 2012, 42, 93–107. [Google Scholar] [CrossRef]
- Orensanz, J.M. Los anelidos poliquetos de la provincia biogeografica Argentina. VI. Arabellidae. Physis 1974, 33, 381–408. [Google Scholar]
- Hartmann-Schröder, G. Die Polychaeten der tropischen Nordwestküste Australiens (zwischen Derby im Norden und Port Hedland im Süden). Teil 2. Zur Kenntnis des Eulittorals der australischen Küsten unter besonderer Berücksichtigung der Polychaeten und Ostracoden. Mitteilungen aus dem Hamburgischen Zoologischen Museum und Institut 1979, 76, 75–218. [Google Scholar]
- Rizzo, A.E.; Almeida dos Santos, S.; da Conceição Guerreiro Couto, E. First report of Labrorostratus caribensis (Annelida, Oenonidae) as endoparasite of Haplosyllis rosenalessoae (Annelida, Syllidae) from Brazil. Int. J. Parasitol. Parasites Wildl. 2020, 12, 64–66. [Google Scholar] [CrossRef]
- Ibarzabal, D.R. Poliquetos de Punta del Este, Isla de la Juventud, Cuba. IV Orden Eunicida. Poeyana 1989, 384, 1–28. [Google Scholar]
- Díaz-Díaz, O.; Liñero-Arana, I. Poliquetos asociados a substratos artificiales sumergidos en la costa noroerental de Venezuela 3. Eunicida (Annelida: Polychaeta). Bol. del Inst. Oceanogr. Venez. 2002, 41, 3–14. [Google Scholar]
- Dean, K.D.; Sibaja-Cordero, J.A.; Cortés, J. Polychaetes (Annelida: Polychaeta) of Cocos Island National Park, Pacific Costa Rica. Pac. Sci. 2012, 66, 347–386. [Google Scholar] [CrossRef]
- Ravara, A.; Cunha, M.R. An unusual new species of Oenonidae (Polychaeta, Eunicida) from the North Atlantic Ocean. Mar. Biodivers. 2018, 48, 1037–1043. [Google Scholar] [CrossRef]
- Uebelacker, J.M. A new parasitic polychaetous annelid (Arabellidae) from the Bahamas. J. Parasitol. 1978, 64, 151–154. [Google Scholar] [CrossRef]
- Uebelacker, J.M. Family Arabellidae Hartman, 1944. In Taxonomic Guide to the Polychaetes of the Northern Gulf of Mexico. Final Report to the Minerals Management Service, Contract 14-12-001-29091. Volume VI; Uebelacker, J.M., Johnson, P.G., Eds.; U.S. Department of the Interior Mineral Management Service Gulf of Mexico OCS.: Mobile, Italy, 1984; pp. 42.1–42.29. [Google Scholar]
- Gilbert, K. Family Lysaretidae Kinberg, 1865. In Taxonomic Guide to the Polychaetes of the Northern Gulf of Mexico. Final Report to the Minerals Management Service, Contract 14-12-001-29091. Volume VI; Uebelacker, J.M., Johnson, P.G., Eds.; U.S. Department of the Interior Mineral Management Service Gulf of Mexico OCS.: Mobile, Italy, 1984; pp. 43.1–43.7. [Google Scholar]
- Imajima, M. Errant polychaetous annelids from Tsukumo Bay and vicinity of Noto Penisula, Japan. Bull. Natl. Sci. Mus. 1967, 10, 403–441. [Google Scholar]
- Imajima, M. Polychaetous annelids of Suruga Bay, central Japan. Natl. Sci. Mus. Monogr. 1997, 12, 149–228. [Google Scholar]
- Imajima, M. Deep-sea benthic polychaetous annelids of Tosa Bay, southwestern Japan. Natl. Sci. Mus. Monogr. 2001, 20, 31–100. [Google Scholar]
- Imajima, M. Polychaetous annelids from Sagami Bay and the Sagami Sea, central Japan. Mem. Natl. Sci. Mus. 2006, 40, 317–408. [Google Scholar]
- Kobayashi, G.; Mukai, R.; Itoh, H. New record of Halla okudai Imajima, 1967 (Annelida, Eunicida, Oenonidae) from Fukue Island in the Goto Islands, Japan. Check List 2020, 16, 1199–1203. [Google Scholar]
- Ramos, J.M. Arabellidae (Polychètes Errantes) de Méditerranée. Ann. l’Institut Océanograph. 1976, 52, 247–257. [Google Scholar]
- San Martín, G.; Sardá, R. Sobre la presencia de un Arabélido (Polychaeta: Arabellidae) parásito de Sílidos (Polychaeta: Syllidae), del género Labrorostratus en las costas espanolas. Boletín la Real Soc. Española Hist. Nat. 1986, 82, 141–146. [Google Scholar]
- Cantone, G. Censimento de policheti de mari Italiani: Arabellidae Hartman, 1944, Oenonidae Kinberg, 1865. Atti Soc. Toscana Sci. Nat. Mem. Ser. B 1996, 103, 99–103. [Google Scholar]
- Mikac, B. A sea of worms: Polychaete checklist of the Adriatic Sea. Zootaxa 2015, 3943, 1–172. [Google Scholar] [CrossRef] [Green Version]
- Knox, G.A.; Green, K.M. The polychaetes of New Zealand. Part 3. Lysaretidae. J. R. Soc. N. Z. 1972, 2, 431–434. [Google Scholar] [CrossRef]
- Fauchald, K. Some Polychaetous Annelids From the Deep Basins in Sognefjorden, Western Norway. Sarsia 1972, 49, 89–106. [Google Scholar] [CrossRef]
- Hartman, O. Deep-water benthic polychaetous annelids off New England to Bermuda and other North Atlantic areas. Occas. Pap. Allan Hancock Found. 1965, 28, 1–378. [Google Scholar]
- Hartman, O.; Fauchald, K. Deep-water benthic polychaetous annelids off New England to Bermuda and other North Atlantic Areas. Part II. Allan Hancock Monogr. Mar. Biol. 1971, 6, 1–327. [Google Scholar]
- Kirkegaard, J.B. Bathyal and abyssal polychaetes (errant species). Galathea Repor 1995, 17, 7–56. [Google Scholar]
- Baeta, M.; Navarrete, A.; Ballesteros, M. First description of a new worm bait fishery in the NW Mediterranean Sea. Mediterr. Mar. Sci. 2019, 20, 302–308. [Google Scholar]
- Saito, H.; Kawai, K.; Umino, T.; Imabayashi, H. Fishing bait worm supplies in Japan in relation to their physiological traits. Mem. Mus. Vic. 2014, 71, 279–287. [Google Scholar] [CrossRef] [Green Version]
- Idris, I.; Arshad, A. Checklist of polychaetous annelids in Malaysia with redescription of two commercially exploited species. Asian J. Anim. Vet. Adv. 2013, 8, 409–436. [Google Scholar] [CrossRef] [Green Version]
- Pettibone, M.H. Polychaeta Errantia of the Siboga Expedition. Part IV. Some additional polychaetes of the Polynoidae, Hesionidae, Nereidae, Goniadidae, Eunicidae, and Onuphidae, selected as new species by the late Dr. Hermann Augener with remarks on other related species. Siboga-Exped. Monogr. 1970, 24, 199–270. [Google Scholar]
- Kucheruk, N.V. Deep-water Onuphidae (Polychaeta) from the collections of 16th cruise of R/V Dmitry Mendeleev (to the generic classification of the family Onuphidae). Tr. Institut. Okeanol. Akad. Nauk SSSR 1978, 113, 88–106. (In Russian) [Google Scholar]
- Fauchald, K. Revision of Onuphis, Nothria, and Paradiopatra (Polychaeta: Onuphidae) based upon type material. Smithson. Contrib. Zool. 1982, 356, 1–109. [Google Scholar] [CrossRef]
- Paxton, H. Generic Revision and Relationships of the Family Onuphidae (Annelida: Polychaeta). Rec. Aust. Mus. 1986, 38, 1–74. [Google Scholar] [CrossRef] [Green Version]
- Paxton, H. Fauchaldonuphis, a new genus for the enigmatic polychaete Diopatra paradoxa Quatrefages, 1866. Mar. Ecol. 2005, 26, 209–215. [Google Scholar] [CrossRef]
- Carrera-Parra, L.F. Onuphidae Kinberg, 1865. In Poliquetos (Annelida: Polychaeta) de México y América Tropical; de León-González, J.Á., Bastida-Zavala, J.R., Carrera-Parra, L.F., García-Garza, M.E., Peña-Rivera, A., Salazar-Vallejo, S.I., Solís-Weiss, V., Eds.; Universidad Autónoma de Nuevo León: Monterrey, Mexico, 2009; pp. 363–378. [Google Scholar]
- de León-González, J.A.; Cornejo-Rodríguez, M.H.; Degraer, S. A new species of Australonuphis (Polychaeta: Onuphidae) from the eastern Pacific. J. Mar. Biol. Assoc. UK 2008, 88, 739–742. [Google Scholar] [CrossRef]
- Paxton, H. Hirsutonuphis (Polychaeta: Onuphidae) from Australia, with a discussion of setal progression in juveniles. Invertebr. Taxon. 1996, 10, 77–96. [Google Scholar] [CrossRef]
- Estrella-Ruiz, A.; Hernández-Alcántara, P.; Solís-Weiss, V. Description of a new species of Hirsutonuphis (Annelida: Onuphidae) from the Mexican Pacific, with notes on the world distribution of the genus. J. Mar. Biol. Assoc. UK 2013, 93, 1777–1783. [Google Scholar] [CrossRef]
- Paxton, H. Revision of the Rhamphobrachium complex (Polychaeta: Onuphidae). Rec. Aust. Mus. 1986, 38, 75–104. [Google Scholar] [CrossRef] [Green Version]
- Paxton, H.; Budaeva, N. Minibrachium, a new subgenus of Rhamphobrachium (Annelida: Onuphidae) from Australia with the description of three new species. Zootaxa 2015, 4019, 621–634. [Google Scholar] [CrossRef] [Green Version]
- Paxton, H.; Budaeva, N. Paradiopatra (Annelida: Onuphidae) from eastern Australian waters, with the description of six new species. Zootaxa 2013, 3686, 140–164. [Google Scholar]
- Meyer, K.S.; Wagner, J.K.S.; Ball, B.; Turner, P.J.; Young, C.M.; Van Dover, C.L. Hyalinoecia artifex: Field notes on a charismatic and abundant epifaunal polychaete on the US Atlantic continental margin. Invertebr. Biol. 2016, 135, 211–224. [Google Scholar] [CrossRef]
- Nishi, E.; Kato, T. Longibrachium arariensis, a new species of Onuphidae (Annelida: Polychaeta) from the shallow water of Izu Peninsula, central Japan, with notes on its feeding behavior. Zootaxa 2009, 2081, 46–56. [Google Scholar] [CrossRef]
- Gaston, G.R.G. Benthic Polychaeta of the Middle Atlantic Bight: Feeding and distribution. Mar. Ecol. Prog. Ser. 1987, 36, 251–262. [Google Scholar] [CrossRef]
- Dayton, P.K.; Hessler, R.R. Role of biological disturbance in maintaining diversity in the deep sea. Deep Sea Res. 1972, 19, 199–208. [Google Scholar] [CrossRef]
- Lipps, J.H.; Ronan, T.E. Predation on Foraminifera by the polychete worm Diopatra. J. Foraminifer. Res. 1974, 4, 139–143. [Google Scholar] [CrossRef]
- Mangum, C.P.; Santos, S.L.; Rhodes, W.R., Jr. Distribution and feeding in the onuphid polychaete, Diopatra cuprea (Bosc). Mar. Biol. 1968, 2, 33–40. [Google Scholar] [CrossRef]
- Brenchley, G.A.; Tidball, J.G. Tube-cap orientations of Diopatra cuprea (Bosc) (Polychaeta): The compromise between physiology and foraging. Mar. Behav. Physiol. 1980, 7, 1–13. [Google Scholar] [CrossRef]
- Arias, A.; Richter, A.; Anadón, N.; Paxton, H. Evidence of simultaneous hermaphroditism in the brooding Diopatra marocensis (Annelida: Onuphidae) from northern Spain. J. Mar. Biol. Assoc. UK 2013, 93, 1533–1542. [Google Scholar] [CrossRef]
- Arias, A.; Paxton, H. The cryptogenic bait worm Diopatra biscayensis Fauchald et al., 2012 (Annelida: Onuphidae)—Revisiting its history, biology and ecology. Estuar. Coast. Shelf Sci. 2015, 163, 22–36. [Google Scholar] [CrossRef]
- Arias, A.; Paxton, H.; Budaeva, N. Redescription and biology of Diopatra neapolitana (Annelida: Onuphidae), a protandric hermaphrodite with external spermaducal papillae. Estuar. Coast. Shelf Sci. 2016, 174, 1–17. [Google Scholar] [CrossRef]
- Hsieh, H.L.; Simon, J.L. The sperm transfer system in Kinbergonuphis simoni (Polychaeta: Onuphidae). Biol. Bull. 1990, 178, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Blake, J.A. The larval development of Polychaeta from the nothern California coast. II. Nothria elegans (Family Onuphidae). Ophelia 1975, 13, 43–61. [Google Scholar] [CrossRef]
- Paxton, H. Diopatra Audouin and Milne Edwards (Polychaeta, Onuphidae) from Australia, with a discussion of developmental patterns in the genus. Beagle Rec. North Territ. Mus. Arts Sci. 1993, 10, 115–154. [Google Scholar]
- Carrasco, F.D. Description of adults and larvae of a new deep water species of Hyalinoecia (Polychaeta, Onuphidae) from the southeastern Pacific Ocean. J. Nat. Hist. 1983, 17, 87–93. [Google Scholar] [CrossRef]
- Hsieh, H.L.; Simon, J.L. Larval development of Kinbergonuphis simoni, with a summary of development patterns in the family Onuphidae (Polychaeta). Bull. Biol. Soc. Wash. 1987, 7, 194–210. [Google Scholar]
- Fadlaoui, S.A.; Lechapt, J.P.; Retière, C. Larval development of the onuphid Diopatra marocensis (Annelida: Polychaeta) from the Atlantic coast of Morocco. J. Mar. Biol. Assoc. UK 1995, 75, 957–966. [Google Scholar] [CrossRef]
- Budaeva, N.; Fauchald, K. Diopatra tuberculantennata, a new species of Onuphidae (Polychaeta) from Belize with a key to onuphids from the Caribbean Sea. Zootaxa 2008, 1795, 29–45. [Google Scholar] [CrossRef]
- Budaeva, N.; Fauchald, K. Larval development of Mooreonuphis stigmatis (Treadwell 1922) (Polychaeta: Onuphidae) from the north-east Pacific. Mar. Biol. Res. 2010, 6, 6–24. [Google Scholar] [CrossRef]
- Budaeva, N.; Pyataeva, S.; Meißner, K. Development of the deep-sea viviparous quill worm Leptoecia vivipara (Hyalinoeciinae, Onuphidae, Annelida). Invertebr. Biol. 2014, 133, 242–260. [Google Scholar] [CrossRef]
- Rangel, L.; Santos, M. Diopatra neapolitana (Polychaeta: Onuphidae) as a second intermediate host of Gymnophallus choledochus (Digenea: Gymnophallidae) in the Aveiro Estuary (Portugal): Distribution within the host and histopathology. J. Parasitol. 2009, 95, 1233–1236. [Google Scholar] [CrossRef] [Green Version]
- Rangel, L.F.; Cech, G.; Székely, C.; Santos, M.J. A new actinospore type unicapsulactinomyxon (Myxozoa), infecting the marine polychaete, Diopatra neapolitana (Polychaeta: Onuphidae) in the Aveiro Estuary (Portugal). Parasitology 2011, 138, 698–712. [Google Scholar] [CrossRef] [PubMed]
- Arias, A.; Anadón, N.; Paxton, H. New records of Diopatra marocensis (Annelida: Onuphidae) from northern Spain. Zootaxa 2010, 2691, 67–68. [Google Scholar] [CrossRef] [Green Version]
- Arias, A.; Paxton, H. Hidden diversity within the polychaete Onuphis eremita sensu lato (Annelida: Onuphidae)—Redescription of O. eremita Audouin & Milne-Edwards, 1833 and reinstatement of Onuphis pancerii Claparède, 1868. Zootaxa 2014, 3861, 145–169. [Google Scholar] [PubMed]
- Jones, C.; Lawton, J.; Shachak, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373–386. [Google Scholar] [CrossRef]
- Berke, S.K.; Mahon, A.R.; Lima, F.P.; Halanych, K.M.; Wethey, D.S.; Woodin, S.A. Range shifts and species diversity in marine ecosystem engineers: Patterns and predictions for European. Glob. Ecol. Biogeogr. 2010, 19, 223–232. [Google Scholar] [CrossRef]
- Berke, S.K. Biogeographic variability in ecosystem engineering: Patterns in the abundance and behavior of the tube-building polychaete Diopatra cuprea. Mar. Ecol. Prog. Ser. 2012, 447, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Bailey-Brock, J. Ecology of the tube-building polychaete Diopatra leuckarti Kinberg, 1865 (Onuphidae) in Hawaii: Community structure, and sediment stabilizing properties. Zool. J. Linn. Soc. Lond. 1984, 80, 191–199. [Google Scholar] [CrossRef]
- McIntosh, W.C. Report on the Annelida Polychaeta collected by H.M.S. Challenger during the years 1873–1876. Rep. Sci. Results Voyag. H.M.S.Chall. Dur. Years 1872–76 1885, 12, 1–554. [Google Scholar]
- Kucheruk, N.V. On the specific composition and distribution of the deep-sea genus Paraonuphis (sic) (Polychaeta, Eunicidae). Tr. Institut. Okeanol. Akad. Nauk SSSR 1977, 108, 44–51. (In Russian) [Google Scholar]
- Jirkov, I.A. Polychaeta of the Arctic Ocean; Yanus-K: Moscow, Russia, 2001. [Google Scholar]
- Hartman, O. Polychaetous annelids collected by the USNS Eltanin and Staten Island cruises, chiefly from Antarctic Seas. Allan Hancock Monogr. Mar. Biol. 1967, 2, 1–387. [Google Scholar]
- Fauchald, K. Onuphidae (Polychaeta) from Western Mexico. Allan Hancock Monogr. Mar. Biol. 1968, 3, 1–82. [Google Scholar]
- Fauchald, K. Polychaetes from Central American sandy beaches. Bull. South Calif. Acad. Sci. 1973, 72, 19–31. [Google Scholar]
- Fauchald, K. Onuphidae (Polychaeta) from Belize, Central America, with notes on related taxa. Biol. Soc. Wash. 1980, 93, 797–829. [Google Scholar]
- de León-González, J.A. de Soft bottom polychaetes from the western coast of Baja California Sur, México. 4. Onuphidae. Cah. Biol. Mar. 1994, 35, 57–67. [Google Scholar]
- Imajima, M. Two New Species of Nothria (Polychaeta, Onuphidae) from off Sanriku, Northeastern Honshu, Japan. Bull. Natl. Sci. Mus. 1989, 15, 1–10. [Google Scholar]
- Imajima, M. Onuphidae (Annelida, Polychaeta) from Japan, excluding the genus Onuphis. Natl. Sci. Mus. Monogr. 1999, 16, 1–115. [Google Scholar]
- Maekawa, N.; Hayashi, I. Onuphid polychaetes from Wakasa Bay, Sea of Japan. Mem. Coll. Agric. 1989, 134, 61–93. [Google Scholar]
- Maekawa, N.; Hayashi, I. Taxonomic study on the genus Onuphis (Polychaeta, Onuphidae) from Japan and adjacent seas, with descriptions of six new species. Bull. Natl. Sci. Mus. 1999, 25, 163–214. [Google Scholar]
- Paxton, H. Three new species of Aponuphis (Annelida: Onuphidae) from eastern Australia. Zootaxa 2017, 4344, 246–260. [Google Scholar] [CrossRef]
- Paxton, H. Two new species of Mooreonuphis (Annelida: Onuphidae) from Australia. Zootaxa 2018, 4486, 589–600. [Google Scholar] [CrossRef]
- Budaeva, N.; Paxton, H. Nothria and Anchinothria (Annelida: Onuphidae) from eastern Australian waters, with a discussion of ontogenetic variation of diagnostic characters. J. Mar. Biol. Assoc. UK 2013, 93, 1487–1502. [Google Scholar] [CrossRef]
- Augener, H. Polychaeta. Beiträge zur Kennt. der Meeresfauna Westafrikas 1918, 2, 67–625. [Google Scholar]
- Arias, A. Onuphis and Mooreonuphis (Annelida: Onuphidae) from West Africa with the description of three new species and the reinstatement of O. landanaensis Augener, 1918. Zootaxa 2016, 4168, 481–511. [Google Scholar] [CrossRef] [PubMed]
- Paxton, H. A new species of Diopatra (Annelida: Onuphidae) from Namibia, southwestern Africa. Mar. Biodivers. 2016, 46, 889–895. [Google Scholar] [CrossRef]
- Orensanz, J.M. Los anelidos poliquetos de la provincia biogeografica Argentina. V. Onuphidae. Physis 1974, 33, 75–122. [Google Scholar]
- Paxton, H. The Diopatra chiliensis confusion—Redescription of D. chiliensis (Polychaeta, Onuphidae) and implicated species. Zool. Scr. 1998, 27, 31–48. [Google Scholar] [CrossRef]
- Arias, A.; Paxton, H.; Anadón, N. Mooreonuphis vespa, a new brooding species of Onuphidae (Annelida) from northern Spain (Bay of Biscay). Zootaxa 2013, 3741, 583–592. [Google Scholar] [CrossRef]
- Arias, A.; Núñez, J.; Paxton, H. Onuphid polychaetes associated with the Cymodocea nodosa meadows of La Gomera (Canary Islands, NW Africa)—New species and new records from the eastern North Atlantic. J. Mar. Biol. Assoc. UK 2017, 97, 857–869. [Google Scholar] [CrossRef]
- Arias, A.; Paxton, H. Onuphis and Aponuphis (Annelida: Onuphidae) from southwestern Europe, with the description of a new species. Zootaxa 2015, 3949, 345–369. [Google Scholar] [CrossRef] [Green Version]
- Arias, A.; Paxton, H. Paradiopatra Ehlers, 1887 (Annelida: Onuphidae) from southwestern Europe with the description of a new species and new ultramorphological data for the genus. Zootaxa 2015, 4040, 149–168. [Google Scholar] [CrossRef]
- Paxton, H.; Arias, A. Unveiling a surprising diversity of the genus Diopatra Audouin & Milne Edwards, 1833 (Annelida: Onuphidae) in the Macaronesian region (eastern North Atlantic) with the description of four new species. Zootaxa 2017, 4300, 505–535. [Google Scholar]
- Kucheruk, N.V. New species of the family Onuphidae (Polychaeta) from the Pacific. Zool. Zhurnal 1979, 58, 1227–1230. (In Russian) [Google Scholar]
- Kucheruk, N.V. A new genus and species of polychaetes (Onuphidae). Zool. Zhurnal 1979, 58, 1585–1587. (In Russian) [Google Scholar]
- Paterson, G.L.J.; Glover, A.G.; Frojan, C.B.; Whitaker, A.; Budaeva, N.; Chimonides, J.; Doner, S. A census of abyssal polychaetes. Deep-Sea Res. II 2009, 56, 1739–1746. [Google Scholar] [CrossRef]
- Bergamo, G.; Carrerette, O.; Nogueira, J.M.M. Continuous and non-seasonal reproductive cycle of the alien species Diopatra neapolitana (Onuphidae, Annelida) in a tropical bay of SW Atlantic. Estuar. Coast. Shelf Sci. 2019, 231, 106479. [Google Scholar] [CrossRef]
- Elgetany, A.H.; van Rensburg, H.; Hektoen, M.; Matthee, C.; Budaeva, N.; Simon, C.A.; Struck, T.H. Species delineation in the speciation grey zone—The case of Diopatra (Annelida, Onuphidae). Zool. Scr. 2020, 49, 516–534. [Google Scholar]
- Budaeva, N.; Borisova, P.; Willassen, E. Nothria conchylega (Onuphidae, Annelida)—An example of a widespread species of polychaetes in the Northern Hemisphere. unpublished.
- Rodrigues, A.M.; Pires, A.; Mendo, S.; Quintino, V. Diopatra neapolitana and Diopatra marocensis from the Portuguese coast: Morphological and genetic comparison. Estuar. Coast. Shelf Sci. 2009, 85, 609–617. [Google Scholar] [CrossRef]
- Pires, A.; Paxton, H.; Quintino, V.; Rodrigues, A.M. Diopatra (Annelida: Onuphidae) diversity in European waters with the description of Diopatra micrura, new species. Zootaxa 2010, 2395, 17–33. [Google Scholar] [CrossRef]
- Nygren, A. Cryptic polychaete diversity: A review. Zool. Scr. 2014, 43, 172–183. [Google Scholar]
- Hektoen, M.M. (Norwegian University of Science and Technology, Trondheim, Norway). Personal communication. 2020.
- Budaeva, N.; Paxton, H.; Ribeiro, P.; Sellanes, J.; Willassen, E. Systematics and diversity of quill worms (Hyalinoeciinae, Onuphidae, Annelida). unpublished.
- Paxton, H.; Budaeva, N. Nothria (Onuphidae, Annelida) from deep Australian waters. unpublished.
- Bors, E.K.; Rowden, A.A.; Maas, E.W.; Clark, M.R.; Shank, T.M. Patterns of Deep-Sea Genetic Connectivity in the New Zealand Region: Implications for Management of Benthic Ecosystems. PLoS ONE 2012, 7, e49474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padovan, A.; Chick, R.C.; Cole, V.J.; Dutoit, L.; Hutchings, P.A.; Jack, C.; Fraser, C.I. Genomic analyses suggest strong population connectivity over large spatial scales of the commercially important baitworm, Australonuphis teres (Onuphidae). Mar. Freshw. Res. 2020, 71, 1549–1556. [Google Scholar] [CrossRef]
- Dağli, E.; Ergen, Z.; Çinar, M.E. One-year observation on the population structure of Diopatra neapolitana Delle Chiaje (Polychaeta, Onuphidae) in Izmir Bay (Aegean Sea, eastern Mediterranean). Mar. Ecol. 2005, 26, 265–272. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zanol, J.; Carrera-Parra, L.F.; Steiner, T.M.; Amaral, A.C.Z.; Wiklund, H.; Ravara, A.; Budaeva, N. The Current State of Eunicida (Annelida) Systematics and Biodiversity. Diversity 2021, 13, 74. https://0-doi-org.brum.beds.ac.uk/10.3390/d13020074
Zanol J, Carrera-Parra LF, Steiner TM, Amaral ACZ, Wiklund H, Ravara A, Budaeva N. The Current State of Eunicida (Annelida) Systematics and Biodiversity. Diversity. 2021; 13(2):74. https://0-doi-org.brum.beds.ac.uk/10.3390/d13020074
Chicago/Turabian StyleZanol, Joana, Luis F. Carrera-Parra, Tatiana Menchini Steiner, Antonia Cecilia Z. Amaral, Helena Wiklund, Ascensão Ravara, and Nataliya Budaeva. 2021. "The Current State of Eunicida (Annelida) Systematics and Biodiversity" Diversity 13, no. 2: 74. https://0-doi-org.brum.beds.ac.uk/10.3390/d13020074