
Orchid Extinction over the Last 150 Years in the Czech Republic


1
Global Change Research Institute, Academy of Sciences of the Czech Republic, Bělidla 986/4a, 60300 Brno, Czech Republic
2
Institute for Environmental Studies, Faculty of Science, Charles University, Benátská 2, 12801 Prague, Czech Republic
*
Author to whom correspondence should be addressed.
Diversity 2021, 13(2), 78; https://0-doi-org.brum.beds.ac.uk/10.3390/d13020078
Submission received: 15 January 2021
/
Revised: 5 February 2021
/
Accepted: 9 February 2021
/
Published: 13 February 2021
(This article belongs to the Special Issue The Ecology and Diversity of Orchids)
Abstract
:Understanding temporal changes in the distribution and abundance of various species is one of the key goals of conservation biology. During recent decades, the abundance and distribution of many species of plants and animals have declined dramatically, mainly because of habitat loss and fragmentation. The purpose of this study is to analyze the rate of extinction of orchids at various sites in different 20-year time intervals over the last 150 years, determined according to changes in society. Using the dataset of the orchid records of the Nature Conservation Agency of the Czech Republic, we determined the disappearance rate of orchids from sites using a grid of 1 × 1 km. We found that the vast majority of orchids disappeared from many of their historical localities in all time intervals analyzed. The number of sites suitable for Czech orchids declined by 8–92%, depending on the species. The most threatened orchid species in the Czech Republic are Spiranthes spiralis, Anacamptis palustris, Epipogium aphyllum and Goodyera repens. This all seems to be closely related with changes in agricultural practices in the open as well as in forest habitats. Preserving suitable orchid habitats seems to be the key for keeping Czech orchid flora alive.
1. Introduction
One of the most important issues of modern conservation biology is to understand temporal changes in the distribution and abundance of various species [1]. During recent decades, the human impact on natural resources and habitats has caused a dramatically large extinction rate and a decline in the number of sites suitable for many plants and animals [2,3,4,5,6,7], with habitat loss and fragmentation being the most important factors [2,3]. Mankind has had a very marked effect on Europe, especially its rapid industrial development in recent centuries, which has resulted in a highly fragmented and degraded landscape [8]. The current land cover in Europe is mainly a result of farming and demographic trends over recent decades [9]. In general, urbanization, changes in land use and the intensification of agriculture are the most important factors that have resulted in habitat destruction in many parts of Europe [5,6,10,11,12].
In the past, there were important changes in the use of land in the Czech Republic (Czechoslovakia, until the splitting of the country in 1993), which differed from those that occurred in Western Europe because of the differences in the political regimes in Central and Western Europe [13,14]. Before 1948, fields and meadows were traditionally managed [15], which involved mowing and grazing, low intensity agriculture of small fields and low application of fertilizers [14]. After 1948, small fields were combined into huge fields [16] and subsidies for fertilizers were provided, which resulted in the amount of chemicals in the soil increasing rapidly [14]. After the change of regime in 1989, the subsidies for fertilizers stopped, which resulted in a great decline in the use of fertilizers [17].
In this study, we concentrate on orchids, because orchids are considered to be one of the most threatened groups of species worldwide [18,19,20]. With approximately 28,500 species [21], Orchidaceae is one of the most diverse and widespread families of flowering plant [18]. Unfortunately, most species of orchids are threatened in the wild [22] and are disappearing from their natural habitats worldwide [7,22,23,24]. In Europe, the main reasons for this dramatic decline are habitat loss, eutrophication and fragmentation [12,23,25,26,27,28]. Their typical low pollination success and low levels of fruit set, as well as environmental heterogeneity, are also linked to their rarity [29,30,31]. Because of the activities of professionals and amateurs, orchids are well recorded and studied in many countries in Europe [32]. The availability of detailed records collected over long periods of time provides opportunities for comparative analyses of the declines of a species over time [23]. Identification of environmental factors and species traits that are correlated with the decline in the orchid’s distribution could result in improvements in the management of valuable habitats [23]. However, despite the increasing number of studies dealing with orchid distribution in different parts of the world e.g., [33,34,35,36,37,38], there is scarce information available on orchid conservation and the drivers of their extinction [31,39].
An important category of orchids that requires special conservation measures is those orchids classified in the most threatened category according either to the IUCN guidelines or other national classifications e.g., [8,40,41]. These lists of species provide essential information on trends in the loss of biodiversity [42,43,44]. Red List assessments often differ among countries [45] and are therefore more appropriate for local conditions than IUCN Red List categories and criteria [46], which were built to be as general as possible. However, some general patterns at the landscape level might be found using national Red List data [43,47]. Therefore, we use the latest Czech Red List [48] here.
The aim of this study was to analyze the trends in the changes in the number of sites of rare species of orchids (those classified as “critically endangered” and “extinct” based on the national Red Lists) [40,41,48]. We attempted to put the available data into context with shifts in agricultural practices.
We asked the following questions: From what percentage of historically known sites did orchids disappear over the time period covered by our database (about 150 years)? Is there any difference in the number of sites from which orchids disappeared before, during and after the communist period in the Czech Republic? Which species of orchids in the Czech Republic are most in danger of becoming extinct, based on the percentage of historical sites still occupied?
2. Materials and Methods
2.1. Study Site
The Czech Republic is in central Europe and its flora is very well studied. Its average altitude is 450 m above sea level, a calculation based on the 30-sec altitude layer [49] and performed in ArcMap 10.1 [50]. The country is covered mainly by highlands of moderate altitude, whereas higher mountains occur at the borders with other countries (mainly in the north and south). The climate of the Czech Republic is typically temperate, with cold, cloudy winters and hot summers. However, there are some regional and local differences due to the relief that forms the complex topography in this area [51]. Because the Czech Republic is a relatively small country in terms of latitude range, temperature and precipitation are mostly affected by local heterogeneity and altitude [52].
2.2. The Database
The dataset of orchid records we used is based on that of the Nature Conservation Agency of the Czech Republic, which is not freely accessible. The records cover a period of about 150 years and include all known literature and reports from people on the records of orchids, the identities of which are subsequently checked by specialists. The database includes more than 115,000 individual records.
Each site is characterized by its GPS coordinates. Because of the high number of records and the extremely long timespan covered, the cross-checking procedure for duplicate records was not consistently conducted, especially in the older data. This means that in many cases, the same site may be called different names and the GPS coordinates may differ for the same sites in different records in the database.
We resolved this problem by using ArcMap 10.1 [50] to create a grid of 1 × 1 km squares. For each square, we then considered any recording with its GPS coordinates falling within this square as the same site. This is reasonable, as typical orchid sites cover several thousand square meters, and small adjacent sites are likely to be sub-sites of a larger site, or, in other words, an orchid meta-population.
For each of the sites (squares in our grid where an orchid population was recorded), we determined the latest year when orchids were still present at the site. If the year was 1990 or later, the site was considered still occupied (except in the case of two species mentioned in Section 3.1, which are commonly considered extinct in the Czech Republic, despite being found during 1990–2020). If the last record of an extant population at a site was prior to 1990, this date was considered to be the date of extinction of the orchid population at this site.
It is unlikely that this resulted in a bias in our calculations. First, in case of rare species of orchids, which is exclusively the case considered here, the Nature Conservation Agency of the Czech Republic meticulously re-checked all of the sites where these orchids were likely to survive. Second, if the date of the last record was prior to 1990, then the population might of course have survived for several additional years after this observation. However, as the dates of extinction are categorized into 20-year intervals (Table 1), it is most likely that the actual date of extinction is in the same 20-year period as the date of the last record.
2.3. Nomenclature
The classification, nomenclature and Red List classification (threat categories) of native taxa are based on Grulich [41,48] and Danihelka et al. [40]. The threat category of Himantoglossum adriaticum was taken from the Botany webpage [53], as there is information that H. adriaticum is still present at one locality in the Czech Republic, despite Danihelka et al. [40] referring to it as an extinct taxon.
2.4. Data Analysis
We categorized a total of 64 species of orchids into threat categories based on the Red List classification for the Czech Republic [40,41,48]. For the purpose of this study, we analyzed only the 34 species that are in the extinct (A1) and critically endangered (C1) categories. We categorized the A1 and C1 species according to the number of sites recorded in the database: species extinct in 2020, species recorded at less than 20 sites, species recorded at 21–100 sites, species recorded at 101–300 sites, species recorded at 301–1400 sites, and the special case of the newly described species Epipactis. Within each of these categories, we ranked the species according to the percentage of sites at which they survived up to the present time, from the lowest to the highest. For each species, we calculated the number of sites at which extinction was recorded during the selected time periods (see Table 1).
Thus, we mainly considered the political events most likely to have affected agricultural practices as milestones, rather than what was recorded at a particular interval of time. We looked for patterns of when orchids disappeared from sites for the different species and attempted to associate them with changes in society and political regimes that resulted in severe changes in agricultural practices.
The results we present in the figures for each interval are the number of sites for which the latest year of the orchid’s presence at the site was in that particular interval. These are referred to as “newly unoccupied sites”. The reason for this notation is that—as we described above—if a site was never recorded as occupied later than this interval, the orchid in question probably went extinct during this interval. The sites recorded as occupied in last interval are considered currently occupied.
3. Results and Discussion
The total number of sites for each species that were included in our database (i.e., which were recorded over the last 150 years), together with the percentages of those that remained occupied up to the present time, is shown in Table 2. In the following sub-sections, the results are individually analyzed for a selected group of orchids in each of the time intervals outlined in the Data Analysis section.
3.1. Extinct Species
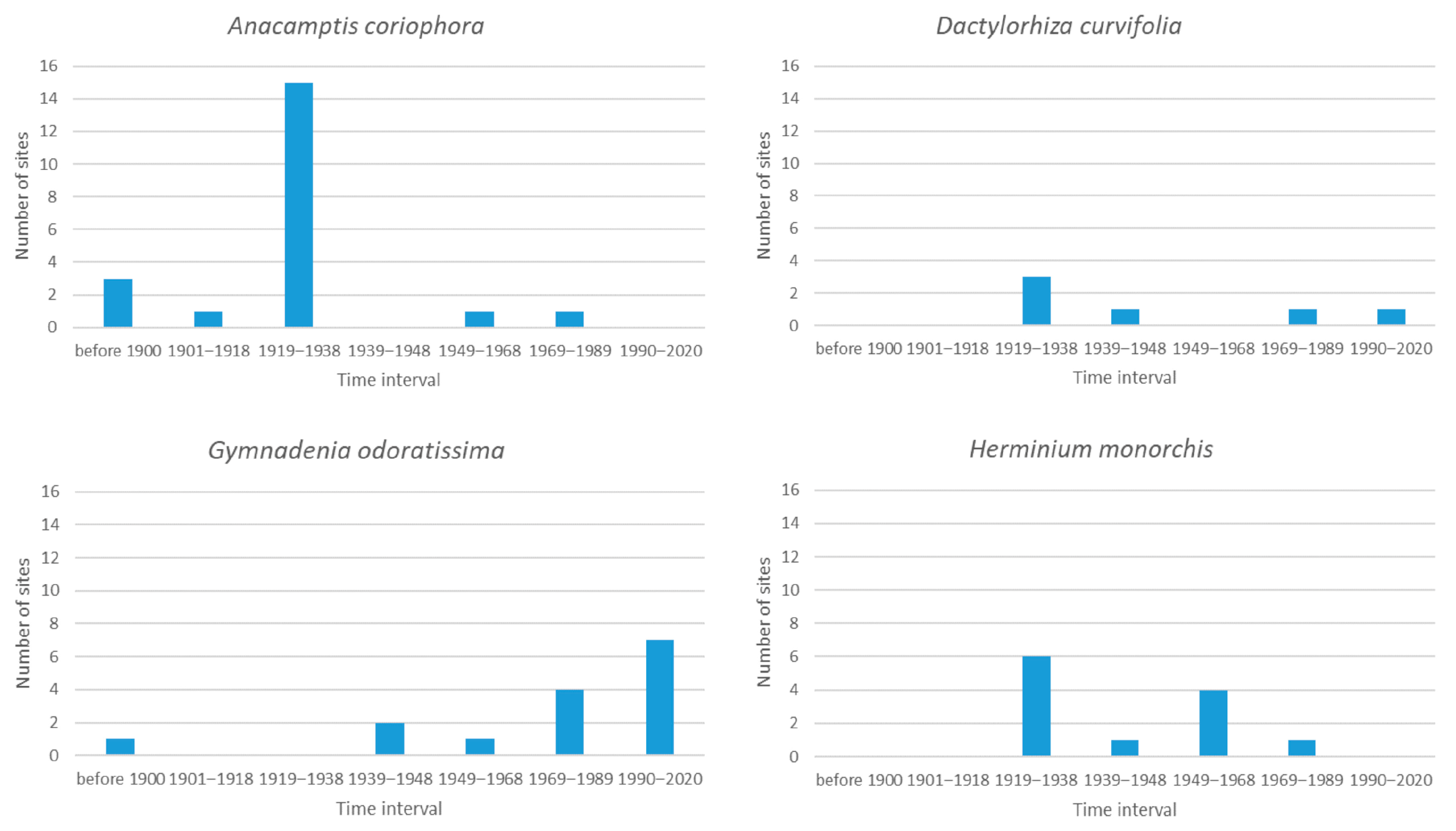
Figure 1 shows the results for the four species that are now considered to be extinct. Although for two species (Dactylorhiza curvifolia and Gymnadenia odoratissima) some sites existed during the beginning of the last period (1990–2020), the recent agreement [40,41,48] is that all of them are now extinct. It is clear that this was dramatic in the case of Anacamptis coriophora, as it disappeared from 15 sites between the First and Second World War. The last two sites that remained after the Second World War were recorded as newly unoccupied before the Velvet revolution in 1989. The reasons for this are probably changes in the management of meadows and overall environmental pollution [54]. Anacamptis coriophora is one of the most threatened orchids in several European countries—it is extinct in Flanders and the Netherlands [55] and is one of the most quickly declining species in France, Belgium and Luxembourg [56].
Dactylorhiza curvifolia and Herminium monorchis were only recorded at a few sites in the Czech Republic (6 and 12, respectively) and disappeared from these sites at almost the same rate over the period of this study. Kull et al. [12] identified H. monorchis as one of the most threatened species in Europe. Herminium monorchis went extinct after the Velvet Revolution in 1989, mainly because of the changes in agriculture and the afforestation of meadows and pastures [53]. The situation with D. curvifolia is a bit complicated. According to the records in the database, it was present even after 1990. However, literature dates the last confirmed locality to the 1930s [53] or 1953 [54]. These inconsistencies could be caused by incorrect identification in the field. Moreover, some historical records of this species from 1930s are classified as the later described Dactylorhiza bohemica [54] or D. traunsteineri in 1970s [53].
The case of Gymnadenia odoratissima is very interesting. This species was also only recorded at 15 sites during the 20th century, and these sites were steadily recorded as newly unoccupied from the beginning of the Second World War, half (seven) of which were newly unoccupied after 1990. The last confirmed locality, in White Carpathians, was destroyed by intensive cattle grazing. However, suitable habitats are still present in White Carpathians, and there is still a small chance of recording this species there in the future [54]. The region of distribution of this species is restricted to Europe. It is classified as a potentially endangered species in Slovakia [53] and is rare in Sweden [57].
3.2. Species with 1 to 20 Sites
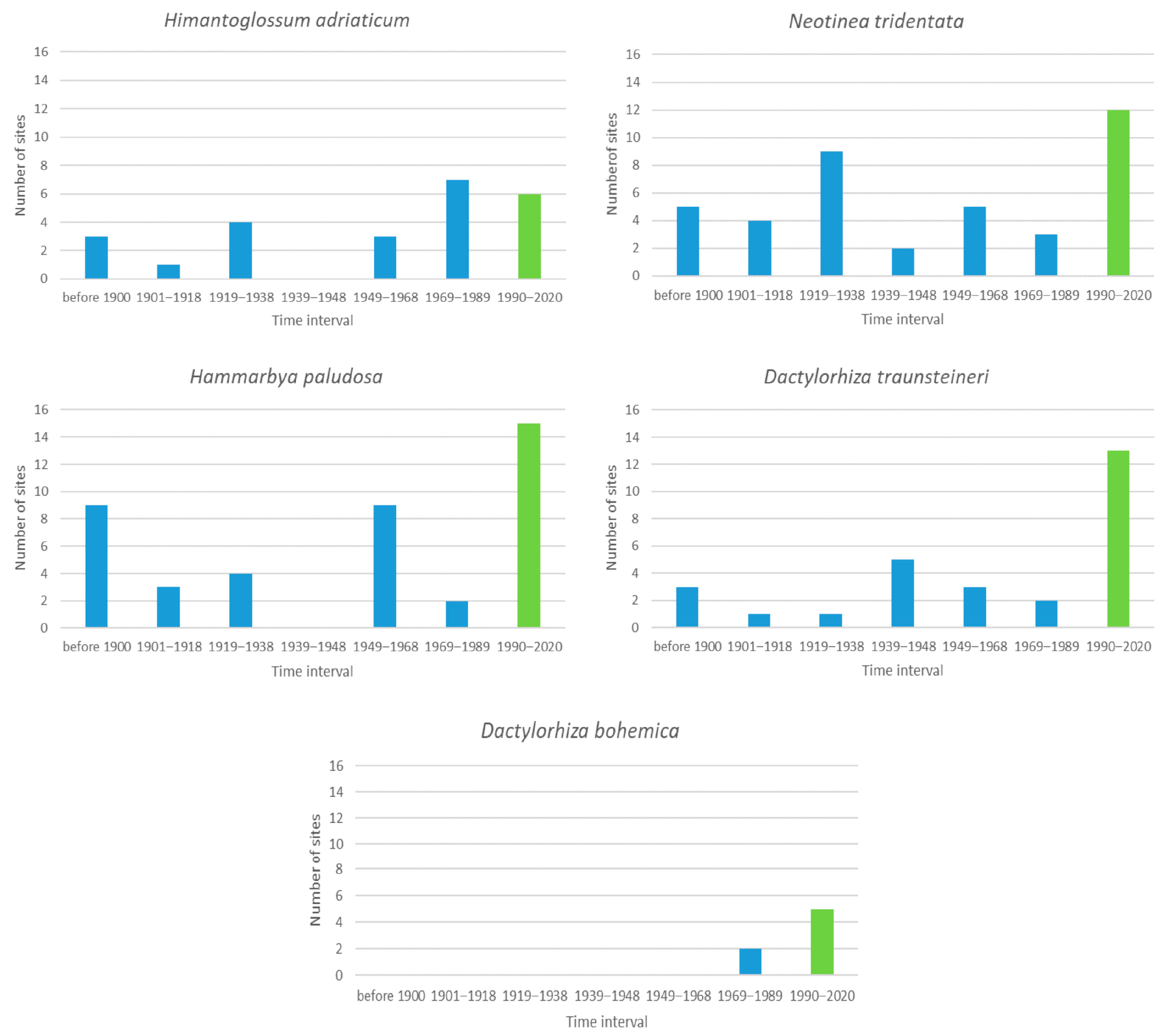
Figure 2 shows the results for the five species that were recorded historically at 1 to 20 sites. For the three species for which more than 40% of the incidence of newly unoccupied sites was recorded, it seems that the number of newly unoccupied sites before the Second World War compared with those after the Second World War (i.e., before or after 1939) is larger for Neotinea tridentata and Hammarbya paludosa, and smaller for Himantoglossum adriaticum. None of these differences, however, were statistically significant (t-test, p > 0.05). In Europe, H. adriaticum is found from the British Isles and France to central European Russia, extending to northern Scandinavia and northernmost Italy; elsewhere, it is very rare [58]. Now, only three sites are extant in the Czech Republic, with the other occurrences having vanished, apparently due to eutrophication followed by succession [58]. Bódis et al. [59] conclude that the decline in dry grasslands due to the abandonment of traditional land-use practices (mowing or grazing) is the most serious threat to H. adriaticum in Hungary.
For Dactylorhiza traunsteineri (46% of sites still occupied) there is no difference between the pre-1939 and post-1939 periods (t-test, p > 0.05). The results for Dactylorhiza bohemica must be treated with caution, because this is a newly described species probably derived from D. traunsteineri [60]. It is considered endemic to the Czech Republic, and the two recently known populations are found within a protected area that seems not to be currently threatened [61].
3.3. Species with 21 to 100 Sites
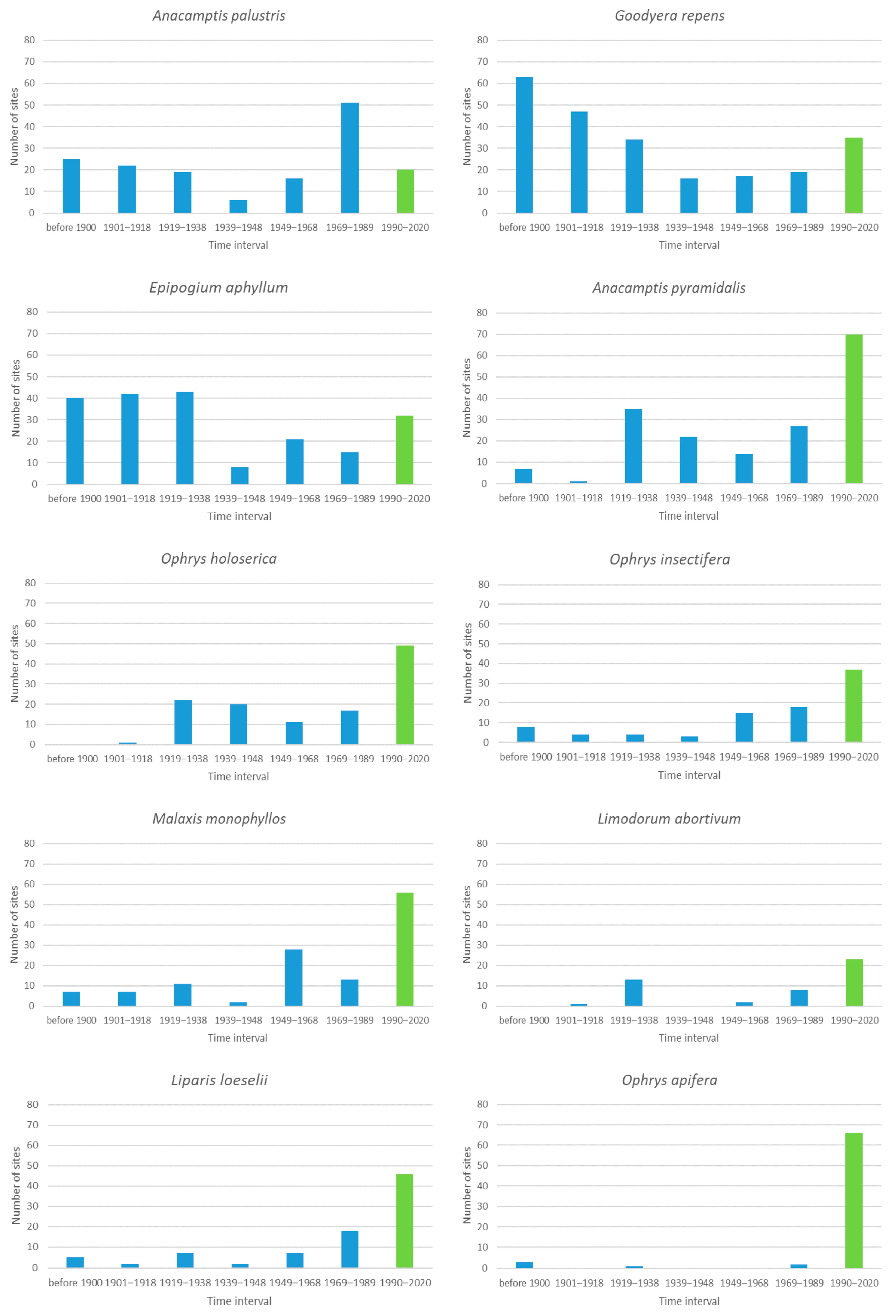
There are 10 species that occur historically at 21 to 100 sites (Figure 3). For three of these (Anacamptis palustris, Epipogium aphyllum, Goodyera repens), less than 20% of the sites are currently occupied. For Anacamptis palustris, the critical period was 1969–1989, when it disappeared from 51 (one third) of its historical sites. For the latter two species, the number of sites newly unoccupied before 1939 was significantly greater than those newly unoccupied after 1939 (t-test, p = 0.02 for G. repens and p = 0.002 for E. aphyllum).
This disparity may be due to both being small forest species and, therefore, easily overlooked by orchid hunters willing to go to forests to look for orchids, which at that time was not a priority. It was found that G. repens is highly specific to its habitat [54,62] and sensitive to soil nutrients, which makes this species a good indicator of forest succession [62]. This might be the reason for its decline in the Czech Republic, where its decrease lies in excessive forest management [54]. In Ukraine, anthropogenic factors eliminate its populations [63].
The results for E. aphyllum should be treated with caution because the appearance of this species above ground is very irregular, and it can remain dormant for several years [64]. However, this species is considered extremely rare in Hungary [65] and one of the rarest orchids in Britain, facing an extinction risk in the near future [66].
A further six species in this group disappeared from 40–53% of their sites. Visual inspection of Figure 3 indicates that there might be some increase in the number of sites newly unoccupied in the more recent time intervals, at least for some species. A t-test (p > 0.05) reveals that if the year 1939 is taken as a breakpoint, there are no differences between the pre-1939 and post-1939 time intervals. In contrast, however, if the year 1948 (the onset of communism in the Czech Republic) is considered as a breakpoint, the number of newly unoccupied species in the post-1948 time intervals is significantly (or almost significantly) larger than those pre-1948 for Ophrys insectifera (t-test, p = 0.003) and Malaxis monophyllos (t-test, p = 0.06—close to significance). For Liparis loeselii, the t-test is not significant, but the p value (p = 0.08) is relatively small, although it exceeds 0.05. It would be interesting to test whether the period of communism really affected the disappearance of O. insectifera, M. monophyllos and maybe even L. loeselii. This goes far beyond the scope of this paper. Possible candidates explaining this potential phenomenon might be the loss of suitable habitats for O. insectifera (and partly also for M. monophyllos) as it mainly occurs in pastures and grasslands, which were often changed into fields in the past. A possible explanation for the loss of many localities of L. loeselii might be hidden in changes in the water regime in the swamps, moors and bogs which this species favors. Similarly, Kaplan et al. [58] stated that the majority of populations disappeared in the late 19th century or during the first half of the 20th century due to direct habitat destruction (including drainage and conversion to arable land) and as a result of mowing abandonment and eutrophication followed by succession. The situation has improved recently [67], but the seed bank in the soil might not be sufficient for re-establishment in its previous localities.
The last species in this group, Ophrys apifera, is a real puzzle. Despite a relatively large number of historical sites (72), it only disappeared from six during the 150 years covered by this study. Why O. apifera has survived so well is unknown. One possible explanation might be that this species is highly valued and so its numbers and localities are closely monitored and its habitats protected. This species was also mentioned as a new orchid for Sweden [68].
3.4. Species with 101 to 300 Sites
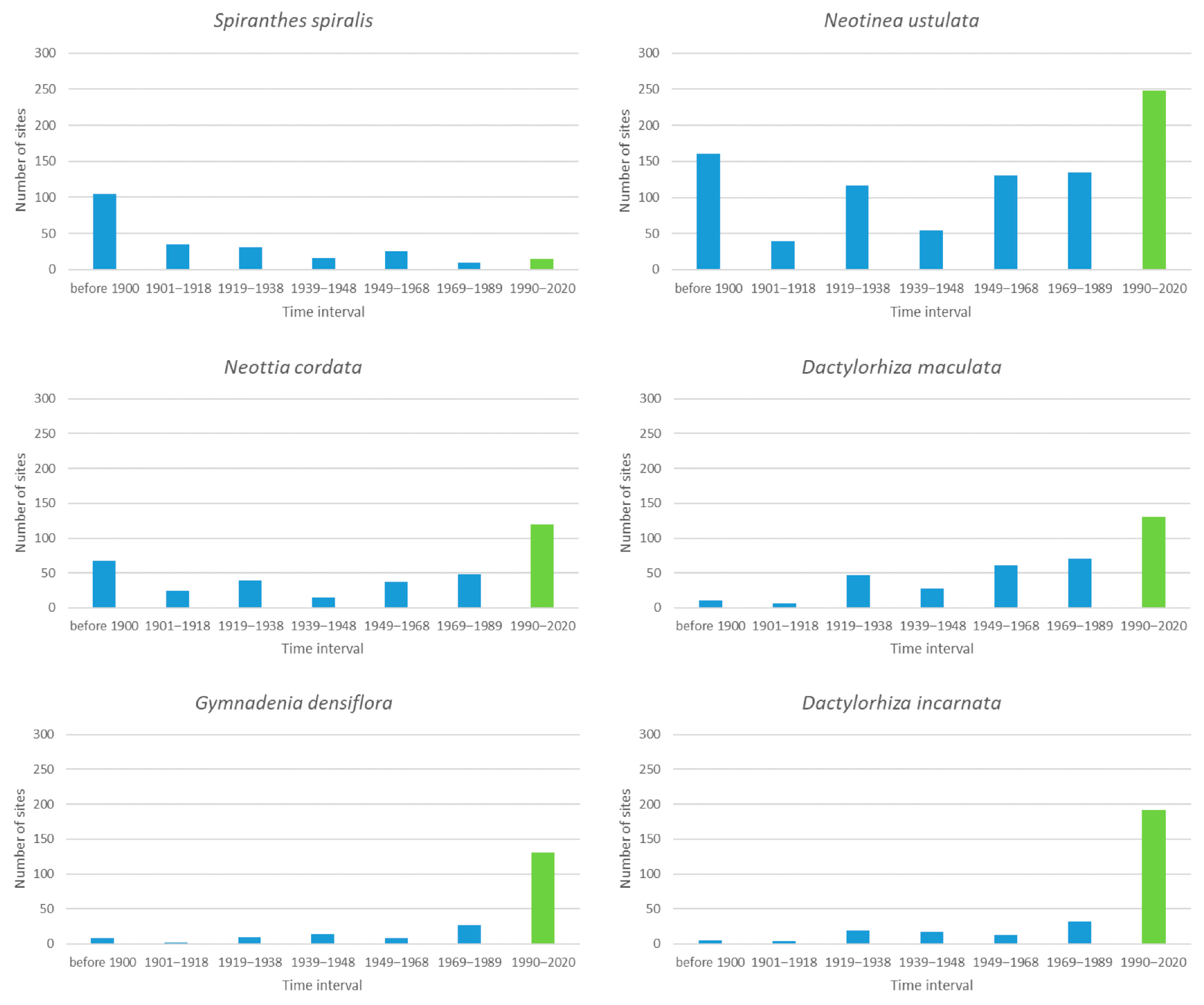
Out of the six species, for which 101–300 sites were recorded in the database (Figure 4), Spiranthes spiralis, with the extremely low survival rate of its historical sites (only 6/%), is outstanding. Almost half of these sites (105 out of 237) went extinct even before 1900. Previous studies from Europe [23,55,69,70,71] suggest that the huge decline in S. spiralis populations is linked to the loss of its natural habitats, mainly because of agricultural intensification and the cessation of traditional management practices. We assume that these changes also play a role in the case of the population loss of S. spiralis in the Czech Republic.
Among other species, in Dactylorhiza maculata, the number of newly unoccupied sites after 1948 seems to be visually larger than that before 1948, but the difference is not statistically significant (t-test, p = 0.05). No trends are observable for the other species, either (t-test, p = 0.05).
The site survival rate of four out of the six species contained in this group (S. spiralis, Neotinea ustulata, Neottia cordata, D. maculata) is smaller than 40%, while the survival rate of the remaining two (Gymnadenia densiflora, Dactylorhiza incarnata) is larger than 60%. We can only speculate as to what is causing these differences.
3.5. Species with More Than 300 Sites
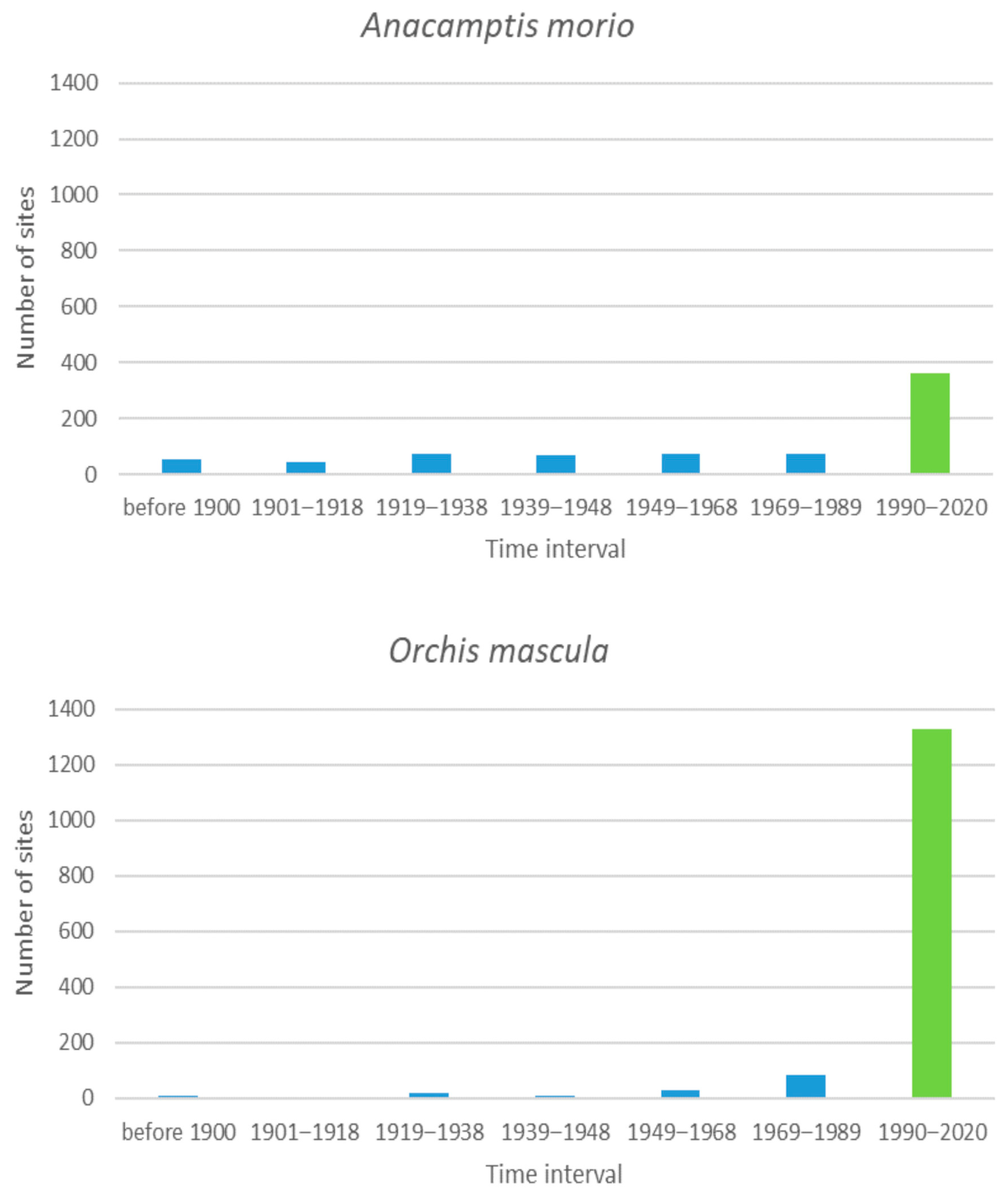
There are only two species with more than 300 sites among the endangered and critically endangered orchids: Anacamptis morio and Orchis mascula (Figure 5). Both of them show a similar pattern: a large survival rate of their sites (about half for A. morio and almost 90% for O. mascula). There are no significant trends between different time intervals in the past (t-test, p = 0.05). The recorded high survival rate of these two species over time in the Czech Republic could be attributed to an increased sampling effort during the last two decades. This is because the number of orchidologists (both experts and amateurs) increased compared to that during previous periods, and both species are now well studied in this country [25,28,72].
3.6. New Species of Epipactis
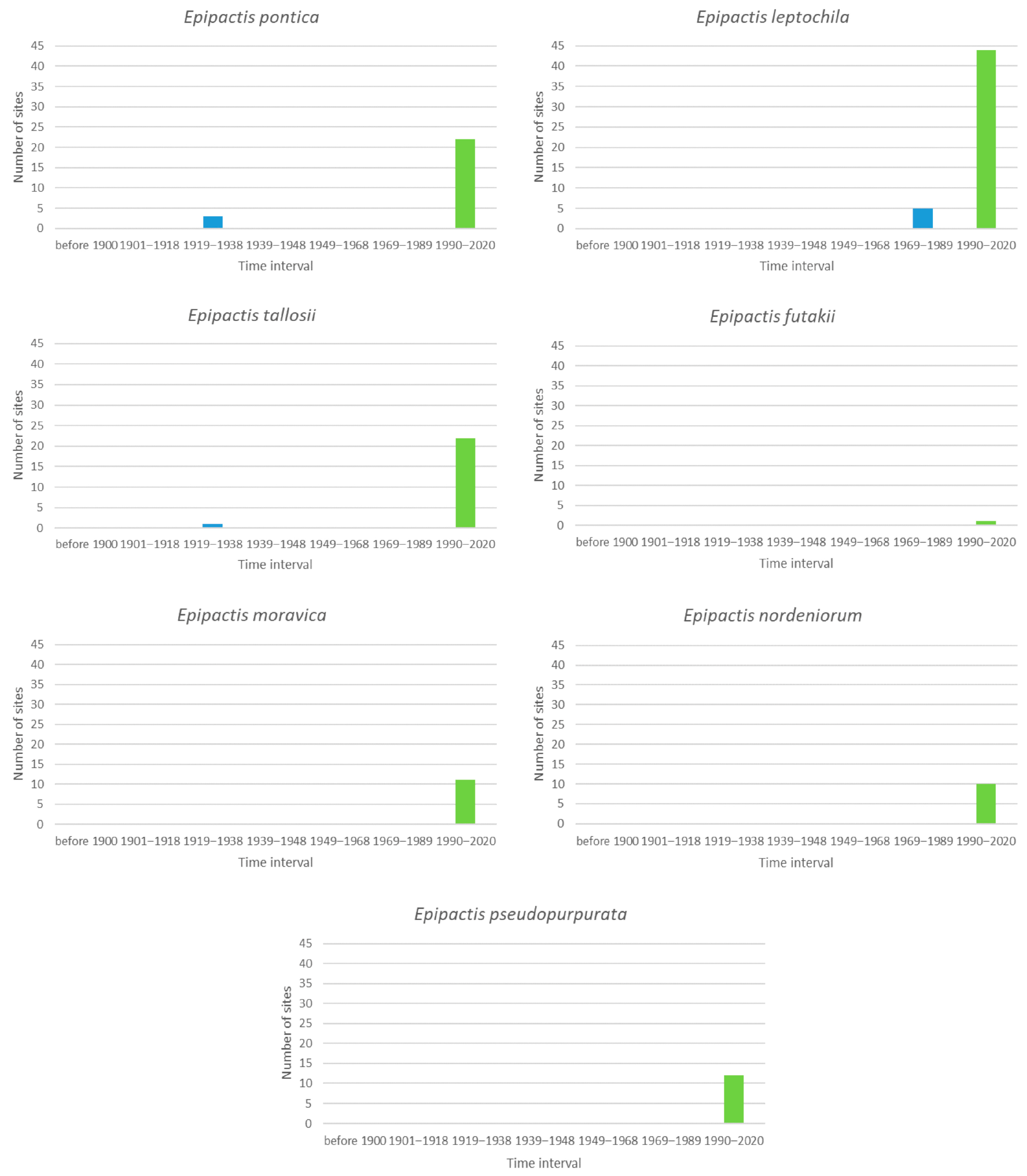
Finally, Figure 6 shows the situation for the newly described species of the genus Epipactis, other than Epipactis pontica and Epipactis tallosii, which were first recorded in the period 1919–1938 in the database. Because the recorded history of these species is very short, one cannot draw any conclusions from these data.
3.7. General Analysis
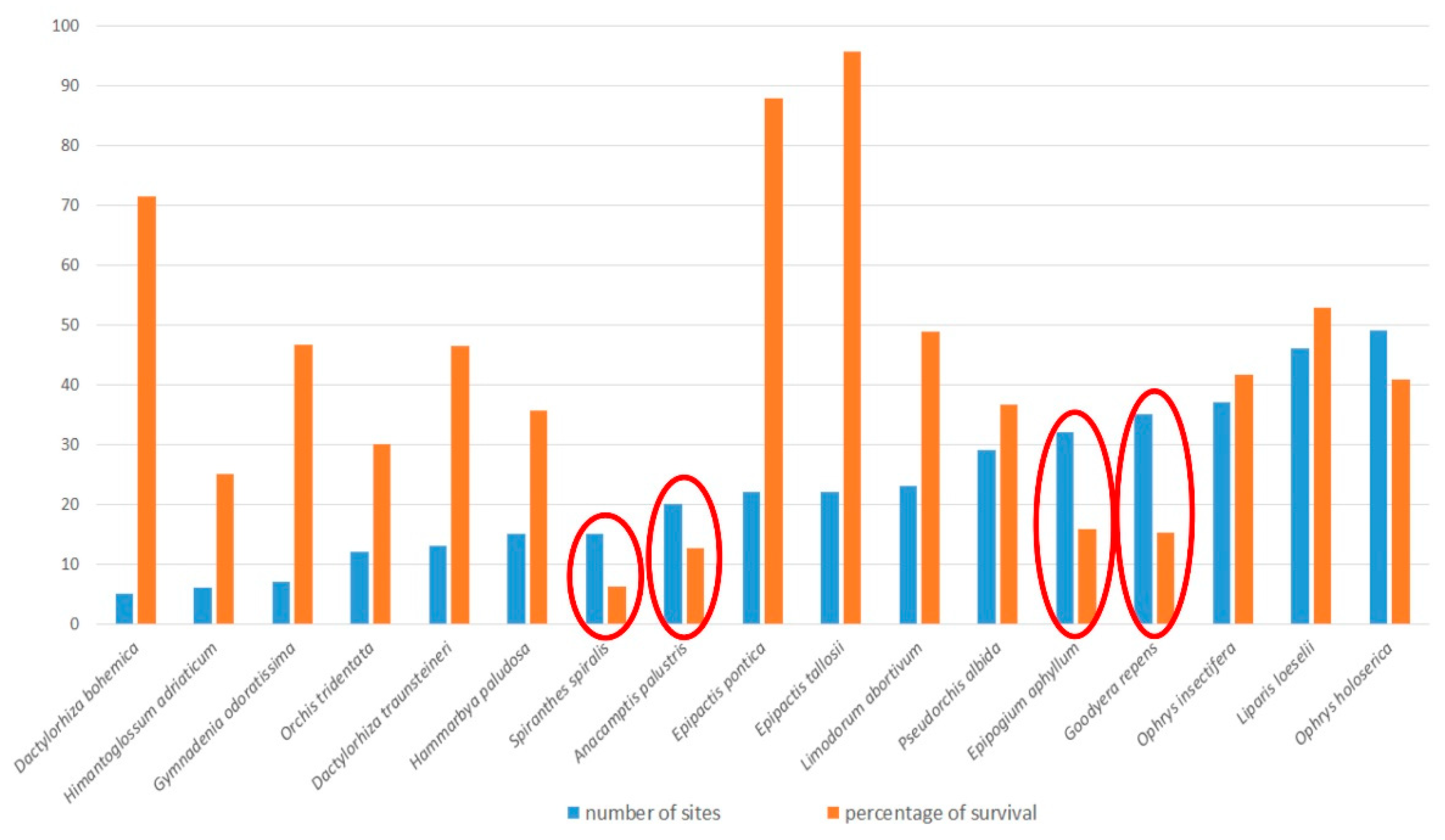
Figure 7 shows the situation for species of orchids in the Czech Republic with less than 50 sites occupied in the last period, ranked in order of the number of occupied sites in the last period (blue bars). The orange bars indicate the percentage of their original sites still occupied (the rate of decline of the number of their sites historically). The four encircled species occupy less than 50 sites in the most recent (last) period and, simultaneously, their present number of sites is less than 20% of that of all their sites in the database. Therefore, we conclude that these species need urgent protection. These species are Spiranthes spiralis, Anacamptis palustris, Epipogium aphyllum and Goodyera repens. The first two of these species occur in open habitats, mainly meadows and pastures, and their decline is caused by the cessation of traditional management and the use of artificial fertilizers. Similar results were also observed in other European countries [55,70,71]. Anacamptis palustris was adversely affected by the changes in water regime on its localities—because of either drainage or excessive wetting and the subsequent expansion of reed. Thus, the remaining localities need to be suitably managed in order to safeguard this species. On the other hand, E. aphyllum and G. repens are forest species, sensitive to changes in soil and the subsequent changes in the mycorrhizal community [54,62]. Preserving ancient forest habitats seems to be the best way to ensure the survival of these species in the Czech Republic.
4. Other Aspects That May Cause Decline in Number of Orchid Sites
Of course, there are aspects other than the changes in agricultural practices that may play an important role in the decline of the number of orchid sites. One such example is climate change. However, the effect of climate change is not thoroughly explored, and it is out of the scope of the current manuscript. Additionally, environmental variability also causes changes in fecundity [31]. In this paper, we found that the four most threatened species are vanishing from Czech orchid flora because of the excessive use or alteration of their habitats, mainly caused by human impact. Thus, we emphasize protecting the natural habitats where Czech orchids occur to preserve them in their natural environment.
Author Contributions
Conceptualization, Z.Š. and P.K.; methodology, Z.Š. and P.K.; software, Z.Š. and P.K.; validation, Z.Š. and P.K.; formal analysis, Z.Š. and P.K.; investigation, Z.Š. and P.K.; resources, Z.Š. and P.K.; data curation, Z.Š. and P.K.; writing—original draft preparation, Z.Š. and P.K.; writing—review and editing, Z.Š. and P.K.; visualization, Z.Š. and P.K.; supervision, P.K.; project administration, P.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Restrictions apply to the availability of these data. Data were obtained from the Nature Conservation Agency of the Czech Republic and are available at https://portal.nature.cz/publik_syst/ctihtmlpage.php?what=3&nabidka=hlavni&variantaPrihlaseni=ISOP (accessed on 1 February 2021) with the permission of the Nature Conservation Agency of the Czech Republic.
Acknowledgments
We thank the Nature Conservation Agency of the Czech Republic for giving us permission to use their dataset. We are greatly indebted to Tony Dixon for uncountable helpful hints as to how to improve the style of English in this paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gaston, K.J.; Blackburn, T.M. Patterns and Process in Macroecology; Blackwell Science Ltd.: Cambridge, UK, 2000. [Google Scholar]
- Fischer, M.; Stöcklin, J. Local extinction of plants in remnants of extensively used calcareous grasslands 1950–1985. Conserv. Biol. 1997, 11, 727–737. [Google Scholar] [CrossRef]
- Kull, T.; Kukk, T.; Leht, M.; Krall, H.; Kukk, Ü.; Kull, K.; Kuusk, V. Distribution trends of rare vascular plant species in Estonia. Biodivers. Conserv. 2002, 11, 171–196. [Google Scholar] [CrossRef]
- Bianchi, F.J.J.A.; Honěk, A.; van der Werf, W. Changes in agricultural land use can explain population decline in a ladybeetle species in the Czech Republic: Evidence from a process-based spatially explicit model. Landsc. Ecol. 2007, 22, 1541–1554. [Google Scholar] [CrossRef]
- Le Roux, J.J.; Hui, C.; Castillo, M.L.; Iriondo, J.M.; Keet, J.-H.; Khapugin, A.A.; Médail, F.; Rejmánek, M.; Theron, G.; Yannelli, F.A.; et al. Recent anthropogenic plant extinctions differ in biodiversity hotspots and coldspots. Curr. Biol. 2019, 29, 2912–2918. [Google Scholar] [CrossRef] [PubMed]
- Khapugin, A.A.; Kuzmin, I.V.; Silaeva, T.B. Anthropogenic drivers leading to reagional extinction of threatened plants: Insights from regional Red Data Books of Russia. Biodivers. Conserv. 2020, 29, 2765–2777. [Google Scholar] [CrossRef]
- Knapp, W.M.; Frances, A.; Noss, R.; Naczi, R.F.C.; Weakley, A.; Gann, G.D.; Baldwin, B.G.; Miller, J.; McIntyre, P.; Mishler, B.D.; et al. Vascular plant extinction in the continental United States and Canada. Conserv. Biol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Bilz, M.; Kell, S.P.; Maxted, N.; Lansdown, R.V. European Red List of Vascular Plants; Publication Office of the European Union: Luxembourg, 2011. [Google Scholar]
- Henle, K.; Alard, D.; Clitherow, J.; Cobb, P.; Firbank, L.; Kull, T.; McCracken, D.; Moritz, R.F.A.; Niemela, J.; Rebane, M.; et al. Identifying and managing the conflicts between agriculture and biodiversity conservation in Europe—A review. Agr. Ecosyst. Environ. 2008, 124, 60–71. [Google Scholar] [CrossRef]
- Stewart, J. The Conservation of European Orchids. Nature and Environment, No. 57; Council of Europe Press: Strasbourg, France, 1992. [Google Scholar]
- Tsiftsis, S.; Tsiripidis, I.; Trigas, P. Identifying important areas for orchid conservation in Crete. Eur. J. Environ. Sci. 2011, 1, 28–37. [Google Scholar] [CrossRef]
- Kull, T.; Selgis, U.; Pecina, M.V.; Metsare, M.; Ilves, A.; Tali, K.; Sepp, K.; Kull, K.; Shefferson, R.P. Factors influencing IUCN threat levels to orchids across Europe on the basis of national Red Lists. Ecol. Evol. 2016, 6, 6245–6265. [Google Scholar] [CrossRef]
- Wädekin, K.E. Agrarian Policies in Communist Europe. A Critical Introduction; Allanheld, Osmun Publishers: Totowa, NJ, USA, 1982. [Google Scholar]
- Adams, A.E.; Adams, J.S. Men Versus Systems. Agriculture in the USSR, Poland, and Czechoslovakia; Free Press: New York, NY, USA, 1971. [Google Scholar]
- Krčmářová, J.; Jeleček, L. Czech traditional agroforestry: Historic and current status. Agroforest Syst. 2017, 91, 1087–1100. [Google Scholar] [CrossRef]
- Skaloš, J.; Wber, M.; Lipský, Z.; Trpáková, I.; Šnatrůčková, M.; Uhlířová, L.; Kukla, P. Using old military survey maps and orthophotograph maps to analyse long-term land cover changes—Case study (Czech Republic). Appl. Geogr. 2011, 31, 426–438. [Google Scholar] [CrossRef]
- Reif, J.; Voříšek, P.; Šťastný, K.; Bejček, V.; Petr, J. Agricultural intensification and farmland birds: New insight from a central European country. Ibis 2008, 150, 596–605. [Google Scholar] [CrossRef]
- Swarts, D.N.; Dixon, K.W. Terrestrial orchid conservation in the age of extinction. Ann. Bot. 2009, 104, 543–556. [Google Scholar] [CrossRef] [Green Version]
- Djordjević, V.; Lukašić, D.; Jovanović, S.; Stevanović, V. Distribution and conservation status of some rare and threatened orchid taxa in the central Balkans and the southern part of the Pannonian Plain. Wulfenia 2017, 24, 143–162. [Google Scholar]
- Khapugin, A.A. A global systematic review on orchid data in protected areas. Nat. Conserv. Res. 2020, 5 (Suppl. 1), 19–33. [Google Scholar] [CrossRef]
- Govaerts, R. World Checklist of Orchidaceae; The Royal Botanic Gardens: Kew, UK, 2020; Available online: http://wcsp.science.kew.org/ (accessed on 31 January 2020).
- Cribb, P.J.; Kell, S.P.; Dixon, K.W.; Barrett, R.L. Orchid conservation: A global perspective. In Orchid Conservation; Dixon, K.W., Kell, S.P., Barrett, R.L., Cribb, P.J., Eds.; Natural History Publications: Kota Kinabalu, Sabah, 2003; pp. 1–2. [Google Scholar]
- Kull, T.; Hutchings, M.J. A comparative analysis of decline in the distribution ranges of orchid species in Estonia and the United Kingdom. Biol. Conserv. 2006, 129, 31–39. [Google Scholar] [CrossRef]
- Wagensommer, R.P.; Medagli, P.; Turco, A.; Perrino, E.V. IUCN Red List evaluation of the Orchidaceae endemic to Apulia (Italy) and considerations of the application of the IUCN protocol to rare species. Nat. Conserv. Res. 2020, 5 (Suppl. 1), 90–101. [Google Scholar] [CrossRef]
- Jersáková, J.; Kindlmann, P.; Stříteský, M. Population dynamics of Orchis morio in the Czech Republic under human influence. In Trends and Fluctuations and Underlying Mechanisms in Terrestrial Orchid Populations; Kindlmann, P., Willems, J.H., Whigham, D.F., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2002; pp. 209–224. [Google Scholar]
- Wotavová, K.; Balounová, Z.; Kindlmann, P. Factors affecting persistence of terrestrial orchids in wet meadows and implications for their conservation in a changing agricultural landscape. Biol. Conserv. 2004, 118, 271–279. [Google Scholar] [CrossRef]
- Janečková, P.; Wotavová, K.; Schödelbauerová, I.; Jersáková, J.; Kindlmann, P. Relative effects of management and environmental conditions on performance and survival of population of a terrestrial orchid, Dactylorhiza majalis. Biol. Conserv. 2006, 129, 40–49. [Google Scholar] [CrossRef]
- Štípková, Z.; Kindlmann, P. Extent and reasons for meadows in South Bohemia becoming unsuitable for orchids. Eur. J. Environ. Sci. 2015, 5, 124–147. [Google Scholar] [CrossRef] [Green Version]
- Darwin, C. The Various Contrivances by which Orchids are Fertilized by Orchids; John Murray: London, UK, 1862. [Google Scholar]
- Neiland, M.R.M.; Wilcock, C.C. Fruit set, nectar reward, and rarity in the Orchidaceae. Am. J. Bot. 1998, 85, 1657–1671. [Google Scholar] [CrossRef] [PubMed]
- Charitonidou, M.; Halley, J.M. What goes up must come down—Why high fecundity orchids challenge conserve beliefs. Biol. Conserv. 2020, 252, 108835. [Google Scholar] [CrossRef]
- Kull, T.; Kindlmann, P.; Hutchings, M.; Primack, R. Conservation biology of orchids: Introduction to the special issue. Biol. Conserv. 2006, 129, 1–3. [Google Scholar] [CrossRef]
- Zhang, Z.; Yan, Y.; Tian, Y.; Li, J.; He, J.-S.; Tang, Z. Distribution and conservation of orchid species richness in China. Biol. Conserv. 2015, 181, 64–72. [Google Scholar] [CrossRef]
- Zheleznaya, E. Results of a study of Cypripedium in several regions of Siberia (Russia). Eur. J. Environ. Sci. 2015, 5, 134–141. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, P.; Van Staden, J. Ansellia africana (Leopard orchid): A medicinal orchid species with untapped reserves of important biomolecules—A mini review. S. Afr. J. Bot. 2016, 106, 181–185. [Google Scholar] [CrossRef]
- Kolanowska, M.; Baranow, P.; Rykaczewski, M. Elleanthus ortizianus—A new orchid species from southern Colombia. Bot. Lett. 2017, 164, 155–158. [Google Scholar] [CrossRef]
- Tsiftsis, S.; Tsiripidis, I. Temporal and spatial patterns of orchid species distribution on Greece: Implications for conservation. Biodivers. Conserv. 2020, 29, 3461–3489. [Google Scholar] [CrossRef]
- Tatarenko, I.; Dodd, M.; Wallace, H.; Bellamy, G.; Fleckney, A. Protecting small populations of rare species. Case study on Dactylorhiza viridis (Orchidaceae) in Fancott Woods and Meadows SSSI, Bedfordshire, UK. Nat. Conserv. Res. 2020, 5 (Suppl. 1), 165–171. [Google Scholar] [CrossRef]
- Zizka, A.; Silvestro, D.; Vitt, P.; Knight, T.M. Automated conservation assessment of the orchid family with deep learning. Conserv. Biol. 2020. [Google Scholar] [CrossRef]
- Danihelka, J.; Chrtek, J., Jr.; Kaplan, Z. Checklist of vascular plants of the Czech Republic. Preslia 2012, 84, 647–811. [Google Scholar]
- Grulich, V. Red list of vascular plants of the Czech Republic: 3rd edition. Preslia 2012, 84, 631–645. [Google Scholar]
- Hoffmann, M.; Brooks, T.M.; da Fonseca, G.A.; Gascon, C.; Hawkins, A.F.A.; James, R.E.; Langhammer, P.; Mittermeier, R.A.; Pilgrim, J.D.; Rodrigues, A.S.L.; et al. Conservation planning and the IUCN Red List. Endanger. Species Res. 2008, 6, 113–125. [Google Scholar] [CrossRef]
- Zamin, T.; Baillie, J.E.M.; Miller, M.; Rodriguez, J.; Ardid, A.; Collen, B. National Red Listing beyond the 2010 Target. Conserv. Biol. 2010, 24, 1012–1020. [Google Scholar] [CrossRef]
- Corlett, R.T. Safeguarding our future by protecting biodiversity. Plant Divers. 2020, 42, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Maes, D.; Isaac, N.J.B.; Harrower, C.A.; Collen, B.; van Strien, A.J.; Roy, D.B. The use of opportunistic data for IUCN Red List assessment. Biol. J. Lin. Soc. 2015, 115, 690–706. [Google Scholar] [CrossRef] [Green Version]
- Sharrock, S.; Jones, M. Conserving Europe´s Threatened Plants: Progress towards Target 8 of the Global Strategy for Plant Conservation; Botanic Gardens Conservation International: Richmond, UK, 2009. [Google Scholar]
- Bachman, S.P.; Field, R.; Reader, T.; Raimondo, D.; Donaldson, J.; Schatz, G.E.; Lughadha, E.N. Progress, challenges and opportunities for Red Listing. Biol. Conserv. 2019, 234, 45–55. [Google Scholar] [CrossRef]
- Grulich, V. The Red List of vascular plants of the Czech Republic. Příroda 2017, 35, 75–132. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1km spatial resolution climate surface for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Desktop: Release 10; Environmental Systems Research Institute: Redlands, CA, USA, 2012. [Google Scholar]
- Climatic Conditions of the Czech Republic. Available online: https://geography.upol.cz/soubory/lide/smolova/GCZ/GCZ_Klima.pdf (accessed on 6 January 2020).
- Štípková, Z.; Tsiftsis, S.; Kindlmann, P. Pollination mechanisms are driving orchid distribution in space. Sci. Rep. 2020, 10, 850. [Google Scholar] [CrossRef] [Green Version]
- Botany.cz. Available online: https://botany.cz/cs/ (accessed on 15 December 2020).
- Průša, D. Orchideje České Republiky; Computer Press: Brno, Czech Republic, 2005. [Google Scholar]
- Jacquemyn, H.; Brys, R.; Hermy, M.; Willems, J. Does nectar reward affect rarity and extinction probabilities of orchid species? As assessment using historical records from Belgium and the Netherlands. Biol. Conserv. 2005, 12, 257–263. [Google Scholar] [CrossRef]
- Vogt-Schilb, H.; Munoz, F.; Richard, F.; Schatz, B. Recent declines and range changes of orchids in Western Europe (France, Belgium and Luxembourg). Biol. Conserv. 2015, 190, 133–141. [Google Scholar] [CrossRef]
- Gustafsson, S.; Sjogren-Gulve, P. Genetic diversity in the rare orchid, Gymnadenia odoratissima and a comparison with the more common congener, G-conopsea. Conserv. Genet. 2002, 3, 225–234. [Google Scholar] [CrossRef]
- Kaplan, Z.; Koutecký, P.; Danihelka, J.; Šumberová, K.; Ducháček, M.; Štěpánková, J.; Ekrt, L.; Grulich, V.; Řepka, R.; Kubát, K.; et al. Distributions of vascular plants in the Czech Republic. Part 6. Preslia 2018, 90, 235–346. [Google Scholar] [CrossRef]
- Bódis, J.; Biró, E.; Nagy, T.; Takács, A.; Sramkó, G.; Bateman, R.M.; Gilian, L.; Illyes, Z.; Tokolyi, J.; Lukacs, B.A.; et al. Biological flora of Central Europe Himantoglossum adriaticum H. Baumann. Perspect. Plant Ecol. 2019, 40, 125461. [Google Scholar] [CrossRef] [Green Version]
- Eccarius, W. Die Orchideengattung Dactylorhiza: Phylogenie, Taxonomie, Morphologie, Biologie, Verbreitung, Ökologie und Hybridisation; Wolfgang Eccarius: Eisenach, Germany, 2016. [Google Scholar]
- Kaplan, Z.; Danihelka, J.; Šumberová, K.; Chrtek, J., Jr.; Rotreklová, O.; Ekrt, L.; Štěpánková, J.; Taraška, V.; Trávníček, B.; Prančl, J.; et al. Distributions of vascular plants in the Czech Republic. Part 5. Preslia 2017, 89, 333–439. [Google Scholar] [CrossRef]
- Tsiftsis, S.; Tsiripidis, I.; Papaioannou, A. Ecology of the orchid Goodyera repens in its southern distribution limits. Plant Biosyst. 2012, 146, 857–866. [Google Scholar] [CrossRef]
- Melnyk, V.I. Geographical distribution, habitats and modern state of populations of Goodyera repens (Orchidaceae) in Ukraine. Ukrayins´kyi Bot. Zhurnal 2015, 72, 364–373. [Google Scholar] [CrossRef] [Green Version]
- Bjørndalen, J.E. Protection of Norwegian orchids—A review of achievements and challenges. Eur. J. Environ. Sci. 2015, 5, 121–133. [Google Scholar] [CrossRef] [Green Version]
- Nagy, T.; Notari, K.; Takacs, A.; Malkocs, T.; Tokolyi, J.; Molnar, A.V. Precipitation and timing of flowering in Ghost orchids (Epipogium aphyllum Sw.). Acta Bot. Hung. 2018, 60, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Taylor, L.; Roberts, D.L. Biological Flora of the British Isles: Epipogium aphyllum Sw. J. Ecol. 2011, 99, 878–890. [Google Scholar] [CrossRef]
- Bufková, I. Náprava narušeného vodního režimu rašelinišť v národním parku Šumava. Ochr. Přírody 2013, 2, 17–19. [Google Scholar]
- Nilsson, S. Ophrys apifera, a new orchid for Sweden. Bot. Not. 2014, 147, 1–4. [Google Scholar]
- Pignatti, S. Flora d´Italia. III; Edagricole: Bologna, Italy, 1982. [Google Scholar]
- Jacquemyn, H.; Hutchings, M.J. Biological flora of the British Isles: Spiranthes spiralis (L.) Chevall. J. Ecol. 2010, 98, 1253–1267. [Google Scholar] [CrossRef]
- Feldmann, P.; Prat, D. Conservation recommendations from a large survey of French orchids. Eur. J. Environ. Sci. 2011, 1, 18–27. [Google Scholar] [CrossRef] [Green Version]
- Vogt-Schilb, H.; Těšitelová, T.; Kotilínek, M.; Sucháček, P.; Kohout, P.; Jersáková, J. Altered rhizoctonia assemblages in grasslands on ex-arable land support germination of mycorrhizal generalist, not specialist orchids. New Phytol. 2020, 227, 1200–1212. [Google Scholar] [CrossRef]
Figure 1.
The number of sites in particular intervals for the four species that are now considered to be extinct in the Czech Republic.
Figure 1.
The number of sites in particular intervals for the four species that are now considered to be extinct in the Czech Republic.

Figure 2.
The blue columns are the number of sites that were occupied by orchids for the last time during the given interval and then no longer occupied for the five species that were found historically at 1 to 20 sites. The green bar indicates the number of sites observed as extant in the last interval, which we consider as extant currently.
Figure 2.
The blue columns are the number of sites that were occupied by orchids for the last time during the given interval and then no longer occupied for the five species that were found historically at 1 to 20 sites. The green bar indicates the number of sites observed as extant in the last interval, which we consider as extant currently.

Figure 3.
The blue columns are the number of sites that were occupied by orchids for the last time during the given interval and then no longer occupied for the 10 species that were found historically at 21 to 100 sites. The green bar indicates the number of sites occupied in the last interval, which we consider to be the current interval.
Figure 3.
The blue columns are the number of sites that were occupied by orchids for the last time during the given interval and then no longer occupied for the 10 species that were found historically at 21 to 100 sites. The green bar indicates the number of sites occupied in the last interval, which we consider to be the current interval.

Figure 4.
The number of sites that were occupied by orchids for the last time during the given interval and then were no longer occupied for the six species that were found historically at 101 to 300 sites. The green bar indicates the number of sites observed as extant in the last interval, which we consider as extant currently.
Figure 4.
The number of sites that were occupied by orchids for the last time during the given interval and then were no longer occupied for the six species that were found historically at 101 to 300 sites. The green bar indicates the number of sites observed as extant in the last interval, which we consider as extant currently.

Figure 5.
The number of sites that were occupied by orchids for the last time during the given interval and then were no longer occupied for the two species that were found historically at more than 300 sites. The green bar indicates the number of sites observed as extant in the last interval, which we consider as extant currently.
Figure 5.
The number of sites that were occupied by orchids for the last time during the given interval and then were no longer occupied for the two species that were found historically at more than 300 sites. The green bar indicates the number of sites observed as extant in the last interval, which we consider as extant currently.

Figure 6.
The blue columns are the number of sites that were occupied by orchids for the last time during the given interval and then were no longer occupied for the seven species of the genus Epipactis. The green bar indicates the number of sites occupied in the last interval, which we consider to be the current situation.
Figure 6.
The blue columns are the number of sites that were occupied by orchids for the last time during the given interval and then were no longer occupied for the seven species of the genus Epipactis. The green bar indicates the number of sites occupied in the last interval, which we consider to be the current situation.

Figure 7.
The highly threatened species of orchids in the Czech Republic with less than 50 sites recorded in the last period, ranked in order of the number of occupied sites in the last period. For each species, the percentage survival (orange bars in the figure) is the percentage of its sites included in the database that have survived until now (were found to be occupied during the last period). The orchid species circled in red are considered to be the most endangered species in the Czech flora (see text for explanation).
Figure 7.
The highly threatened species of orchids in the Czech Republic with less than 50 sites recorded in the last period, ranked in order of the number of occupied sites in the last period. For each species, the percentage survival (orange bars in the figure) is the percentage of its sites included in the database that have survived until now (were found to be occupied during the last period). The orchid species circled in red are considered to be the most endangered species in the Czech flora (see text for explanation).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Explanation of the selected time periods.
| Time Period | Explanation |
|---|---|
| Before 1900 | Very old data from the 19th century |
| 1901–1918 | Prior to the end of the 1st World War and the founding of Czechoslovakia |
| 1919–1938 | During the existence of the “1st” Czechoslovak Republic |
| 1939–1948 | 2nd World War and the period before the Communist Putsch in 1948 |
| 1949–1968 | Czechoslovakia under the Communist regime, before the Prague Spring |
| 1969–1989 | The period after the Soviet invasion and before the Velvet Revolution |
| 1990–2020 | The period after the Velvet Revolution and the free market economy in Czechoslovakia |
Table 2.
The total number of sites and their proportion occupied at the last interval for each orchid species analyzed in the Czech Republic. Species in red are considered to be extinct (category A1 in the national Red Lists) in the Czech Republic in 2020 according to available literature [40,41,53,54].
Table 2.
The total number of sites and their proportion occupied at the last interval for each orchid species analyzed in the Czech Republic. Species in red are considered to be extinct (category A1 in the national Red Lists) in the Czech Republic in 2020 according to available literature [40,41,53,54].
| Species | Total Number of Sites | Sites Occupied (%) | Species | Total Number of Sites | Sites Occupied (%) |
|---|---|---|---|---|---|
| Anacamptis coriophora | 21 | 0 | Goodyera repens | 231 | 15 |
| Anacamptis morio | 750 | 49 | Gymnadenia densiflora | 199 | 65 |
| Anacamptis palustris | 159 | 13 | Gymnadenia odoratissima | 15 | 47 |
| Anacamptis pyramidalis | 176 | 40 | Hammarbya paludosa | 42 | 36 |
| Dactylorhiza bohemica | 7 | 71 | Herminium monorchis | 12 | 0 |
| Dactylorhiza curvifolia | 6 | 17 | Himantglossum adriaticum | 24 | 25 |
| Dactylorhiza incarnata | 282 | 68 | Limodorum abortivum | 47 | 49 |
| Dactylorhiza maculata | 355 | 37 | Liparis loeselii | 87 | 53 |
| Dactylorhiza traunsteineri | 28 | 46 | Malaxis monophyllos | 124 | 45 |
| Epipactis futakii | 1 | 100 | Neotinea tridentata | 40 | 30 |
| Epipactis leptochila | 49 | 90 | Neotinea ustulata | 885 | 28 |
| Epipactis moravica | 11 | 100 | Neottia cordata | 350 | 34 |
| Epipactis nordeniorum | 10 | 100 | Ophrys apifera | 72 | 92 |
| Epipactis pontica | 25 | 88 | Ophrys holoserica | 120 | 41 |
| Epipactis pseudopurpurata | 12 | 100 | Ophrys insectifera | 89 | 42 |
| Epipactis talosii | 23 | 96 | Orchis mascula | 1483 | 89 |
| Epipogium aphyllum | 201 | 16 | Spiranthes spiralis | 237 | 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Štípková, Z.; Kindlmann, P. Orchid Extinction over the Last 150 Years in the Czech Republic. Diversity 2021, 13, 78. https://0-doi-org.brum.beds.ac.uk/10.3390/d13020078
AMA Style
Štípková Z, Kindlmann P. Orchid Extinction over the Last 150 Years in the Czech Republic. Diversity. 2021; 13(2):78. https://0-doi-org.brum.beds.ac.uk/10.3390/d13020078
Chicago/Turabian StyleŠtípková, Zuzana, and Pavel Kindlmann. 2021. "Orchid Extinction over the Last 150 Years in the Czech Republic" Diversity 13, no. 2: 78. https://0-doi-org.brum.beds.ac.uk/10.3390/d13020078
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.