
Multivariate Abundance Analysis of Multi-Host/Multi-Parasite Lungworms in a Sympatric Wild Ruminant Population


 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
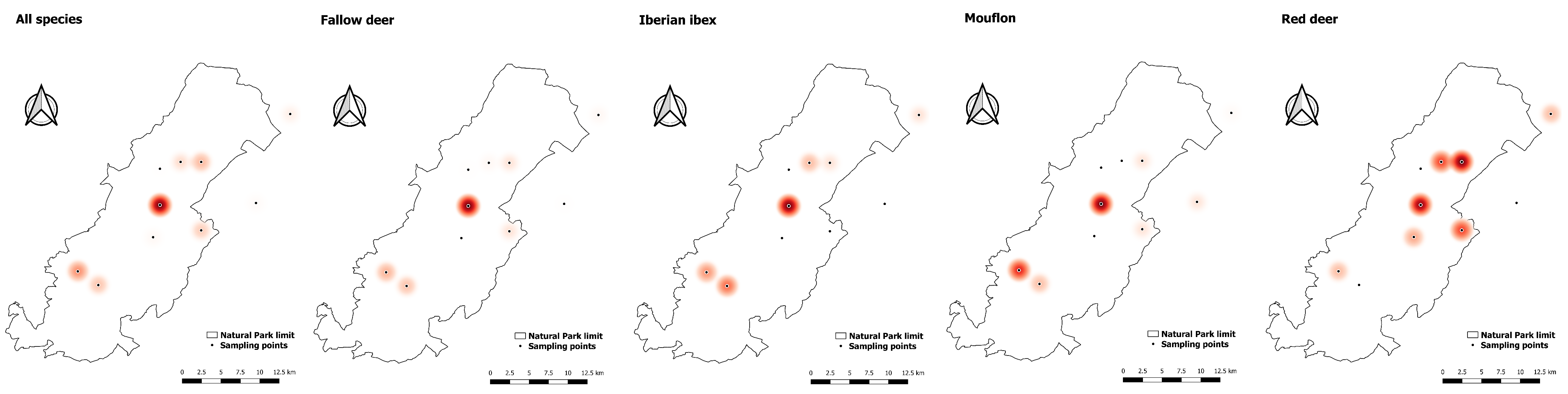
2.1. Study Area and Wild Ruminants
2.2. Sampling Protocol
2.3. Epidemiological Parameters and Statistical Analysis
3. Results
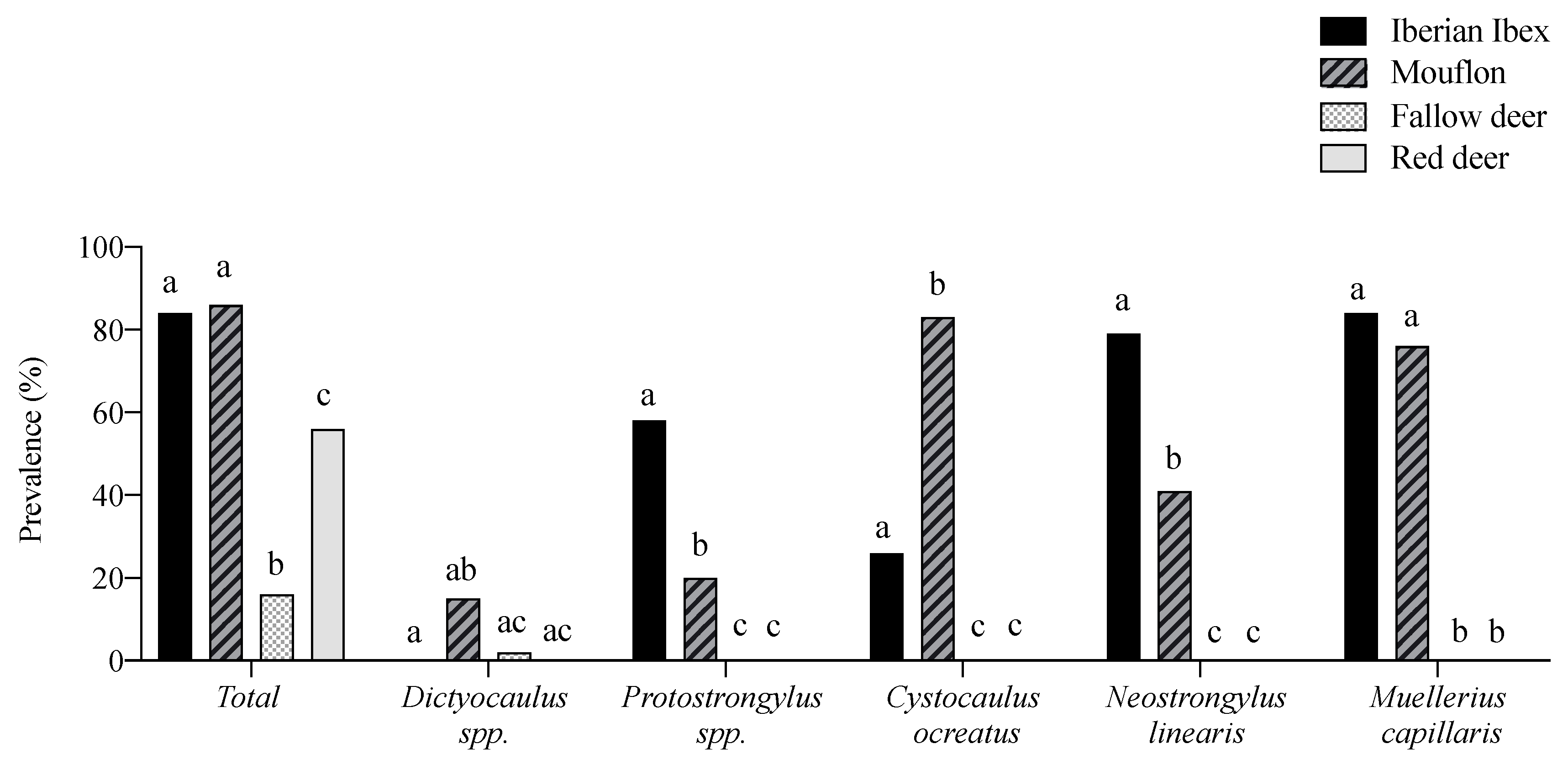
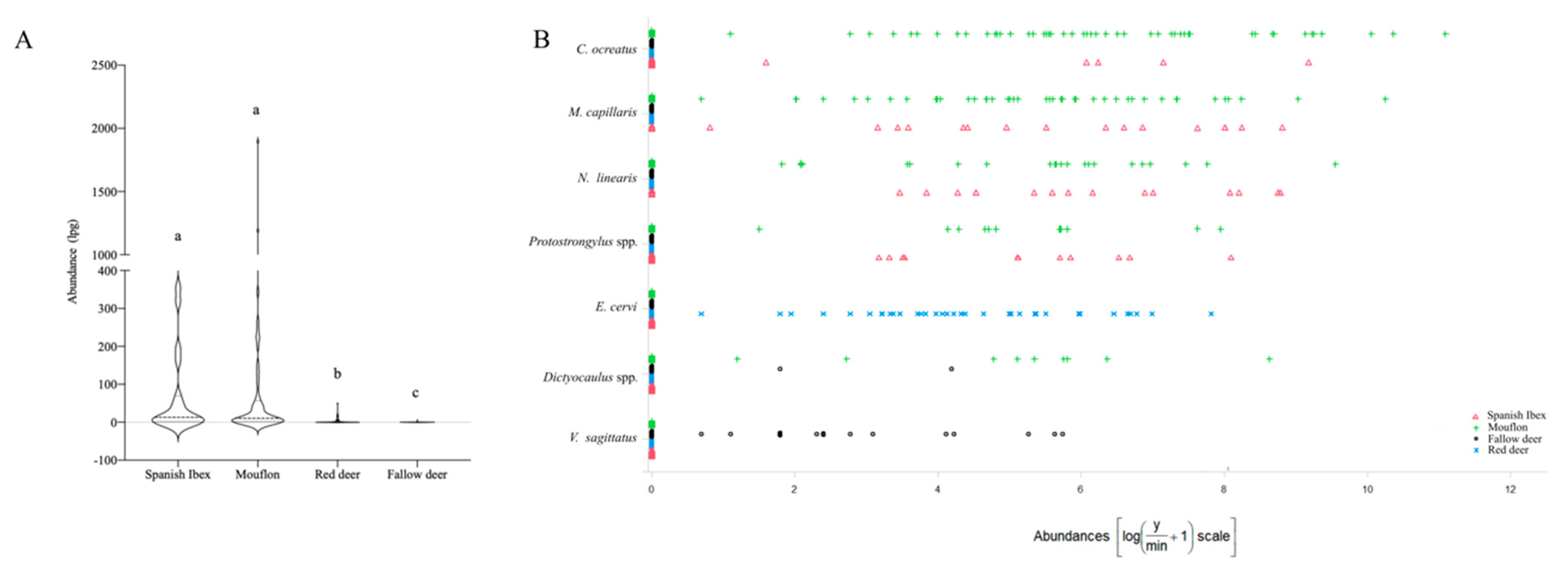
3.1. Overall Descriptive Patterns of the Lungworms
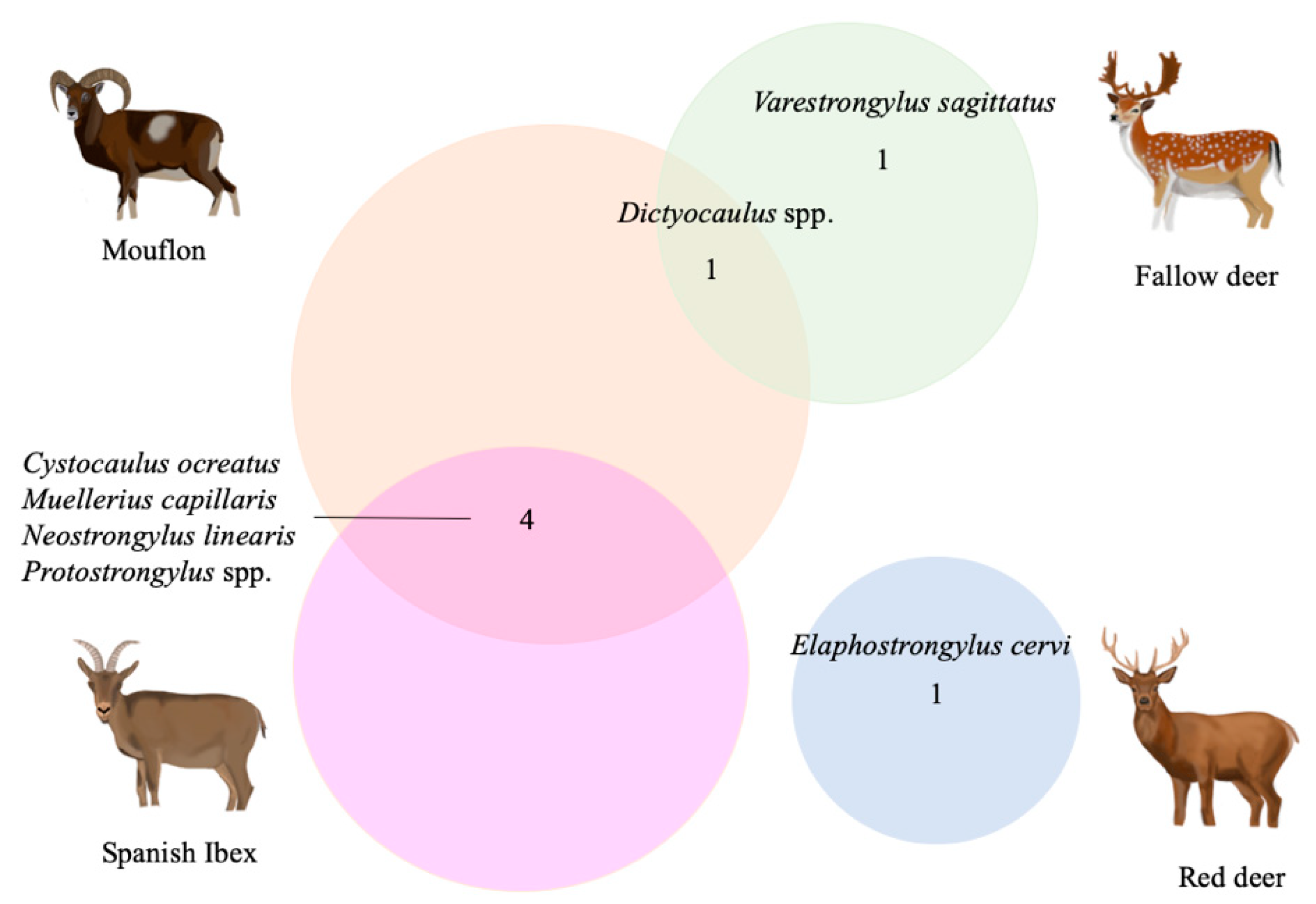
3.2. Multi-Host Lungworm Distribution
3.3. Single-Host Lungworms Distribution
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Panayotova-Pencheva, M.S.; Alexandrov, M.T. Some Pathological Features of Lungs from Domestic and Wild Ruminants with Single and Mixed Protostrongylid Infections. Vet. Med. Int. 2010, 2010, e741062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoberg, E.P.; Kocan, A.A.; Rickard, L.G. Gastrointestinal Strongyles in Wild Ruminants. In Parasitic Diseases of Wild Mammals; Samuel, W.M., Pybus, M.J., Kocan, A.A., Eds.; Manson Publishing/Veterinary Press: London, UK, 2001; pp. 193–227. [Google Scholar]
- Gunn, A.; Irvine, R.J. Subclinical Parasitism and Ruminant Foraging Strategies: A Review. Wildl. Soc. Bull. 2003, 31, 117–126. [Google Scholar]
- Jenkins, E.J.; Veitch, A.M.; Kutz, S.J.; Bollinger, T.K.; Chirino-Trejo, J.M.; Elkin, B.T.; West, K.H.; Hoberg, E.P.; Polley, L. Protostrongylid Parasites and Pneumonia in Captive and Wild Thinhorn Sheep (Ovis dalli). J. Wildl. Dis. 2007, 43, 189–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutz, S.J.; Hoberg, E.P.; Polley, L. A New Lungworm in Muskoxen: An Exploration in Arctic Parasitology. Trends Parasitol. 2001, 17, 276–280. [Google Scholar] [CrossRef] [Green Version]
- Panadero, R.; Carrillo, E.B.; López, C.; Díez-Baños, N.; Díez-Baños, P.; Morrondo, M.P. Bronchopulmonary Helminths of Roe Deer (Capreolus capreolus) in the Northwest of Spain. Vet. Parasitol. 2001, 99, 221–229. [Google Scholar] [CrossRef]
- Böhm, M.; White, P.C.L.; Daniels, M.J.; Allcroft, D.J.; Munro, R.; Hutchings, M.R. The Health of Wild Red and Sika Deer in Scotland: An Analysis of Key Endoparasites and Recommendations for Monitoring Disease. Vet. J. 2006, 171, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Carreno, R.A.; Diez-Baños, N.; del Rosario Hidalgo-Argüello, M.; Nadler, S.A. Characterization of Dictyocaulus Species (Nematoda: Trichostrongyloidea) from Three Species of Wild Ruminants in Northwestern Spain. J. Parasitol. 2009, 95, 966–970. [Google Scholar] [CrossRef] [Green Version]
- Díez-Baños, P.; Morrondo-Pelayo, P.; Feijoo-Penela, A.; Carrillo-González, B.; López-Sández, C. Relationship between the Excretion of Protostrongylid Larvae in Sheep in North-West Spain and Climatic Conditions. J. Helminthol. 1994, 68, 197–201. [Google Scholar] [CrossRef]
- López, C.M.; Fernández, G.; Viña, M.; Cienfuegos, S.; Panadero, R.; Vázquez, L.; Díaz, P.; Pato, J.; Lago, N.; Dacal, V.; et al. Protostrongylid Infection in Meat Sheep from Northwestern Spain: Prevalence and Risk Factors. Vet. Parasitol. 2011, 178, 108–114. [Google Scholar] [CrossRef]
- Winter, J.; Rehbein, S.; Joachim, A. Transmission of Helminths between Species of Ruminants in Austria Appears More Likely to Occur than Generally Assumed. Front. Vet. Sci. 2018, 5, 30. [Google Scholar] [CrossRef] [Green Version]
- Carrau, T.; Martínez-Carrasco, C.; Garijo, M.M.; Alonso, F.; León-Vizcaíno, L.; Herrera-Russert, J.; Tizzani, P.; Ruiz De Ybáñez, R. Epidemiological Approach to Nematode Polyparasitism Occurring in a Sympatric Wild Ruminant Multi-Host Scenario. J. Helminthol. 2021. accepted manuscript. [Google Scholar]
- Delogu, M.; Ghetti, G.; Gugiatti, A.; Cotti, C.; Piredda, I.; Frasnelli, M.; De Marco, M.A. Virological Investigation of Avian Influenza Virus on Postglacial Species of Phasianidae and Tetraonidae in the Italian Alps. ISRN Vet. Sci. 2013, 2013, 601732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leivesley, J.A.; Bussière, L.F.; Pemberton, J.M.; Pilkington, J.G.; Wilson, K.; Hayward, A.D. Survival Costs of Reproduction Are Mediated by Parasite Infection in Wild Soay Sheep. Ecol. Lett. 2019, 22, 1203–1213. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.F.; Sax, D.F.; Lafferty, K.D. Evidence for the Role of Infectious Disease in Species Extinction and Endangerment. Conserv. Biol. 2006, 20, 1349–1357. [Google Scholar] [CrossRef] [PubMed]
- Fandos, P. La Cabra Montés (Capra pyrenaica) en el Parque Natural de las Sierras de Cazorla, Segura y las Villas; ICONA, D.L.: Madrid, Spain, 1991; p. 176. [Google Scholar]
- IGME Relieve y Climatología. Atlas Hidrogeológico de la Provincia de Jaén; Diputación Provincial De Jaén: Jaén, Spain, 1997; Volume 76, pp. 19–22. ISBN mkt0006228463. [Google Scholar]
- Carrau, T.; Martínez-Carrasco, C.; Garijo, M.M.; Alonso, F.; de Ybáñez, R.R.; Tizzani, P. Evaluation of the Baermann–Wetzel Method for Detecting Lungworm Larvae in Wild Ruminants from Faecal Samples. J. Helminthol. 2021, 95, E13. [Google Scholar] [CrossRef] [PubMed]
- Commonwealth Institute of Helminthology; Anderson, R.C.; Chabaud, A.; Willmott, S. CIH Keys to the Nematode Parasites of Vertebrates; Commonwealth Agricultural Bureaux; Farnham Royal: Wallingford, UK, 1974; p. 480. [Google Scholar]
- Margolis, L.; Esch, G.W.; Holmes, J.C.; Kuris, A.M.; Schad, G.A. The Use of Ecological Terms in Parasitology (Report of an Ad Hoc Committee of the American Society of Parasitologists). J. Parasitol. 1982, 68, 131–133. [Google Scholar] [CrossRef]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology Meets Ecology on Its Own Terms: Margolis et al., Revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Wang, Y.; Naumann, U.; Wright, S.T.; Warton, D.I. Mvabund—An R Package for Model-Based Analysis of Multivariate Abundance Data. Methods Ecol. Evol. 2012, 3, 471–474. [Google Scholar] [CrossRef]
- RStudio Team. Studio: Integrated Development for R; RStudio, Inc.: Boston, MA, USA, 2015. [Google Scholar]
- Alasaad, S.; Morrondo, P.; Dacal-Rivas, V.; Soriguer, R.C.; Granados, J.E.; Serrano, E.; Zhu, X.Q.; Rossi, L.; Pérez, J.M. Bronchopulmonary Nematode Infection of Capra pyrenaica in the Sierra Nevada Massif, Spain. Vet. Parasitol. 2009, 164, 340–343. [Google Scholar] [CrossRef] [Green Version]
- Luzón, M.; Moreno, J.S.; Meana, A.; Díaz, A.T.; Pastor, A.P.; Brunet, A.G.; Sebastián, A.L. Parasitism and Horn Quality in Male Spanish Ibex (“Capra pyrenaica hispanica”) from Andalucía Based on Coprological Analysis and Muscle Biopsy. Span. J. Agric. Res. 2008, 353–361. [Google Scholar] [CrossRef] [Green Version]
- Meana, A.; Luzón-Peña, M.; Santiago-Moreno, J.; De Bulnes, A.; Gómez-Bautista, M. Natural Infection by Gastrointestinal and Bronchopulmonary Nematodes in Mouflons (Ovis musimon) and Their Response to Netobimin Treatment. J. Wildl. Dis. 1996, 32, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Panayotova-Pencheva, M.S. New Records of Protostrongylid Lungworms from Wild Ruminants in Bulgaria. Vet. Med. Praha. 2006, 51, 477–484. [Google Scholar]
- Kowal, J.; Kornaś, S.; Nosal, P.; Basiaga, M.; Wajdzik, M.; Skalska, M.; Wyrobisz, A. Lungworm (Nematoda: Protostrongylidae) Infection in Wild and Domestic Ruminants from Małopolska Region of Poland. Ann. Parasitol. 2016, 62, 63–66. [Google Scholar] [CrossRef] [PubMed]
- San Miguel, J.; Álvarez, G.; Luzón, M. Procesos Parasitarios Detectados En Ciervos (Cervus elaphus) Abatidos En La Provincia de Toledo. Galemys 2001, 13, 11–24. [Google Scholar]
- Redjadj, C.; Darmon, G.; Maillard, D.; Chevrier, T.; Bastianelli, D.; Verheyden, H.; Loison, A.; Saïd, S. Intra- and Interspecific Differences in Diet Quality and Composition in a Large Herbivore Community. PLoS ONE 2014, 9, e84756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago-Moreno, J.; Toledano-Díaz, A. El muflón europeo (Ovis orientalis musimon Schreber, 1782) en España: Consideraciones. Galemys 2004, 16, 3–20. [Google Scholar]
- Martínez, T.; Fandos, P. Solapamiento Entre La Dieta de La Cabra Montés (Capra pyrenaica) y La Del Muflón (Ovis musimon). Doñana. Acta Vertebr. 1989, 16, 315–318. [Google Scholar]
- Miranda, M.; Sicilia, M.; Bartolomé, J.; Molina-Alcaide, E.; Gálvez-Bravo, L.; Cassinello, J. Contrasting Feeding Patterns of Native Red Deer and Two Exotic Ungulates in a Mediterranean Ecosystem. Wildl. Res. 2012, 39, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Ezenwa, V.O. Habitat Overlap and Gastrointestinal Parasitism in Sympatric African Bovids. Parasitology 2003, 126, 379–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- León-Vizcaíno, L.; Ruíz de Ybáñez, M.R.; Cubero, M.J.; Ortíz, J.M.; Espinosa, J.; Pérez, L.; Simón, M.A.; Alonso, F. Sarcoptic Mange in Spanish Ibex from Spain. J. Wildl. Dis. 1999, 35, 647–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fandos, G.; Enrique, J.; Granados-Torres, J.E.; Burón, D.; Vega, E.; Perez, J.M.; Soriguer, R.C.; Fandos, P. Changes in the Co-Occurrence of the Ungulates of Cazorla after the Collapse of the Spanish Ibex. In Proceedings of the 7th World Mountain Ungulate Conference, Bozeman, MT, USA, 10–13 September 2019. [Google Scholar]
- Cabaret, J. Réceptivité Des Mollusques Terrestres de La Région de Rabat à l’infestation Par Les Protostrongles Dans Les Conditions Expérimentales et Naturelles. Ph.D. Thesis, Univeristy of Paris, Paris, France, 1981. [Google Scholar]
- Handeland, K.; Slettbakk, T. Outbreaks of Clinical Cerebrospinal Elaphostrongylosis in Reindeer (Rangifer tarandus tarandus) in Finnmark, Norway, and Their Relation to Climatic Conditions. J. Vet. Med. Ser. B 1994, 41, 407–410. [Google Scholar] [CrossRef]
- Vicente, J.; Fierro, Y.; Gortazar, C. Seasonal Dynamics of the Fecal Excretion of Elaphostrongylus cervi (Nematoda, Metastrongyloidea) First-Stage Larvae in Iberian Red Deer (Cervus elaphus hispanicus) from Southern Spain. Parasitol. Res. 2005, 95, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Vicente, J.; de Mera, I.G.F.; Gortazar, C. Epidemiology and Risk Factors Analysis of Elaphostrongylosis in Red Deer (Cervus elaphus) from Spain. Parasitol. Res. 2006, 98, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Nocture, M.; Cabaret, J.; Hugonnet-Chapelle, L. Protostrongylid Nematode Infection of Chamois (Rupicapra rupicapra) At The Bauges Massif (French Alps). Vet. Parasitol. 1998, 77, 153–161. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total | Iberian Ibex n = 19 | Mouflon n = 59 | Fallow Deer n = 108 | Red Deer n = 64 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P (%) | I.R. | P (%) | A | I.R. | P (%) | A | I.R. | P (%) | A | I.R. | P (%) | A | I.R. | |
| Total | 48.0 | 0–1896.0 | 84.2 | 6.59 | 0.1–35.5 | 86.4 | 106.2 | 0.1–189 | 15.7 | 2.0 | 0.1–0.6 | 56.3 | 3.0 | 0.1–49.5 |
| C. ocreatus | 22.0 | 0.1–1309.0 | 26.3 | 12.5 | 01–193.5 | 83.1 | 71.9 | 0.1–1309 | -- | -- | -- | -- | -- | -- |
| Dictyocaulus spp. | 4.4 | 0.1–111.7 | 0.0 | 0.0 | 0.0 | 15.3 | 2.5 | 0.1–111.7 | 1.8 | 0.1 | 0.1–1.3 | 0.0 | 0.0 | 0.0 |
| E. cervi | 14.4 | 0.1–49.5 | -- | -- | -- | -- | -- | -- | -- | -- | -- | 56.3 | 0.3 | 0.1–49.5 |
| M. capillaris | 24.4 | 0.1–565.7 | 84.2 | 22.6 | 0.1–134.2 | 76.3 | 21.1 | 0.1–565.7 | -- | -- | -- | 0.0 | 0.0 | 0.0 |
| N. linearis | 15.6 | 0.1–280.1 | 78.9 | 24.6 | 0.6–130.6 | 40.7 | 8.5 | 0.1–280.1 | -- | -- | -- | 0.0 | 0.0 | 0.0 |
| Protostrongylus spp. | 9.2 | 0.1–65.2 | 57.9 | 6.1 | 0.5–65.2 | 20.3 | 2.2 | 0.1–56.4 | -- | -- | -- | 0.0 | 0.0 | 0.0 |
| V. sagittatus | 6.0 | 0.1–6.2 | -- | -- | -- | -- | -- | -- | 13.9 | 0.2 | 0.1–6.2 | 0.0 | 0.0 | 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carrau, T.; Martínez-Carrasco, C.; Garijo, M.M.; Alonso, F.; de Ybáñez, R.R.; Tizzani, P. Multivariate Abundance Analysis of Multi-Host/Multi-Parasite Lungworms in a Sympatric Wild Ruminant Population. Diversity 2021, 13, 227. https://0-doi-org.brum.beds.ac.uk/10.3390/d13060227
Carrau T, Martínez-Carrasco C, Garijo MM, Alonso F, de Ybáñez RR, Tizzani P. Multivariate Abundance Analysis of Multi-Host/Multi-Parasite Lungworms in a Sympatric Wild Ruminant Population. Diversity. 2021; 13(6):227. https://0-doi-org.brum.beds.ac.uk/10.3390/d13060227
Chicago/Turabian StyleCarrau, Tessa, Carlos Martínez-Carrasco, María Magdalena Garijo, Francisco Alonso, Rocío Ruiz de Ybáñez, and Paolo Tizzani. 2021. "Multivariate Abundance Analysis of Multi-Host/Multi-Parasite Lungworms in a Sympatric Wild Ruminant Population" Diversity 13, no. 6: 227. https://0-doi-org.brum.beds.ac.uk/10.3390/d13060227