
Increasing Survival of Wild Macaw Chicks Using Foster Parents and Supplemental Feeding


1
The Macaw Society-Sociedad Pro Guacamayos, Department of Ecology and Conservation Biology, Texas A&M University, TAMU 4467, College Station, TX 77843, USA
2
Wildlife Conservation Society–Guatemala, Avenida 15 de Marzo, Casa No. 3. Flores, Petén CP 17001, Guatemala
3
The Macaw Society-Sociedad Pro Guacamayos, Schubot Center for Avian Health, Department of Veterinary Pathobiology, Texas A&M University, TAMU 4467, College Station, TX 77843, USA
*
Author to whom correspondence should be addressed.
†
Current affiliation: Schubot Center for Avian Health, TAMU 4457, College Station, TX 77843, USA.
Diversity 2021, 13(3), 121; https://0-doi-org.brum.beds.ac.uk/10.3390/d13030121
Submission received: 31 December 2020
/
Revised: 2 March 2021
/
Accepted: 3 March 2021
/
Published: 12 March 2021
(This article belongs to the Special Issue Ecology and Conservation of Parrots in Their Native and Non-Native Ranges)
Abstract
:The use of foster parents has great potential to help the recovery of highly endangered bird species. However, few studies have shown how to successfully use these techniques in wild populations. Scarlet Macaws (Ara macao macao) in Perú hatch 2–4 chicks per nest but about 24% of all chicks die of starvation and on average just 1.4 of them fledge per successful nest. In this study we develop and test new techniques to increase survival of wild Scarlet Macaw chicks by reducing chick starvation. We hypothesized that using foster parents would increase the survival of chicks at risk of starvation and increase overall reproductive success. Our results show that all relocated macaw chicks were successfully accepted by their foster parents (n = 28 chicks over 3 consecutive breeding seasons) and 89% of the relocated chicks fledged. Overall, we increased fledging success per available nest from 17% (2000 to 2016 average) to 25% (2017 to 2019) and decreased chick death by starvation from 19% to 4%. These findings show that the macaw foster parents technique and post relocation supplemental feeding provide a promising management tool to aid wild parrot population recovery in areas with low reproductive success.
1. Introduction
The use of foster parents in avian population management is a technique with great potential to aid in the recovery of highly endangered species in the wild [1]. Foster parenting, the use of breeding pairs to raise young that were not part of their -own broods, is a well-known avicultural technique that has been intensively used in captive breeding and reintroduction programs over several decades [2] and also in conservation captive breeding programs to increase reproduction [1,3,4,5]. However, few studies have systematically studied how to successfully use this tool in the wild.
The topic of increasing productivity in parrots for conservation is not new. In the early 1990s’, it was suggested that managing intensively the factors that limit species’ population growth was the key to productivity maximization [6]. One of the techniques proposed at the time was to increase fledging success [7]. In psittacines, the majority of species hatch their eggs asynchronously over a period of 1 to 14 days [5,8,9,10,11,12,13,14] which results in a size-based hierarchy among brood members [9,15,16] which often leads to the death of younger chicks [8,9,13,14,17]. In this scenario, decreasing hatching asynchrony has been proposed as a potential management tool to increase numbers of young for harvesting for conservation purposes [7]. These harvested last and penultimate chicks could be relocated in foster nests to increase overall reproductive output. This technique has great potential for in situ conservation efforts because there is strong evidence that psittacines can be successfully used as foster parents and they are able to raise and fledge additional chicks (RG-A unpublished data [18]).
Chick fostering has been successfully used in commercial aviculture to raise finches (Lonchura ssp. and others) and with captive psittacines of the genera Cyclopsitta, Alisterus, Amazona, Pionus and Cacatua [4,5,8,19,20], mainly as an emergency tool when chicks were rejected by parents or fell out of the nest [19,20,21]. It has also been used in captive breeding programs for psittacines [4,5]. In the wild, it has proven to be useful for recovering the Puerto Rican Amazon (Amazona vittata) and increasing population recruitment in the Yellow-shouldered Parrot (Amazona barbadensis) in Venezuela [18]. It has also been used in the wild as a tool to study parent/offspring interactions in Crimson Rosellas (Platycercus elegan) [22] and Galahs (Eolophus roseicapillus) [8] in Australia. Most recently, it has been used by RG-A and collaborators from the Wildlife Conservation Society (WCS)-Guatemala as part of the efforts to recover Scarlet Macaw (Ara macao cyanoptera) populations around Laguna del Tigre National Park.
The Scarlet Macaw (Ara macao), one of the most iconic members of the Psittacidae family and an important flagship species of the tropical forest, is widely distributed in the Americas from Southeastern Mexico to Peru and Bolivia [23]. However, most populations in Central America are currently declining due to a combination of habitat loss and poaching for the local pet trade [14,24,25,26,27,28]. As with many other members of the family Psittacidae, the species shows brood reduction by chick starvation [11,14]. This starvation can result in the death of >22% of all hatched chicks and is the most common cause of chick death [15]. In Tambopata, Peru, an area with no nest poaching, clutches have on average three eggs, resulting in broods of about two chicks but just a mean of 1.4 chicks fledge per nest per season [29]. Overall, 27% of second chicks and all third and fourth chicks die by starvation, which results in a substantial loss of hatchlings. In areas where Scarlet Macaw populations are declining, valuable chicks that could help increase population numbers starve to death. Increasing survival of those starving chicks could provide significant numbers of young that can directly increase wild populations.
There is little information published on Scarlet Macaws as foster parents. In the late 1990s’ in Carara National Park in Costa Rica one chick rescued from poachers was placed in a wild nest that had just one chick and both chicks fledged [11]. There are also reports from captivity where a Scarlet Macaw pair was used as a surrogate to raise chicks of the Blue-headed Macaw (Primolious couloni [30]). The most comprehensive information related to Scarlet Macaws as foster parents in the wild comes from Guatemala where the technique has been used to place 60 chicks during seven breeding seasons since 2011 [RG-A unpublished data]. In this case, foster chicks averaged 41 days old (range 12–85 days old) and 78% of them were successfully adopted and fledged [RG-A unpublished data]. The technique was used when: (1) chicks did not gain weight as expected, (2) third and fourth chicks hatched, (3) chicks lost their parents, (4) chicks hatched in the field station after eggs were rescued from nest poaching) [RG-A unpublished data].
The use of Scarlet Macaws as foster parents in the wild offers a good system not only to test the technique in situ, but also to test the main drivers of chick death by starvation. The main driver behind death by starvation appears to be brood members’ age differences: first chicks apparently do not die of starvation and the chance of younger chicks starving is directly proportional to age difference in relation to the first chick of the brood [29]. In the case of second chicks, when the age difference was 3 to 4 days the probability of death was 24% but if this difference was 5 days or more, the probability of death by starvation jumped to 80% [29]. If age difference among brood members is the main reason why the younger member of the brood starved to death [29], age differences in foster broods would need to be less than 5 days to ensure that none of the brood members would perish.
The main objective of this experiment was to develop and test techniques to increase survival of Scarlet Macaw chicks in the wild by reducing chick starvation using wild foster parents. In addition, we wanted to test if the age difference among brood members was the sole driver of chick death by starvation. To do this, we tested the following main hypotheses: (1) Wild Scarlet Macaws accept chicks that are not their offspring and raise them to fledging, (2) Using foster parents increases the survival of chicks at risk of starvation and increases the overall population reproductive success, and (3) Age differences among brood members is not the only driver of chick death by starvation and (4) Wild Scarlet Macaws are able of fledge a brood of three chicks.
2. Materials and Methods
This research was conducted in the forests surrounding the Tambopata Research Center (13°8′ S, 69°36′ W), located in the Tambopata National Reserve (275,000 ha) adjacent to the Bahuaja-Sonene National Park (1,091,416) in the department of Madre de Dios, southeastern Perú. The forest adjacent to the research station is classified as tropical moist forest (Holdridge life zones system) and is a combination of flood plain, terra firme, successional, and palm swamp forests that receives around 3200 mm of rain annually [31,32].
2.1. Background Methodology
We conducted this research from October 2016 to March 2019, during three consecutive macaw breeding seasons, as part of a program of investigation on Scarlet Macaw breeding ecology, nesting behavior, and health run by The Macaw Society -Sociedad Pro Guacamayos (www.TheMacawSociety.org and http://vetmed.tamu.edu/macawproject, accessed on 1 October 2016) [29,33,34,35,36]. This program has been monitoring macaw nests intensively since 1999. Macaw breeding season is from mid-October to mid-April, annually. Each season we monitored about 40 macaw nests (16 natural, 24 artificial) in a 5 km radius area, using single rope climbing systems [37,38]. Artificial nests were a combination of wooden boxes and PVC pipes (16” diameter) and were hung one per tree [See [33] for a detail explanation about artificial nest used] (Figure 1). Eight artificial nests in a 3 km radius were equipped with video surveillance cameras each season. Due to the high humidity of the rainforest, video systems frequently suffered intermittent malfunctions that alter sample sizes for data reported from nest videos. Not all nests with video systems received foster chicks. All nests were checked once every 2–3 days until the first egg was found. After an egg was found, nest monitoring ceased until 26 days later and continued daily until all viable eggs had hatched. Due to this frequent monitoring, done over almost two decades, nesting macaws were habituated to human intervention and did not flee or abandon nests during nest checks. However, nesting macaws still displayed a few disturbance behaviors in the presence of climbers (i.e., alarm calls and calls they use when fighting with other macaws). They also showed aggressive behaviors towards climbers (i.e., lunging at climbers from inside nest, flying at and even hitting the climber, etc.). Despite this high level of acclimatization of the birds, we did not weigh, measure or manipulate broods until second chick hatched in each brood in order to reduce disturbance at the nest and maximize hatching success.
2.2. Chick Relocation Procedures
2.2.1. Criteria and Timing to Remove Chicks for Relocation Procedures
A total of 32 macaw chicks were removed from their original nests. Four of them perished in our nursery. Two of them died the same day they hatched probably due to the fact that they hatched underweight. Two other chicks died at <5 days old, probably due to slow digestion problems. Macaw chicks were removed from their original nests according to the criteria and timing shown in Table 1.
2.2.2. Removed Chicks’ Initial Conditions
The majority of the chicks (67%) were healthy when removed from the nest. Eleven first chicks and nine third chicks weighed as expected for their ages (see Vigo et al. [14]). Two third chicks arrived underweight (17.9 g each), one with early signs of dehydration and the other one apparently in good condition. Three second chicks were brought in as soon as they did not gain weight as expected but were still in the weight range for their ages. One of the second chicks arrived underweight for its age, showing signs of starvation (empty crop, grayish color, dry skin and prominent ocular area). Another second chick that had a congenital foot malformation was brought in as soon as it did not gain weight as expected for its age (2 days of age). The three chicks showing signs of botfly related infection did not have botflies when arrived to the nursery because they were removed in the field. Both fourth chicks arrived underweight. One arrived right after hatching at 17.1 g. The other one, was left in its original nest with a sibling 9 days older, and it was removed at age 5 days when it started to showed early signs of starvation. We removed 32 chicks in total (Season 1 = 5 chicks, season 2 = 11 chicks, season 3 = 16 chicks, Table 1).
2.2.3. Macaw Chick Rearing in the Nursery
Chicks were kept in boxes (40 cm × 40 cm × 50 cm) with three solid wood sides, a solid wood base, and a wire mesh front and top. We used a Brinsea EcoGlow Brooder as a source of heat for the chicks. Heat and humidity in each box were monitored with an off the shelf digital thermometer-hygrometer. Chicks were separated in two different boxes according to their age. Chicks under 15 days old were kept in one box, each in a separate plastic cup on wood chips and a soft piece of cloth. Chicks over 15 days old were kept together in a separate box with a woodchip substrate and a 1/2” square mesh tray on top of substrate. Chicks were maintained at age-appropriate temperature and humidity conditions following the recommendations in Voren and Jordan [39]. In general, chicks were syringe fed Zupreem Embrace baby bird hand feeding formula prepared following the age-specific manufacture recommendations (https://www.zupreem.com/products/birds/embrace-plus/ accessed on 1 October 2016). Chicks that came in sick, weak or underweight were given custom diets used commonly in commercial aviculture (Table A1). All chicks that arrived showing signs of botfly related infection (n = 4) were treated with oral antibiotics and/or local antibiotic cream.
2.2.4. Criteria to Assign Macaw Chicks to Wild Macaw Nests
2.2.5. Criteria to Select Foster Parents
We used the literature [18,40] and our previous knowledge of the species to create selection criteria for pairs to host foster chicks. We preferentially chose pairs with the following characteristics: (1) Pairs nesting in artificial nests; (2) Known pairs with banded individuals older than 8 years old; (3) Pairs with at least one chick that fledged in a previous season; (4) Pairs with no history of chick death by unknown causes; (5) Pairs with no history of chick death due to poor parental care in solo chicks, such as hypothermia or low daily feeding rates; and (6) Pairs with no records suggesting they have little breeding experience, such as slow growth/poor body condition in chicks. Due to a lack of suitable nesting pairs, one foster chick was placed with a nesting pair with an unknown breeding history. In total, 12 macaw pairs were used as foster parents. In 10 of them at least one individual of the pair was banded. Adult macaw genders were determined either by DNA testing of at least one individual of the pair (67% of total macaw foster pairs) or by comparing nesting behavior to known gender macaw pairs (33% of total macaw foster pairs). Seven macaw pairs were used as foster parents in more than one season and one was used in all three seasons of this experiment.
2.2.6. Foster Chick Relocation Procedures and Timing
We conducted four complementary macaw chick relocation procedures during this study. Each one was designed to test a different hypothesis related the capacity of wild macaws to foster chicks in wild conditions (Table 3). A chick was considered relocated when it was taken out of its original nest and moved to another nest, either the same day or days after. Due to the fact that we were working in natural conditions in the wild, we had little ability to set up identical conditions in each specific foster nest case. Foster nest candidates for procedures 1, 2 and 4 were checked every day from hatch to ensure nest requirements were maintained. In these procedures, the key moment to place foster chicks in foster nests was when the younger chick opened its eyes. By having both chicks with open eyes, all chicks were able to see the parents and effectively beg for food in a similar fashion. Foster nests for procedure 3 were chosen in an opportunistic way as long as foster parents fulfilled the selection criteria explained in 2.2.6. In total, we worked with 28 foster chicks in 23 foster broods.
2.2.7. Foster Chick Relocation Schedule
In all but three cases, foster chicks were placed in nests between 8:00 and 9:30 AM with a crop half-full of food. At the time of the relocation, the resident chick was pulled from the nest weighed and measured and then both chicks, the resident and foster were placed back into the nest at the same time. In the three relocations following chick/egg predation, foster chicks were placed in the nest between 11 AM and 2 PM. No chicks were relocated on rainy days as adult movements and feeding rates are lower during rain (GV-T and DJB unpublished data). Just 2 foster chicks (7% of total foster chicks) were relocated the same day they were removed from the original nest.
2.2.8. Observations of Foster Parents/Foster Chick Interactions
In our first and second seasons working with macaw foster parents, video camera systems were installed in the majority of foster our nests (n = 12 foster nests with cameras of 14 foster nest total; 13 foster chicks). In the third season (n = 9 foster nests, 15 foster chicks), we did not work with video cameras in foster nest. All relocation procedures except #4 (“Triple brood”) had at least one foster nest with a video camera (Table 3). At the foster nests with video cameras, an experienced observer arrived at the foster nest at about 5:00 AM on relocation day and took observations of parent/chick interactions using the nest video system until 5 PM. Observers took focal group observations of known individuals to record all contact and feeding behaviors between the parent and the chicks. The recorded behaviors were (1) feeding, (2) preening and (3) brooding. Feeding refers to when adults grasp the bill of the chick crosswise from above and bob during regurgitation. Preening refers to when an adult gently touches the chicks’ body with its beak in a continuous manner. Brooding refers to when an adult positions its body in direct contact with the chick’s body. We considered that the time the nesting individuals (nesting females or nesting males) entered the nest and are visible on the video camera as the moment that the adults became aware of the foster chick’s presence in the nest. Nests with camera sample sizes varies slightly due to intermittent video camera malfunction during key behavioral interactions. One foster nest with video camera had problems with image (but not sound) at the time of foster chick placement and initial interactions were not recorded. Similar issues happened in another foster nest right after first interactions but before the first feeding. We had behavioral observations inside the nest of 9 different foster parent pairs.
2.2.9. Monitoring of Foster Chicks
We intensively monitored each foster nest for 10 days after each foster chick relocation. The monitoring process included (1) checking the foster chick’s crop content twice per day (5 AM and 5 PM); (2) providing supplemental food to the foster chick any time we checked the nest and found its crop was more than half-empty (in this way foster chicks were fed from 0 to 2 times per day), (3) monitoring weight gain by weighing both foster and resident chicks at 5:00 AM daily; (4) monitoring interactions between the foster chicks, resident chicks, and foster parents using video cameras when available (14 foster chicks in 12 foster nests with video cameras); (5) counting feedings per day of both foster and resident chicks as seen through the video cameras both live in the field and later from video recordings when cameras were available (See Table A2 for details of hours analyzed). To count feedings, we performed focal group observations of known individuals and recorded all feedings of each particular chick in a continuous manner each time they happened.
2.2.10. Supplemental Feeding Plan after Relocation of Foster Chick
Our objective with the supplemental feeding was to allow foster parents, foster siblings, and foster chicks to learn how to interact with each other without compromising the foster chick’s nutrition and overall health. We assumed that it would take time for foster parents to adjust to the new brood size and feeding requirements. We also assumed that it would take time for foster chicks that were syringe fed prior to placement, to learn how to consume chunky food regurgitated by the adult macaws. Our supplemental feeding plan had three stages: (1) Intensive supplemental feeding period. This was during the first 10 days after the foster chick was placed. The foster chick was checked twice per day and fed until the crop was 75% full. We slowly decreased the number of supplemental feedings, until by day 5 after relocation we left the chick with the crop only 50% full. This reduction in feeding was done in order to stimulate chick begging from the foster parents. In extreme cases (n = 2 chicks), foster chicks were just fed once (in the afternoon) in order to promote hunger and begging by the foster chicks. (2) Moderate supplemental feeding period. This was from day 10 after foster chick placement until the foster chick was 40 days old. In this stage, we checked the foster chick every day and provided supplemental food when its weight gain was less than 50% of the expected weight for its age on two consecutive days (expected weight was for [14] and unpublished data). (3) Passive supplemental feeding period. This was from 40 days old until the foster chick fledged. We checked the foster chick every other day and provided supplemental food if its keel was perceivable but there was moderate breast muscle development still found around it. In the 2017 breeding season, resident chicks were not supplemental fed. In the 2018 and 2019 breeding seasons, 75% of resident chicks lost weight during the 10-day adaptation period (n = 12 resident chicks). To address this, resident chicks were given supplemental food when the daily weight gain was 50% less than expected for its age. In all resident chick supplemental feeding cases, the chick was fed only until the crop was 50% full.
2.2.11. Foster Chick Acceptance Criteria
We established three levels of foster parent acceptance of foster chicks. (1) Initial acceptance: foster parents preen foster chick repeatedly and/or start attempting to feed foster chick. Attempts to feed refers to when the foster parent grabs the foster chick’s beak in an attempt to start regurgitation and then releases the beak. (2) Intermediate acceptance: foster parents consistently feed foster chick (as seen by video camera) and/or foster chick shows a half-full crop on daily checkups. (3) Full acceptance: foster parents feed both foster chick and resident chick similarly. The half-full crop criterion was chosen because our previous observations on wild macaw chicks that fledge show that chicks in the 19 to 33 days old age range (Stage 2B) have on average a half-full crop (mean = 2.1 in a 0 to 4 scale, n = 515 chick crop observations, from 61 macaw chicks that fledged, during 16 breeding seasons, GV-T and DJB unpublished data). When the foster chick was the only chick in the nest, we established just one level of foster parent acceptance: relocation was considered successful when the foster chick was being fed by foster parents and was gaining weight as expected.
2.2.12. Foster Chick Acceptance Analysis
In order to better analyze the process of foster chick acceptance by their new parents we quantitatively measured acceptance using chick feeding ratios and foster chick growth.
2.2.13. Chick Feeding Ratios
In order to show how foster parents were accepting foster chicks we calculated a ratio of feedings per day (foster/resident chick) for each day in each nest during the first 10 days after relocation. We collected feeding ratio data in two different ways: from direct field observations (one season) and from video recordings (two seasons). A total of 418 h of observations were conducted live in the field by multiple observers and 573 h of video were scored by a single observer (Appendix B). Feeding ratios from video observations include 227 h of nocturnal observations. To determine if chick feeding ratios increased over the first 10 days after relocation we conducted a least squares regression with feeding ratio as the dependent variable, day post relocation as the independent variable and nest ID as a random variable.
2.2.14. Foster Chick Growth
In order to evaluate foster chick quality and acceptance we calculated the logistic growth curves for foster chicks that fledged and compared them with the growth curves of wild macaws that fledged in our study area during the previous 19 breeding seasons [14]. For this analysis, we only included chicks with 25 or more daily weight measurements and ≥ 1 measurement taken during the first week of life [14]. A total of 23 foster chicks fulfilled this criterion. The non-manipulated chicks used in this analysis were individuals that fledged from nests with no foster chicks, had no major health issues, did not receive supplemental feedings, and had fully wild parents that were not-hand raised and released (see [40] for history of releases in our site). A total of 81 wild chicks fulfilled these criteria.
To calculate the logistic growth curves, we used the chick weights and a logistic model with the equation
where W = weight in grams, T = age of the chick in days, A = the asymptotic body mass, B = growth rate constant, C = age in days for which the growth rate is maximal, and e = the natural constant [14]. We also compared growth parameters of foster chicks and wild chicks grouped by brood size and hatch order. Wild chick groups were as follows: single chicks (n = 17), first chicks (n = 38) and, second chicks (n = 26). Curves were fitted using Data Fit 9.1.32 (Oakdale Engineering, 2014, Oakdale, Pennsylvania, USA). To determine if growth differed between wild chicks and foster chicks, we compared the three growth parameters using a Mann Whitney U test with p-values calculated using a χ2 approximation. The differences between groups were tested with Wilcoxon pairwise comparisons.
W = A/(1 + e (−B ∗ (T − C))),
2.2.15. Foster Chick Influence on Breeding Success
Previous studies on breeding ecology of psittacines have used population level breeding parameters to make comparisons within and among seasons [10]. In order to measure the impact of chick relocations on overall breeding success we compared the overall breeding success for our monitored nests during the three seasons working with foster chicks and the previous 17 seasons with no foster chicks. Macaw nests were monitored from mid-October to mid-April every breeding season. We used five breeding success parameters: (1) Chicks that fledged per available nest (# chicks that fledge/# available nests), (2) Chicks that fledged per nest with eggs (# chicks that fledge/# nests with at least one egg), (3) Chicks that fledged per nest with at least one chick (# chicks that fledge/# nests with at least one chick), (4) Percentage of younger chicks that died from starvation (# of chick starved/# second chicks, third chicks and fourth chicks hatched), and (5) Percentage of chicks that fledged (# fledged chicks/# total chicks). Macaw nests included for this part of the experiment included natural and artificial cavities [33]. However, nests where total clutch size and total number hatched were not known exactly were removed from the analysis. As mentioned above, some of the adult macaws at our site were hand-raised as chicks, released and continue to consume food at the lodge [40]. Offspring of those individuals are not included in this analysis. Wild chicks that received supplemental food for any reason at some point in their lives were also excluded. To determine if breeding success differed between seasons with foster chicks and seasons without foster chicks we compared the parameters for both groups using a Mann Whitney U test with p-values calculated using a χ2 approximation. All statistical comparisons were done using JMP Pro 15, with a confidence interval of 0.95 and α = 0.05. All results are presented as mean ± standard deviation unless otherwise indicated.
3. Results
Twenty-eight foster Scarlet Macaw chicks were placed in nests with wild macaw foster parents. All of them were successfully accepted and 89% of them (n = 25 chicks) fledged from their foster nests. This included 23 successful foster broods (4 single foster broods, 18 double foster broods and 1 triple brood). In general, foster chicks were placed back in wild nests at 22 ± 9 days old (min = 14, max = 46 days old, n = 28 chicks). In five double foster broods, both chicks were foster chicks.
Overall, we had 15% (4 of n = 28 chicks) of foster chicks placed as solo chicks and 75% of them fledged. All 36% of foster chicks (10 of n = 28 of chicks) placed as first chicks fledged and all but two of the 46 % (13 of n = 28 chicks) placed as second chicks fledged. The one foster chick placed as a third chick successfully fledged.
The three foster chicks that did not fledge died of depredation (n = 1), lightning (n = 1) and what was probably an unknown disease (n = 1, Appendix C). We prematurely terminated one chick translocation only 3.5 h after the chick was placed in the nest because sweat bees (family Halictidae) from a nearby beehive started entering the nest cavity and attacking the foster chick.
In the “Acceptance” relocation procedure, 67% of foster chicks were placed as the younger member of the brood and 33% of foster chicks were placed as the older chick (n = 11 foster chicks). In this group, four chicks hatched as third chicks in their original nests, were placed as 2nd chicks (n = 3) and 1st chicks (n = 1) and were all successfully accepted.
In the “Age difference” relocation procedure, the age difference range was 4 to 9 days. All chicks in this procedure were accepted. Here, 60% of foster chicks were placed as 1st chick (n = 6 foster chicks) and 40% as 2nd chicks (n = 4 foster chicks). In four cases, chicks were true siblings that hatched in the same nests with a five-day difference which in normal conditions would have signified the death by starvation of the younger chick. Indeed, from all the procedures combined, 29% (8 of n = 28 chicks) were members of multiple broods with age differences > 4 days (4 foster chicks as first chicks, 3 foster chicks as second chicks and 1 foster chick as a third chick). In all of these eight foster broods, all chicks successfully fledged.
In the “empty nest” relocation procedure, 60% of the foster chicks were placed as solo chicks (n = 5 foster chicks). One of them was 46 days old. Two foster chicks of the same age were placed in the same foster nest but in different days. The heavier chick was placed first at age 31 and second chick was placed at age 39. All chicks in this procedure were accepted.
For the “triple brood” relocation procedure all chicks were true siblings. Here, the third chick was successfully accepted and all three chicks fledged. However, all three chicks showed inconsistent weight gain, even > 10 days after relocation of the third chick. Due to the weight gain problems, we intensively managed this brood giving them a high fat content supplemental feeding when we found them with empty crops, until the youngest chick was 45 days old. We planned to conduct more than one triple brood relocation, but were unable due to depredation events in our other chosen foster nests.
In all foster broods in the first two seasons (n = 14 broods: 12 broods with video cameras and 2 without video camera), we placed the foster chick when the nesting female was within sight of the nest (range 0 to 10 m from the nest). In two cases, the nesting female stayed inside the nest covered with a towel when we placed the foster chick. In all but two cases, the nesting male was not present. In the majority of the cases, nesting pairs were present during the precise moment when foster chicks were placed in their nest (100% females, 86% males, n = 14 nesting pairs). In these two first seasons, two double foster broods had both chicks as foster chicks.
3.1. Foster Chick Acceptance
The behavior of foster parents when seeing the foster chick for the first time followed the same pattern in all cases. In all foster broods with video cameras (n = 12 broods), the first foster parent to have physical contact with the foster chick was the nesting female. First contact behavior was usually preening (61% of the time, n = 14 chicks with video cameras) but some foster parents first attempted to feed the foster chick (39% of the time, n = 14 chicks with video cameras). None of the females showed aggression towards the chick. On average, first contact was made 4.2 min after the foster mother arrived to the nest (n = 14 chicks with video available, range = 0.8 to 14 min) and first feeding was given on average 13 min after arrival (Time range = 0.8 to 76 min, n = 13 of 14 chicks with video available).
In all nests with cameras (n = 14), the first physical contact between an adult and foster chick happened in the first 15 min after the nesting female arrived at the nest. In the two cases when females took the longest to touch the foster chick (14 min) it was because she was paying attention to the climber getting ready to repel down from the tree. After an average of 4.6 ± 3.3 days foster chicks that were members of multiple chick broods consistently had half-full crops when checked (n = 23 chicks, min = 2 days, max = 15 days).
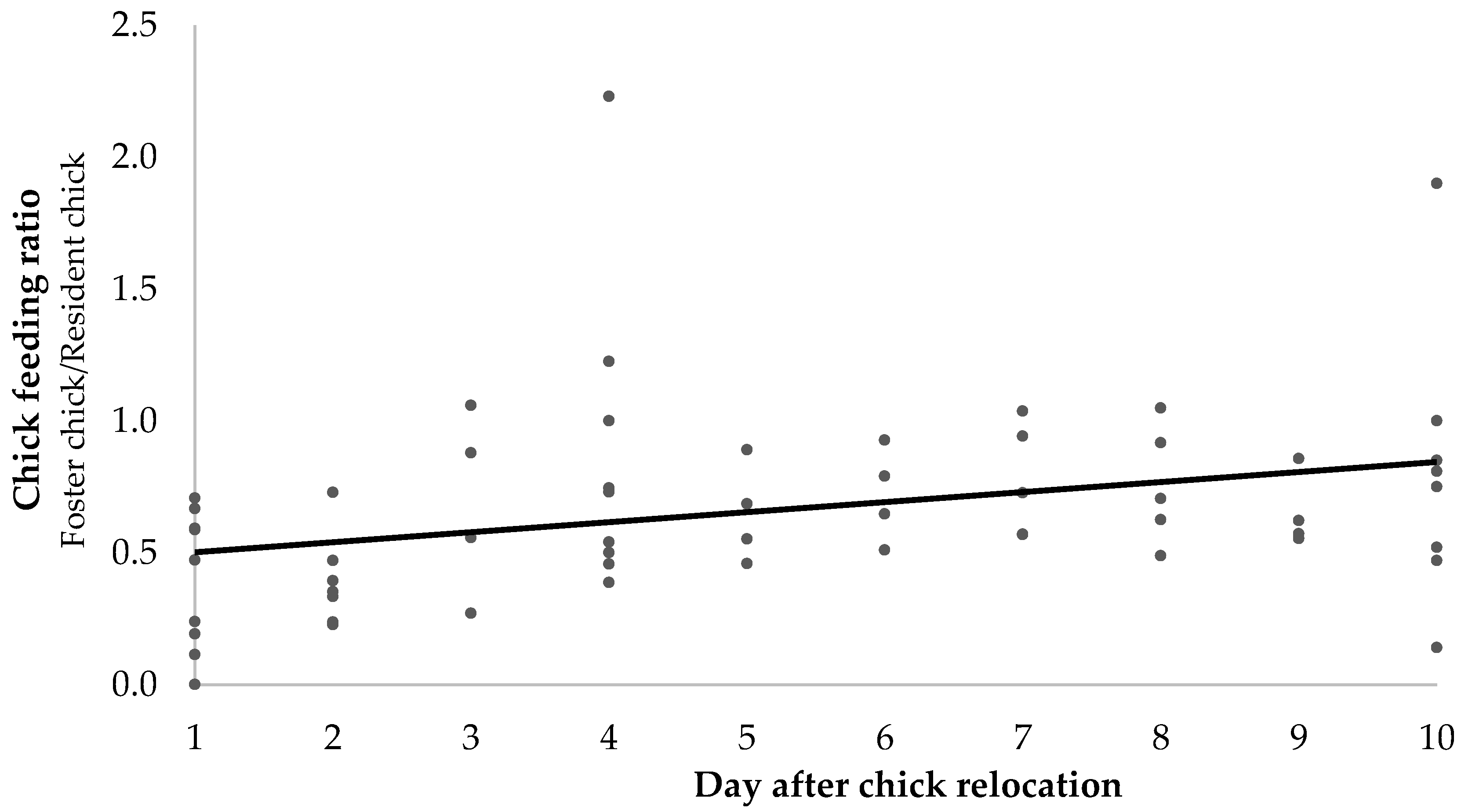
3.2. Foster Chick-Feeding Ratios
Daily feeding ratios from observations done in the field by multiple observers and from recorded observations done by one single observer showed similar patterns. Foster chicks were initially fed less than resident chicks (daily feeding ratio of 0.37 ± 0.25 on relocation day, n = 10 chick pairs), but feeding ratio increased progressively until feedings were similar for both chicks 10 days after relocation (daily feeding ratio = 0.8 ± 0.4, n = 10 chick pairs, Figure 3). The combination of day and nest (as a random variable) explained about 22% of the variation in the data and the relationship with day post relocation was highly significant (least squares regression: R2 = 0.22, df = 53, t-ratio 2.56, p = 0.013).
3.3. Foster Chick Growth
Foster chicks reached similar maximum weights compared to wild chicks (asymptotic size from logistic equation: all wild chicks combined: 1014.2 ± 79.7 g, n = 81, foster chicks 1020.3 ± 81.1 g, n = 23, Mann–Whitney U: χ2 = 0.14, df = 1, p = 0.7) and both grew at a similar rate (growth rate from logistic equation: all wild chicks combined 0.116 ± 0.016, n = 81, foster chicks 0.120 ± 0.014, n = 23, Mann–Whitney U: χ2 = 0.47 df = 1, p = 0.5). However, foster chicks reached maximum growth rate at a significantly younger age (Age at maximum growth rate from logistic equation: all wild chicks combined 26.3 ± 3 days, n = 81, foster chicks: 23.9 ± 1.7 days, Mann–Whitney U: χ2 = 13.6, n = 23, df = 1, p = 0.0022, Table 4).
Foster chicks grew significantly faster than second chicks (growth rate from logistic equation: foster chicks 0.111 ± 0.015, n = 23 chicks, second chicks, 0.121 ± 0.014, n = 26 chicks. Wilcoxon pairwise comparisons: Z = −1.96, p = 0.05). Foster chick growth was not significantly different than solo chicks or first chicks (Wilcoxon pairwise comparisons: Z > 0.84, p > 0.06, Table 4). Foster chicks reached maximum growth rate at a significantly younger age than both first and second chicks: 1.5 days younger than first chicks (first chicks: 25.7 ± 2.4 days old, n = 38 chicks; foster chicks: 23.9 ± 1.7 days old, n = 23 chicks, Wilcoxon pairwise comparisons: Z = −2.61, p = 0.009) and 3.5 days younger than second chicks (second chicks: 28.2 ± 3.2 days old, n = 26 chicks, foster chicks 23.9 ± 1.7 days old, n = 23 chicks. Wilcoxon pairwise comparisons: Z = 5.18, p ≤ 0.001, Table 4).
3.4. Foster Chick Influence on Breeding Success
In general, during the three seasons with chick relocations, more chicks fledged, more nests had chicks that fledged and fewer chicks died of starvation (Table 5).
4. Discussion
Our use of wild Scarlet macaws as foster parents along with supplemental feeding and veterinary care was categorically successful: all foster chicks were accepted by the foster parents with no chick rejection, foster chicks were fed at rates similar to resident chicks, foster chick growth was similar to wild chick growth, and almost 90% of all foster chicks fledged. Moreover, the use of foster parents dramatically reduced chick mortality due to starvation and increased overall reproductive success in the study area.
4.1. Scarlet Macaws as Foster Parents in the Wild
There are few studies of foster parents in wild psittacines but most of them are quite complete. Their objectives varied from a management tool to increase population recruitment (RGA unpublished data, [5,18]), to a scientific technique to understand behaviors such a parent/chick recognition [8], hunger response [42] and food allocation [22]. None of these previous studies addressed the potential conflict of increasing brood size in species that show brood reduction strategies in the early stages of the nesting period nor why pairs allow their own chicks to starve at the beginning of the nesting period but then accept additional unrelated chicks later in the same nesting event.
We designed our experimental procedures to avoid placing foster chicks during the starvation risk period for the species. According to our investigations on brood reduction by chick starvation of Scarlet Macaws [29], we observed that fourth chicks are always left to starve in the first week of life and third chicks in their first two weeks. For second chicks, no death by starvation was recorded after 25 days of age. In fact, 88% of second chicks that starved were younger than 20 days old. For that reason, we consider “the starvation risk period” in Scarlet Macaw is from zero to 20 days old [29]. We did not place additional chicks in foster nests when the youngest member of the foster brood was on average younger than 22 days old. A similar strategy was used in relocating Yellow-shouldered Parrot foster chicks, where only chicks older than two weeks were used as foster chicks because mortality rates are higher in the first weeks of life [18]. Evidence from Scarlet Macaw fostering work in Guatemala support this suggestion, as foster chicks < 15 days old seemed to be rejected at higher rates than older chicks (RG-A personal observations). In foster chicks of Puerto Rican Amazons [5] and the Galahs [8] individuals as young as one week old were accepted in whole brood swaps. These two studies warn about using older foster chicks due to the evidence that adults do not recognize small chicks as individuals but they do recognize older chicks. No warnings are made about placing young foster chicks during starvation risk periods.
A difference between our foster parent experiment and previous studies with wild psittacine foster parents is the presence or absence of the nesting pair when foster chicks were placed in nests. In the two Amazon parrot studies in Puerto Rico and Venezuela and in the Galah study in Australia, foster chicks were placed in nests when parents were absent in order to minimize disturbance and possibly nest abandonment [5,8,18]. Multiple authors stated that they thought the foster parents did not detect the placement of the foster chick [5,42], but in our study that was clearly not the case. In our case, the majority of nesting pairs were present during the precise moment when foster chicks were placed in their nest. In a few cases, the female nesting individual was even inside the nest, so the argument that foster parents did not detect an additional chick is not valid in our case. It is worth emphasizing that in our experiment, we did not consider the nesting pair presence or absence at the nest as an important factor because at our study site, we have been monitoring macaw nests intensively for the last 20 years and Scarlet Macaw pairs are very accustomed to our nest checks and rarely display typical disturbance behaviors when researchers visit the nests and manipulate the chicks [5]. However, in other areas with little or no history of nest checks or chick manipulations, human presence may disturb nesting pairs and alter the results of chick fostering attempts.
4.2. Foster Chick Acceptance and Rejection
Overall acceptance of foster chicks in our investigation was excellent, as expected. There are published records of wild parrots in the genera Cacatua, Amazona, Platycercus and Ara ([8,18,42] and RG-A unpublished data) and captive Cylopsitta, Alisterus, Amazona, Pionus, Cacatua and Melopsittacus ([4,5,8,19,20] and GV-T personal observations) accepting and raising foster chicks. In all the studies done in the wild, including ours, when foster chicks were accepted, fostering manipulations caused no major disruption of adult nesting behaviors [5]. In the case of the Galah, the main disruption of adult nesting behavior happened when whole broods were swapped for unrelated broods [8]. Some pairs hesitated for several hours before first entering the nest, but once one adult entered the nest (generally the female), the other nesting adult followed. This hesitation is explained by the fact that older Galah chicks (few weeks from fledgling) reply to their own parent’s calls when they arrive at the nest and foster chicks did not respond to foster parent’s calls when they arrived [8]. In our case, we did not detect hesitation to enter the nest as in the Galahs. Usually Scarlet Macaw nesting females in our site were eager to come back and check on their chicks as soon as they were returned to the nest by the researchers.
High acceptance of foster chicks after chick predation or egg hatching failure was surprising but also not unexpected; mainly because it has been reported in studies with foster chicks in Amazon parrots in the wild. In Yellow-shouldered parrots, three of four foster chicks placed after full predation events were accepted [18]. In wild Puerto Rican Amazons two foster chicks placed after eggs failed to hatch were accepted, even though the foster chicks were another species: Hispaniolan Amazons (Amazona ventralis [5]). In our experiment in Tambopata, in the cases of foster chick acceptance after the resident brood was depredated, after chicks were killed by lightning, and after egg hatching failure, nesting pair behavior was very similar to that reported by the previous studies with Amazon parrots [5,18,24]. There was some initial hesitation, especially the very first time the foster chick was seen, but once it was fed, the nesting pair behavior fell into the normal attendance pattern according to the foster chick’s age.
In all three studies, including ours, the timing in which the foster chick was placed after the nest was emptied was likely a key aspect [5,18]. In the case of replacing unviable eggs with a foster chick, the swap probably needs to be done as close to the estimated hatching date as possible. The hatching period is a very sensitive period for the nesting pair. It offers a very small window to replace eggs for foster chicks. Nesting individuals, especially nesting females, tend to decline in attentiveness a few days after the end of the normal incubation period if the eggs fail to hatch [5]. Even though the nesting pair keeps visiting the nest after egg failure, visits are likely more related to a desire to defend the nest cavity and maintain ownership GV personal observations, [43,44].
An unexpected result in our experiment was that all foster chicks were accepted. In three of four foster parent studies from the literature, a few foster chicks were rejected in each. In the Puerto Rican Amazon case [5], two older chicks that were swapped for one foster chick were rejected, even when the foster chick was an offspring of the foster parents. In the Galah study [8] 5% of foster chicks (n = 10 chicks, 3 broods) were rejected; perhaps because all of them were placed very close to fledgling time and the foster parents seemed able to recognize their own chicks either by vocalizations or by unique physical cues [8]. In the Yellow-shouldered Parrot [18], 9.3% (n = 5 chicks) were rejected. Here, rejection was attributed to a low feeding response of foster chicks and different developmental stages between foster chicks and wild offspring. In the Scarlet Macaw in Guatemala, foster chicks were occasionally rejected as well, presumably because they were too young (between 10 to 20 days old) or because brood size was increased over the maximum brood size of the species in the area (broods of three or four chicks, RG-A unpublished data). The Crimson Rosella study did not mention chick rejection at all [22].
A tentative explanation for the zero foster chick rejection found in our experiment is that we measured rejection in a different way than in the previous studies [5,8,18]. We considered that a foster chick was rejected when daily feeding ratios (foster/resident chick) were not similar and when the tendency of feeding-ratios was not positive after 10 days of foster chick relocation. In the Puerto Rican Amazon, the indicator of acceptance was also feedings but based on crop size observations after a few days and not direct observations of feeding [5]. In addition, rejection happened when whole broods were swapped but not when chicks were added to a brood. We did not swap entire broods, so these rejections are not comparable to our case. In the Yellow-shouldered Parrot, the foster chick acceptance indicator was also feedings based on crop size and observations of the absence of injuries at the end of day of relocation. Here, when foster chicks did not have large crops until the next morning after relocations, they were removed and placed in another nest [18]. Interestingly, in our experiment, the only cases in which foster chicks showed large crops in the day after relocation were when the foster chick was the only chick in the nest. In the Scarlet Macaw in Guatemala, foster chick acceptance indicator was also crop size and crop content after a maximum of three days post relocation. After that period, chicks found with crops with no macaw food content were relocated to another foster nests (RG-A unpublished data). In our case, all our foster chicks that were members of multiple broods needed on average five days to show half-full crops. They were fed by foster parents, as clearly observed in videos, but did not have large crops. Under our acceptance/rejection criteria those chicks were not considered rejected.
Zero foster chick rejection in our experiment might be due to the fact that we matched ages/developmental stages between foster chicks and resident chicks. Developmental stages in our experiment were defined based on our extensive knowledge on the nesting biology of the species [14]. Therefore, our foster chicks looked very similar to the resident chicks in nearly all cases. The importance of matching similar ages between foster chicks and resident chicks in chick additions, chick swaps, and whole brood swaps was mentioned in all the previous studies (RG-A personal observations, [5,8,18]). All investigations that worked with psittacine foster parents address the fact that foster chick acceptance and especially rejection were related to age differences among chicks involved (RG-A personal observations, [5,8,18]). In the Yellow-shouldered Amazon [18] and in the Galah [8], foster chicks and foster broods that differed in age from the resident chicks and broods were rejected. In the Puerto Rican Amazon [5] and in the Crimson Rosella [22] studies the authors considered that pairing chicks that “look similar” to be very important. In the Scarlet Macaw in Guatemala, it was considered a key aspect in order to warrant chick acceptance (RG-A unpublished data).
4.3. Foster Chick Chick-Feeding Rates
No previous studies of wild Psittacidae as foster parents have analyzed acceptance of foster chicks using chick feeding rates as an indicator of acceptance. The daily feeding ratios in our experiment showed that foster chick acceptance was a slow process that needed more than one day of post-relocation monitoring before concluding failure. In the Crimson Rosella study in Australia, feedings (food transfers) were used as a tool to: (1) quantify hunger response when broods or individual chicks were placed back in the original nest [22], and (2) to understand food allocation among brood members [42]. However, in both experiments, resident broods and chicks were placed back in their original nest three hours later, so there was no way to analyze daily feeding rates. In the Galah [8] and Puerto Rican Amazon studies [5] feedings were used in a descriptive manner, not in an analytical manner.
Other studies have addressed the first response of foster chicks to foster parents. In our experiment the results were unexpected because the foster chick reaction we observed most commonly could be considered a “distress” response. The majority of foster chicks (n = 23) were syringe fed from a few days old to 20 days of age. Because of that, they showed low or even nonexistent feeding response when approached by an adult macaw. When foster parents, grabbed foster chicks’ beaks in an attempt to feed, the foster chick usually shook its head and pulled away. This pulling away behavior was consistently observed during the first days after chick relocations, even twelve hours after last supplemental feeding, when the chicks had little to no food in the crop. Our observations showed that this pulling away decreased slowly during the first 10 days post relocation as the foster chicks learned to receive food from the adult macaws. This same sequence of behaviors was also observed in Guatemala (RG-A personal observations). We consider this behavior a distress response because we have never seen any wild chicks pull away from parents in this fashion (even when their crops are nearly 100% full) in either Peru or Guatemala.
A more intense reaction was observed with the Puerto Rican Amazon in which some hand-raised chicks gave fright responses and distress vocalizations when they were placed into the wild nests and first encountered adult parrots attempting to brood and feed them [5]. The first response to foster parents in Galah Cockatoo chicks was not a distress response, instead, the chicks gave little or no response, especially for chicks in the second half of their nesting period. At that age, Galah chicks start to respond to parent calls when they arrive at the nest and foster chicks did not reply to foster parent call when they first arrived to the nest. However, after a few hours of not being fed, the tendency for rejection by the nestling was overruled by hunger. Once the nestling was hungry, begging and vocalizations attracted foster parents that proceeded to enter the nest and feed them [8].
4.4. Foster Chick Supplemental Feeding
In our experiment, chicks were given supplemental food once or twice per day during the transition period when foster chicks were learning how to be fed by wild macaws. Even though they were not responding when foster parents tried to feed them, they were not losing weight or showing signs of nutritional deficiency due to our supplemental feedings. This is similar to the fostering protocols used in Guatemala which obtained similar results (RG-A personal observations). This evidence from Peru and Guatemala leads us to conclude that supplemental feeding gave foster chicks time to learn appropriate feeding response behaviors and increased chick acceptance and the success of this technique.
Our experiment raises the question of whether or not we could have obtained similar results by just feeding the chicks in situ, without pulling them out of the nest and relocating them. Multiple lines of evidence suggest that just feeding may not have been successful. In previous breeding seasons, we provided supplemental feeding to starving younger Scarlet Macaw chicks in the nest (n = 5 chicks: 1 s chick and 4 third chicks) on average three times per day (range = 1 to 4 times per day) for an average of 4 days (range = 3 to 7 days) and this failed to prevent starvation. In addition, supplemental feeding provided to younger Green-rumped Parrotlet chicks upon hatch in the nest three times per day marginally increase the probability of survival of last-hatched chicks but they still experienced significantly higher mortality than early hatched chicks and it did not improved probability of survival of penultimate hatched chicks [45]. Moreover, our observations suggest that feeding alone may be insufficient to save younger macaw hatchlings as parents may selectively exclude them from brooding: second and third chicks that starved were excluded from brooding from 6 to 35 times more than second chicks that fledged (n = 9 macaw broods, 3 breeding seasons, 250 video hours analyzed, GV-T unpublished data). This is problematic because improper brooding of captive macaw hatchlings and young chicks can cause abrupt temperature fluctuations that may result in thermal stress and death [39].
4.5. Foster Chick Growth Rates
An interesting finding was that foster chicks in our experiment were not only accepted by foster parents, but also raised as wild chicks. Some foster chicks were in poor condition when removed from their nests; either underweight, not gaining weight as expected or sick, and they received special treatments in order to recover. Even though these individuals grew slowly when young, their overall growth rate ended up similar to wild chicks. These results fit with the compensatory growth principle that states that given adequate conditions, slow development as a result of poor nutrition is followed by accelerated growth. Growth rates become similar to nestlings that did not experience nutritional stress at all [39,46]. Besides, our hand-raising procedures in the nursery and supplemental feeding plan during the first 10 days after relocation provided enough nutrition to foster chicks, so they were able to compensate for the low caloric intake received from foster parents during the first days in foster nests, and this allowed them to catch-up and attain maximum growth rates and maximum weights similar to the wild chicks.
The fact that foster chicks reached the maximum growth rate almost two days earlier than resident chicks is likely a direct consequence of our hand-feeding procedures. Captive raised Scarlet Macaw chicks grow differently than their wild equivalents. Indeed, purely captive-raised chicks reached the maximum growth rate even sooner than foster chicks in this experiment [15]. These differences in growth might be related to differences in the consistency of macaw chicks’ diets. In the wild, the diet of macaw chicks contains full seeds and even tree bark [47] so it may take more time and energy to digest than the puree like formula that is used in captivity [48]. In order to get foster chicks extra fat in preparation for the adaptation process in their new foster nest, we provided a high fat diet (formula with nuts and peanut butter added), large portions, and high feeding frequencies [29].
4.6. Testing Starvation Drivers
The main driver behind brood reduction in two chick broods appears to be the age difference between brood members [29]. This age difference effect predicts that the greater the age difference between brood members the higher the risk of starvation of younger brood members. However, work in Costa Rica and specially in Guatemala [RG-A unpublished data] suggests that macaws that allowed their own chicks to starve at the beginning of the nesting period accepted, raised, and fledged additional unrelated chicks later in the same nesting event even when age differences were >5 days (average age difference 9 days, maximum = 14 days, n = 60 chicks). In our foster parent work, we confirmed that these age difference effects on starvation did not apply when brood members were older than 20 days. We had seven foster chicks with age differences > 5 days from their foster sibling. In all seven foster broods, both chick members successfully fledged. One reason why age difference might not be correlated with starvation in multiple broods with chicks older than 20 days old is that the younger the chicks, the more age specific the parental care requirements (aka: brooding and feeding). Recommendations for brooding temperatures in captivity indicate that newly hatched chicks need to be kept 2 °C warmer than 5–9 days old chicks and extreme temperature fluctuations at this time can be harmful or even fatal to the chicks [39]. When the pinfeathers of chicks start to show, around 18 days old, chicks are less affected by temperature and when they are heavily pin feathered, around 30 days old, heat requirements diminish considerably [39]. The recommended feeding frequency also varies from every hour for hatchlings to every 3 to 4 h for 5 to 9-day old chicks and are even more variable as the birds age (Appendix A, [39,49]). Hence, when chicks are older than 20 days, chicks that are >5 days apart “look similar” and their parental care requirements, brooding and feeding, are similar. The fact that foster parents were able to fledge chicks that were over 5 days apart suggests that the developmental stage in which the foster chicks were placed may have been a key factor. Since both foster siblings were at the age at which parental care requirements were very similar, even though foster parents needed time to adjust their food provisioning and foster chicks needed to learn how to be fed, death by starvation was no longer a major risk.
4.7. “Triple Brood” Chick Relocation Procedure
In our triple brood procedure, the fact that all foster chicks did not gain weight as expected suggests that the parents were unable to provide sufficient food. Even the resident chick, that regularly had the largest crop of the trio, could not consistently reach the average weight for its age. It seems like even though all chicks where being fed, the macaw foster parents were not able to feed three chicks properly. According to our work on Scarlet Macaws in southeastern Peru a maximum of 2 chicks fledge under natural circumstances. In fact, only 37% of nesting pairs managed to fledge 2 chicks and 1.3 chicks per nest is the average chick production per successful nest in the area [29]. However, there are reports of rare successful natural triple broods from Costa Rica [50] and Guatemala [51] suggesting that conditions may vary geographically. In Guatemala, RG-A’s team has created a total of four triple and two quadruple foster broods across at total of three breeding seasons. From the four triple foster broods created, all chicks died in one, all chicks fledged in another and one fledged only two chicks. In the two quadruple foster broods created, only three chicks fledged from both (RG-A unpublished data).
In the Yellow-shouldered Parrot foster parent research [18] it was recommended to not create foster broods that were bigger than the optimal brood size of the species. Our results agree with this conclusion, and show that it is important to calibrate foster brood size using as a general indicator the optimal brood size of the species in the area.
5. Conclusions
Our technique of macaw foster parents and post relocation supplemental feeding was categorically successful. All relocated foster chicks were successfully accepted by their foster parents (n = 28 chicks across three seasons) and 89% of them fledged. The only three foster chick fatalities were due to unknown disease, predation, and lightning. Foster chick acceptance by foster parents was a slow process. Foster chicks were initially fed less than resident chicks, but feeding ratio increased progressively until feedings were similar for both chicks 10 days after relocation. Foster chicks needed on average about 5 days to consistently have half-full crops when checked. Growth rates of foster chicks were similar to wild chicks and both chick groups reached similar maximum weights. However, foster chicks reached maximum growth rate at a younger age. These differences were likely due to differences in diet and feeding schedule. Our foster parents technique increased the reproductive success of our studied population: fledging success per available nest increased from 23% (2000–2016) to 43% (2017–2019) and chick death by starvation decreased from 35% to 6%.
Interdisciplinary Collaboration in Parrot Conservation
Our ability to produce foster chicks that were successfully accepted and that were very similar to wild chicks by fledgling time, is the result of the integration of three different fields: parrot ecology, avian veterinary medicine, and aviculture. Psittacines have been the heart of aviculture for centuries and there are many well know breeding techniques that can be easily modified and adjusted for use in the wild [52]. In fact, the Scarlet Macaw is considered one of the most productive species of macaws in captivity (Mark Moore, co-owner of Hill Country Aviaries, USA. Personal communications, [53]). In our experiment, we used information from the aviculture literature [15,39,49,54] and worked closely with experienced psittacine breeders. We also worked with avian veterinarians that took care of chick health issues and provided insights from their experiences with captive psittacines. Lastly, we integrated our knowledge on breeding ecology and nesting behavior of the species [14,33,47,55,56]. As has been demonstrated with the Puerto Rican Parrot [5], Spix’s Macaw [57,58] and our work with Scarlet Macaw, the integration of ecologists, veterinarians and aviculturists has great potential to assist management actions in the wild.
Author Contributions
Conceptualization, G.V.-T., R.G.-A. and D.J.B.; methodology, G.V.-T., R.G.-A. and D.J.B.; software, G.V.-T.; validation, G.V.-T., formal analysis, G.V.-T. and D.J.B.; investigation, G.V.-T. and D.J.B.; resources, G.V.-T.; data curation, G.V.-T.; writing—original draft preparation, G.V.-T.; writing—review and editing, G.V.-T., R.G.-A. and D.J.B.; visualization, G.V.-T.; supervision, D.J.B.; project administration, G.V.-T.; funding acquisition, G.V.-T. and D.J.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was part of G.V.-T.’s graduate studies supported by a National Science Foundation (NSF) Graduate Research Fellowship, by a Diversity fellowship from Texas A&M University and a Regents Fellowship from the College of Agriculture and Life Science from Texas A&M University. Chick rearing work was funded by generous private donations to two crowdfunding campaigns run by Experiment.com in December 2017 and December 2018 along with donations from Chris Biro from Bird Recovery International, and Janice Boyd from Amigos de las Aves USA and The Parrot Fund. Macaw chick formula was donated by ZuPreem. Discounted rates for lodging expenses and boat transportation were given by Rainforest Expeditions S.A.C. thanks to Kurt Holle. Data collection was funded in part by the Schubot Center for Avian Health thanks to Dr. Ian Tizard and funds from DJB lab.
Institutional Review Board Statement
This study follows the Animal Use Permits: AUP 2012-145 “Macaw and parrot biology in southeastern Peru” given by Texas A&M University. The work has been authorized by the Peruvian government under the following research permits: Resolución Jefatural de la Reserva Nacional Tambopata, N 16-2018-SERNAMP-JEF, Resolución Jefatural de la Reserva Nacional Tambopata, N 22-2016-SERNAMP-JEF, Resolución 33-2015-SERNAMP- JEF, Resolución 13-2014-SERNAMP-JEF, Resolución 19-2013-SERNAMP- JEF, Resolución 18-2012-SERNAMP- JEF.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to permit restrictions from the National Service of Protected Areas of Perú (SERNANP).
Acknowledgments
Thanks to Mark Moore, María Belen Aguirre, Sharman Hoppes, Sophie Hebert, Liz Villanueva Paipay, Gustavo Martinez, Carlos Huamaní, Shannan Courtenay, Dylan Whitaker, Roshan Tailor, George Olah, Bruce Nixon, Deysi Delgado, Jorge León, Susanne Vorbrüggen, Lauren Bazley, Patricia Gray, Simon Kiacz, Maximo Gonzales and the whole BS 2017, BS 2018 & BS 2019 “guacamayeros” volunteer teams for their invaluable hard work. Thanks to Vanesa Hilares from AIDER permit support. To Janice Boyd and Rodrigo Leon for the scientific advice. To Thomas Lacher, Ian Tizard and especially to Jane Packard for the mentoring and the corrections provided to this manuscript. To Keith Regelmann for his collaboration on our crowdfunding campaigns. Special thanks to Amanda Lucile (Mandy Lu) Brightsmith and Samantha King Brightsmith. Dedicated to Lucille Smith, Mónica Trauco and Oscar Vigo.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Appendix A
Diet details for Scarlet Macaw wild chicks in the nursery: Throughout the project macaw chicks were fed based on their age and other special circumstances as outlined here and in Table A1. Formula for neonates (<4 days old) was prepared as 1-part Zupreem formula to 4 parts water. For chicks ≥4 days old regular chick formula was 1-part Zupreem formula to 3 parts water plus peanut butter in the majority of the cases. For chicks ≥12 days old a mix of shredded raw Brazil nuts, pecans, and peanuts was added to the regular chick formula (Table A1). The majority of the time chicks were fed when their crops were empty or close to empty resulting in a feeding frequency of about once every 2.7 h when they were under 4 days of age to about once every 5 h when they were between 15 and 20 days old [modified from 39]. This protocol was followed for 21 chicks. For one chick we added shredded peanuts and peanut butter to the neonate formula starting at age 2 days and four chicks that had additional health problems received customized feeding regimes.
One underweight third chick was fed neonatal formula until it was 11 days old because its digestion was slow. From 12 days on it was fed regular chick formula. By age 15, it showed slower growth and slower development for its age but by 24 days old, its weight was as expected for its age. A similar situation happened with the underweight forth chick that arrived to the nursery right after hatching. The chick was fed neonatal emergency formula on its first day of life and neonate formula its second day of life. Subsequently it was moved up to neonate formula plus until it was 11 days old. At age 7 and age 8 chick showed early signs of slow digestion and it was given a mix of warm papaya juice and cinnamon added to its usual food until crop size increased to half crop full, once per day. From hatch, this chick showed a slower growth and slower development for its age but by age 25, it weighed as expected for its age. A second chick with signs of starvation, was given a special neonatal emergency formula (1 part Formula One by Avitech and 4 parts water; (http://www.avitec.com/Formula-One-for-hand-feeding-hatchlings-s/70.htm, accessed on 1 October 2016) and subcutaneous fluids for its first 12 h in the nursery. In these first 12 h it gained 78% of its arrival weight and after that the chick was fed according to its age.
One chick with a large botfly infection and low weight received two feedings that were a mix of neonatal emergency food and regular neonatal formula.
One first chick that was brought in as part of the acclimation process to create a triple brood, had food aspiration problems in its second day in the nursery when it was 19 days old. The chick was under antibiotics, anti-inflammatory and antifungal oral treatment for the following 20 days (15 days in the nursery, 5 days after nest relocation). This chick’s weight gain was always as expected for its age. All of the remaining chicks were fed normally according to their age.

{kind=link}
{kind=link}
{kind=link}
Table A1.
Summary of diet of wild macaw chicks in the nursery. Food names were assigned to differentiate among five different types of food provided. Neonate food was given to younger chicks (under 4 days old) and Regular food to older ones (over 4 days old). Emergency food was given to chicks showing signs of starvation. PLUS foods contain peanut butter and EXTRA PLUS foods contain peanut butter plus shredded raw Brazil nuts, pecans, and peanuts. Chick age is given in days. Formulas used are well-known commercial formulas use to raised macaw chicks in captivity: Zupreem Embrace (https://www.zupreem.com/products/birds/embrace-plus/, accessed on 1 October 2016) and Formula One by Avitech (http://www.avitec.com/Formula-One-for-hand-feeding-hatchlings-s/70.htm, accessed on 1 October 2016). Proportion used to prepare formula was the recommended by the manufacture.
Table A1.
Summary of diet of wild macaw chicks in the nursery. Food names were assigned to differentiate among five different types of food provided. Neonate food was given to younger chicks (under 4 days old) and Regular food to older ones (over 4 days old). Emergency food was given to chicks showing signs of starvation. PLUS foods contain peanut butter and EXTRA PLUS foods contain peanut butter plus shredded raw Brazil nuts, pecans, and peanuts. Chick age is given in days. Formulas used are well-known commercial formulas use to raised macaw chicks in captivity: Zupreem Embrace (https://www.zupreem.com/products/birds/embrace-plus/, accessed on 1 October 2016) and Formula One by Avitech (http://www.avitec.com/Formula-One-for-hand-feeding-hatchlings-s/70.htm, accessed on 1 October 2016). Proportion used to prepare formula was the recommended by the manufacture.
| Food Name | Age Range | Ingredients | Formula/Water Proportion | |||||
|---|---|---|---|---|---|---|---|---|
| Min (days) | Max (days) | Formula Zupreem Embrace Baby Bird | Formula One Avitech | Peanut Butter | Sheered Seeds | Water | ||
| Neonates Formula | 0 | 9 | Yes | No | No | No | Yes | 1 to 4 |
| Neonates Formula Plus | 2 | 20 | Yes | No | Yes | No | Yes | 1 to 4 |
| Regular Formula | 4 | 43 | Yes | No | No | No | Yes | 1 to 3 |
| Regular Formula Plus | 4 | 74 | Yes | No | Yes | No | Yes | 1 to 3 |
| Regular Formula Extra Plus | 12 | 28 | Yes | No | Yes | Yes | Yes | 1 to 3 |
| Special Emergency Formula | 7 | 13 | No | Yes | No | No | Yes | 1 to 4 |
Appendix B
Table A2.
Video observations of scarlet macaw behavior in foster nests. Field observations were done by a mix of 20 different assistants watching live video feeds in the field. Recorded observations were done by one experienced observer using video recordings and included recordings of both diurnal and nocturnal activity. A total of 10 chicks were observed with video, 3 in 2017 and 7 in 2018.
Table A2.
Video observations of scarlet macaw behavior in foster nests. Field observations were done by a mix of 20 different assistants watching live video feeds in the field. Recorded observations were done by one experienced observer using video recordings and included recordings of both diurnal and nocturnal activity. A total of 10 chicks were observed with video, 3 in 2017 and 7 in 2018.
| Type of Observation | Seasons | # Total Chicks | # Observers | Total Hours Observed | Hours Observed Per Day | |||
|---|---|---|---|---|---|---|---|---|
| Max | Min | Average | St Dev | |||||
| Field Observations | 2018 | 7 | 20 | 417.9 | 12.0 | 4.3 | 8.4 | 2.8 |
| Recorded Observations | 2017 and 2018 | 10 | 1 | 573.4 | 23.6 | 3.7 | 9.0 | 4.2 |
Appendix C
Foster chick fatalities. During the three years of work with wild macaws as foster parents, three foster chicks perished in their foster nests.
The first one died five days after being placed possibly because of an unknown disease. This foster chick had half-full crop by the day after relocation but just ¼ full on the following days. On those days, it was fed supplemental food. Starting on the day after relocation, this foster chick showed small red hematomas, first on the right flank, then the left flank, next to the keel with a scratch-like wound on the right leg. The foster chick was the third chick in its original nest. Both chicks in the original nest died with the same type of hematomas: the second hatched chick at 6 days of age and the first hatched at 12 days.
The second case of a foster chick death was due a combination of predation and lightning hitting the video cable systems installed in artificial nest (PVC pipe). Nest was found with cable system burned and the access door blown off. The foster chick (39 days old) and the resident chicks (41 days old) were not found inside the nest or in the surroundings. Marks of large claws were found around the door of the PVC nest box and on the tree branches from where nest was hung.
A third case of a foster chick death was due to lightning hitting the artificial nest (wooden box) that blew the base and top off the nest. The foster chick was 36 days old. Both foster and resident chick were found dead on the ground, below the nest tree, right after the thunderstorm stopped. Necropsy suggested that the foster chick’s death was due to electrocution and resident chick’s due to the fall.
References
- Cade, T.J. The husbandry of falcons for return to the wild. Int. Zoo Yearbook 1980, 20, 3. [Google Scholar] [CrossRef]
- Saint Jalme, M. Endangered avian species captive propagation: An overview of functions and techniques. Avian Poult. Biol. Rev. 1999, 13, 16. [Google Scholar] [CrossRef]
- Fentzloff, C. Breeding, artificial incubation and release of White-tailed sea eagles. Int. Zoo Yearb. 1984, 23, 17. [Google Scholar] [CrossRef]
- Romer, L. Management of the Double-eyed or Red-Browed fig parrot. Zool. Soc. Lond. 2000, 37, 6. [Google Scholar]
- Snyder, N.R.R.; Wiley, J.W.; Kepler, C.B. The Parrots of Luquillo: Natural History and Conservation of the Puerto Rican Parrot; Western Foundation of Vertebrate Zoology: Los Angeles, CA, USA, 1987. [Google Scholar]
- Beissinger, S.R.; Snyder, N.F.R. New World Parrots In Crisis; Smithsonian Institution Press: Washington, DC, USA, 1992. [Google Scholar]
- Beissinger, S.R.; Butcher, E.H. Sustainable harvesting of parrots. In New World Parrots in Crisis; Beissinger, S.R., Snyder, N.F.R., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1992; pp. 73–115. [Google Scholar]
- Rowley, I. Parent-offspring Recognition in a Cockatoo, the Galah Cacatua roseicapilla. Aust. J. Zool. 1980, 28, 9. [Google Scholar] [CrossRef]
- Beissinger, S.R.; Waltman, J.R. Extraordinary clutch size and hatching asynchrony of a Neotropical parrot. Auk 1991, 108, 863–871. [Google Scholar]
- Sanz, V.; Rodriguez-Ferraro, A. Reproductive parameters and productivity of the Yellow-shouldered Parrot on Margarita Island, Venezuela: A long-term study. Condor 2006, 108, 178–192. [Google Scholar] [CrossRef]
- Vaughan, C.; Nemeth, N.; Marineros, L. Ecology and management of natural and artificial scarlet macaw (Ara macao) nest cavities in Costa Rica. Ornithol. Neotrop. 2003, 14, 381–396. [Google Scholar]
- Marineros, L.; Vaughan, C. Scarlet macaws in Carara. In The Large Macaws: Their Care, Breeding and Conservation; Abramson, J., Spear, B.L., Thomsen, J.B., Eds.; Raintree Publications: Ft. Bragg, CA, USA, 1995; pp. 445–468. [Google Scholar]
- Krebs, E. Breeding biology of crimson rosellas (Platycercus elegans) on Black Mountain Australian Capital Territory. Aust. J. Zool. 1998, 46, 17. [Google Scholar] [CrossRef]
- Vigo, G.; Williams, M.; Brightsmith, D.J. Growth of Scarlet Macaw (Ara macao) chicks in southeastern Peru. Neotrop. Ornithol. 2011, 22, 143–153. [Google Scholar]
- Vigo Trauco, G. Crecimiento de Pichones de Guacamayo Escarlata, Ara macao (Linneus: 1758) en la Reserva Nacional Tambopata—Madre de Dios—Perú. Bachelor’s Thesis, Universidad Nacional Agraria La Molina, Lima, Peru, 2007. [Google Scholar]
- Smith, G.A. Systematics of parrots. Ibis 1975, 117, 99. [Google Scholar] [CrossRef]
- Raso, T.D.; Seixas, G.H.F.; Guedes, N.M.R.; Pinto, A.A. Chlamydophila psittaci in free-living Blue-fronted Amazon parrots (Amazona aestiva) and Hyacinth macaws (Anodorhynchus hyacinthinus) in the Pantanal of Mato Grosso do Sul, Brazil. Vet. Microbiol. 2006, 117, 235–241. [Google Scholar] [CrossRef]
- Sanz, V.; Rojas-Suárez, F. Los nidos nodriza como técnica para incrementar el reclutamiento de la cotorra cabeciamarilla (Amazona barbadensis, Aves: Psittacidae). Vida Silv. Neotrop. 1997, 6, 8–14. [Google Scholar]
- Stoodley, A.A. Surrogate Parents. In AFA Watchbid; American Federation of Aviculture: Austin, TX, USA, 1986; Volume 13, p. 6. Available online: https://journals.tdl.org/watchbird/index.php/watchbird/article/view/2455 (accessed on 1 October 2016).
- Dingle, S. Olde Tymer Bill Rattray: King of the King Parrots. In AFA Wathcbird; American Federation of Aviculture: Austin, TX, USA, 1998; Volume 25, Available online: https://journals.tdl.org/watchbird/index.php/watchbird/article/view/1407 (accessed on 1 October 2016).
- Yantz, J. Notes on fostering finches. In AFA Watchbird Magazine; American Federationof Aviculture: Austin, TX, USA, 1986; Available online: https://journals.tdl.org/watchbird/index.php/watchbird/article/view/2299 (accessed on 1 October 2016).
- Krebs, E.A.; Cunningham, R.B.; Donnelly, C.F. Complex patterns of food allocation in asynchronously hatching broods of Crimson Rosellas. Anim. Behavior 1999, 57, 753–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forshaw, J.M. Parrots of the World, 3rd ed.; Landsdowne Editions: Melbourne, Australia, 1989; p. 672. [Google Scholar]
- Iñigo-Elias, E.E. Ecology and Breeding Biology of the Scarlet Macaw (Ara macao) in the Usumacinta Drainage Basin of Mexico and Guatemala. Ph.D. Dissertation, University of Florida, Gainesville, FL, USA, 1996. [Google Scholar]
- Wiedenfeld, D.A. A new subspecies of Scarlet Macaw and its status and conservation. Ornitol. Neotrop. 1994, 5, 99–104. [Google Scholar]
- Portillo Reyes, H. Distribucion actual de la guara (lapa) roja (Ara macao) en Honduras. Zeledonia 2005, 9, 3. [Google Scholar]
- Tella, J.L.; Hiraldo, F. Illegal and legal parrot trade shows a long-term, cross-cultural preference for the most attractive species increasing their risk of extinction. PLoS ONE 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero-Vidal, P.; Hidalgo, F.; Rosetto, F.; Blanco, G.; Carrete, M.; Tella, J.L. Opportunistic or Non-random Wildlife Crime? Attractiveness rather than Abundances in the Wild Leads to Selective Parrot Poaching. Diversity 2020, 12, 314. [Google Scholar] [CrossRef]
- Vigo, G. Scarlet Macaw Nesting Ecology and Behavior: Implications for Conservation Management. Ph.D.Thesis, Texas A&M University, College Station, TX, USA, 2020. [Google Scholar]
- Bentin, M.; Leyva, J.C. Model of assisted breeding of two Primolious couloni chicks by Ara macao adults. AFA Watchbird Magazine, 5 September 2018; Volume XLV, 23–29. [Google Scholar]
- Brightsmith, D.J. Effects of diet, migration, and breeding on clay lick use by parrots in Southeastern Peru. In Proceedings of the Annual Convention 2004, American Federation of Aviculture, San Francisco, CA, USA, 31 March–3 April 2004; pp. 13–14. [Google Scholar]
- Tosi, J.A. Zonas de Vida Natural en el Perú. Memoria Explicativa Sobre el Mapa Ecológico del Perú; Instituto Interamericano de las Ciencias Agricolas de la Organización de los Estados Americanos: Lima, Peru, 1960; p. 271. [Google Scholar]
- Olah, G.; Vigo, G.; Heinsohn, R.; Brightsmith, D.J. Nest site selection and efficacy of artificial nests for breeding success of Scarlet Macaws (Ara macao) in lowland Peru. J. Nat. Conserv. 2014, 22, 176–185. [Google Scholar] [CrossRef]
- Brightsmith, D.J.; Holle, K.M.; Stronza, A. Ecotourism, conservation biology, and volunteer tourism: A mutually beneficial triumvirate. Biol. Conserv. 2008, 141, 2832–2842. [Google Scholar] [CrossRef]
- Gish, E. The Case for Bio-Centric Development: An Ethnographic Study of the Tambopata Macaw Project, Ecotourism and Volunteer Tourism in the Peruvian Amazon. Master’s Dissertation, Wageningen University, Wageningen, The Netherlands, 2009. [Google Scholar]
- Brightsmith, D.J. The Tambopata Macaw Project: Developing techniques to increase reproductive success of large macaws. AFA Watchbird 2001, 28, 24–32. [Google Scholar]
- Perry, D.R.; Williams, J. The tropical rain forest canopy: A method providing total access. Biotropica 1981, 13, 283–285. [Google Scholar] [CrossRef]
- Perry, D.R. A method of access into the crowns of emergent and canopy trees. Biotropica 1978, 10, 155–157. [Google Scholar] [CrossRef]
- Voren, H.; Jordan, R. Parrots: Hand Feeding & Nursery Management; Mattachione, Silvio, & Company: Toronto, ON, Cananda, 1992. [Google Scholar]
- Brightsmith, D.J.; Hilburn, J.; Del Campo, A.; Boyd, J.; Frisius, M.; Frisius, R.; Janik, D.; Guillén, F. The use of hand-raised Psittacines for reintroduction: A case study of Scarlet Macaws (Ara macao) in Peru and Costa Rica. Biol. Conserv. 2005, 121, 465–472. [Google Scholar] [CrossRef]
- Ricklefs, R.E. Patterns of growth in birds. Ibis 1968, 419–451. [Google Scholar] [CrossRef]
- Krebs, E. Begging and food distribution in Crimson Rosella (Platycercus elegans) broods: Why dont hungry chicks beg more? Behav. Ecol. Sociobiol. 2001, 50, 10. [Google Scholar] [CrossRef]
- Renton, K.; Brightsmith, D.J. Cavity use and reproductive success of nesting macaws in lowland forest of southeast Peru. J. Field Ornithol. 2009, 80, 1–8. [Google Scholar] [CrossRef]
- Renton, K. Agonistic interactions of nesting and nonbreeding macaws. Condor 2004, 106, 354–362. [Google Scholar] [CrossRef]
- Stoleson, S.H.; Beissinger, S.R. Hatching asynchrony, brood reduction, and food limitation in a Neotropical parrot. Ecol. Monogr. 1997, 76, 131–154. [Google Scholar] [CrossRef]
- Schew, W.A.; Ricklefs, R.E. Developmental plasticity. In Avian Growth and Development-Evolution within the Altricial-Precocial Spectrum; Starck, M., Ricklefs, R.E., Eds.; Oxford University Press: Oxford, UK, 1998; pp. 288–303. [Google Scholar]
- Brightsmith, D.J.; Matsufuji, D.; McDonald, D.; Bailey, C.A. Nutritional content of free-living Scarlet Macaw chick diets in southeastern Peru. J. Avian Med. Surg. 2010, 24, 9–23. [Google Scholar] [CrossRef]
- Cornejo, J.; Dierenfeld, E.S.; Bailey, C.A.; Brightsmith, D.J. Nutritional and physical characteristics of commercial hand-feeding formulas for parrots. Zoo Biol. 2013. [Google Scholar] [CrossRef]
- Clubb, S.L.; Clubb, K.J.; Skidmore, D.; Wolf, S.; Phillips, A. Psittacine neonatal care and hand-feeding. In Psittacine Aviculture: Perspectives, TECHNIQUES and Research; Shubot, R.M., Clubb, K.J., Clubb, S.L., Eds.; Avicultural Breeding and Research Center: Loxahatchee, FL, USA, 1992. [Google Scholar]
- Myers, M.; Vaughan, C. Movement and behavior of Scarlet Macaws (Ara macao) during the post-fledging dependence period: Implications for in situ versus ex situ management. Biol. Conserv. 2004, 118, 9. [Google Scholar] [CrossRef]
- Boyd, J.; McNab, R.B. The Scarlet Macaw in Guatemala and El Salvador: 2008 status and future possibilities. Findings and Recommendations from a Species Recovery Workshop 9–15 March 2008; Wildlife Conservation Society-Guatemala Program: Guatemala City and Flores City, Peten, Guatemala, 2008; p. 178. [Google Scholar]
- Clubb, S.L. The role of private avicultre in the conservation of Neotropical psittacines. In New World Parrots in Crisis; Snyder, N.F.R., Beissinger, S.R., Eds.; Smithsonian: Washington, DC, USA, 1992; pp. 117–132. [Google Scholar]
- Clubb, K.J.; Clubb, S.L. Status of macaws in aviculture. In Psittacine Aviculture: Perspectives, Techniques and Research; Shubot, R.M., Clubb, K.J., Clubb, S.L., Eds.; Avicultural Breeding and Research Center: Loxahatchee, FL, USA, 1992. [Google Scholar]
- Abramson, J.; Spear, B.L.; Thomsen, J.B. The Large Macaws: Their Care, Breeding and Conservation; Raintree Publications: Fort Bragg, CA, USA, 1995. [Google Scholar]
- Brightsmith, D.J. Parrot nesting in southeastern Peru: Seasonal patterns and keystone trees. Wilson Bull. 2005, 117, 296–305. [Google Scholar] [CrossRef]
- Olah, G.; Vigo, G.; Ortiz, L.; Rozsa, L.; Brightsmith, D.J. Philornis sp. bot fly larvae in free living Scarlet macaw nestlings and a new technique for their extraction. Vet. Parasitol. 2013. [Google Scholar] [CrossRef]
- Juniper, T. Spix’s Macaw: The Race to Save the World’s Rarest Bird; Fourth Estate: London, UK, 2003; p. 296. [Google Scholar]
- Schischakin, N. Special Report: The Spix’s Macaw Conservation Program. AFA Watchb. 1999, 26, 46–55. Available online: https://journals.tdl.org/watchbird/index.php/watchbird/article/view/1398 (accessed on 1 October 2016).
Figure 1.
Nest check of an artificial Scarlet Macaw nest (wooden box) using a single rope climbing system. Both macaws are displaying nest defense behaviors. A stuffed leather glove attached to a metal stick is used as a tool to cover the nest entrance and prevent macaws from reentering the nest. In the picture, the researcher access door is open. Researcher: Gustavo Martinez Sovero MSc. Photo Credit: Liz Villanueva Paipay.
Figure 1.
Nest check of an artificial Scarlet Macaw nest (wooden box) using a single rope climbing system. Both macaws are displaying nest defense behaviors. A stuffed leather glove attached to a metal stick is used as a tool to cover the nest entrance and prevent macaws from reentering the nest. In the picture, the researcher access door is open. Researcher: Gustavo Martinez Sovero MSc. Photo Credit: Liz Villanueva Paipay.

Figure 2.
Developmental stages of Scarlet Macaw chicks. (a) Stage 1: 0 to 2 days old; (b) Stage 2A: from 3 to 18 day old; (c) Stage 2B: 19 to 33 days old; (d) Stage 3: 34 to 65 days old; (e) Stage 4: 66 days old to fledge. Photo credits: The Macaw Society- Sociedad Pro Guacamayos. For more details, see Table 2.
Figure 2.
Developmental stages of Scarlet Macaw chicks. (a) Stage 1: 0 to 2 days old; (b) Stage 2A: from 3 to 18 day old; (c) Stage 2B: 19 to 33 days old; (d) Stage 3: 34 to 65 days old; (e) Stage 4: 66 days old to fledge. Photo credits: The Macaw Society- Sociedad Pro Guacamayos. For more details, see Table 2.

Figure 3.
Acceptance of Scarlet Macaw foster chicks during the first 10 days after placement in foster nests. Acceptance of foster chicks in their new nests was measured by counting feedings per day of both foster and resident chicks and calculating daily feeding ratios (n = 10 chick pairs). Each point on the graph corresponds to the feeding ratio in one foster nest on one day. The solid line represents the positive linear trend observed and shows that foster chick feedings increased during the first 10 days in the foster nest. Day one on the X-axis indicates the day of relocation. These data are from the recorded observations (see Appendix B and Methods for additional descriptions of these data).
Figure 3.
Acceptance of Scarlet Macaw foster chicks during the first 10 days after placement in foster nests. Acceptance of foster chicks in their new nests was measured by counting feedings per day of both foster and resident chicks and calculating daily feeding ratios (n = 10 chick pairs). Each point on the graph corresponds to the feeding ratio in one foster nest on one day. The solid line represents the positive linear trend observed and shows that foster chick feedings increased during the first 10 days in the foster nest. Day one on the X-axis indicates the day of relocation. These data are from the recorded observations (see Appendix B and Methods for additional descriptions of these data).

Table 1.
Criteria and timing to remove Scarlet Macaw chicks from original nests for relocation procedures in Tambopata.
Table 1.
Criteria and timing to remove Scarlet Macaw chicks from original nests for relocation procedures in Tambopata.
| Hatch Order of Removed Chicks and Criteria for When to Remove Chicks | Timing to Remove Chick from Original Nest | # Chicks Removed |
|---|---|---|
| First Chicks (11 chicks removed) | ||
| When second chick hatched > 3 days after first chick | As soon as second chick hatched | 7 |
| If chick showed signs of life-threatening botfly related infection | When infection was clearly getting worse but still localized in one area | 2 |
| To create conditions for a triple brood by adding a third chick as the younger brood member | When third chick was placed in foster nest | 2 |
| Second Chicks (7 chicks removed) | ||
| If chick showed signs of starvation | When chick was still active and begging. Usually within 3 days of not gaining weight as expected, before they started to lose weight | 5 |
| If chick showed signs of life-threatening botfly related infection | When infection was clearly getting worse but still localized in one area | 1 |
| If chick was needed for relocation to another nest where the clutch or brood was lost due to damage to the nest | As soon as damaged nest was fixed | 1 |
| Third Chicks (12 chicks removed) | ||
| All third chicks were removed | <24 h after hatching | 12 |
| Fourth Chick (2 chicks removed) | ||
| All fourth chicks were removed | <24 h after hatching | 2 |
| Chicks removed from their original nests | 32 | |
| Chicks that perished in our nursery before relocation | 4 | |
| Total chicks relocated | 28 | |
Table 2.
Developmental stages in wild Scarlet macaw chicks. To define the Scarlet Macaw chick developmental stages, we used a modified version of the mass growth stages presented in Vigo et al. [14]. Scarlet macaw chicks fledge on average at 88 days old (n = 104 chicks, range = 79 to 99 day old, [15]); see Figure 2 for images of each developmental stage.
Table 2.
Developmental stages in wild Scarlet macaw chicks. To define the Scarlet Macaw chick developmental stages, we used a modified version of the mass growth stages presented in Vigo et al. [14]. Scarlet macaw chicks fledge on average at 88 days old (n = 104 chicks, range = 79 to 99 day old, [15]); see Figure 2 for images of each developmental stage.
| Developmental Stages in Wild Scarlet Macaw Chicks | ||
|---|---|---|
| Stage | Age Range | Description |
| Stage 1 | 0 to 2 | Hatchling |
| Stage 2A | 3 to 18 | Naked to light pinfeathers and eyes closed |
| Stage 2B | 19 to 33 | Light pinfeathers to heavy pinfeathers and eyes open |
| Stage 3 | 34 to 65 | Heavy pinfeathers to mostly feathered |
| Stage 4 | 66 to fledged | Mostly feathered to fully feathered |
Table 3.
Macaw foster chick relocation procedures and timing in Tambopata, Peru. Procedures outlined below were designed to evaluate the capacity of nesting wild Scarlet Macaws to accept and raise foster chicks. “Foster nest requirements” refers to the characteristics of nests needed to be eligible to foster chicks. “Foster chick age at relocation criterion” was established considering the reproductive ecology and behavior of the species [29]. In smaller font, below the number of foster chicks is the number of foster chicks in nests with video cameras. The number of foster broods is less than the number of foster chicks because some foster broods contained two foster chicks.
Table 3.
Macaw foster chick relocation procedures and timing in Tambopata, Peru. Procedures outlined below were designed to evaluate the capacity of nesting wild Scarlet Macaws to accept and raise foster chicks. “Foster nest requirements” refers to the characteristics of nests needed to be eligible to foster chicks. “Foster chick age at relocation criterion” was established considering the reproductive ecology and behavior of the species [29]. In smaller font, below the number of foster chicks is the number of foster chicks in nests with video cameras. The number of foster broods is less than the number of foster chicks because some foster broods contained two foster chicks.
| Macaw Chick Relocation Procedures | Hypothesis | Foster Nest Requirements | Foster Chick Age at Relocation Criterion | Timing to Place Foster Chick | Foster Chick Age at Relocation | # Foster Broods | # Foster Chicks | # Foster Chicks that Fledged | |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Acceptance | Wild macaws will foster unrelated chicks and fledge them as their own | Nest with single resident chick | ≤2 days older or younger than resident chick | Within 24 h of both foster and resident chicks, opening their eyes | ~18 days old | 9 | 11 5 w/video | 9 |
| 2 | Age difference | Age difference is the absolute driver of death by starvation | Nest with single resident chick | From 4 to 9 days older or younger than the resident chick. | Within 48 h of the resident chick opening its eyes | ~18 days old | 9 | 10 7 w/video | 10 |
| 3 | Empty nest | Wild macaws will foster and fledge chicks after losing their own brood or clutch | Nest that lost eggs due to depredation but pair was still incubating an artificial egg | Foster chick was a solo chick | At expected hatching day | <3 days old | 2 | 2 1 w/video | 4 |
| Nest that lost chicks due to depredation or accident (lighting) | Similar developmental stage as lost chicks | <36 h of original brood disappearance | <46 days old | 2 | 3 No video | ||||
| 4 | Triple brood | Wild macaws will accept an additional chick in a brood of two chicks and fledge three chicks. | Nest that hatched three chicks | <5 days between first and third chick | Third chick swapped for first chick when eyes fully opened. | 18 days old | 1 | 2 No video | 2 |
| First chick was placed back in foster nest 5 days after. | 24 days old | ||||||||
Table 4.
Effect of hatch order on growth parameters for Scarlet Macaw. All parameters were calculated using the logistic growth model [14,41]. “Solo chicks” are wild chicks in one-chick broods. “First chicks” are older chicks in wild two chicks’ broods and “second chicks” are younger chicks in wild two chicks’ broods. Within a column, values followed by a different superscript letter differed significantly using a Mann–Whitney U (DF = 1, p < 0.05).
Table 4.
Effect of hatch order on growth parameters for Scarlet Macaw. All parameters were calculated using the logistic growth model [14,41]. “Solo chicks” are wild chicks in one-chick broods. “First chicks” are older chicks in wild two chicks’ broods and “second chicks” are younger chicks in wild two chicks’ broods. Within a column, values followed by a different superscript letter differed significantly using a Mann–Whitney U (DF = 1, p < 0.05).
| Chick Type | Number of Individuals | Maximum Growth | Growth | Age at Maximum Growth * | |
|---|---|---|---|---|---|
| Rate | |||||
| A | B | C | |||
| (Mean) | (Mean) | (Mean) | |||
| Wild chicks | Solo chick | 17 | 1028 ± 83.2 A | 0.126 ± 0.013 B | 24.7 ± 2.3 D |
| First chick | 38 | 1022 ± 75.4 A | 0.115 ± 0.014 B | 25.7 ± 2.4 E | |
| Second chick | 26 | 993 ± 77.7 A | 0.111 ± 0.015 C | 28.2 ± 3.2 F | |
| All combined | 81 | 1014 ± 79.7 | 0.116 ± 0.016 | 26.3 ± 3 | |
| Foster chicks | 23 | 1020 ± 81.1 A | 0.121 ± 0.014 B | 23.9 ±1.7 D | |
* The only growth parameter that differed significantly between wild chicks (all combined) and foster chicks is indicated by an asterisk, based on Wilcoxon pairwise comparisons (DF = 1, p < 0.05).
Table 5.
Effect of foster nests on seasonal breeding success of Scarlet Macaw. Seasons with no foster chicks (17 seasons, 2000 to 2016) were compared to seasons with chick fostering (3 seasons, 2017 to 2019). In the fostering experiments, we placed, swapped or added, foster chicks to specific nests (see text). The breeding season was mid-October to mid-April. Available nests refer to cavities (n = 40) that were not occupied by other species by the beginning of the breeding season. We calculated χ2 and p-values using Mann–Whitney U (DF = 1).
Table 5.
Effect of foster nests on seasonal breeding success of Scarlet Macaw. Seasons with no foster chicks (17 seasons, 2000 to 2016) were compared to seasons with chick fostering (3 seasons, 2017 to 2019). In the fostering experiments, we placed, swapped or added, foster chicks to specific nests (see text). The breeding season was mid-October to mid-April. Available nests refer to cavities (n = 40) that were not occupied by other species by the beginning of the breeding season. We calculated χ2 and p-values using Mann–Whitney U (DF = 1).
| Breeding Success Parameters | Seasons with Foster Nests (n = 3) | Seasons without Foster Nests (n = 17) | χ2 | p-Value |
|---|---|---|---|---|
| Chicks that fledged per available cavity | 0.43 ± 0.05 | 0.23 ± 0.86 | 7.2 | 0.036 |
| Chicks that fledged per nest with eggs | 1.13 ± 0.15 | 0.56 ± 0.21 | 7.1 | 0.007 |
| Chicks that fledged per nest where at least one chick hatched | 1.5 ± 0.3 | 0.86 ± 0.24 | 7.1 | 0.01 |
| Percentage of younger chicks that starved | 0.06 ± 0.03 | 0.35 ± 0.22 | 5.5 | 0.02 |
| Percentage of chicks that fledged | 0.7 ± 0.05 | 0.46 ± 0.14 | 5.5 | 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Vigo-Trauco, G.; Garcia-Anleu, R.; Brightsmith, D.J. Increasing Survival of Wild Macaw Chicks Using Foster Parents and Supplemental Feeding. Diversity 2021, 13, 121. https://0-doi-org.brum.beds.ac.uk/10.3390/d13030121
AMA Style
Vigo-Trauco G, Garcia-Anleu R, Brightsmith DJ. Increasing Survival of Wild Macaw Chicks Using Foster Parents and Supplemental Feeding. Diversity. 2021; 13(3):121. https://0-doi-org.brum.beds.ac.uk/10.3390/d13030121
Chicago/Turabian StyleVigo-Trauco, Gabriela, Rony Garcia-Anleu, and Donald J. Brightsmith. 2021. "Increasing Survival of Wild Macaw Chicks Using Foster Parents and Supplemental Feeding" Diversity 13, no. 3: 121. https://0-doi-org.brum.beds.ac.uk/10.3390/d13030121
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.