
Mediterranean Red Macro Algae Mats as Habitat for High Abundances of Serpulid Polychaetes




1
Marine Ecology Department, Faculty of Biology and Chemistry, University of Bremen, 28359 Bremen, Germany
2
Department of Environmental Biology, Sapienza University of Rome, 00185 Rome, Italy
*
Author to whom correspondence should be addressed.
Diversity 2021, 13(6), 265; https://0-doi-org.brum.beds.ac.uk/10.3390/d13060265
Submission received: 25 April 2021
/
Revised: 30 May 2021
/
Accepted: 8 June 2021
/
Published: 11 June 2021
(This article belongs to the Special Issue Biodiversity Conservation in Mediterranean Sea)
Abstract
:The Mediterranean Sea harbors more than 17,000 eukaryotic marine species, with several ecosystems recognized as biodiversity hotspots, such as Posidonia oceanica meadows. Recent research indicates that benthic mats formed by the fleshy red alga Phyllophora crispa are also associated with high species richness. Among key groups found in these mats are sessile polychaetes, which live as epiphytes on the red algae thalli. Knowledge of abundance, species richness, and spatial variation of polychaetes associated with these habitats is still scarce. We carried out a comparative assessment focusing on serpulid polychaetes within samples from P. crispa mats and neighboring P. oceanica meadows at six different sampling sites around Giglio Island (Tyrrhenian Sea, Italy). A total of 17 serpulid taxa were identified. The abundance of serpulids (5665 individuals m−2 of P. crispa mat) were similar to neighboring P. oceanica meadows (2304 individuals m−2 leaves and 5890 individuals m−2 shoots). The number of serpulid taxa was significantly higher in P. crispa mats (average 6.63 ± 1.32 taxa) compared to P. oceanica beds (average 1.56 ± 0.63 and 1.84 ± 1.04 taxa in leaves and shoots, respectively). Within habitat type, there were no significant differences in species richness between sites. The most abundant species found was Josephella marenzelleri (61% of individuals), while Vermiliopsis spp. and Bathyvermilia sp. were exclusively found in P. crispa samples. Our results highlight that P. crispa mats host an exceptional diversity and that these habitats should be included in conservation strategies. Further research should focus on the significance of other important taxonomic groups within these mats and evaluate the distribution of P. crispa in different regions of the Mediterranean Sea.
1. Introduction
With almost 17,000 described eukaryotic species [1], the Mediterranean Sea harbors high levels of biological diversity accompanied by a high rate of endemism (estimated 20%) accounting for the term “biodiversity hotspot” [1,2,3]. Its relatively small size compared to the world’s oceans (less than 1%), in combination with this prevalent endemism, underlines the Mediterranean Sea’s ecological relevance [4]. In particular, many Mediterranean biogenic habitats’ structural complexity (e.g., Posidonia oceanica seagrass meadows) facilitates their role as biodiversity hotspots by providing spawning grounds, nurseries, and permanent settling space for many species [5,6,7,8,9]. Furthermore, this structural complexity results in a wide range of ecological niches formed by temporal and spatial gradients (e.g., light and temperature). These gradients are often created by engineering or foundation species [10] that modify their environment through their growth (e.g., macroalgae), while others alter local conditions collectively (e.g., tropical corals). This habitat modification often results in a shift of ecological zonation (e.g., seaweeds providing wet layers in tidal zones) by mitigating stressors for inhabiting species [11]. Finally, associated mobile species often enhance the engineer’s growth by driving out competitors or protecting against grazers [12,13].
Some of the most studied biodiversity hotspots in the Mediterranean basin include Posidonia oceanica (L.) Delile, 1813 meadows (Figure 1C–E) and coralligenous habitats. The marine angiosperm P. oceanica promotes high biodiversity and provides nursery grounds for commercially important vertebrate species [14]. Furthermore, the structural complexity of the meadow serves as a secondary substrate for diverse communities [15] of sessile invertebrates (e.g., Serpulidae) [5]. Coralligenous habitats are bioconstructions primarily built by encrusting red algae and secondarily by calcifying invertebrate species, which also build a structurally complex habitat and provide biogenic substrate for a wide range of epibiota [7].
An additional potential and relatively unexplored biodiversity hotspot is the habitat created by mat-forming, fleshy red alga Phyllophora crispa (Hudson) P.S.Dixon, 1964 (Figure 1A,B) [16,17]. In the Black Sea, P. crispa mats harbor a rich community of associated fauna, including a diverse invertebrate epifauna [17]. The documented importance of these habitats led to the establishment of a marine reserve [18]. Marine polychaetes are one of the main groups of associated invertebrate epifauna of these mats [19].
P. crispa mats colonize light-exposed rock surfaces all around Giglio Island. Together with P. oceanica meadows and coralligenous reefs, it represents one of the most abundant biogenic habitats in this area. Hemisciaphilic assemblages dominated by P. crispa were reported in the Tyrrhenian Sea, both along Giglio Island [16,17] and the North-East coast of Sardinia [20,21]. While differences in diversity and abundance of associated epifauna between P. oceanica meadows and other sciaphilic (adapted to low light conditions) hardbottom communities have been reported [22,23], only a few studies describe the associated biodiversity of P. crispa mats.
Polychaetes are among the most diverse metazoan groups, with an estimated number of 2481 species in the Mediterranean Sea, accounting for 20.5% of the 12,088 species reported globally [24]. They show a wide functional variety and adaptation to different environmental conditions [25,26,27]. Polychaete abundance and diversity are often used to assess benthic communities’ states and dynamics [28].
Serpulid polychaetes are sessile organisms that colonize various marine habitats, from the shallow infralittoral to abyssal depths [29] and contain great taxonomic diversity [30]. Key drivers for the abundance and diversity of sessile polychaetes are environmental gradients (e.g., light and depth), as well as suitable space for larval settlement [19,31]. They contribute a considerable amount of carbonate bioconstructions from tropical to boreal latitudes, and shape the seafloor by acting as secondary builders [32,33]. Their ability to precipitate carbonate emphasizes their pivotal role as bioengineering species [34]. In the Mediterranean Sea, serpulids have been relatively well-studied [35,36,37,38], and several species have been recognized as lessepsian migrants (i.e., immigrated species from the Red Sea) [30,39,40,41]. Furthermore, the habitats’ structural heterogeneity provided by the algae influences serpulid assemblages (i.e., by offering settling ground and accumulating food particles), as demonstrated by Casoli et al. (2016) [19]. Therefore, the study of serpulids distribution as proxy groups for the occurrence of spatial microhabitats [42,43] is pivotal to understanding the biological diversity hosted in poorly investigated habitats, such as P. crispa mats. However, the distribution and role of serpulid worms in Mediterranean P. crispa mats are still not well understood. With this work, we aim to answer the following research questions:
- (1)
- What are the abundance and species richness of serpulid polychaetes in P. crispa mats compared to P. oceanica meadows?
- (2)
- Which species are found in both habitats and which species are unique to P. crispa mats?
- (3)
- What is the spatial variability of serpulid polychaetes associated with red algae mats?
2. Materials and Methods
2.1. Study Area and Sampling Activities
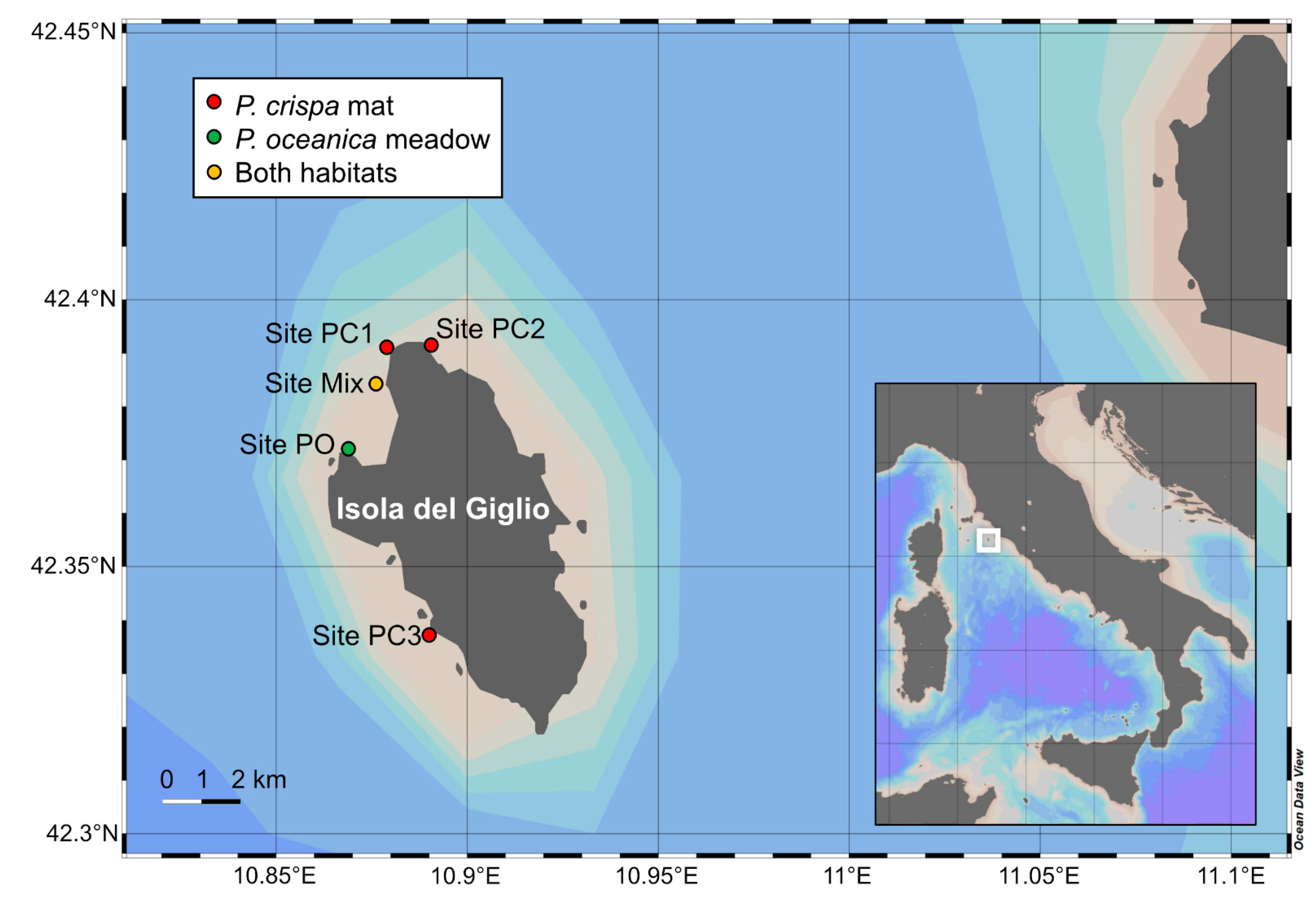
The study was carried out at five sites distributed along the North-eastern and North-western coasts of Giglio Island, in the Tuscan Archipelago National Park (42°21′19.4″ N 10°54′06.1″ E, Tyrrhenian Sea) (Figure 2). The islands’ underwater seabeds are characterized by granite slopes, alternating with sand bottoms, where P. oceanica meadows, P. crispa mats and coralligenous habitats colonize infralittoral seabeds. All samples were collected by SCUBA divers at a water depth of 30 m. Samplings were carried out at three sites with P. crispa mats (Site PC1, PC2 and PC3), one with only P. oceanica meadows (Site PO), and one site (Site mix) with both habitats being present (Figure 2).
Temporally randomized sampling took place during late spring, between May and July 2019. For sampling P. crispa mats, a standardized maximum mat thickness of 5 cm was defined. After randomly defining a sampling spot in the target area, a metal frame (size 30 × 30 cm) was placed in the mat and all algal thalli within, including the holdfasts, were carefully removed using a spatula. Sampled material was then placed into 1 L Kautex jars (each holding approx. 1/3 algae, 2/3 seawater). Every site was sampled four times, resulting in a total of 16 replicate samples for this habitat.
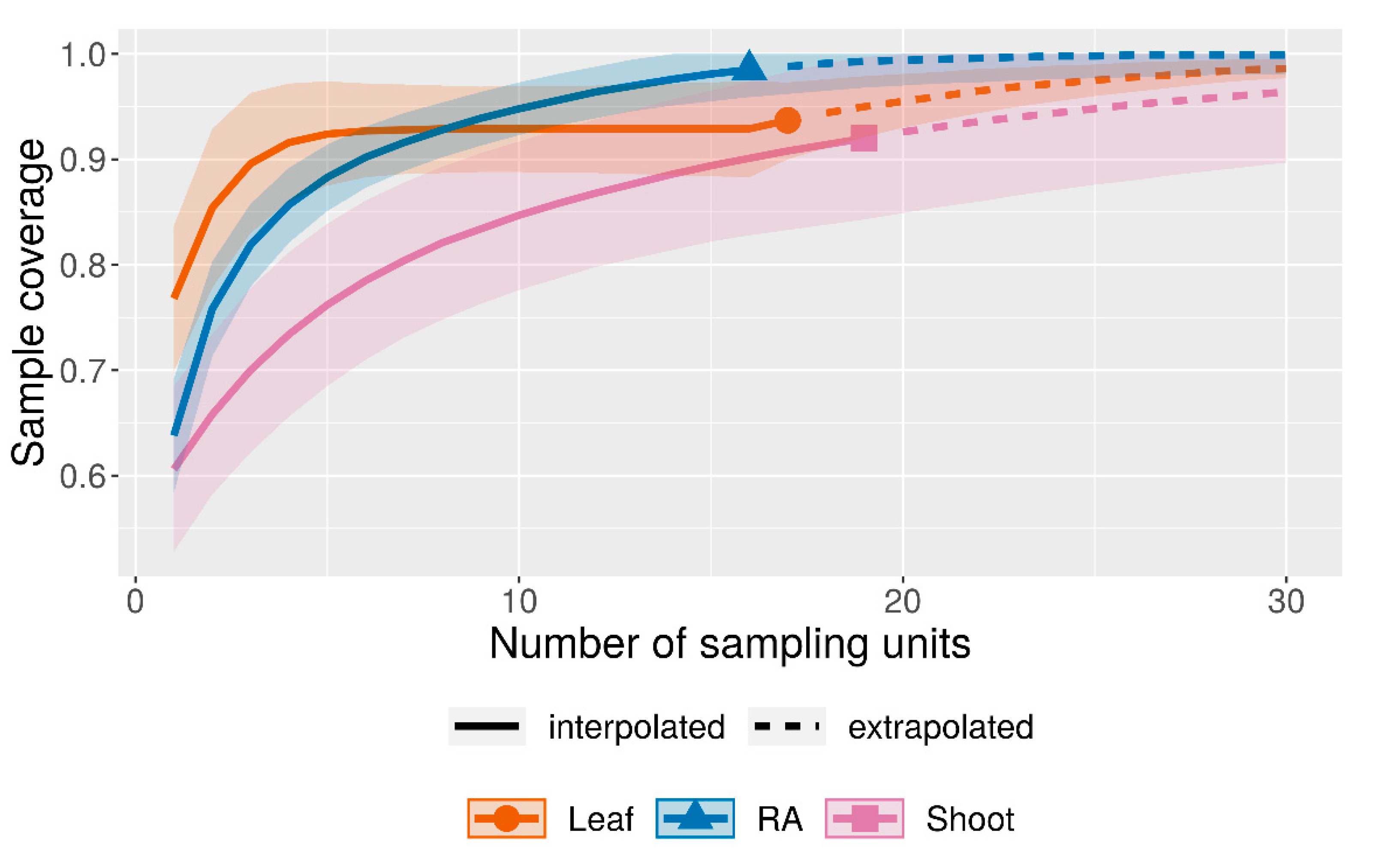
In total, 19 P. oceanica shoots and 17 leaves (site PO: 10 shoots and 9 leaves; site mix: 9 shoots and 8 leaves) were sampled separately into 1 L Kautex jars. The leaves were cut at the sheath of the shoot and shoots were cut at the rhizome node. Samples were immediately transferred into seawater holding tanks at the Institute for Marine Biology (IfMB, Campese, Italy) and kept at a constant temperature of 18 °C. Shoots and leaves were treated as separate subhabitats taking into account their different ecological traits, particularly regarding their longevity as a fundamental trait for serpulid settlement [44,45]. With this approach, we followed recent studies on P. oceanica epifauna [46,47,48]. The number of samples at each of the sites represents the whole epiphytic community, as demonstrated by previous studies on epiphytic communities associated with P. oceanica meadows that used 15 shoots as a significative sampling effort [49,50]. We used this number as a minimum goal for our sampling efforts. Sample completeness was confirmed using the approach of Chao et al. (2016) (Appendix A Figure A1).
In addition, we counted the number of P. oceanica shoots per m² (total n = 74 quadrats; using a plastic tube frame of 40 × 40 cm) and leaves per shoot (total n = 32 shoots) in the sampling area.
2.2. Species Identification
For the analysis of P. crispa associated serpulid polychaetes, a subsample of approximately 10 g wet weight was taken from the main sample. All replicated samples were processed within three days after collection. Algal thalli were transferred into small bowls and cut into single phylloids for analysis under a stereomicroscope (maximum 40× magnification). Following the analysis, the wet weights of the main and subsamples were measured after removing adherent water. The P. oceanica shoots were analyzed as a whole under a stereo microscope, while leaves were cut into pieces of approximately 8 cm for easier handling and to avoid double counting.
All specimens were identified using relevant literature (Appendix A Table A1) and crosschecking with online resources (WORMS; marinespecies.org, accessed on 31 July 2019). The number of individuals per species were also recorded for quantitative analysis. Counting and sample processing were standardized, and every observer was trained to a high level of taxonomic proficiency prior to processing the samples.
The data were then correlated to the surface area, as individuals per m2 surface of P. crispa and P. oceanica. Thalli of P. crispa were placed in a bowl of water on a laminated graph paper sheet and flattened with a glass pane. Pictures were taken from a 90° angle using a Canon G12 camera and a tripod stand to ensure a constant distance and angle to the sample. The surface area was then calculated from the picture with ImageJ (version 1.52o, https://imagej.nih.gov/ij/, accessed on 23 April 2019) and multiplied by two to account for both sides of the thalli. The measured surface area was then extrapolated to the main sample, using the wet weight measured after the analysis. This relation allowed for the extrapolation of the density of serpulids to the surface area of underlying rock (Appendix A Formula (A1)). For P. oceanica shoots, we assumed a cylindrical shape and calculated the surface area using diameter and length. For P. oceanica leaves, we assumed a rectangular shape, calculating its surface area with length and width multiplied by two to account for both sides of the leaf. The total surface area of P. oceanica substrate was then extrapolated using the field observations of leaves per shoot and shoots per m² (Appendix A Formulas (A2) and (A3)), to assess the number of serpulids per m² of seafloor.
2.3. Diversity Descriptors
Diversity was assessed using four descriptors: total numbers of serpulid taxa per site and habitat, total abundances of individuals per m² of substrate, Shannon diversity index [51] and Pielou evenness index [52]. The descriptors were calculated as means per site and then reported with the respective standard error.
2.4. Statistical Analysis
Pairwise Wilcoxon–Mann–Whitney tests were carried out to assess differences in diversity descriptors among sites and habitats (Appendix A, Table A2). Differences in the composition of serpulid assemblages among sites and habitats were tested through multivariate permutational analysis of variance (PERMANOVA [53]). The northern sites’ data were pooled to compare among habitats and further analyze differences among P. crispa sites (Table 1). Pairwise comparisons were conducted using Tukey’s honestly significant difference (HSD) test. Data was not transformed to stress the importance of the abundance of taxa in determining the differences among habitats. The Serpulid assemblages were hierarchically clustered with Spearman ranked correlation (average linkage) using the software ’heatmapper’ [54] to reveal differences in the species composition among sites and visually highlight variances in the serpulid assemblages among habitats. Analyses and plots were made with R (version 3.5.3) [55].
3. Results
3.1. Diversity Descriptors
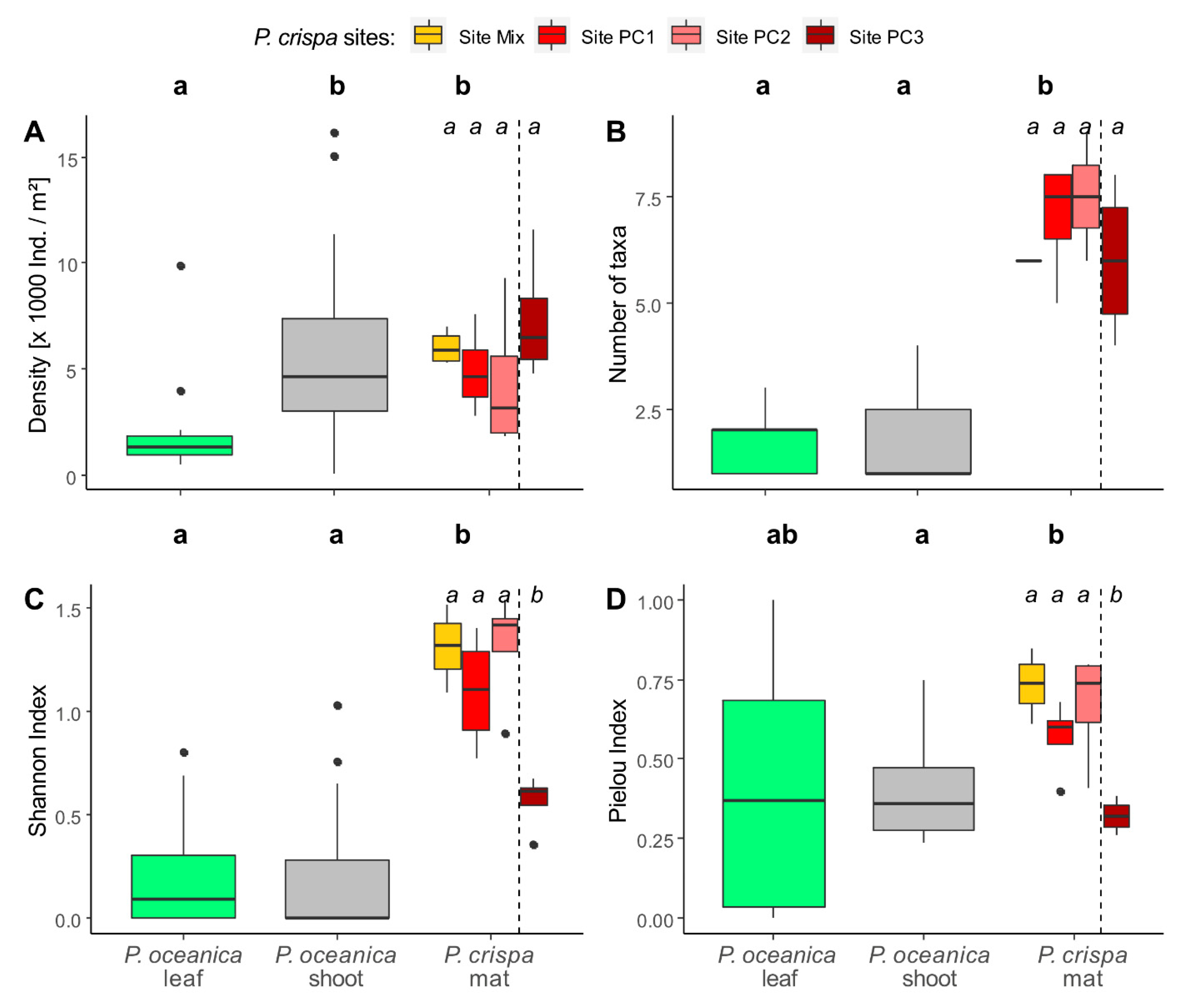
A total of 2403 Serpulidae specimens belonging to 17 taxa were collected. Overall abundances of individuals m−2 and numbers of taxa were significantly higher in P. crispa mats than in P. oceanica shoot and leaf samples (Figure 3).
The density of individuals was highest in P. oceanica shoots at site mix (average 8197 ± 1549 individuals m−2) and lowest in P. oceanica leaves at site PO (average 1714 individuals m−2). The comparison with P. oceanica leaves showed significantly higher densities for P. crispa mats (p < 0.001). The shoots of P. oceanica hosted a similar number of individuals (average 5890 ± 815 individuals m−2) compared to P. crispa samples (average 5664 ± 622 individuals m−2) (Figure 3A). The calculated numbers of serpulids per m2 seafloor showed a similar trend of highest values for P. crispa and P. oceanica shoots (average 1,239,728 ± 784,455 and 864,444 ± 1,081,419, respectively), and a lower density for P. oceanica leaves (average 238,567 ± 179,055). The combined averages of both P. oceanica subhabitats add up to 1,103,011 ± 1,096,142 individuals per m² seafloor.
All of the 17 identified taxa were found in P. crispa samples and only 10 taxa were found in P. oceanica samples. The highest number of P. crispa associated taxa were found in the samples from Site PC2 (average 8 ± 1 taxa) and the lowest in samples from site mix (average 6 ± 0 taxa) and site PC3 (average 6 ± 2 taxa). In comparison, P. oceanica samples harbored 2 ± 1 different taxa on the leaves and shoots, respectively (Figure 3B).
Shannon diversity index was higher in P. crispa mats (average 1.1 ± 0.09) compared to P. oceanica habitats (average leaves: 0.2 ± 0.06; average shoots: 0.2 ± 0.07; Figure 3C). Site PC3 showed a significantly lower diversity compared to other P. crispa samples (p < 0.05).
Pielou evenness index was highest in P. crispa samples (average 0.6 ± 0.05) compared to P. oceanica habitats (average leaves: 0.4 ± 0.09; average shoots: 0.4 ± 0.04; Figure 3D). Site PC3 showed a significantly lower evenness compared to other P. crispa samples (p < 0.05).
3.2. Analysis of Serpulid Assemblages
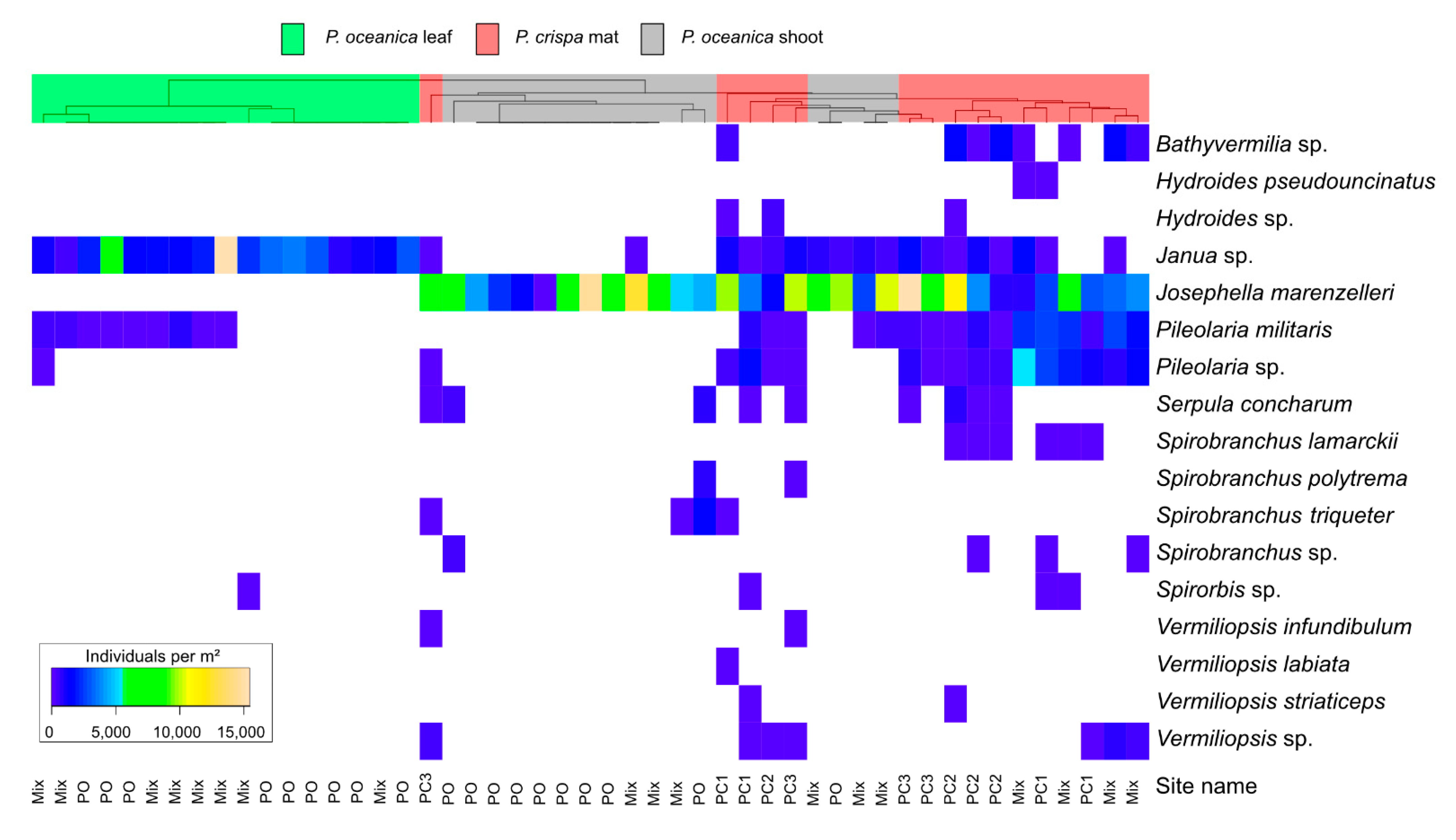
The multivariate analyses (PERMANOVA) showed significant differences among the two P. oceanica habitats without an effect of the site (Table 1). The analysis of P. crispa sites showed a significant difference among sites; however, the pairwise comparison did not confirm this result (Table 2). We further analyzed differences among the habitats for the northern sites only, avoiding potential effects of the location around the island. The cluster analysis confirms these results and shows differences in the species composition of the different habitats (Figure 4). The dendrogram of Spearman rank correlation across sites and habitats shows clustering according to habitat, with a lower coefficient for P. crispa and P. oceanica shoots (r < 0.6) than P. oceanica leaves compared to P. oceanica shoots and P. crispa mat (r > 0.8).
The most frequent species found in all samples of P. crispa mats was Josephella marenzelleri. This species was also largely present on P. oceanica shoots, however, it was not found on P. oceanica leaves. In contrast, the most abundant species on the leaves was Janua sp., which also occurred in 13 P. crispa replicates and on 7 investigated P. oceanica shoots. In addition, J. marenzelleri was also the most abundant species in terms of individuals found m−2 of P. crispa and P. oceanica shoots, while Janua sp. showed the highest density on P. oceanica leaves. Out of the 17 species, two were exclusively found on P. crispa samples: Bathymermilia sp. and Vermiliopsis labiata (Figure 4). The species Pileolaria militaris showed similar abundances in P. crispa and P. oceanica samples.
4. Discussion
This study highlights the high diversity and density of serpulid polychaetes for all investigated habitats. The total number of species found associated with P. crispa mats (17 species) was consistent with previous studies from other areas in the Mediterranean Sea [37,56], including the results of Casoli et al. (2016), which investigated P. crispa mats along a water depth gradient and found 16 species (Table 3) [19]. Here, we focused on the water depth range with the highest P. crispa mat development and identified potential differences among sites. Species richness and abundance were high across all sites; however, the southernmost site (Site PC3) showed a lower diversity and evenness than the northern sites.
The extrapolated abundances per m2 seafloor were comparable between P. crispa (average 1,239,728 ± 784,455) and P. oceanica, (1,103,011 ± 1,096,142) with the shoots mainly contributing to these numbers. However, the calculations likely cause some bias due to the extrapolation based on average values, as indicated by the large standard deviations.
Comparing P. crispa mats and neighboring P. oceanica meadows revealed significant differences in serpulid assemblage composition. By modifying the substratum through the creation of spatial microhabitats and influencing feeding habits, seagrass beds and algae mats appear to favor the establishment of different polychaete assemblages [43,57,58]. Habitat formation and the substantial improvement of food source accessibility are pivotal ecological drivers that influence the epiphytic serpulid assemblages’ diversity and structure. Overall abundance, species richness, diversity and evenness were significantly lower in most P. oceanica samples compared to P. crispa. Exceptions are P. oceanica shoots, where similar densities of individuals were found compared to P. crispa mats. The species composition was also similar, with the same dominant species (J. marenzelleri), which resulted in a shared cluster disparate from the P. oceanica leaf samples (Figure 4). This clustering could be related to the two habitats’ semi-hemisciaphilious conditions, which leads to reduced competition with algal epiphytes and influences rhizome communities [31]. Lower light conditions inside P. crispa mats and P. oceanica shoots are also reflected by the lower numbers of the photophilic Spirorbinae (e.g., Janua sp.) [59], mostly found on the leaves of P. oceanica. The perennial P. crispa thalli provide an extension of colonizable surfaces that are less flexible and long-lived compared to P. oceanica leaves. Constant motion and a one-year life cycle favor species with well-developed strategies to cope with stresses on P. oceanica leaves, such as Spirorbidae (Janua sp.). These are recognized as pioneer species, characterized by fast and cinsistent recruitment on smaller surfaces [60,61]. Thus, dense red algae mats composed of both prostrate and erect thalli (heterotrichous species) [17] constitute a sheltered and long-lived habitat rather than oscillating structures of shallow algae or P. oceanica leaves.
In comparison to previous reports of neighboring (i.e., coralligenous reefs) habitats, we see that J. marenzelleri is a ubiquitous species that is found from the shallow infralittoral to deeper coralligenous habitats (Table 3). This species is known to be a pioneer species with the ability to colonize available settling grounds quickly and efficiently [36]. Furthermore, J. marenzelleri, with its articulate tube, is particularly adapted to colonize rigid algal thalli as epiphytes, being found both on Cystoseira spp. canopy and encrusting red algae [59,62]. Other widely distributed taxa characterized the serpulid assemblage on P. crispa samples. However, the presence of species with dark or deep-habitat affinity (Bathyvermilia sp. and V. infundibulum) confirmed the hemisciaphilic conditions provided within the algal mat.
In contrast, serpulids with a larger and more erect growth form are only found in coralligenous habitats (e.g., Protula sp., Table 3). These slow-growing species are sensitive to water movement and prefer hard settling grounds over flexible algae and plants [59]. However, the overall high abundance of serpulids on P. crispa thalli supports their rigid structure and long-term stability as key characteristics for the establishment of these species. The laminal thalli of P. crispa represent a living substratum suitable for the colonization of sessile solitary species, increasing the surface available for settlement. Sedimentation processes on horizontal and gently sloped sea beds, where P. crispa is abundant, can help understand the patterns described in this study [17]. Serpulidae comprise exclusively filter-feeding species that can benefit from organic and inorganic particle sedimentation as trophic sources.
Communities of the brown algae Cystoseira sp. in the Ionian Sea host a notably higher diversity of serpulids (Shannon of 2.4) than we found in P. crispa mats (average Shannon of 1.1) [59]. This shows dominance of a few species in the P. crispa mats, as also seen in the abundance values. Interestingly, the abundances exhibit a trend of higher numbers on the southern site (PC3) while diversity and evenness are significantly lower (Figure 3). This variation could reflect the serpulid assemblage’s heterogeneity over a larger spatial scale due to the physical exposure of different P. crispa mats. While the diversity was still considerably high in both areas, it also points out that the western Peninsula (Figure 2) separates the southern P. crispa mats from the northern patches. This effect could be driven by the rather exposed situation of PC3 to the prevalent southern currents, while the northern sites are situated on the leeward side of the island [63]. Thus, this situation could also impact larval as well as food particle supply.
We conclude that P. crispa mats harbor a rich serpulid assemblage, comparable to or even exceeding other neighboring biodiversity hotspots (i.e., P. oceanica meadows). Furthermore, we found two species that were unique to this habitat in our study. This high diversity underlines the vital function of P. crispa as a habitat-forming species and host for exceptional biodiversity. Therefore, we recommend evaluating this habitat for future conservation actions to prevent habitat and biodiversity loss. Future research should also address the role of other taxonomic groups associated with the P. crispa mats and assess differences in the composition of infauna communities on a regional scale to confirm its role as an essential benthic habitat in the (western) Mediterranean Sea. Our results revealed potential regional differences in the P. crispa associated fauna that need to be further addressed by sampling other areas along the Mediterranean coast. This information is crucial to explore how connected or isolated the different patches of P. crispa mats are in terms of exchange of biodiversity and larval settlement. Potential drivers of connectivity for serpulid communities are prevalent water currents [33] as indicated by the differences between the northern sites and the rather exposed southern site [64].
Author Contributions
Conceptualization, F.I.R., M.B. and C.W.; methodology, F.I.R. and M.B; validation, F.I.R., M.B. and C.W.; formal analysis, F.I.R. and M.B.; investigation, F.I.R. and M.B.; resources, C.W.; data curation, F.I.R.; writing—original draft preparation, F.I.R. and E.C.; writing—review and editing, F.I.R., E.C., M.B. and C.W.; visualization, F.I.R.; supervision, C.W.; project administration, C.W.; funding acquisition, C.W. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by baseline funding of the Marine Ecology department, University of Bremen and by the Institute for Marine Biology (Campese, Italy). M.B. received funding via the ERASMUS+ program.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The corresponding dataset will be openly available in the PANGAEA data repository (www.pangaea.de) upon publication.
Acknowledgments
The authors would like to thank Jenny Tuček (Institute for marine biology) as well as Reiner and Regina Krumbach (Campese Diving Center) for logistical support throughout our study. We are also thankful to Susann Roßbach for providing helpful feedback on the manuscript, Mischa Schwarzmeier and Anette Reh for their support in sampling activities, and Matthew Tietbohl and Lucy Fitzgerald for language improvement.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Figure A1.
Sample completeness curve based on incidence data according to Chao et al. (2016) [64]. Number of bootstraps used: 500; level of confidence: 0.95. RA = P. crispa mats, Leaf = P. oceanica leaves, Shoot = P. oceanica shoots with sample coverages (C.hat) of 0.98, 0.93 and 0.92, respectively.
Figure A1.
Sample completeness curve based on incidence data according to Chao et al. (2016) [64]. Number of bootstraps used: 500; level of confidence: 0.95. RA = P. crispa mats, Leaf = P. oceanica leaves, Shoot = P. oceanica shoots with sample coverages (C.hat) of 0.98, 0.93 and 0.92, respectively.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table A1.
Literature used for identification of species.
| Author(s) | Year | Title |
|---|---|---|
| Riedl, R. | 2011 | Fauna und Flora des Mittelmeeres |
| Stresemann, E. | 1992 | Wirbellose |
| Hayward, P.J. and Ryland, J.S. | 1999 | Handbook of the Marine Fauna of North-West Europe |
| Bianchi et al. | 1981 | Guide per il riconoscimento delle specie animali delle acque lagunari e costiere italiane |
| Ten Hove et al. | 2009 | Taxonomy of Serpulidae (Annelida, Polychaeta): the state of affairs |
| Zibrowius H. | 1968 | Etude morphologique, systématique et écologique des Serpulidae (Annelida Polychaeta) de la région de Marseille |
| Zibrowius H. | 1972 | Mise au point sur les especes mediterraneennes de Serpulidae (Annelida Polychaeta) déecrites par Stefano delle Chiaje (1822–1829, 1841–1844) et Oronzio Gabriele Costa (1861) |
Table A2.
Results of univariate tests (Wilcoxon–Mann–Whitney) among P. crispa sites (ns = p > 0.05; * = 0.01 < p < 0.05).
Table A2.
Results of univariate tests (Wilcoxon–Mann–Whitney) among P. crispa sites (ns = p > 0.05; * = 0.01 < p < 0.05).
| Group 1 | Group 2 | p-Value | p-Signif | |
|---|---|---|---|---|
| Density | Site Mix | Site PC1 | 0.34 | ns |
| Site Mix | Site PC2 | 0.34 | ns | |
| Site Mix | Site PC3 | 0.69 | ns | |
| Site PC1 | Site PC2 | 0.69 | ns | |
| Site PC1 | Site PC3 | 0.34 | ns | |
| Site PC2 | Site PC3 | 0.20 | ns | |
| No of taxa | Site Mix | Site PC1 | 0.278 | ns |
| Site Mix | Site PC2 | 0.069 | ns | |
| Site Mix | Site PC3 | 1.000 | ns | |
| Site PC1 | Site PC2 | 0.766 | ns | |
| Site PC1 | Site PC3 | 0.454 | ns | |
| Site PC2 | Site PC3 | 0.306 | ns | |
| Shannon index | Site Mix | Site PC1 | 0.486 | ns |
| Site Mix | Site PC2 | 0.686 | ns | |
| Site Mix | Site PC3 | 0.029 | * | |
| Site PC1 | Site PC2 | 0.200 | ns | |
| Site PC1 | Site PC3 | 0.029 | * | |
| Site PC2 | Site PC3 | 0.029 | * | |
| Pielou index | Site Mix | Site PC1 | 0.057 | ns |
| Site Mix | Site PC2 | 0.886 | ns | |
| Site Mix | Site PC3 | 0.029 | * | |
| Site PC1 | Site PC2 | 0.200 | ns | |
| Site PC1 | Site PC3 | 0.029 | * | |
| Site PC2 | Site PC3 | 0.029 | * |
Formula (A1): Calculation of serpulid individuals on P. crispa per m² seafloor (IndSF) from individuals per m² substrate (IndSS), using wet weights (WW) and surface area of the sub sample (SASS) (0.09 m² corresponds to the size of the sampling frame:
Formula (A2): Calculation of serpulid individuals on P. oceanica leaves per m² seafloor using the average leaf surface area (SAleafAVG), average number of leaves per m² (162), surface area of investigated leaf sample (SA leafSS) and individual count per m² substrate (IndSS):
Formula (A3): Calculation of serpulid individuals on P. oceanica shoots per m² seafloor using the shoot surface area (SAshootSS), average number of shoots per m² (40.5) and individual count per m² substrate (IndSS):
References
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Lasram, F.B.R.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The Biodiversity of the Mediterranean Sea: Estimates, Patterns, and Threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [Green Version]
- Medail, F.; Quezel, P. Biodiversity Hotspots in the Mediterranean. Conserv. Biol. 1999, 13, 1510. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Bianchi, C.N.; Morri, C. Marine Biodiversity of the Mediterranean Sea: Situation, Problems and Prospects for Future. Mar. Pollut. Bull. 2016, 40, 367–376. [Google Scholar] [CrossRef]
- Defant, A. Physical Oceanography; Pergamon: New York, NY, USA, 1961; Volume 1. [Google Scholar]
- Boudouresque, C.F.; Bernard, G.; Bonhomme, P.; Charbonnel, E.; Diviacco, G.; Meinesz, A.; Pergent, G.; Pergent-Martini, C.; Ruitton, S.; Tunesi, L. Préservation et Conservation des Herbiers à Posidonia Oceanica; Ramoge and RAC/SPA: Tunis, Tunesia, 2006; ISBN 2905540303. [Google Scholar]
- Mazzella, L.; Buia, M.C.; Gambi, M.C.; Lorenti, M.; Russo, G.F.; Scipione, M.B.; Zupo, V. Plant-animal trophic relationships in the Posidonia oceanica ecosystem of the Mediterranean Sea: A review. Plant Anim. Interact. Mar. Benthos 1992, 46, 165–187. [Google Scholar]
- Ballesteros, E. Mediterranean coralligenous assemblages: A synthesis of present knowledge. In Oceanography and Marine Biology: An Annual Review; Gibson, R.N., Atkinson, R.J.A., Gordon, J.D.M., Eds.; Taylor & Francis: London, UK, 2006; pp. 123–195. [Google Scholar]
- Cocito, S. Bioconstruction and biodiversity: Their mutual influence. Sci. Mar. 2004, 68, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Ingrosso, G.; Abbiati, M.; Badalamenti, F.; Bavestrello, G.; Belmonte, G.; Cannas, R.; Benedetti-Cecchi, L.; Bertolino, M.; Bevilacqua, S.; Bianchi, C.N.; et al. Mediterranean Bioconstructions Along the Italian Coast, 1st ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2018; Volume 79, ISBN 9780128151013. [Google Scholar]
- Lepoint, G.; Balancier, B.; Gobert, S. Seasonal and depth-related biodiversity of leaf epiphytic Cheilostome Bryozoa in a Mediterranean Posidonia oceanica meadow. Cah. Biol. Mar. 2014, 55, 57–67. [Google Scholar]
- Stachowicz, J.J. The Structure of Ecological Communities. Bioscience 2001, 51, 235–246. [Google Scholar] [CrossRef]
- Stachowicz, J.J.; Hay, M.E. Facultative mutualism between an herbivorous crab and a coralline alga: Advantages of eating noxious seaweeds. Oecologia 1996, 105, 377–387. [Google Scholar] [CrossRef]
- Duffy, J.E. Amphipods on seaweeds: Partners or pests? Oecologia 1990, 83, 267–276. [Google Scholar] [CrossRef]
- Tomas, F.; Turon, X. Seasonal and small-scale spatial variability of herbivory pressure on the temperate seagrass Posidonia oceanica. Mar. Ecol. Prog. Ser. 2005, 301, 95–107. [Google Scholar] [CrossRef]
- Piazzi, L.; Balata, D.; Ceccherelli, G. Epiphyte assemblages of the Mediterranean seagrass Posidonia oceanica: An overview. Mar. Ecol. 2016, 37, 3–41. [Google Scholar] [CrossRef]
- Casoli, E.; Bonifazi, A.; Gravina, M.F.; Russo, G.F.; Sandulli, R.; Donnarumma, L. Comparative Analysis of Mollusc Assemblages from Different Hard Bottom Habitats in the Central Tyrrhenian Sea. Diversity 2019, 11, 74. [Google Scholar] [CrossRef] [Green Version]
- Bonifazi, A.; Ventura, D.; Gravina, M.F.; Lasinio, G.J.; Belluscio, A.; Ardizzone, G.D. Unusual algal turfs associated with the rhodophyta Phyllophora crispa: Benthic assemblages along a depth gradient in the Central Mediterranean Sea. Estuar. Coast. Shelf Sci. 2017, 185, 77–93. [Google Scholar] [CrossRef]
- Kostylev, E.F.; Tkachenko, F.P.; Tretiak, I.P. Establishment of “Zernov’s Phyllophora field” marine reserve: Protection and restoration of a unique ecosystem. Ocean Coast. Manag. 2010, 53, 203–208. [Google Scholar] [CrossRef]
- Casoli, E.; Bonifazi, A.; Ardizzone, G.; Gravina, M.F. How algae influence sessile marine organisms: The tube worms case of study. Estuar. Coast. Shelf Sci. 2016, 178, 12–20. [Google Scholar] [CrossRef]
- Navone, A.; Bianchi, C.N.; Orru, P.; Ulzega, A. Saggio di cartografia geomorfologica e bionomica nel parco marino di Tavolara-Capo Coda di Cavallo (Sardegna nord-orientale). Oebalia 1992, XVII, 469–478. [Google Scholar]
- Bianchi, C.N.; Morri, C.; Navone, A. I popolamenti delle scogliere rocciose sommerse dell’Area Marina Protetta di Tavolara Punta Coda Cavallo (Sardegna nord-orientale). Sci. Rep. Port Cros Natl. Park 2010, 24, 39–85. [Google Scholar]
- Donnarumma, L.; Sandulli, R.; Appolloni, L.; Russo, G.F. Assessing molluscs functional diversity within different coastal habitats of Mediterranean marine protected areas. Ecol. Quest. 2018, 29, 35–51. [Google Scholar] [CrossRef] [Green Version]
- Buonocore, E.; Donnarumma, L.; Appolloni, L.; Miccio, A.; Russo, G.F.; Franzese, P.P. Marine natural capital and ecosystem services: An environmental accounting model. Ecol. Modell. 2020, 424, 109029. [Google Scholar] [CrossRef]
- Horton, T.; Kroh, A.; Ahyong, S.; Bailly, N.; Boyko, C.B.; Brandão, S.N.; Gofas, S.; Hooper, J.N.A.; Hernandez, F.; Holovachov, O.; et al. World Register of Marine Species (WoRMS). Available online: https://www.marinespecies.org (accessed on 27 January 2021).
- Giangrande, A.; Gravina, M.F. Brackish-water polychaetes, good descriptors of environmental changes in space and time. Transit. Waters Bull. 2015, 9, 42–55. [Google Scholar]
- Casoli, E.; Ricci, S.; Antonelli, F.; Sacco Perasso, C.; Ardizzone, G.; Gravina, M.F. Colonization dynamic on experimental limestone substrata: The role of encrusting epilithics favouring boring polychaetes. Hydrobiologia 2019, 842, 101–112. [Google Scholar] [CrossRef]
- Musco, L. Ecology and diversity of Mediterranean hard-bottom Syllidae (Annelida): A community-level approach. Mar. Ecol. Prog. Ser. 2012, 461, 107–119. [Google Scholar] [CrossRef] [Green Version]
- Giangrande, A.; Licciano, M.; Musco, L. Polychaetes as environmental indicators revisited. Mar. Pollut. Bull. 2005, 50, 1153–1162. [Google Scholar] [CrossRef]
- Watson, D.I.; Barnes, D.K.A. Quantifying assemblage distinctness with time: An example using temperate epibenthos. J. Exp. Mar. Bio. Ecol. 2004, 312, 367–383. [Google Scholar] [CrossRef]
- Ben-Eliahu, M.N.; Fiege, D. Serpulid tube-worms (Annelida: Polychaeta) of the Central and Eastern Mediterranean with particular attention to the Levant Basin. Senckenberg. Marit. 1996, 28, 1–51. [Google Scholar] [CrossRef]
- Piazzi, L.; Balata, D.; Cinelli, F. Epiphytic macroalgal assemblages of Posidonia oceanica rhizomes in the western Mediterranean. Eur. J. Phycol. 2002, 37, 69–76. [Google Scholar] [CrossRef]
- Fagerstrom, J.A. Reef-building guilds and a checklist for determining guild membership—A new approach for study of communities. Coral Reefs 1991, 10, 47–52. [Google Scholar] [CrossRef]
- Sanfilippo, R.; Vertino, A.; Rosso, A.; Beuck, L.; Freiwald, A.; Taviani, M. Serpula aggregates and their role in deep-sea coral communities in the southern Adriatic Sea. Facies 2013, 59, 663–677. [Google Scholar] [CrossRef]
- Vinn, O.; Ten Hove, H.A.; Mutvei, H.; Kirsimäe, K. Ultrastructure and mineral composition of serpulid tubes (Polychaeta, Annelida). Zool. J. Linn. Soc. 2008, 154, 633–650. [Google Scholar] [CrossRef]
- Ten Hove, H.A.; Kupriyanova, E.K. Taxonomy of Serpulidae (Annelida, Polychaeta): The State of Affairs. Zootaxa 2009, 2036, 1–126. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, C.N. Ecologia dei Serpuloidea (Annelida, Polychaeta) del piano infralitorale presso Portofino (Genova). Boll. Musei Inst. Biol. Univ. Genova 1979, 47, 101–115. [Google Scholar]
- Çinar, M.E. Serpulid species (Polychaeta: Serpulidae) from the Levantine coast of Turkey (eastern Mediterranean), with special emphasis on alien species. Aquat. Invasions 2006, 1, 223–240. [Google Scholar] [CrossRef]
- Balduzzi, A.; Bianchi, C.N.; Burlando, B.; Cattaneo-Vietti, R.; Manconi, R.; Morri, C.; Pansini, M.; Pronzato, R.; Sara, M. Zoobenthos di substrato duro delle isole di Capraia e del Giglio (Arcipelago Toscano). Atti Soc. Toscana Sci. Nat. Resid. 1995, 52, 12. [Google Scholar]
- Zenetos, A.; Çinar, M.E.; Panucci-Papadopoulou, M.A.; Harmelin, J.G.; Furnari, G.; Andaloro, F.; Bellou, N.; Streftaris, N.; Zibrowius, H. Annotated list of marine alien species in the Mediterranean with records of the worst invasive species. Mediterr. Mar. Sci. 2005, 6, 63–118. [Google Scholar] [CrossRef]
- Zibrowius, H. Ongoing modification of the Mediterranean marine fauna and flora by the establishment of exotic species. Mésogée 1991, 51, 83–107. [Google Scholar]
- Hopkins, G.A.; Forrest, B.M. Management options for vessel hull fouling: An overview of risks posed by in-water cleaning. ICES J. Mar. Sci. 2008, 65, 811–815. [Google Scholar] [CrossRef] [Green Version]
- Abbiati, M.; Bianchi, C.N.; Castelli, A. Polychaete Vertical Zonation along a Littoral Cliff in the Western Méditerranean. Mar. Ecol. 1987, 8, 33–48. [Google Scholar] [CrossRef]
- Giangrande, A. Polychaete zonation and its relation to algal distribution down a vertical cliff in the western Mediterranean (Italy): A structural analysis. J. Exp. Mar. Bio. Ecol. 1988, 120, 263–276. [Google Scholar] [CrossRef]
- Kikuchi, T. Handbook of Seagrass Biology: An Ecosystem Perspective; Phillips, R.C., McRoy, C.P., Eds.; Garland STPM Press: New York, NY, USA, 1980. [Google Scholar]
- Kikuchi, T.; Pérès, J.M. Animal communities in seagrass beds: A review. In Seagrass Ecosystems: A Scientific Perspective; McRoy, C.P., Helfferich, C., Eds.; Marcel Dekker: New York, NY, USA, 1967; pp. 147–193. [Google Scholar]
- Mounir, B.B.; Asma, H.; Sana, B.I.; Lotfi, M.; Abderrahmen, B.; Lotfi, A. What factors drive seasonal variation of phytoplankton, protozoans and metazoans on leaves of Posidonia oceanica and in the water column along the coast of the Kerkennah Islands, Tunisia? Mar. Pollut. Bull. 2013, 71, 286–298. [Google Scholar] [CrossRef]
- Donnarumma, L.; Lombardi, C.; Cocito, S.; Gambi, M.C. Settlement pattern of Posidonia oceanica epibionts along a gradient of ocean acidification: An approach with mimics. Mediterr. Mar. Sci. 2014, 15, 498–509. [Google Scholar] [CrossRef] [Green Version]
- Albano, P.G.; Sabelli, B. The molluscan assemblages inhabiting the leaves and rhizomes of a deep water Posidonia oceanica settlement in the central Tyrrhenian Sea. Sci. Mar. 2012, 76, 721–732. [Google Scholar] [CrossRef] [Green Version]
- Piazzi, L.; Balata, D.; Cinelli, F.; Benedetti-Cecchi, L. Patterns of spatial variability in epiphytes of Posidonia oceanica: Differences between a disturbed and two reference locations. Aquat. Bot. 2004, 79, 345–356. [Google Scholar] [CrossRef]
- Mecca, S.; Casoli, E.; Ardizzone, G.; Gambi, M.C. Effects of ocean acidification on phenology and epiphytes of the seagrass Posidonia oceanica at two CO2 vent systems of Ischia (Italy). Mediterr. Mar. Sci. 2020, 21, 70–83. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- McArdle, B.H.; Anderson, M.J. Fitting multivariate models to community data: A comment on distance-based redundancy analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef]
- Free Software Foundation The R Project for Statistical Computing. Available online: http://www.r-project.org (accessed on 10 June 2020).
- Marzialetti, S.; Nicoletti, L.; Ardizzone, G.D. The polychaete community of the Fregene artificial reef (Tyrrhenian Sea, Italy): A 20-year study (1981–2001). Zoosymposia 2009, 2, 551–566. [Google Scholar] [CrossRef]
- Giangrande, A. Trophic structure changes of a polychaete community along a vertical cliff. In Rapport Commission Internationale pour L’exploration Scientifique de la Mer Méditerranée; CIESM: Villa Girasole, Monaco, 1986; Volume 30, p. 252. [Google Scholar]
- Mikac, B.; Licciano, M.; Jaklin, A.; Iveša, L.; Giangrande, A.; Musco, L. Diversity and distribution patterns of hard bottom polychaete assemblages in the north adriatic sea (Mediterranean). Diversity 2020, 12, 408. [Google Scholar] [CrossRef]
- Sanfilippo, R.; Rosso, A.; Sciuto, F.; Serio, D.; Catra, M.; Alongi, G.; Negri, M.P.; Leonardi, R.; Viola, A. Serpulid polychaetes from Cystoseira communities in the Ionian Sea, Mediterranean. Vie Milieu 2017, 67, 217–226. [Google Scholar]
- Mabrouk, L.; Ben Brahim, M.; Hamza, A.; Bradai, M.N. Temporal and spatial zonation of macroepiphytes on Posidonia oceanica (L.) Delile leaves in a meadow off Tunisia. Mar. Ecol. 2015, 36, 77–92. [Google Scholar] [CrossRef]
- Ippolitov, A.P.; Rzhavsky, A.V. Tube morphology, ultrastructures and mineralogy in recent Spirorbinae (Annelida: Polychaeta: Serpulidae). III. Tribe Circeini. Invertebr. Zool. 2015, 12, 151–173. [Google Scholar] [CrossRef]
- Sanfilippo, R. Micromorphology, microstructure and functional morphology of the Josephella marenzelleri (Polychaeta Serpulidae) tube. In Autoecology of Selected Organisms: Achievements and Problems; Bollettino della Società Paleontologica Italiana: Milan, Italy, 1996; Volume 3, pp. 205–211. [Google Scholar]
- Boero, F.; De Leo, F.; Fraschetti, S.; Ingrosso, G. The Cells of Ecosystem Functioning: Towards a Holistic Vision of Marine Space, 1st ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2019; Volume 82. [Google Scholar]
- Chao, A.; Ma, K.H.; Hsieh, T.C. iNEXT (iNterpolation and EXTrapolation) Online: Software for Interpolation and Extrapolation of Species Diversity. Program and User’s Guide. 2016. Available online: http://chao.stat.nthu.edu.tw/wordpress/software_download/ (accessed on 28 May 2021).
Figure 1.
Top: P. crispa mat in full view (A) and close up including sessile polychaetes and other epifauna (B). Bottom: P. oceanica mat as seen from the water column (C) and close-ups of the leaves (D) and shoots (E), showing different sessile epifauna. Pictures: F. I. Rossbach.
Figure 1.
Top: P. crispa mat in full view (A) and close up including sessile polychaetes and other epifauna (B). Bottom: P. oceanica mat as seen from the water column (C) and close-ups of the leaves (D) and shoots (E), showing different sessile epifauna. Pictures: F. I. Rossbach.

Figure 2.
Location of Giglio Island in the northern part of the Tyrrhenian Sea and sampling sites, for P. crispa mats (red), P. oceanica meadows (green) and sites where both habitats were sampled (yellow).
Figure 2.
Location of Giglio Island in the northern part of the Tyrrhenian Sea and sampling sites, for P. crispa mats (red), P. oceanica meadows (green) and sites where both habitats were sampled (yellow).

Figure 3.
(A) Total numbers of identified species per site, (B) mean densities of serpulids per m2 substrate per site, (C) Shannon diversity index and (D) Pielou evenness index reported for the investigated P. crispa (red) sites in comparison to P. oceanica leaves (green) and shoots (grey). Letters a and b indicate the results of pairwise Wilcoxon–Mann–Whitney tests performed for comparison among habitats in northern sites (excluding PC3; bold letters) and among all P. crispa sites. Black dots resemble outliers.
Figure 3.
(A) Total numbers of identified species per site, (B) mean densities of serpulids per m2 substrate per site, (C) Shannon diversity index and (D) Pielou evenness index reported for the investigated P. crispa (red) sites in comparison to P. oceanica leaves (green) and shoots (grey). Letters a and b indicate the results of pairwise Wilcoxon–Mann–Whitney tests performed for comparison among habitats in northern sites (excluding PC3; bold letters) and among all P. crispa sites. Black dots resemble outliers.

Figure 4.
Hierarchical clustering (average linkage, Spearman ranked correlation) of taxa abundances among sites (individuals per m²) found in P. crispa (red) and P. oceanica habitat northern sites (green = leaf; grey = shoot).
Figure 4.
Hierarchical clustering (average linkage, Spearman ranked correlation) of taxa abundances among sites (individuals per m²) found in P. crispa (red) and P. oceanica habitat northern sites (green = leaf; grey = shoot).

Table 1.
Results of permutational multivariate analysis of variance (PERMANOVA) of Serpulid assemblages among P. oceanica subhabitats (shoots and leaves), and pairwise comparison of all habitats on the northern sites (site PC1, PC2, PO and mix). Significant results are indicated in bold.
Table 1.
Results of permutational multivariate analysis of variance (PERMANOVA) of Serpulid assemblages among P. oceanica subhabitats (shoots and leaves), and pairwise comparison of all habitats on the northern sites (site PC1, PC2, PO and mix). Significant results are indicated in bold.
| Serpulid Assemblages on P. oceanica Subhabitats | |||||
|---|---|---|---|---|---|
| Source | Df | SS | R2 | F | p |
| Habitat | 1 | 6.1885 | 0.5350 | 50.946 | 0.001 |
| Site | 1 | 0.2829 | 0.0267 | 2.329 | 0.094 |
| Habitat:Site | 1 | 0.2474 | 0.2333 | 2.037 | 0.117 |
| Residuals | 32 | 3.8871 | 0.3665 | ||
| Total | 35 | 10.6059 | 1.0000 | ||
| Pairwise Comparison (All Habitats, N Sites) | |||||
| Pairs | p | p adj | |||
| P. crispa mat | P. oceanica leaf | 0.001 | 0.003 | ||
| P. crispa mat | P. oceanica shoot | 0.001 | 0.003 | ||
| P. oceanica leaf | P. oceanica shoot | 0.001 | 0.003 | ||
Table 2.
Results of permutational multivariate analysis of variance (PERMANOVA) and pairwise comparison of Serpulid assemblages among P. crispa sites. Significant results are indicated in bold.
Table 2.
Results of permutational multivariate analysis of variance (PERMANOVA) and pairwise comparison of Serpulid assemblages among P. crispa sites. Significant results are indicated in bold.
| Serpulid Assemblages on P. crispa Mats | |||||
|---|---|---|---|---|---|
| Source | Df | SS | R2 | F | p |
| Site | 3 | 0.9182 | 0.4066 | 2.7407 | 0.012 |
| Residual | 12 | 1.3401 | 0.5934 | ||
| Total | 15 | 2.2583 | 1.0000 | ||
| Pairwise Comparison (P. crispa Sites) | |||||
| Pairs | p | p adj. | |||
| Mix | PC1 | 0.444 | 1.000 | ||
| Mix | PC2 | 0.032 | 0.192 | ||
| Mix | PC3 | 0.033 | 0.198 | ||
| PC1 | PC2 | 0.351 | 1.000 | ||
| PC1 | PC3 | 0.104 | 0.624 | ||
| PC2 | PC3 | 0.062 | 0.372 | ||
Table 3.
Serpulidae species found in the samples compared to other habitats, x indicates presence; Data from (1) Casoli et al. (2016) [19] and (2) this study (grey columns).
Table 3.
Serpulidae species found in the samples compared to other habitats, x indicates presence; Data from (1) Casoli et al. (2016) [19] and (2) this study (grey columns).
| Species | Shallow Infralittoral (1) | P. crispa Mat (1),(2) | Coralligènous Habitat (2) | P. oceanica Meadow (2) | |
|---|---|---|---|---|---|
| Bathyvermilia sp. | x | ||||
| Filograna implexa (Berkeley) | x | x | |||
| Hydroides spp. (Gunnerus) | x | x | x | x | x |
| Janita fimbriata (Delle Chiaje) | x | ||||
| Janua sp. | x | x | x | x | |
| Josephella marenzelleri (Caullery and Mesnil) | x | x | x | x | x |
| Metavermilia multicristata (Philippi) | x | ||||
| Pileolaria spp. | x | x | x | x | x |
| Protula sp. (Risso) | x | ||||
| Semivermilia crenata (O. G. Costa) | x | x | |||
| Semivermilia cribrata (O. G. Costa) | x | ||||
| Serpula spp. | x | x | x | x | x |
| Serpula vermicularis (Linnaeus) | x | x | |||
| Spiraserpula massiliensis (Zibrowius) | x | ||||
| Spirobranchus spp. | x | x | x | x | x |
| Spirorbis spp. | x | x | x | ||
| Vermiliopsis spp. | x | x | x | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rossbach, F.I.; Casoli, E.; Beck, M.; Wild, C. Mediterranean Red Macro Algae Mats as Habitat for High Abundances of Serpulid Polychaetes. Diversity 2021, 13, 265. https://0-doi-org.brum.beds.ac.uk/10.3390/d13060265
AMA Style
Rossbach FI, Casoli E, Beck M, Wild C. Mediterranean Red Macro Algae Mats as Habitat for High Abundances of Serpulid Polychaetes. Diversity. 2021; 13(6):265. https://0-doi-org.brum.beds.ac.uk/10.3390/d13060265
Chicago/Turabian StyleRossbach, Felix I., Edoardo Casoli, Milan Beck, and Christian Wild. 2021. "Mediterranean Red Macro Algae Mats as Habitat for High Abundances of Serpulid Polychaetes" Diversity 13, no. 6: 265. https://0-doi-org.brum.beds.ac.uk/10.3390/d13060265
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.