
Do Weeds Hinder the Establishment of Native Plants on a Reclaimed North American Boreal Mine Site?


Department of Renewable Resources, University of Alberta, 442 Earth Sciences Building, Edmonton, AB T6G 2E3, Canada
*
Authors to whom correspondence should be addressed.
Diversity 2021, 13(2), 76; https://0-doi-org.brum.beds.ac.uk/10.3390/d13020076
Submission received: 11 December 2020
/
Revised: 3 February 2021
/
Accepted: 7 February 2021
/
Published: 12 February 2021
(This article belongs to the Special Issue Weed Ecology and Diversity)
Abstract
:The majority of plant diversity in the boreal forest of northern Alberta, Canada is comprised of native understory plant species that are continuously facing competition from other species, including both undesirable native and weedy species. In oil sands mine reclamation, cover soils rich in organic matter are used to cap overburden materials. The aim of this study is to understand the role of weeds on different reclamation cover soils (forest floor-mineral mix and peat-mineral mix) and determine if they hinder the establishment of the native plant community. This study was conducted four growing seasons after site establishment in June 2019. At that time, both soil types had approximately 45% total cover, had 21 species per plot, and were composed of mainly native vegetation. Competition from non-native forbs (11% average cover, mainly Sonchus arvensis and Melilotus alba) did not seem to impact the development of the native vegetation community on either soil type given the high cover and richness of native forbs. However, native graminoids (predominantly Calamagrostis canadensis) were associated with reduced native forb cover and richness at graminoid cover greater than 17%. Overall, non-native forbs appeared to have little impact on the native forb community on either soil type while native graminoids had a negative influence. We suggest that the classification of what is considered an undesirable weedy species should be evaluated in the context of ecosystem management goals rather than simply the presence of non-native species.
1. Introduction
Weed invasion on mine sites post-reclamation is an ongoing management issue worldwide, particularly if weeds dominate to the point of altering the desired plant community. For example, the introduced forb Tripleurospermum perforatum (scentless chamomile) is establishing and dominating on reclaimed well sites in Alberta [1,2], Andropogon gayanus (Gamba grass) is invading reclaimed mine sites in northern Australia [3], and the encroachment of a non-native shrub, Elaeagnus umbellate, is dominating coal mining reclamation sites across the eastern United States [4]. These species, and many others around the world [5,6], inhibit revegetation of desirable species post-disturbance and are considered detrimental to the reclamation goal of a native plant community. Weeds have the potential to alter the successional trajectory of a reclamation site and hinder the establishment of a native plant community [1,2,7]. Weeds typically reproduce prolifically, spread rapidly, are disturbance-adapted [8], and outcompete other plants via utilization of vital resources required for successful establishment and growth; they may also migrate outwards into the surrounding landscape [9]. Consequently, in this context of post-mining land use a “weed” is any undesirable plant species, native or introduced, that interferes with ecological, economic, and management objectives for revegetation post-disturbance and may prevent sites from meeting certification requirements [7].
“Weedy” species, including both native and non-native types in the North American boreal region, are predominantly disturbance-adapted forbs and perennial grasses capable of thriving in a variety of growing conditions [10,11]. Many of the boreal weeds may be acting as temporary placeholders in the early stage of vegetation community development and can potentially be outcompeted over time. A study done in Alberta’s oil sands region on a reclaimed upland forest suggests that within early successional communities, weedy species can act as pioneers and may be facilitating native revegetation long-term [12]. However, facilitative interactions from non-native plants on the native vegetation community are scarce and generally occur in high stress environments [13,14,15,16]. In many cases, non-native species in higher stress environments can be crucial in the recovery of optimal soil conditions which may be uninhabitable by native trees or understory species [12,16]. Through facilitation, weedy species have been shown to create better soil conditions and structure due to their ability to efficiently capture limited resources and in turn enhance aboveground and belowground biomass, leading to the addition of organic matter to the soil [12,15,16]. This positive interaction has been seen in a weedy graminoid species, Stipa tenacissim, in the degraded steppes of Spain, where it has directly enhanced seedling survival through the alteration of microclimate, soil fertility, and moisture availability [16]. However, it is not clear if competition or facilitation processes are dominating on our site.
Anthropogenic activities, particularly oil sands mining, represent a high severity disturbance that both removes the vegetation and modifies the soil [7,10,11,17]. Current reclamation operations in the oil sands mining region of northern Alberta include constructing landforms from overburden or tailings and then capping these with cover soil rich in organic matter. The reclamation cover soils used in northern Alberta are either upland-derived forest floor-mineral mix (FFMM) or peatland-derived peat-mineral mix (PMM). Due to the high stress environments created post-mining, the direction and interactions that will occur within the novel vegetation communities are relatively unknown. The interactions between weedy and native species are, in part, largely dependent on soil quality and type. FFMM often displays higher initial species diversity than PMM, resulting in more potential competitive and facilitative interactions. Cover and diversity dynamics on PMM, on the other hand, likely have a more neutral relationship, as there is initially more growing space available.
Weed research tends to be focused on the impact of non-native weedy species, but in the boreal region there is evidence that native graminoid species pose a risk to the ecosystem, tree, and plant community. However, these may not be a concern to land managers as they are native species. Further research is needed, therefore, to understand and help evaluate the differences in the plant community in response to various levels of competition from both native and non-native species on reclamation sites. The goal of this project is to gain a greater understanding of the role that weedy species, both native and non-native, play on reclaimed sites and determine if weeds hinder the establishment of the vegetation community, in particular native forbs. Alternatively, the weeds may simply be utilizing available resources with no long-term impacts on plant community development. The specific research questions include: Are weeds impeding the successful establishment of a native plant community? What are the impacts of weeds on the different soil types? Which species are acting as weeds?
2. Materials and Methods
This study was conducted on a reclamation site located at an oil sands mine approximately 75 km northwest of Fort McMurray, Alberta, Canada (57.3377° N 111.7552° W) within the boreal mixedwood ecoregion [18]. This area experiences annual precipitation of 419 mm with an annual average temperature of 1 °C [19]. Reclamation occurred in 2015 on an overburden-based tailings dyke extending over 19 ha. The reclamation site was capped with two distinct cover soils, upland-derived FFMM and peatland-derived PMM, to a depth of 0.25 m. FFMM is an upland forest-based cover soil that incorporates the seedbank rich organic forest floor layer with underlying mineral soil resulting in high levels of initial plant diversity and biomass [10,11]. PMM is derived from peatlands and is a mixture of lowland peat deposits and underlying mineral soil (ratio of 60:40 peat-to-soil volume) [17]. PMM has high organic matter content, water holding capacity, and surface roughness, making it ideal for tree and woody species establishment [20,21].
Plant community composition was measured in June 2019 at 40 (20 FFMM and 20 PMM) plots. Within each plot, four 1 m2 quadrats were averaged to assess the percent cover and richness of all vascular plants identified to species-level to a minimum of 0.5% cover [22,23,24,25]. Bare soil (%) was calculated as 100—total cover. Species were also grouped into functional groups: native forbs, non-native forbs, graminoids (grasses, rushes, and sedges), shrubs, and trees [22,23,24,25]. All graminoids, shrubs, and trees were native species. Species nomenclature follows USDA [26]. The main competitive native graminoid species on reclamation sites in Alberta is Calamagrostis canadensis [27,28,29,30]. Calamagrostis canadensis is an aggressive rhizomatous grass that grows to be 60–120 cm tall, is highly competitive for light and nutrient resources, and establishes earlier in the growing season than other plant species [23,29,30]. The large amounts of dense aboveground litter and belowground root mass produced by Calamagrostis canadensis negatively influence light availability, surface access, and soil temperature, which can prevent understory plant and tree regeneration [31,32].
General soil characteristics were measured in all plots. Volumetric soil water content (FieldScout TDR 300, Spectrum Technologies Inc., Aurora, IL, USA), soil temperature (Fisher Scientific digital thermometers), and pH (400-m, Field Scout, Spectrum Technologies Inc., Aurora, IL, USA) were measured within a short period (3–5 days) to minimize variation [33]. Soil measurements were not performed within 24 h after a rain event [33,34]. Volumetric water content (VWC) was collected from three random locations within the inner plot to acquire an average reading. Soil temperature and pH readings were collected at the plot center; the thermometer and probes were left in the ground to stabilize for 2–3 min before the reading was taken. All soil measurements were taken at a depth of 12 cm. VWC was higher by 10.5% on PMM compared to FFMM, while pH and soil temperature were similar between soil types (Table 1). All reclamation topsoil must meet chemical quality guidelines for pH, electrical conductivity, and other variables as outlined in [17].
All data analysis was done using R software (version R.3.1.1, R Core Team 2019). The main response variable of interest is native forb cover in relation to soil type and competing weed cover. Species cover, volumetric water content, and pH data had non-normal distribution based on the Shapiro–Wilk test and residual plots and could not be normalized; therefore, non-parametric tests were completed. To determine the differences between functional group cover, richness (native forbs, non-native, graminoids, shrubs, and trees), pH, VWC, and tree regeneration by soil type, a two-way permutational analysis of variance (permANOVA) (significance level of 0.05) was completed using the lmPerm package (version 2.1.0) [35]. ANOVA was used to compare soil temperature between soil types. Spearman correlation was used to compare the cover of non-native forbs and graminoids to native forb cover and richness on both soil types. Negative correlations are interpreted to indicate competition, while positive correlations indicate facilitation or no direct interaction. Regression tree analysis using the mvpart package (Version 1.6–2) [36] was used to evaluate the potential thresholds of weedy, graminoid, and individual species cover that would be potentially limiting on native forb cover.
3. Results
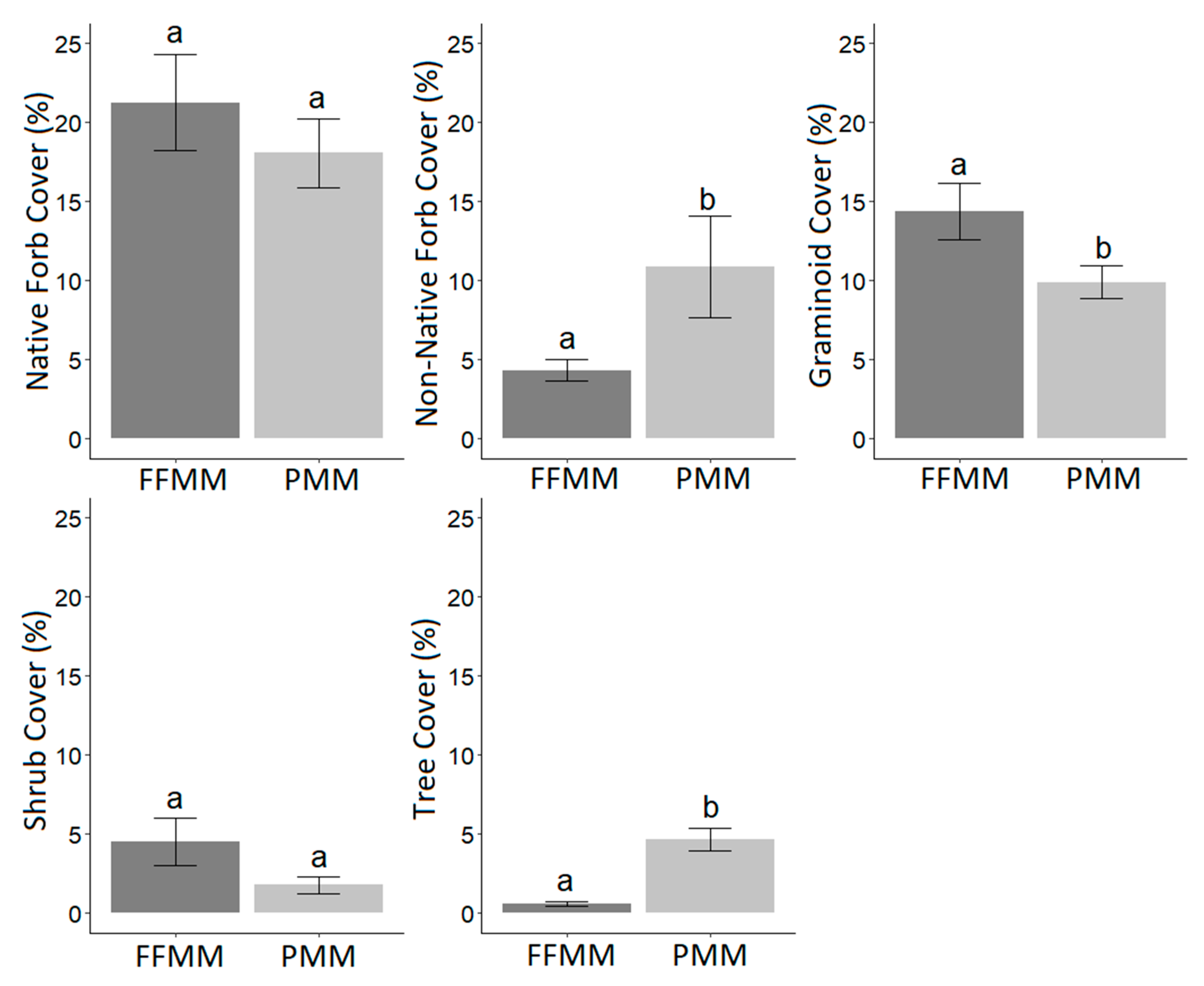
Four growing seasons after establishment, species richness (average = 21 species per 1 m2 quadrat, p = 1) and total cover (average = 45%, p = 1) were similar between FFMM and PMM. The largest component of the plant community in terms of both richness and cover was from native forbs on both soil types (Table 1, Figure 1). Native forb cover was similar on both soil types (p = 0.408) with an average cover of 21.3% on FFMM and 18.1% on PMM. Shrub cover was similar between soil types (p = 0.076) with an average cover of 4.5% on FFMM and 1.8% on PMM (Figure 1). Non-native forb cover (p < 0.001) was higher on PMM, which had a cover of 10.9%, compared to FFMM, which had an average cover of 4.3% (Figure 1). Tree cover (p < 0.001) was higher on PMM, which had an average cover of 4.7%, while FFMM had 0.6% (Figure 1). Graminoid cover was higher on the FFMM (p = 0.034), which had an average cover of 14.4%, while PMM had 9.9%.
On FFMM, the species with the highest cover were Calamagrostis canadensis (native grass), Chamerion angustifolium (native forb), Rubus idaeus (native shrub), and Sonchus arvensis (non-native forb) (Table 2). On PMM, the species with the highest cover were Chamerion angustifolium, Calamagrostis canadensis, Equisetum arvense (native forb), and Melilotus alba (non-native forb) (Table 2). Calamagrostis canadensis accounted for the largest proportion of graminoid cover on both soil types; 47% of graminoid cover was Calamagrostis on FFMM and 53% on PMM. Sonchus arvensis accounted for 60% of non-native forb cover on FFMM and 34% on PMM, while Melilotus alba accounted for 14% of non-native cover on FFMM and 40% on PMM. These three weedy species were clearly the dominant weedy species on both soil types.
On both soil types, neither native forb cover (rho = −0.307, p = 0.056) nor richness (FFMM rho = −0.131, p = 0.582; PMM rho = −0.044, p = 0.854) showed a significant relationship related to non-native forb cover. Similarly, no relationships occurred between native forb cover and the two main non-native forb species, Sonchus arvensis (FFMM rho = −0.278, p = 0.235; PMM rho = −0.223, p = 0.344) and Melilotus alba (FFMM rho = −0.149, p = 0.530; PMM rho = −0.166, p = 0.483). There was also no relationship between non-native forb cover and either tree (FFMM rho = 0.298 p = 0.202; PMM rho = −0.070 p = 0.768) or shrub (FFMM rho = 0.040 p = 0.868; PMM rho = −0.071 p = 0.767) cover.
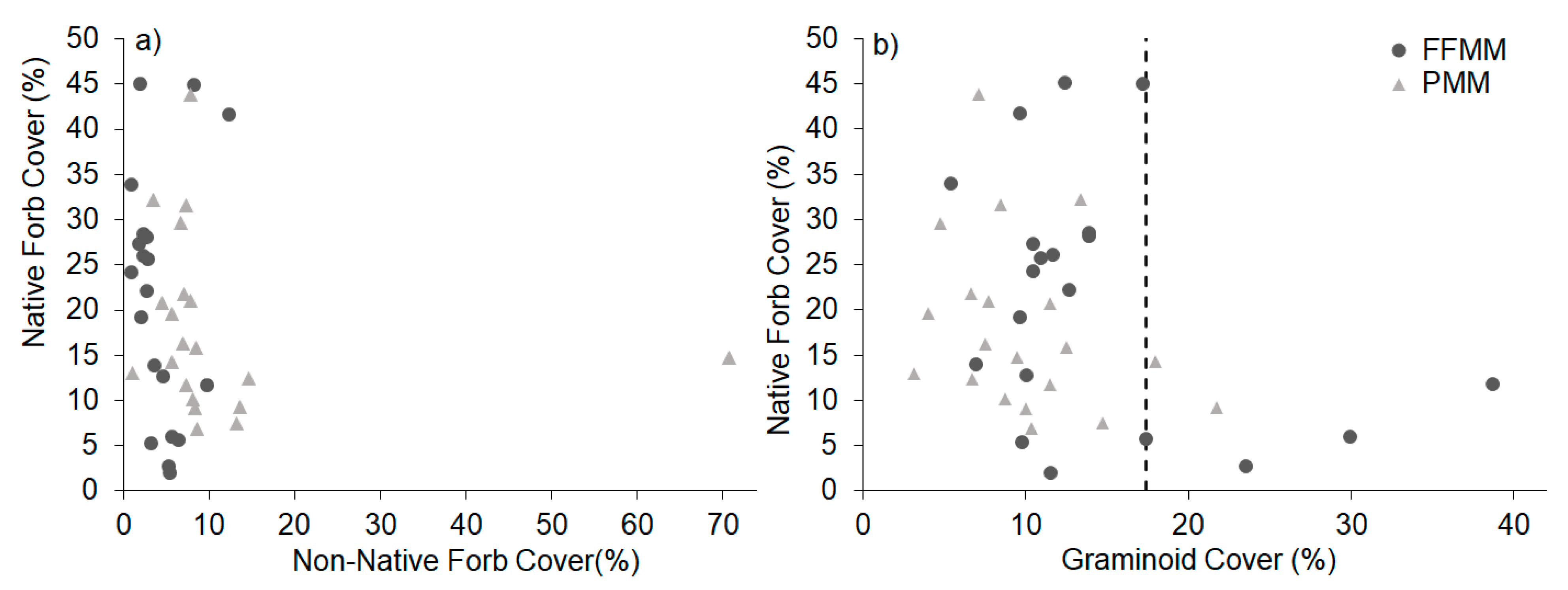
In contrast to non-native forbs, the relationship between graminoid cover and native forb cover showed a clear threshold relationship with reduced cover of native forbs when graminoid cover was greater than 17.4% (Figure 2b). Native forb richness, however, was not impacted by graminoid cover on either soil type (graminoid FFMM rho = 0.104, p = 0.663; PMM rho = 0.278, p = 0.235). There was no relationship between graminoid cover and either tree (FFMM rho = −0.043, p = 0.857; PMM rho = −0.027, p = 0.910) or shrub (FFMM rho = 0.0928, p = 0.697; PMM rho = −0.027, p = 0.513) cover on either soil type. These relationships were unaffected by soil pH, temperature, and VWC at the plot level.
4. Discussion
There is little evidence of competition among functional groups or of non-native forbs hindering the development of native forbs. On the other hand, there is also no evidence of facilitation with non-native species enhancing the establishment of native forbs such as has been hypothesized for the nitrogen-fixing Melilotus [37,38,39,40]. Based on the results of this study, Melilotus alba does not appear to be a factor in understory growth, displaying little to no impact on abundance or diversity of native species on either soil type; however, this may also be due to lifecycle, as Melilotus spp. are biennials [28,39]. Overall, it appears that non-native forbs could simply be acting as placeholders or niche fillers alongside other pioneer species within the native plant community and may not impede the establishment and development of other species [38,41]. This random vegetation community structure is common in early successional communities [39,42] and is evidenced in our study by the abundance of available seedbeds (55% bare soil on average) and growing space. Competition may play a larger role in structuring the vegetation community once all available growing space is occupied, but competition from non-native forbs does not appear to be negatively influencing the plant community at this stage.
On oil sands mine sites that utilized either FFMM or PMM as cover soil, Sonchus arvensis (Perennial sow-thistle, non-native perennial forb), Melilotus alba (White sweet clover, non-native biennial forb), and Calamagrostis canadensis (Bluejoint reedgrass, native perennial grass) have been identified as potentially having the most impact on the desired native forb community post-disturbance [1,2,27]. Listed on Alberta’s noxious weed list [20], Sonchus arvensis is 40–200 cm tall and forms large, aggressive root networks [22,43] capable of capitalizing on belowground resources, which can result in an aboveground competitive advantage deleteriously affecting the native plant community development and diversity [27]. Melilotus alba establishes quickly, grows to 250 cm tall, spreads rapidly, and acts as a nitrogen-fixer [22,28,44], making it potentially useful as a cover crop. However, some of these same characteristics enable Melilotus alba to become weedy and difficult to control [28,44]. On a reclaimed coal mine in Alberta utilizing Melilotus spp. as a cover crop, findings show the establishment of the native understory was not affected and may have been facilitated via repression of other weeds [28]. Therefore, whether or not Melilous alba should be considered a weed is dependent on many factors.
Sonchus arvensis, the other main non-native species found in our study, is listed on Alberta’s noxious weed list [45], meaning that it is legally required to be controlled to prevent the spread. However, based on our results Sonchus arvensis showed no evidence of competition with native forbs four years after site establishment and has less than 5% cover on both soil types. Species like Sonchus arvensis, and many other weeds, have high initial cover after reclamation and then decrease in abundance within a few years. This is because they often originate from the seedbank, particularly on FFMM, and have rapid expansion [28,35,46]. However, it appears that this initially high cover of Sonchus arvensis does not inhibit the growth and development of native forbs and is a low risk to the long-term ecosystem development on the site [2].
Although all graminoids, including the dominant Calamagrostis canadensis, present on our site are native species and are not considered noxious weeds, they appear to have a greater potential to reduce native forb cover [47,48]. Graminoids reduced cover of native forbs on both FFMM and PMM at graminoid cover levels greater than 17%. In other studies, the abundance of native forb species was reduced by 10% in the presence of native perennial graminoids, in particular Calamagrostis canadensis, and the growth and establishment of native trees were substantially reduced [25,43]. Graminoids have the potential to impede other native plants through a decrease in soil temperature, reduced light availability, and snow press damage due to the dense leaf litter [31,32]. A recent study from northeastern Alberta found that perennial grasses showed the greatest impact on tree growth and competition dynamics in this system [49]. We see a similar relationship in our study with higher graminoid cover and lower tree cover on FFMM, with the opposite being true on PMM.
The future of weeds on this site is still relatively unknown. On some mine reclamation sites, non-native forbs can still be a large problem, showing 95% cover more than 20 years post reclamation [50]. In other studies, weeds average only 10% cover after 20 years [21]. In our study, non-native forbs were around 10% cover after four years. Non-native species still have the potential to drive ecosystem development when native species richness is low and non-native forb cover is high [21]. In other cases, such as in our study, when non-native cover is low and native species richness is high, non-native forbs are not likely to have a large impact in structuring the vegetation community [51].
The future role of non-native forbs is generally dependent on the development of an overstory canopy [51,52,53]. As long as trees can develop into the initial community on reclamation sites, the long-term competitive relationships will likely be similar to what is found in nearby natural ecosystems [46,48]. In both mature natural stands and post-disturbance stands, the tree canopy has a strong influence on the altering of the understory plant community, on soil nutrients, and on light availability [46,48]. With the presence of an overstory canopy, the understory will be less likely to be invaded by weedy species due to the barrier for wind-dispersal and heavy shading [50,52]. However, as in natural forests, graminoids are capable of remaining in the understory canopy throughout the entire rotation [29].
We expected the weed relationships to vary by soil type, but we did not see this trend. This indicates that the vegetation communities on both soil types are behaving in a similar manner. The initial vegetation community on this site was different between soil types due to the differences in the seedbank [46]. Within a few years, however, other factors such as seed rain alter the vegetation community on both soil types [46,53]. It also appears that even at this early successional stage, the competitive processes between plant functional groups are similar between soil types; in the future, we expect the successional patterns to be similar as well.
5. Management Implications
From a management perspective, if weeds are not posing a problem by reducing desirable plant cover or altering successional pathways, they can likely be monitored rather than eradicated, resulting in saved time and money on weed control. Instead of focusing on the presence and subsequent control of a particular weed species, reclamation requirements and policies may need to focus more on ecosystem health and productivity outcomes, such as the cover and richness of a desirable native plant community. This approach will better account for “novel” ecosystems, which are a more realistic and economically achievable outcome [51]. A viable weed management option for our site appears to be simply allowing the desirable plant community to develop over time with no further interventions required. However, on sites where native graminoids appear to be acting as a more serious weedy competitor than non-native forbs, more active interventions may be needed.
Sonchus arvensis, Melilotus alba, and Calamagrostis canadensis on this reclamation site established through the seedbank. Determining the pre-salvage abundance of these species would allow managers to identify the probability of reclamation sites having higher weed pressures and potentially prevent them from establishing on site at all [2,29]. Although competition did not seem to have a large influence in structuring the vegetation community in our study, caution must be used to extrapolate to other reclamation sites and successional stages. This study was a short-term experiment during the summer season of 2019 and was only one time period on a successional stage. In addition, our study was intended to identify the actual impact of weeds on native forbs on a specific site, and we believe we have accomplished this goal using relatively simple plant community metrics, specifically cover and richness. However, future studies using more sophisticated metrics such as plant volume and competition indices may be able to better quantify plant interactions [54]. Of course, all reclamation sites are unique; therefore, continued monitoring is needed to gain a better understanding of the long-term implications of different weed management techniques on different site types.
Author Contributions
Conceptualization, B.B., B.D.P., and K.E.T.; methodology, B.B., B.D.P., and K.E.T.; validation, B.B., B.D.P., and K.E.T.; formal analysis, K.E.T.; investigation, B.B., B.D.P., and K.E.T.; data curation, B.B., B.D.P., and K.E.T.; writing—original draft preparation, K.E.T., B.B., and B.D.P.; writing—review and editing, K.E.T., B.B., and B.D.P.; visualization, K.E.T. and B.D.P.; supervision, B.D.P. and K.E.T.; funding acquisition, B.D.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Canadian Natural Resources Limited.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to privacy concerns.
Acknowledgments
We would like to thank Sofia Toledo and Mathew Anderson for help with lab and field work. Thank you, Sebastian Dietrich for the comments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cole, D.E.; King, J.R.; Oyarzun, D.A.; Dietzler, T.H.; McClay, A. Experiences with invasive plant management and ecology in Alberta. Can. J. Plant Sci. 2007, 87, 1013–1022. [Google Scholar] [CrossRef] [Green Version]
- Small, C.; Degenhardt, D.; Drozdowski, B.; Thacker, S.; Powter, C.; Schoonmaker, A.; Schreiber, S. Optimizing Weed Control for Progressive Reclamation: Literature Review; Canada’s Oil Sands Innovation Alliance: Calgary, AB, Canada, 2018; 48p. [Google Scholar]
- Luck, L.; Bellairs, S.M.; Rossiter-Rachor, N.A. Residual herbicide treatments reduce Andropogon gayanus (Gamba Grass) recruitment for mine site restoration in northern Australia. Ecol. Manag. Restor. 2019, 20, 214–221. [Google Scholar] [CrossRef] [Green Version]
- Franke, M.E.; Zipper, C.; Barney, J.N. Native hardwood tree seedling establishment following invasive autumn-olive (elaeagnus umbellata) removal on a reclaimed coal mine. Invasive Plant Sci. Manag. 2018, 11, 155–161. [Google Scholar] [CrossRef]
- Holmes, P.M. Shrubland restoration following woody alien invasion and mining: Effects of topsoil depth, seed source, and fertilizer addition. Restor. Ecol. 2001, 9, 71–84. [Google Scholar] [CrossRef]
- González-Muñoz, N.; Costa-Tenorio, M.; Espigares, T. Invasion of alien Acacia dealbata on Spanish Quercus robur forests: Impact on soils and vegetation. For. Ecol. Manag. 2012, 269, 214–221. [Google Scholar] [CrossRef]
- Alberta Environment. Guidelines for Reclamation to Forest Vegetation in the Athabasca Oil Sands Region, 2nd ed.; Terrestrial Subgroup of the Reclamation Working Group of the Cumulative Environmental Management Association: Fort McMurray, AB, Canada, 2010. [Google Scholar]
- Baker, H.G. The evolution of weeds. Annu. Rev. Ecol. Syst. 1974, 5, 1–24. [Google Scholar] [CrossRef]
- Espeland, E.; Perkins, L. Weed establishment and persistence after water pipeline installation and reclamation in the mixed grass prairie of Western North Dakota. Ecol. Restor. 2017, 35, 303–310. [Google Scholar] [CrossRef]
- Errington, R.C.; Pinno, B.D. Early successional plant community dynamics on a reclaimed oil sands mine in comparison with natural boreal forest communities. Ecoscience 2015, 22, 133–144. [Google Scholar] [CrossRef]
- Mackenzie, D.D.; Naeth, M.A. The role of the forest soil propagule bank in assisted natural recovery after oil sands mining. Restor. Ecol. 2009, 18, 418–427. [Google Scholar] [CrossRef]
- Gingras-Hill, T.; Nwaishi, F.C.; Macrae, M.; Price, J.S.; Petrone, R.M. Ecohydrological functioning of an upland undergoing reclamation on post-mining landscape of the Athabasca oil sands region, Canada. Ecohydrology 2018, 11, 1–12. [Google Scholar] [CrossRef]
- Cavieres, L.; Arroyo, M.T.K.; Peñaloza, A.; Molina-Montenegro, M.; Torres, C. Nurse effect of Bolax gummifera cushion plants in the alpine vegetation of the Chilean Patagonian Andes. J. Veg. Sci. 2002, 13, 547–554. [Google Scholar] [CrossRef]
- Bruno, J.F.; Stachowicz, J.J.; Bertness, M.D. Inclusion of facilitation into ecological theory. Trends Ecol. Evol. 2003, 18, 119–125. [Google Scholar] [CrossRef]
- Jordan, N.; Larson, D.L.; Huerd, S.C. Soil modification by invasive plants: Effects on native and invasive species of mixed-grass prairies. Biol. Invasions 2007, 10, 177–190. [Google Scholar] [CrossRef]
- Maestre, F.T.; Bautista, S.; Cortina, J.; Bellot, J. Potential for using facilitation by grasses to establish shrubs on a semiarid degraded steppe. Ecol. Appl. 2001, 11, 1641–1655. [Google Scholar] [CrossRef]
- Alberta Environment and Water. Best Management Practices for Conservation of Reclamation Materials in the Mineable Oil Sands Region of Alberta; Province of Alberta: Edmonton, AB, Canada, 2012; pp. 1–161. [Google Scholar]
- Natural Regions Committee. Natural Regions and Subregions of Alberta; Natural Regions Committee: Edmonton, AB, Canada, 2006.
- Canadian Climate Normals 1981–2010 Station Data. Available online: https://climate.weather.gc.ca/climate_normals/results_1981_2010_e.html?searchType=stnProv&lstProvince=AB&txtCentralLatMin=0&txtCentralLatSec=0&txtCentralLongMin=0&txtCentralLongSec=0&stnID=2704&dispBack=0 (accessed on 22 April 2020).
- Pinno, B.D.; Errington, R.C. Maximizing natural trembling aspen seedling establishment on a reclaimed boreal oil sands site. Ecol. Restor. 2015, 33, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Pinno, B.D.; Hawkes, V.C. Temporal trends of ecosystem development on different site types in reclaimed boreal forests. Forests 2015, 6, 2109–2124. [Google Scholar] [CrossRef] [Green Version]
- Tannas, K.E. Common Plants of the Western Rangelands—Volume 3: Forbs, 1st ed.; Alberta Agriculture, Food and Rural Development: Lethbridge, AB, Canada, 2004. [Google Scholar]
- Tannas, K.E. Common Plants of the Western Rangelands—Volume 1: Grasses and Grass-Like Species, 1st ed.; Alberta Agriculture, Food and Rural Development: Lethbridge, AB, Canada, 2001. [Google Scholar]
- Moss, E.H.; Packer, J. Flora of Alberta, 2nd ed.; University of Toronto Press: Toronto, ON, Canada, 1994. [Google Scholar]
- Tannas, K.E. Common Plants of the Western Rangelands—Volume 2: Trees and Shrubs, 1st ed.; Alberta Agriculture, Food and Rural Development: Lethbridge, AB, Canada, 2003. [Google Scholar]
- United States Department of Agriculture. Available online: https://plants.sc.egov.usda.gov/adv_search.html (accessed on 7 December 2019).
- de Bortoli, L.A.; Pinno, B.D.; Mackenzie, M.D.; Li, E.H. Plant community composition and tree seedling establishment in response to seeding and weeding treatments on different reclamation cover soils. Can. J. For. Res. 2019, 49, 836–843. [Google Scholar] [CrossRef] [Green Version]
- Macdonald, S.E.; Snively, A.E.K.; Fair, J.M.; Landhausser, S.M. Early trajectories of forest understory development on reclamation sites: Influence of forest floor placement and a cover crop. Restor. Ecol. 2015, 23, 698–706. [Google Scholar] [CrossRef]
- Lieffers, V.J.; Macdonald, S.E.; Hogg, E.H. Ecology of and control strategies for Calamagrostis canadensis in boreal forest sites. Can. J. For. Res. 1993, 23, 2070–2077. [Google Scholar] [CrossRef]
- Macdonald, S.E.; Lieffers, V.J. Rhizome plasticity and clonal foraging of Calamagrostis canadensis in response to habitat heterogeneity. J. Ecol. 1993, 81, 769–776. [Google Scholar] [CrossRef]
- Landhäusser, S.M.; Mulak, T.L.; Lieffers, V.J. The effect of roots and litter of Calamagrostis canadensis on root sucker regeneration of Populus tremuloides. Forestry 2007, 80, 481–488. [Google Scholar] [CrossRef]
- Landhausser, S.M.; Stadt, K.J.; Lieffers, V.J. Screening for control of a forest weed: Early competition between three replacement species and Calamagrostis canadensis of Picea glauca. J. Appl. Ecol. 1996, 33, 1517. [Google Scholar] [CrossRef]
- Macdonald, S.E.; Fenniak, T.E. Understory plant communities of boreal mixedwood forests in western Canada: Natural patterns and response to variable-retention harvesting. For. Ecol. Manag. 2007, 242, 34–48. [Google Scholar] [CrossRef]
- Pinno, B.D.; Sherr, I.; Errington, R.C.; Shea, K. Islands – Soil Patches and Plant Community Dynamics on a New Oil Sands Reclamation Design. J. Am. Soc. Min. Reclam. 2016, 5, 28–44. [Google Scholar] [CrossRef]
- Package ‘lmperm’. Available online: https://cran.r-project.org/web/packages/lmPerm/lmPerm.pdf (accessed on 7 December 2019).
- Package ‘mvpart’. Available online: https://mran.microsoft.com/snapshot/2014-12-11/web/packages/mvpart/mvpart.pdf (accessed on 7 December 2019).
- Foster, B.L.; Tilman, D. Dynamic and static views of succession: Testing the descriptive power of the chronosequence approach. Plant Ecol. 2000, 146, 1–10. [Google Scholar] [CrossRef]
- Alday, J.G.; Pallavicini, Y.; Marrs, R.H.; Martínez-Ruiz, C. Functional groups and dispersal strategies as guides for predicting vegetation dynamics on reclaimed mines. Plant Ecol. 2011, 212, 1759–1775. [Google Scholar] [CrossRef]
- Spellman, K.V.; Schneller, L.C.; Mulder, C.P.H.; Carlson, M.L. Effects of non-native Melilotus albus on pollination and reproduction in two boreal shrubs. Oecologia 2015, 179, 495–507. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Wedin, D. Dynamics of nitrogen competition between successional grasses. Ecology 1991, 72, 1038–1049. [Google Scholar] [CrossRef]
- MacDougall, A.S.; Turkington, R. Are invasive species the drivers or passengers of change in degraded ecosystems? Ecology 2005, 86, 42–55. [Google Scholar] [CrossRef] [Green Version]
- Dhar, A.; Comeau, P.G.; Karst, J.; Pinno, B.D.; Chang, S.X.; Naeth, A.M.; Vassov, R.; Bampfylde, C. Plant community development following reclamation of oil sands mine sites in the boreal forest: A review. Environ. Rev. 2018, 298, 1–13. [Google Scholar] [CrossRef]
- Plants Database: Sonchus arvensis, L. Field Sowthistle. Available online: https://plants.usda.gov/core/profile?symbol=SOAR2 (accessed on 7 December 2019).
- Turkington, R.A.; Cavers, P.B.; Rempel, E. The biology of Canadian weeds. 29. Melilotus alba Desr. and M. offtcinalis (L.) Lam. ROY. Can. J. Planr Sci. 1978, 58, 523–537. [Google Scholar] [CrossRef]
- Provincially Regulated Weeds. Available online: https://www.alberta.ca/provincially-regulated-weeds.aspx (accessed on 7 December 2019).
- Trepanier, K.E.; Pinno, B.D.; Errington, R.C. Dominant drivers of plant community assembly vary by soil type and time in reclaimed forests. Plant Ecol. 2020. [Google Scholar] [CrossRef]
- Brown, C.S.; Bugg, R.L. Effects of established perennial grasses on introduction of native forbs in California. Restor. Ecol. 2001, 9, 38–48. [Google Scholar] [CrossRef]
- Landhäusser, S.M.; Lieffers, V.J. Growth of Populus tremuloides in association with Calamagrostis canadensis. Can. J. For. Res. 1998, 28, 396–401. [Google Scholar] [CrossRef]
- Henkel-Johnson, D.; Macdonald, S.E.; Bork, E.W.; Thomas, B.R. Influence of weed composition, abundance, and spatial proximity on growth in young hybrid poplar plantations. For. Ecol. Manag. 2016, 362, 55–68. [Google Scholar] [CrossRef]
- Strong, W.L. Vegetation development on reclaimed lands in the Coal Valley Mine of western Alberta, Canada. Can. J. Bot. 2000, 78, 110–118. [Google Scholar] [CrossRef]
- Audet, P.; Pinno, B.D.; Thiffault, E. Reclamation of boreal forest after oil sands mining: Anticipating novel challenges in novel environments. Can. J. For. Res. 2015, 45, 364–371. [Google Scholar] [CrossRef] [Green Version]
- Hart, S.A.; Chen, H.Y.H. Understory vegetation dynamics of North American boreal forests. CRC. Crit. Rev. Plant Sci. 2006, 25, 381–397. [Google Scholar] [CrossRef]
- Cole, R.J.; Holl, K.D.; Zahawi, R.A. Seed rain under tree islands planted to restore degraded lands in a tropical agricultural landscape. Ecol. Appl. 2010, 20, 1255–1269. [Google Scholar] [CrossRef] [PubMed]
- Weigelt, A.; Jolliffe, P. Indices of plant competition. J. Ecol. 2003, 91, 707–720. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Average percent cover by functional group comparison of forest floor-mineral mix (FFMM) and peat-mineral mix (PMM) soil types with standard error bars and significant letters based on permutational analysis of variance (permANOVA) (FFMM n = 20; PMM n = 20).
Figure 1.
Average percent cover by functional group comparison of forest floor-mineral mix (FFMM) and peat-mineral mix (PMM) soil types with standard error bars and significant letters based on permutational analysis of variance (permANOVA) (FFMM n = 20; PMM n = 20).

Figure 2.
Scatter plot between native forb cover (%), (a) non-native forbs, and (b) graminoid cover (%) with a threshold created from a regression tree analysis (threshold for graminoid cover ≥17.4% n = 6, <17.4% n = 34).
Figure 2.
Scatter plot between native forb cover (%), (a) non-native forbs, and (b) graminoid cover (%) with a threshold created from a regression tree analysis (threshold for graminoid cover ≥17.4% n = 6, <17.4% n = 34).


{kind=link}
{kind=link}
Table 1.
Study site characteristics for showing average soil temperature, VWC, pH, (standard deviation), and species richness for both soil types (FFMM n = 20; PMM n = 20). p-values for soil temperature are based on ANOVA and all other p-values are based on permANOVA.
Table 1.
Study site characteristics for showing average soil temperature, VWC, pH, (standard deviation), and species richness for both soil types (FFMM n = 20; PMM n = 20). p-values for soil temperature are based on ANOVA and all other p-values are based on permANOVA.
| FFMM | PMM | p | ||
|---|---|---|---|---|
| Soil | Soil Temperature (°C) | 14.1 (2.2) | 15.4 (1.4) | 0.090 |
| VWC (%) | 16.5 (4.4) | 27.0 (6.4) | <0.001 | |
| pH | 6.9 (0.7) | 6.5 (0.9) | 0.176 | |
| Average Species Richness Per 1 m2 Quadrat | Tree | 1 | 2 | <0.001 |
| Native Forb | 10 | 9 | 0.118 | |
| Non-Native Forb | 3 | 4 | 0.270 | |
| Graminoid | 5 | 5 | 1 | |
| Shrub | 2 | 1 | 0.980 |
Table 2.
Four species with the highest average percent cover per plot found within each functional group and soil type showing cover (%).
Table 2.
Four species with the highest average percent cover per plot found within each functional group and soil type showing cover (%).
| FFMM | PMM | |||
|---|---|---|---|---|
| Functional Group | Species | Average Cover Per Plot (%) | Species | Average Cover Per Plot (%) |
| Non-Native Forbs | Sonchus arvensis | 2.6 | Melilotus alba | 4.4 |
| Melilotus alba | <1.0 | Sonchus arvensis | 3.7 | |
| Taraxacum officinale | <1.0 | Crepis tectorum | 1.5 | |
| Crepis tectorum | <1.0 | Taraxacum officinale | <1.0 | |
| Native Forbs | Chamerion angustifolium | 6.2 | Chamerion angustifolium | 6.5 |
| Equisetum arvense | 3.9 | Equisetum arvense | 5.8 | |
| Lathyrus ochroleucus | 1.9 | Achillea millefolium | 1.2 | |
| Fragaria virginiana | 1.7 | Lathyrus ochroleucus | <1.0 | |
| Graminoids | Calamagrostis canadensis | 6.8 | Calamagrostis canadensis | 5.3 |
| Poa pratensis | 2.7 | Agropyron trachycaulum | 1.0 | |
| Agropyron trachycaulum | 2.5 | Carex spp. | <1.0 | |
| Leymus innovatus | 1.3 | Poa pratensis | <1.0 | |
| Shrubs | Rubus idaeus | 3.7 | Salix spp. | 1.0 |
| Rosa acicularis | <1.0 | Rubus idaeus | <1.0 | |
| Cornus sericea | <1.0 | Rosa acicularis | <1.0 | |
| Ribes oxyacanthoides | <1.0 | Shepherdia canadensis | <1.0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Trepanier, K.E.; Burton, B.; Pinno, B.D. Do Weeds Hinder the Establishment of Native Plants on a Reclaimed North American Boreal Mine Site? Diversity 2021, 13, 76. https://0-doi-org.brum.beds.ac.uk/10.3390/d13020076
AMA Style
Trepanier KE, Burton B, Pinno BD. Do Weeds Hinder the Establishment of Native Plants on a Reclaimed North American Boreal Mine Site? Diversity. 2021; 13(2):76. https://0-doi-org.brum.beds.ac.uk/10.3390/d13020076
Chicago/Turabian StyleTrepanier, Kaitlyn E., Brea Burton, and Bradley D. Pinno. 2021. "Do Weeds Hinder the Establishment of Native Plants on a Reclaimed North American Boreal Mine Site?" Diversity 13, no. 2: 76. https://0-doi-org.brum.beds.ac.uk/10.3390/d13020076
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.