
Detection of Target-Site Herbicide Resistance in the Common Ragweed: Nucleotide Polymorphism Genotyping by Targeted Amplicon Sequencing

 , , ,
, , , 
Abstract
:1. Introduction
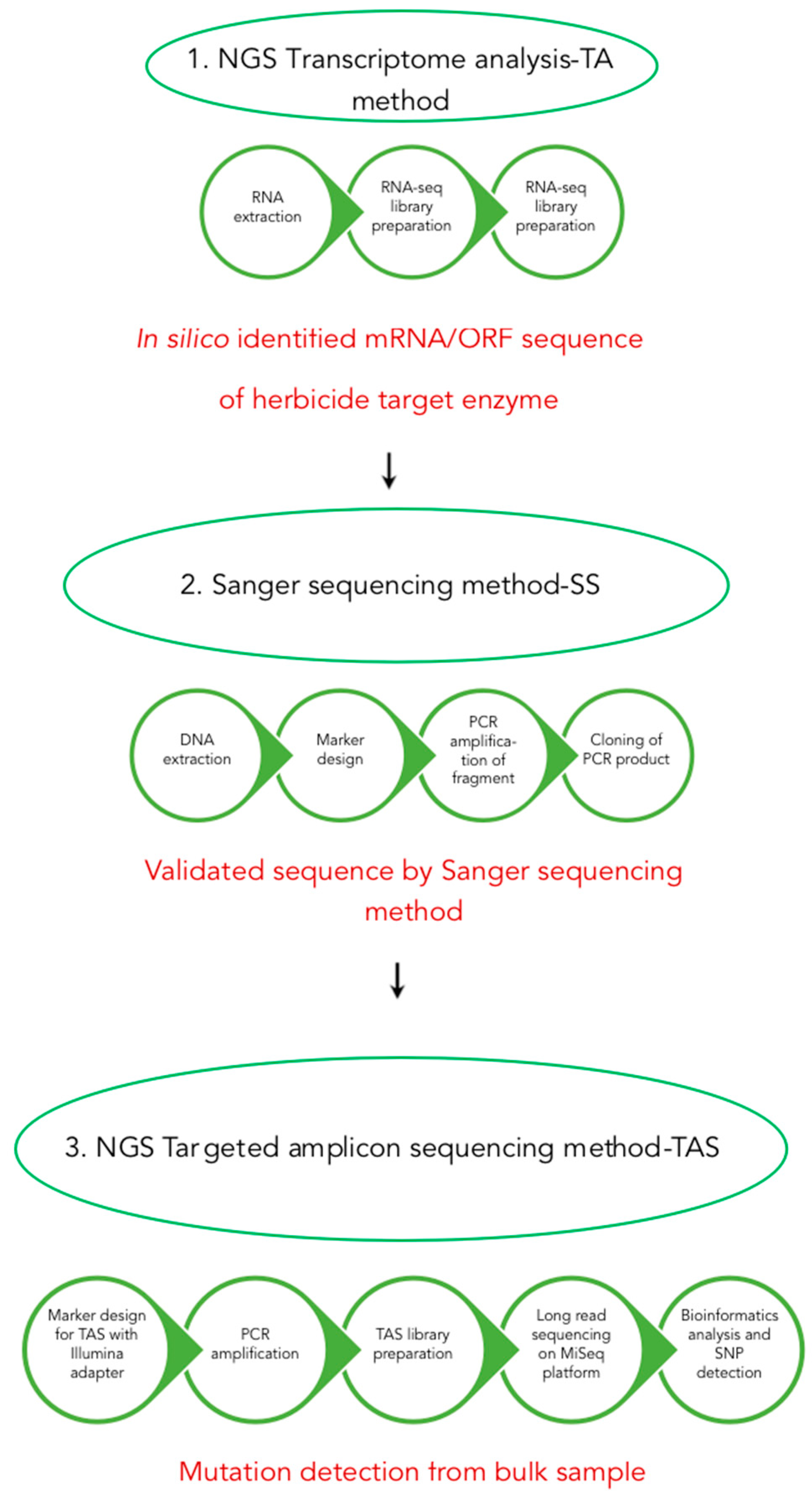
2. Materials and Methods
2.1. Plant Materials and DNA Extraction
2.2. Identification of A. artemisiifolia Herbicide Target Enzyme mRNAs
2.3. Intron Analysis, Amplification and Cloning of A. artemisiifolia Herbicide Target Enzyme Genes
2.4. PCR Amplification for NGS-TAS Experiments
2.5. NGS-TAS Experiments
2.6. Bioinformatics Analysis
3. Results
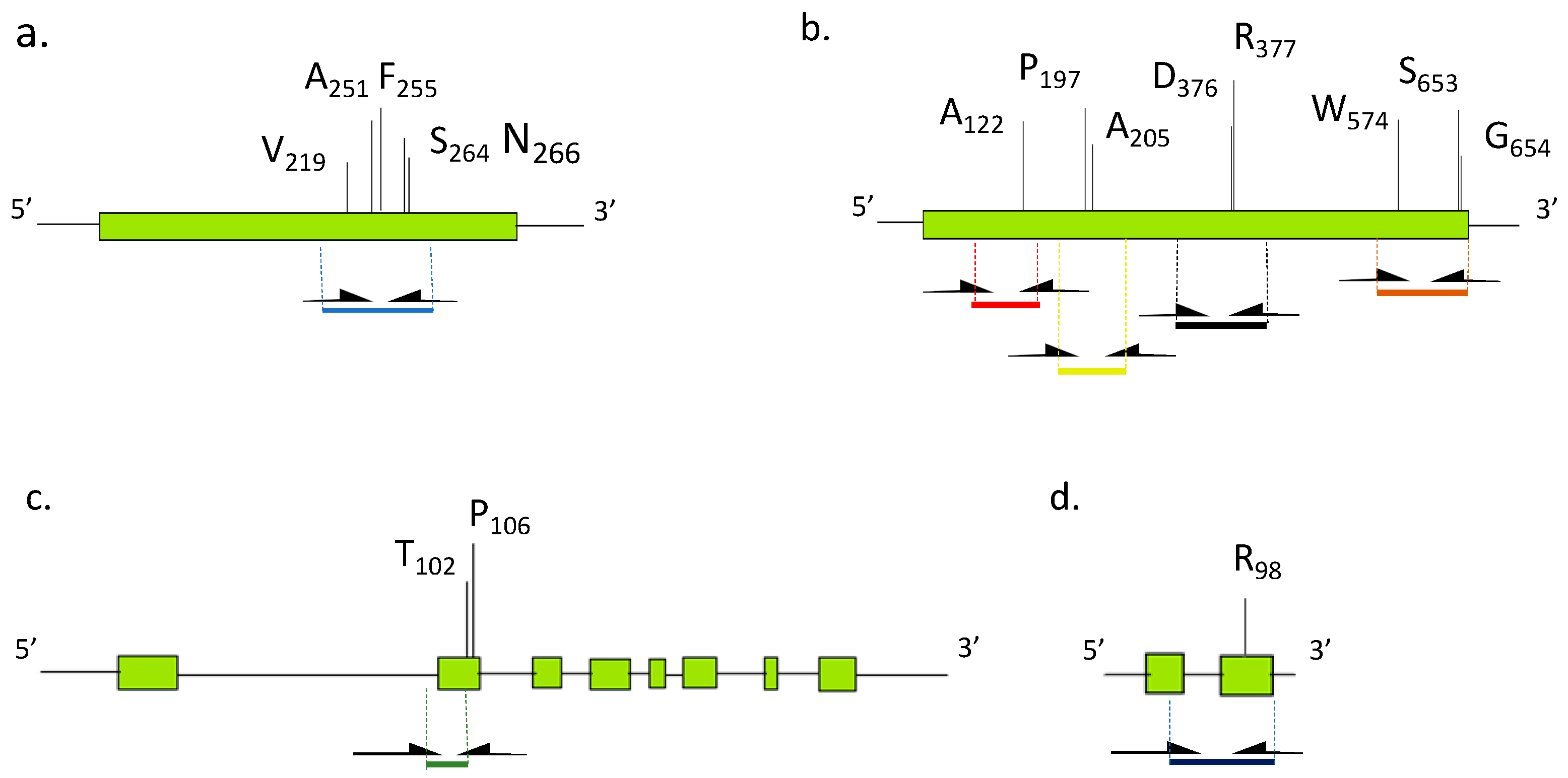
3.1. Identification of A. artemisiifolia Herbicide Target Enzyme cds and Genes
3.2. NGS-TAS Experiments of AMI and AMU Biological Repeats
3.3. Bioinformatics Analysis of AMI and AMU Groups
3.4. Detection of Mutation Points in Resistant Biotypes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baessler, C.; Klotz, S. Effects of changes in agricultural land-use on landscape structure and arable weed vegetation over the last 50 years. Agric. Ecosyst. Environ. 2006, 115, 43–50. [Google Scholar] [CrossRef]
- Lommen, S.T.E.; Hallmann, C.A.; Jongejans, E.; Chauvel, B.; Leitsch-Vitalos, M.; Aleksanyan, A.; Tóth, P.; Preda, C.; Šćepanović, M.; Onen, H.; et al. Explaining variability in the production of seed and allergenic pollen by invasive Ambrosia artemisiifolia across Europe. Biol. Invasions 2018, 20, 1475–1491. [Google Scholar] [CrossRef] [Green Version]
- Pinke, G.; Király, G.; Barina, Z.; Mesterházy, Á.; Balogh, L.; Csiky, J.; Schmotzer, A.; Molnár, A.V.; Pál, R.W. Assessment of endangered synanthropic plants of Hungary with special attention to arable weeds. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2011, 145, 426–435. [Google Scholar] [CrossRef]
- Alberternst, B.; Nawrath, S.; Starfinger, U. Biodiversity impacts of common ragweed. Julius-Kühn-Archiv 2016. [Google Scholar] [CrossRef]
- Essl, F.; Biró, K.; Brandes, D.; Broennimann, O.; Bullock, J.M.; Chapman, D.S.; Chauvel, B.; Dullinger, S.; Fumanal, B.; Guisan, A.; et al. Biological Flora of the British Isles: Ambrosia artemisiifolia. J. Ecol. 2015, 103, 1069–1098. [Google Scholar] [CrossRef] [Green Version]
- Fumanal, B.; Girod, C.; Fried, G.; Bretagnolle, F.; Chauvel, B. Can the large ecological amplitude of Ambrosia artemisiifolia explain its invasive success in France? Weed Res. 2008, 48, 349–359. [Google Scholar] [CrossRef]
- Bassett, I.J.; Crompton, C.W. The biology of Canadian weeds.: 11. Ambrosia artemisiifolia L. and A. psilostachya DC. Can. J. Plant Sci. 1975, 55, 463–476. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, J.; DiTommaso, A.; Zhang, C.; Zheng, G.; Liang, W.; Islam, F.; Yang, C.; Chen, X.; Zhou, W. Weed research status, challenges, and opportunities in China. Crop. Prot. 2020, 134, 104449. [Google Scholar] [CrossRef]
- Kiss, L.; Beres, I. Anthropogenic factors behind the recent population expansion of common ragweed (Ambrosia artemisiifolia L.) in Eastern Europe: Is there a correlation with political transitions? J. Biogeogr. 2006, 33, 2156–2157. [Google Scholar] [CrossRef]
- Csontos, P.; Vitalos, M.; Barina, Z.; Kiss, L. Early distribution and spread of Ambrosia artemisiifolia in Central and Eastern Europe. Bot. Helv. 2010, 120, 75–78. [Google Scholar] [CrossRef]
- Chauvel, B.; Dessaint, F.; Cardinal-Legrand, C.; Bretagnolle, F. The historical spread of Ambrosia artemisiifolia L. in France from herbarium records. J. Biogeogr. 2006, 33, 665–673. [Google Scholar] [CrossRef]
- Hamaoui-Laguel, L.; Vautard, R.; Liu, L.; Solmon, F.; Viovy, N.; Khvorostyanov, D.; Essl, F.; Chuine, I.; Colette, A.; Semenov, M.A.; et al. Effects of climate change and seed dispersal on airborne ragweed pollen loads in Europe. Nat. Clim. Chang. 2015, 5, 766–771. [Google Scholar] [CrossRef]
- Simard, M.-J.; Benoit, D.L. Distribution and abundance of an allergenic weed, common ragweed (Ambrosia artemisiifolia L.), in rural settings of southern Quebec, Canada. Can. J. Plant Sci. 2010, 90, 549–557. [Google Scholar] [CrossRef] [Green Version]
- Müller-Schärer, H.; Sun, Y.; Chauvel, B.; Karrer, G.; Kazinczi, G.; Kudsk, P.; Oude, A.L.; Schaffner, U.; Skjoth, C.; Smith, M.; et al. Cross-fertilizing weed science and plant invasion science to improve efficient management: A European challenge. Basic Appl. Ecol. 2018, 33, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Neve, P.; Barney, J.N.; Buckley, Y.; Cousens, R.D.; Graham, S.; Jordan, N.R.; Lawton-Rauh, A.; Liebman, M.; Mesgaran, M.B.; Schut, M.; et al. Reviewing research priorities in weed ecology, evolution and management: A horizon scan. Weed Res. 2018, 58, 250–258. [Google Scholar] [CrossRef] [Green Version]
- Westwood, J.H.; Charudattan, R.; Duke, S.O.; Fennimore, S.A.; Marrone, P.; Slaughter, D.C.; Swanton, C.; Zollinger, R. Weed Management in 2050: Perspectives on the Future of Weed Science. Weed Sci. 2018, 66, 275–285. [Google Scholar] [CrossRef] [Green Version]
- Moss, S.R.; Storkey, J.; Cussans, J.W.; Perryman, S.A.M.; Hewitt, M.V. Symposium the Broadbalk long-term experiment at Rothamsted: What has it told us about weeds? Weed Sci. 2004, 52, 864–873. [Google Scholar] [CrossRef]
- Coble, H.D. Future Directions and Needs for Weed Science Research. Weed Technol. 1994, 8, 410–412. [Google Scholar] [CrossRef]
- Kazinczi, G.; Novák, R.; Pathy, Z.; Béres, I. Common ragweed (Ambrosia artemisiifolia L.): A review with special regards to the results in Hungary. III. Resistant biotypes, control methods and authority arrangements. Herbologia 2008, 9, 119–144. [Google Scholar]
- Powles, S.B.; Yu, Q. Evolution in Action: Plants Resistant to Herbicides. Annu. Rev. Plant Biol. 2010, 61, 317–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harker, K.N.; O’Donovan, J.T. Recent Weed Control, Weed Management, and Integrated Weed Management. Weed Technol. 2013, 27, 1–11. [Google Scholar] [CrossRef]
- Délye, C.; Jasieniuk, M.; Le Corre, V. Deciphering the evolution of herbicide resistance in weeds. Trends Genet. 2013, 29, 649–658. [Google Scholar] [CrossRef]
- Vila-Aiub, M.M.; Neve, P.; Powles, S.B. Fitness costs associated with evolved herbicide resistance alleles in plants. New Phytol. 2009, 184, 751–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powles, S.B.; Preston, C. Evolved Glyphosate Resistance in Plants: Biochemical and Genetic Basis of Resistance. Weed Technol. 2006, 20, 282–289. [Google Scholar] [CrossRef]
- Bernasconi, P.; Woodworth, A.R.; Rosen, B.A.; Subramanian, M.V.; Siehl, D.L. A Naturally Occurring Point Mutation Confers Broad Range Tolerance to Herbicides That Target Acetolactate Synthase. J. Biol. Chem. 1995, 270, 17381–17385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, H.; Yu, Q.; Purba, E.; Li, M.; Walsh, M.; Friesen, S.; Powles, S.B. A novel amino acid substitution Ala-122-Tyr in ALS confers high-level and broad resistance across ALS-inhibiting herbicides. Pest Manag. Sci. 2012, 68, 1164–1170. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, M.; Gao, X.; Fang, F. A novel amino acid substitution Trp574Arg in acetolactate synthase (ALS) confers broad resistance to ALS-inhibiting herbicides in crabgrass (Digitaria sanguinalis). Pest Manag. Sci. 2017, 73, 2538–2543. [Google Scholar] [CrossRef]
- McNaughton, K.E.; Letarte, J.; Lee, E.A.; Tardif, F.J. Mutations inALSconfer herbicide resistance in redroot pigweed (Amaranthus retroflexus) and Powell amaranth (Amaranthus powellii). Weed Sci. 2005, 53, 17–22. [Google Scholar] [CrossRef]
- Riar, D.S.; Norsworthy, J.K.; Srivastava, V.; Nandula, V.; Bond, J.A.; Scott, R.C. Physiological and Molecular Basis of Acetolactate Synthase-Inhibiting Herbicide Resistance in Barnyardgrass (Echinochloa crus-galli). J. Agric. Food Chem. 2013, 61, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Warwick, S.I.; Xu, R.; Sauder, C.; Beckie, H.J. Acetolactate Synthase Target-Site Mutations and Single Nucleotide Polymorphism Genotyping in ALS-Resistant Kochia (Kochia scoparia). Weed Sci. 2008, 56, 797–806. [Google Scholar] [CrossRef]
- Yu, Q.; Jalaludin, A.; Han, H.; Chen, M.; Sammons, R.D.; Powles, S.B. Evolution of a Double Amino Acid Substitution in the 5-Enolpyruvylshikimate-3-Phosphate Synthase in Eleusine indica Conferring High-Level Glyphosate Resistance. Plant Physiol. 2015, 167, 1440–1447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brosnan, J.T.; Vargas, J.J.; Breeden, G.K.; Grier, L.; Aponte, R.A.; Tresch, S.; Laforest, M. A new amino acid substitution (Ala-205-Phe) in acetolactate synthase (ALS) confers broad spectrum resistance to ALS-inhibiting herbicides. Planta 2016, 243, 149–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diebold, R.S.; McNaughton, K.E.; Lee, E.A.; Tardif, F.J. Multiple resistance to imazethapyr and atrazine in Powell amaranth (Amaranthus powellii). Weed Sci. 2003, 51, 312–318. [Google Scholar] [CrossRef]
- Liu, M.; Hulting, A.G.; A Mallory-Smith, C. Characterization of multiple-herbicide-resistant Italian ryegrass (Lolium perenne spp. multiflorum). Pest Manag. Sci. 2014, 70, 1145–1150. [Google Scholar] [CrossRef]
- Stephenson, G.R.; Dykstra, M.D.; McLaren, R.D.; Hamill, A.S. Agronomic Practices Influencing Triazine-Resistant Weed Distribution in Ontario. Weed Technol. 1990, 4, 199–207. [Google Scholar] [CrossRef]
- Alfonso, M.; Pueyo, J.J.; Gaddour, K.; Etienne, A.L.; Kirilovsky, D.; Picorel, R. Induced New Mutation of D1 Serine-268 in Soybean Photosynthetic Cell Cultures Produced Atrazine Resistance, Increased Stability of S2QB- and S3QB-States, and Increased Sensitivity to Light Stress. Plant Physiol. 1996, 112, 1499–1508. [Google Scholar] [CrossRef] [Green Version]
- Cseh, A.; Cernák, I.; Taller, J. Molecular characterization of atrazine resistance in common ragweed (Ambrosia artemisiifolia L.). J. Appl. Genet. 2009, 50, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Mátyás, K.K.; Taller, J.; Cseh, A.; Poczai, P.; Cernák, I. Development of a simple PCR-based assay for the identification of triazine resistance in the noxious plant common ragweed (Ambrosia artemisiifolia) and its applicability in higher plants. Biotechnol. Lett. 2011, 33, 2509–2515. [Google Scholar] [CrossRef] [PubMed]
- Saint-Louis, S.; DiTommaso, A.; Watson, A.K. A Common Ragweed (Ambrosia artemisiifolia) Biotype in Southwestern Québec Resistant to Linuron. Weed Technol. 2005, 19, 737–743. [Google Scholar] [CrossRef]
- Beckie, H.J.; Hall, L.; Tardif, F.; Blackshaw, R. Herbicide resistance in Canada—Where are we today. In Integrated Weed Management: Explore the Potential; Expert Committee on Weeds: Sainte-Anne-de-Bellevue, QC, Canada, 2001; pp. 1–36. [Google Scholar]
- Simard, M.-J.; Laforest, M.; Soufiane, B.; Benoit, D.L.; Tardif, F. Linuron resistant common ragweed (Ambrosia artemisiifolia) populations in Québec carrot fields: Presence and distribution of Target site and non-target site resistant biotypes. Can. J. Plant Sci. 2017, 98, 345–352. [Google Scholar] [CrossRef]
- Beckie, H.J.; Tardif, F.J. Herbicide cross resistance in weeds. Crop. Prot. 2012, 35, 15–28. [Google Scholar] [CrossRef]
- Duggleby, R.G.; Pang, S.S. Acetohydroxyacid synthase. J. Biochem. Mol. Biol. 2000, 33, 1–36. [Google Scholar]
- Tranel, P.J.; Wright, T.R. Resistance of weeds to ALS-inhibiting herbicides: What have we learned? Weed Sci. 2002, 50, 700–712. [Google Scholar] [CrossRef]
- Yu, Q.; Powles, S.B. Resistance to AHAS inhibitor herbicides: Current understanding. Pest Manag. Sci. 2014, 70, 1340–1350. [Google Scholar] [CrossRef] [PubMed]
- Heap, I. The International Survey of Herbicide Resistant Weeds. 2021. Available online: www.weedscience.org (accessed on 10 February 2021).
- Patzoldt, W.L.; Tranel, P.J.; Alexander, A.L.; Schmitzer, P.R. A common ragweed population resistant to cloransulam-methyl. Weed Sci. 2001, 49, 485–490. [Google Scholar] [CrossRef]
- Massa, D.; Krenz, B.; Gerhards, R. Target-site resistance to ALS-inhibiting herbicides in Apera spica-venti populations is conferred by documented and previously unknown mutations. Weed Res. 2011, 51, 294–303. [Google Scholar] [CrossRef]
- Ng, C.H.; Wickneswari, R.; Salmijah, S.; Teng, Y.T.; Ismail, B.S. Gene polymorphisms in glyphosate-resistant and -susceptible biotypes of Eleusine indica from Malaysia. Weed Res. 2003, 43, 108–115. [Google Scholar] [CrossRef]
- Lee, L.J.; Ngim, J. A first report of glyphosate-resistant goosegrass (Eleusine indica (L) Gaertn) in Malaysia. Pest Manag. Sci. Former. Pestic. Sci. 2000, 56, 336–339. [Google Scholar] [CrossRef]
- Baerson, S.R.; Rodriguez, D.J.; Tran, M.; Feng, Y.; Biest, N.A.; Dill, G.M. Glyphosate-Resistant Goosegrass. Identification of a Mutation in the Target Enzyme 5-Enolpyruvylshikimate-3-Phosphate Synthase. Plant Physiol. 2002, 129, 1265–1275. [Google Scholar] [CrossRef] [Green Version]
- Perotti, V.E.; Larran, A.S.; E Palmieri, V.; Martinatto, A.K.; E Alvarez, C.; Tuesca, D.; Permingeat, H.R. A novel triple amino acid substitution in the EPSPS found in a high-level glyphosate-resistant Amaranthus hybridus population from Argentina. Pest Manag. Sci. 2019, 75, 1242–1251. [Google Scholar] [CrossRef]
- Brewer, C.E.; Oliver, L.R. Confirmation and Resistance Mechanisms in Glyphosate-Resistant Common Ragweed (Ambrosia artemisiifolia) in Arkansas. Weed Sci. 2009, 57, 567–573. [Google Scholar] [CrossRef]
- Smith, A.G.; Marsh, O.; Elder, G.H. Investigation of the subcellular location of the tetrapyrrole-biosynthesis enzyme coproporphyrinogen oxidase in higher plants. Biochem. J. 1993, 292, 503–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lermontova, I.; Kruse, E.; Mock, H.-P.; Grimm, B. Cloning and characterization of a plastidal and a mitochondrial isoform of tobacco protoporphyrinogen IX oxidase. Proc. Natl. Acad. Sci. USA 1997, 94, 8895–8900. [Google Scholar] [CrossRef] [Green Version]
- Patzoldt, W.L.; Hager, A.G.; McCormick, J.S.; Tranel, P.J. A codon deletion confers resistance to herbicides inhibiting protoporphyrinogen oxidase. Proc. Natl. Acad. Sci. USA 2006, 103, 12329–12334. [Google Scholar] [CrossRef] [Green Version]
- Rousonelos, S.L.; Lee, R.M.; Moreira, M.S.; Vangessel, M.J.; Tranel, P.J. Characterization of a Common Ragweed (Ambrosia artemisiifolia) Population Resistant to ALS- and PPO-Inhibiting Herbicides. Weed Sci. 2012, 60, 335–344. [Google Scholar] [CrossRef]
- Shirasawa, K.; Kuwata, C.; Watanabe, M.; Fukami, M.; Hirakawa, H.; Isobe, S. Target Amplicon Sequencing for Genotyping Genome-Wide Single Nucleotide Polymorphisms Identified by Whole-Genome Resequencing in Peanut. Plant Genome 2016, 9, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Virág, E.; Hegedűs, G.; Barta, E.; Nagy, E.; Mátyás, K.; Kolics, B.; Taller, J. Illumina Sequencing of Common (Short) Ragweed (Ambrosia artemisiifolia L.) Reproductive Organs and Leaves. Front. Plant Sci. 2016, 7, 1506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Untergasser, A.; Nijveen, H.; Rao, X.; Bisseling, T.; Geurts, R.; Leunissen, J.A.M. Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Res. 2007, 35, W71–W74. [Google Scholar] [CrossRef] [Green Version]
- Gaines, T.A.; Wright, A.A.; Molin, W.T.; Lorentz, L.; Riggins, C.W.; Tranel, P.J.; Beffa, R.; Westra, P.; Powles, S.B. Identification of Genetic Elements Associated with EPSPS Gene Amplification. PLoS ONE 2013, 8, e65819. [Google Scholar] [CrossRef] [Green Version]
- Marcus, L.; Hartnett, J.; Storts, D.R. The pGEM®-T and pGEM®-T Easy Vector Systems. Promega Notes Mag. 1996, 58, 36. [Google Scholar]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rombel, I.T.; Sykes, K.F.; Rayner, S.; Johnston, S.A. ORF-FINDER: A vector for high-throughput gene identification. Gene 2002, 282, 33–41. [Google Scholar] [CrossRef]
- Kniss, A.R. Genetically Engineered Herbicide-Resistant Crops and Herbicide-Resistant Weed Evolution in the United States. Weed Sci. 2017, 66, 260–273. [Google Scholar] [CrossRef] [Green Version]
- Gaines, T.A.; Zhang, W.; Wang, D.; Bukun, B.; Chisholm, S.T.; Shaner, D.L.; Nissen, S.J.; Patzoldt, W.L.; Tranel, P.J.; Culpepper, A.S.; et al. Gene amplification confers glyphosate resistance in Amaranthus palmeri. Proc. Natl. Acad. Sci. USA 2009, 107, 1029–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajwa, A.A.; Mahajan, G.; Chauhan, B.S. Nonconventional Weed Management Strategies for Modern Agriculture. Weed Sci. 2015, 63, 723–747. [Google Scholar] [CrossRef]
- Pearson, D.E.; Ortega, Y.K.; Runyon, J.B.; Butler, J.L. Secondary invasion: The bane of weed management. Biol. Conserv. 2016, 197, 8–17. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Gene Name | cds | Complete Gene | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Length (bp) | Lenght (aa) | Similarity (%) | Reference ID NCBI GeneBank | Contig ID NCBI GeneBank GEZL01000001 | NCBI Accession | GC Content (%) | Length (bp) | Intron Number | Intron Position from…to | NCBI Accession | GC Content (%) | |
| psbA | 1062 | 353 | 99% | A. artemisiifolia AB427162.1 | TR92155|c0_g1_i1 | MT425203 | 41.7 | 1062 | - | - | MT879746 | 41 |
| ahas | 1965 | 654 | 95% | Xanthium sp. AAA74913.1 | TR49503|c0_g3_i1 TR49503|c0_g8_i1 | MK096760 | 48.1 | 1965 | - | - | MT415954 | 48.3 |
| epsps | 1539 | 512 | 96% | Helianthus annuus XP_022017499.1 | TR44247|c0_g1_i1 | MK096765 | 45 | 3539 | 7 | 316…1311 1557…1832 1987…2114 2330…2410 2529…2656 2868…3006 3069…3400 | MT415955 MT409110 | 34 37.5 |
| ppx2 | 1476 | 491 | 95% | Helianthus annuus XP_021982414.1 | TR33881|c0_g1_i1 | MK096762 | 42.8 | Partial 325 | 1 | 56–276 | MT879747 MT879748 | 34.3 33.1 |
| Fragments | Length of Fragment with Adapter (bp) | AMI | AMU | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 Sample | 2 Sample | 3 Sample | 1 Control Sample | 1 Sample | 2 Sample | 3 Sample | 1 Control Sample | ||||||||||
| cc | Number of Contig | cc | Number of Contig | cc | Number of Contig | cc | Number of Contig | cc | Number of Contig | cc | Number of Contig | cc | Number of Contig | cc | Number of Reads | ||
| psbA | 377 | 50.3 | 26,989 | 32.8 | 34,594 | 40.8 | 30,560 | 72.1 | 32,592 | 24.3 | 28,106 | 19.1 | 28,523 | 19.9 | 30,492 | 48.3 | 40,178 |
| ahas 1 | 288 | 10,009 | 25,817 | 11,802 | 60.3 | 9129 | 8066 | 21,401 | 12,709 | 24.3 | 11,426 | ||||||
| ahas 2 | 273 | 45,613 | 32,641 | 45,887 | 50.3 | 34,158 | 43,411 | 47,525 | 50,635 | 43.4 | 52,289 | ||||||
| ahas 3 | 319 | 18,488 | 15,106 | 23,639 | 74.3 | 20,210 | 25,580 | 21,738 | 19,612 | 37 | 23,140 | ||||||
| ahas 4 | 453 | 12,563 | 13,668 | 11,955 | 88.6 | 17,567 | 13,133 | 15,549 | 15,854 | 37.1 | 12,032 | ||||||
| epsps | 310 | 21,543 | 34,053 | 29,779 | 51.9 | 16,599 | 18,103 | 10,839 | 17,476 | 31.7 | 15,737 | ||||||
| ppx2 | 390 | 11,580 | 9280 | 26,659 | 58.8 | 10,546 | 15,205 | 10,108 | 9346 | 37.5 | 15,583 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kutasy, B.; Farkas, Z.; Kolics, B.; Decsi, K.; Hegedűs, G.; Kovács, J.; Taller, J.; Tóth, Z.; Kálmán, N.; Kazinczi, G.; et al. Detection of Target-Site Herbicide Resistance in the Common Ragweed: Nucleotide Polymorphism Genotyping by Targeted Amplicon Sequencing. Diversity 2021, 13, 118. https://0-doi-org.brum.beds.ac.uk/10.3390/d13030118
Kutasy B, Farkas Z, Kolics B, Decsi K, Hegedűs G, Kovács J, Taller J, Tóth Z, Kálmán N, Kazinczi G, et al. Detection of Target-Site Herbicide Resistance in the Common Ragweed: Nucleotide Polymorphism Genotyping by Targeted Amplicon Sequencing. Diversity. 2021; 13(3):118. https://0-doi-org.brum.beds.ac.uk/10.3390/d13030118
Chicago/Turabian StyleKutasy, Barbara, Zoltán Farkas, Balázs Kolics, Kincső Decsi, Géza Hegedűs, Judit Kovács, János Taller, Zoltán Tóth, Nikoletta Kálmán, Gabriella Kazinczi, and et al. 2021. "Detection of Target-Site Herbicide Resistance in the Common Ragweed: Nucleotide Polymorphism Genotyping by Targeted Amplicon Sequencing" Diversity 13, no. 3: 118. https://0-doi-org.brum.beds.ac.uk/10.3390/d13030118