
Quantifying Southern Pacific Rattlesnake (Crotalus oreganus helleri) Hunting Behavior through Community Science


Department of Biology, California State University San Bernardino, 5500 University Parkway, San Bernardino, CA 92407, USA
*
Author to whom correspondence should be addressed.
Diversity 2021, 13(8), 349; https://0-doi-org.brum.beds.ac.uk/10.3390/d13080349
Submission received: 24 June 2021
/
Revised: 17 July 2021
/
Accepted: 28 July 2021
/
Published: 29 July 2021
(This article belongs to the Special Issue Advances in the Conservation and Ecology of Rattlesnakes)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:It is increasingly important to study animal behaviors as these are the first responses organisms mount against environmental changes. Rattlesnakes, in particular, are threatened by habitat loss and human activity, and require costly tracking by researchers to quantify the behaviors of wild individuals. Here, we show how photo-vouchered observations submitted by community members can be used to study cryptic predators like rattlesnakes. We utilized two platforms, iNaturalist and HerpMapper, to study the hunting behaviors of wild Southern Pacific Rattlesnakes. From 220 observation photos, we quantified the direction of the hunting coil (i.e., “handedness”), microhabitat use, timing of observations, and age of the snake. With these data, we looked at whether snakes exhibited an ontogenetic shift in behaviors. We found no age differences in coil direction. However, there was a difference in the microhabitats used by juveniles and adults while hunting. We also found that juveniles were most commonly observed during the spring, while adults were more consistently observed throughout the year. Overall, our study shows the potential of using community science to study the behaviors of cryptic predators.
1. Introduction
Predators play a vital role in ecosystems, exhibiting top-down effects within their respective food chains [1]. They regulate prey abundance and competition [2], and have been shown to affect prey foraging behavior and microhabitat use [3]. Because a predator’s reproduction and survival are linked to acquiring prey, many predators have evolved diverse adaptations to capture prey, including morphological, physiological, and behavioral traits [4,5]. Thus, the ways in which predators hunt (i.e., their behavior) can affect their relative fitness. Yet, wild predators are often logistically difficult to track and observe and therefore most studies on wild predator behavior have relatively large research teams and large research costs, and yet comparatively small sample sizes. Due to this, we often lack information on key traits such as how predators hunt and whether hunting behaviors change ontogenetically. With an increase of data and the use of community science, the general public could generate information on how predatory organisms impact ecosystems [6], and can also lead to interest in predator-based conservation [7].
Rattlesnakes are top predators within the Americas, but habitat loss (e.g., due to urbanization) and human activities (e.g., rattlesnake roundups) are threatening many species. For example, Red Diamond Rattlesnakes (Crotalus ruber), Timber Rattlesnakes (C. horridus), Twin-spotted Rattlesnakes (C. pricei), Banded Rock Rattlesnakes (C. lepidus klauberi), and Ridgenosed Rattlesnakes (C. willardi) have lost much of their suitable habitats due to human activity and land development [8,9,10,11,12]. Eastern Massasaugas (Sistrurus catenatus) are considered endangered in all of their range [13], and Eastern Diamondbacks (C. adamanteus) are on the verge of becoming listed as a threatened species [14,15]. Understanding rattlesnake behaviors could be key for better understanding the ecologies of these species. Yet, very few studies focus on wild juvenile rattlesnake behavior, so conservation efforts may gear towards adult tendencies, which could be ineffective. Understanding ontogeny of behavior is often important. For example, Hellbenders (Cryptobranchus alleganiensis) show ontogeny in diet, suggesting the importance of aquatic insect microhabitats to protect juvenile Hellbenders [16]. American Alligators (Alligator mississippiensis) have been shown to utilize different microhabitats throughout their lifetime, therefore, all these microhabitats should be recognized in conservation plans [17]. Studies on juvenile individuals could reveal different needs of rattlesnakes throughout their life stages.
Rattlesnakes are ambush hunters that remain coiled at a single hunting site and wait hours to days to surprise-attack unsuspecting prey [18,19]. Rattlesnakes heavily rely on crypsis while hunting in order to remain undetected from predators and prey [20]. Many rattlesnake species exhibit an ontogenetic shift in diet with a shift from lizard to rodent prey as juveniles transition to adults [21,22,23]. Yet, even though these dietary shifts are well-known, we lack detailed information on if and how hunting behaviors of free-ranging rattlesnakes change ontogenetically. Tracking juveniles can be difficult due to their small size, which is why most dietary studies utilize dead specimens [21,24]. Due to the low quantity of juvenile observations within studies of free-ranging individuals, very little is known about their behavior. This makes it difficult for scientists to fully grasp the concept of ontogeny of many rattlesnake behaviors. It has been shown that adult sidewinder snakes (C. cerastes) utilize better hunting sites and exhibit relatively different activity cycles when compared to juveniles, suggesting an ontogenetic shift in hunting behaviors [25]. Juvenile Santa Catalina Rattlesnakes (C. catalinensis) can often be found in arroyos where lizard activity is high, while adult rattlesnakes are more often found in areas with high mammal activity [26]. Other studies have hypothesized similar behaviors based on fecal samples of juvenile specimens [21,24]. Aside from few studies, little is known about rattlesnake hunting behavior, likely because of the time, money, and other logistical difficulties in studying completely free-ranging individuals, especially small-bodied juveniles which are usually too small for radio transmitters (i.e., to track individuals in the wild).
In traditional field studies, time and money often create logistical obstacles that influence the collection of data and limit our ability to effectively learn about wild animal behaviors. As with most predators, rattlesnakes are notoriously difficult to track because of their cryptic habits, and this can lead to relatively low sample sizes. Specific equipment and tracking methods are usually needed to efficiently study individuals within a population. When using traditional field methods, rattlesnakes are commonly captured and given a radio transmitter or GPS tracker via surgical implantation or external attachment [25,27,28,29,30]. The equipment needed for surgery and tracking can be expensive and require a large crew to manage. Tracking can be a time-consuming process, and an “on-the-ground” research team is often needed in order to maximize sampling rates. Some studies take place in remote areas that are difficult to perform frequent revisits and can lead to studies taking longer amounts of time than anticipated [31]. Data collection can take years and can still result in low sample sizes because of the above-mentioned constraints. For example, previous studies have spent three or more years and obtained sample sizes of fewer than 50 individuals [28,32,33]. Some studies with sample sizes of more than 50 individuals have taken 10 years or more to complete [34,35,36]. With rapid habitat loss or modifications affecting many species, we need a more effective way of collecting basic ecological and behavioral data at broad spatial and temporal scales.
Community science (also called citizen science) is a promising new approach to study the behaviors of hard-to-study species, while reducing the above-mentioned barriers. Community science is when the general public assists in a scientific investigation utilizing the scientific method as defined by the Cornell Lab of Ornithology (www.birds.cornell.edu accessed on 20 June 2021). Community science collaborations have increased over the past years [37,38,39] and could provide new avenues for data collection beyond costly traditional field techniques. Through community science, we can access large crowd-sourced data that are taken over broad environmental scales. Thus far, community science has been useful for conservation-based research. For instance, it has led to the rediscovery of rare species [40] or the documentation of new populations of nonnative/invasive species [41]. Researchers have used it to track urbanization’s effects on bird diversity [42]. This study showed that green areas are critical to bird biodiversity within urban areas, information that could be directly used by conservation managers. Community science also gives researchers the ability to collect data from private properties, for instance, by tracking bumblebee nesting abundance within residential gardens [43]. A recent study demonstrated a novel method to use photo-vouchered observations by community scientists to quantify the effects of urbanization on tail loss (a proxy for predation risk) and parasitism in Southern Alligator Lizards (Elgaria multicarinata) [44]. Like rattlesnakes, Alligator Lizards are a reclusive species that rely on crypsis for survival, and because of this, very little is known about their ecology, yet the researchers were able to download and evaluate more than 700 observations of individual alligator lizards in less than a month. This study provided initial support for the promising use of community science to crowdsource data on secretive, cryptic, or rare species for which the ecology and behaviors are little known or hard to study. Community science may become an ideal approach to overcome barriers that often coincide with predator-focused research.
Here, we assess the potential for using community science to study rattlesnake hunting behaviors. Rattlesnakes are a good species for this work as they are generally tolerant of human observers (i.e., it is often easy for people to approach and take relatively good photographs of them), and their behaviors are relatively easy to quantify based on body posture. When hunting, rattlesnakes coil very tightly and exhibit a distinct kink within the neck region [25,45]. Furthermore, the age class (juvenile vs. adult) of some rattlesnake species is distinguishable by color patterns, and across all species, relative age can be determined through the shape of the rattle and number of rattle segments (detailed in the Methods below). Because it is relatively easy to visually quantify the behaviors and age class of rattlesnakes, they make a good study system in which to use community science photographs to understand the nature and ontogeny of their hunting behaviors. We took advantage of the large sample sizes obtained through photo-vouchered community science observations to address two key questions: (1) whether snakes exhibit behavioral lateralization when hunting, and (2) whether hunting behaviors change ontogenetically.
Behavioral lateralization, also known as “handedness”, refers to preferentiation of a specific side of the body (left or right) when it comes to behaviors that have a directional component, such as locomotion or appendage use [46]. Behavioral lateralization is important because it can affect multiple facets of an organism’s life such as mating, feeding, and hunting [47]. Various snake species are speculated to exhibit behavioral lateralization in mating behaviors (which hemipenis males use during copulation), and while hunting [48,49]. Cottonmouths (Agkistrodon piscivorus) display handedness in hunting coil direction (e.g., clockwise or anticlockwise) at the population level, and age is speculated to be a contributing factor in handedness [49]. Further research is needed to determine why snakes display handedness, for instance, whether it is related to the microhabitat of the hunting site and therefore associated with hunting success within specific microhabitats. Some studies suggest that it may affect hunting success, such as in snail eating snakes, but future research is needed to confirm this claim [50].
The present study utilized community science as a way to quantify Southern Pacific Rattlesnake (Crotalus oreganus helleri) hunting behavior. We focused on this species (C. oreganus) as it is one of the most commonly observed rattlesnake species on the community science platform iNaturalist (www.inaturalist.org accessed on 20 June 2021—one of two community science platforms that we used) with over 5000 observations (as of 22 April 2021). We narrowed results to the subspecies C. o. helleri to reduce variations in hunting behaviors that could be associated with geographic range and because the authors are most familiar with the biology of this species. These snakes are found in southwestern California as far east as San Bernardino County, and can also be found in Baja California. They can be found in various habitats including seaside dunes, desert scrub, and open woodlands [51]. Using photo-vouchered community science data, we evaluated if hunting rattlesnakes exhibited behavioral lateralization (handedness) in ambush coil direction, and if an ontogenetic shift exists in hunting behaviors (coil direction, timing of observations, and microhabitat use).
2. Materials and Methods
2.1. Identification of Community Science Data
We searched for photo-vouchered observations of Southern Pacific Rattlesnakes on two community science platforms: iNaturalist (www.inaturalist.org accessed on 30 March 2020) and HerpMapper (www.herpmapper.org accessed on 30 March 2020). We recognize that many community science platforms exist, and we chose these two for specific reasons. First, iNaturalist is taxonomically diverse and is used by people around the world (notably, it is the platform of choice for the global City Nature Challenge event—www.citynaturechallenge.org accessed on 30 March 2020), making it the platform with the most data observations that we are aware of. Furthermore, data undergo quality assessments and are readily available for downloading. HerpMapper is one of the largest platforms for observations of reptiles and amphibians. Like iNaturalist, HerpMapper is used by people around the world, data are photo-vouchered to verify species identity, and data can be easily filtered and downloaded from the website. Although our study is focused on a single species, we felt that both these platforms were useful to include because they are global in extent, taxonomically diverse (for iNaturalist), and data are accessible for research, allowing the methods presented in this study to be adopted by scientists that work in different locations and on different taxa.
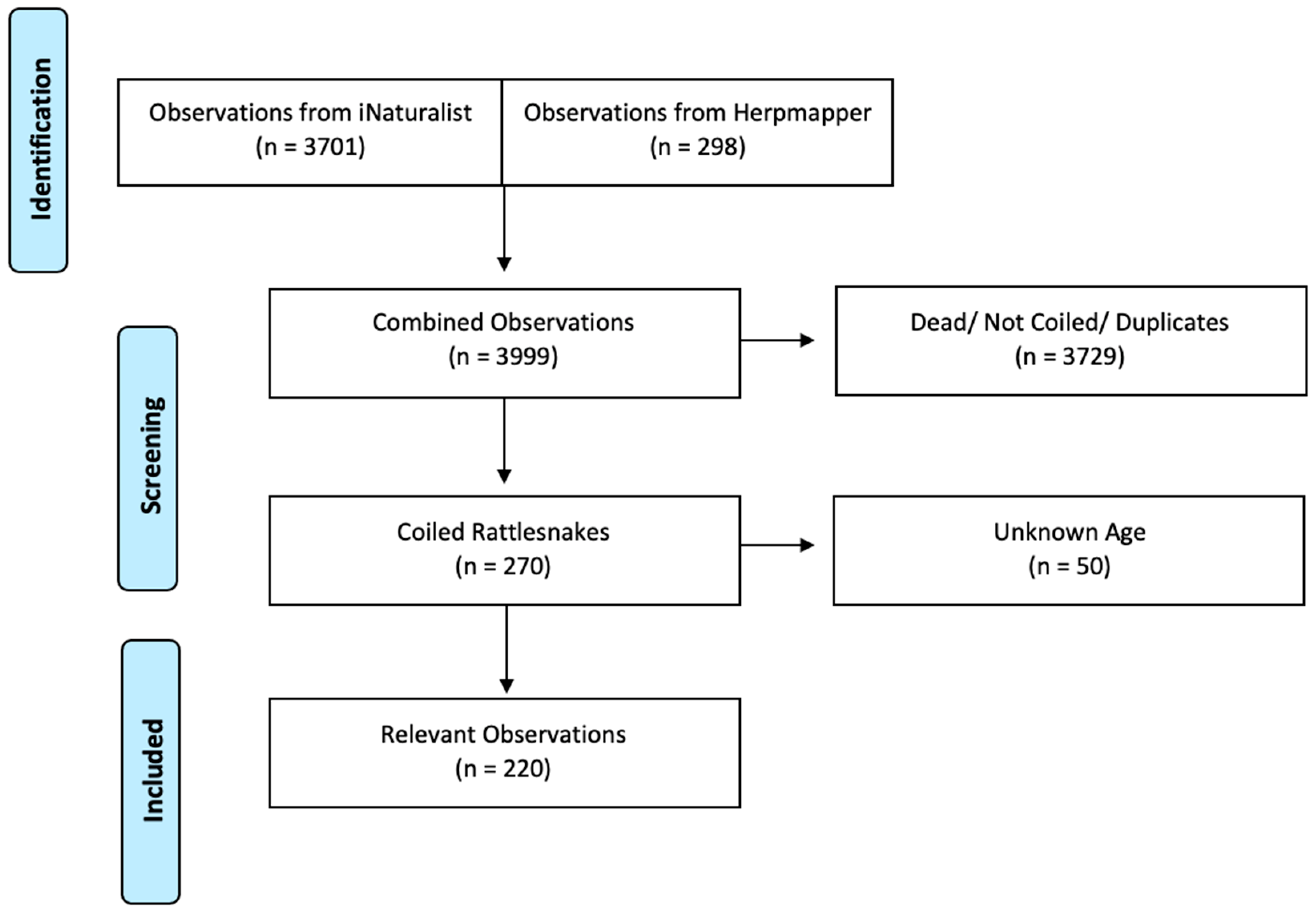
To identify appropriate observations for our study, we filtered the data using dates from the initiations of the platforms to 30 March 2020 (Year of creation: HerpMapper 2013; iNaturalist 2008). Only photo-vouchered observations with confirmed taxonomic identification were used. Observations on iNaturalist become ResearchGrade when they have media (photo or sound), a location, a date, and a community consensus on a precise taxonomic identification. More than two-thirds of the community identifiers must agree on the taxonomic ID. HerpMapper uses a similar method, as users can choose the taxonomic ID, but it can be corrected or changed by a panel of taxonomic reviewers who are experienced herpetologists. In HerpMapper, we also were able to filter out observations of deceased individuals. Combined, our iNaturalist and HerpMapper searches yielded 3999 Southern Pacific Rattlesnake observations. We visually inspected the photographs of each observation to select those with snakes in an ambush hunting posture (tight coil with kink in neck). After filtering out observations of snakes that were dead, not in a hunting posture, duplicates, or in which age could not be identified (see below), we ended up with 220 Southern Pacific Rattlesnake hunting observations (Figure 1).
2.2. Data Extraction
From observations, we extracted various data, including age class of the snake (adult or juvenile), coil direction (clockwise or anticlockwise), microhabitat of the ambush site, whether the snake was hunting on the ground or on an elevated perch, location (latitude and longitude), and date and time of the observation.
Ages of Southern Pacific Rattlesnakes were classified as either adult or juvenile and identified using three methods: (1) the life stage was indicated by the observer, (2) body pattern and color variation (we classified juveniles as having a more defined and contrasting pattern and a yellow tail), and/or (3) shape of rattle and number of rattle segments (we classified juveniles as having a tapered rattle with an intact neonatal button and no more than 5 segments). Due to the fact that it is hard to obtain sex without observing the actual specimen, sex was classified as unknown for all data. Past studies showed that sex has little to no effect on rattlesnake hunting behavior [52,53].
For our study, only tightly coiled body postures with a kinked neck were considered as hunting, while any snake elongated or loosely coiled was ignored [25]. If coiled, the snakes were categorized further into clockwise or anticlockwise formation, in terms of coil direction of the body. A hunting snake will be tightly coiled, with a kink in its neck, followed by its body going either to the left (anticlockwise) or the right (clockwise). A preference in coil direction can be referred to as lateralization or handedness.
The microhabitat of each snake’s hunting site was identified by using the following categories: dirt, brush, rock, wood, and human-made substrate. Dirt was classified as anything that had a particle-like consistency; mud and sand were included in the dirt category. Brush included plants, foliage, or plant debris such as fallen leaves. Rock included boulders and ground cover that had a gravel-like consistency. Logs, tree trunks, and areas with high amounts of fallen bark were classified as wood microhabitats. Human-made microhabitats contained anything that was constructed of human-made material (concrete, asphalt, etc.). If a photograph captured several microhabitats, the observation was categorized using the microhabitat that the snake utilized the highest percentage of (in terms of contact with its body). If we could not access the surrounding microhabitat, we defined it as Not Applicable, only 2 observations of the total 220 fell in this category. We also determined whether snakes were hunting above ground level (i.e., on a perch) or not (i.e., on the ground). Photos were only considered for this category if we could assess the habitat within a coil’s length above, below, and to the sides of the snakes. If the snake’s whole body was on a surface that was elevated on top of natural substrate (logs, concrete, large rocks, etc.), it was considered off the ground. Following our data extraction methods, only 78 out of the 220 observations followed the criteria of this category.
We used the date of each observation to assess seasonal differences in observations. We based our seasons on the biological behaviors of rattlesnakes. Winter included November–February, months which have traditionally been characterized as an inactive period for rattlesnakes [10]. Spring included March–May, which was considered the mating season (based on hormonal studies and observations of mating behavior) [54,55]. Summer included June–July which is considered the foraging season [52,56]. Fall included August–October which is a second mating period, and is when female snakes give birth [51]. We used the date of observation to collect sunrise times and from these, calculated the time of each observation relative to hours after sunrise. We did this as snakes may use the position of the sun and associated ambient temperatures as their hunting cues [25].
2.3. Statistical Analyses
All analyses were done in R (v. 4.0.3, The R Foundation for Statistical Computing) and alpha was set to 0.05. We analyzed data on 220 community science observations of Southern Pacific Rattlesnakes, 129 juveniles and 91 adults (Figure 1). We first looked at whether snakes were found coiled in a direction that deviated from an expected 50% probability (i.e., if they had a “preference”) by using Chi-squared goodness of fit tests, and we tested adults and juveniles separately. Then, we used a logistic regression to look at the effects of age, microhabitat, and their interaction on the coil direction of ambushing snakes. We used Chi-squared tests of independence to look for the effect of age on microhabitat use, season observed, and use of elevated ambush sites (on ground vs. off ground). We used a Mann-Whitney U test to look at whether the hour of day for observations (time since sunrise) differed between juvenile and adult snakes. We only included daytime observations for this analysis as we assume most community scientists are not active at night and so we subsequently had very few nighttime observations (only 15 out of 220). We chose a non-parametric test because we were unable to meet the assumptions of parametric tests.
We recognize that community science observations can be biased as observers are usually not following standardized research protocols. First, spatial and temporal biases within the data will occur because community scientists may be more likely to sample certain locations or during certain times of day or year. These temporal and spatial biases should be standardized across both age classes of rattlesnakes and therefore should not introduce significant biases for the questions we are attempting to address. Other biases could include multiple observations of the same individual snake or by the same user (i.e., one or a few community scientists contributing the majority of the data). We assessed these biases in a few ways. First, we calculated the number of observations that occurred within 500 m of each other. We chose this distance as a home range and movement study on C. oreganus showed that the maximum home range size of 17 tracked individuals was approximately 10 ha in area, which converts to a diameter of roughly 357 m [57]. We chose 500 m as a conservation estimate. All pairwise observations that occurred within 500 m of each other were visually examined to determine whether they might be of the same individual. We looked at age class and dorsal pattern, which can be used to identify individuals in rattlesnakes [57]. Only 20 of the 220 observations occurred within 500 m of each other. None appeared to be of the same individual. Next, we calculated the mean number of observations contributed by each community scientist to determine whether observations might be biased toward certain individual observers. Observations in our dataset came from 131 unique community scientists. We found that the mean ± SD of observations per observer was 1.68 ± 2.71 suggesting that most community scientists contributed only 1–2 observations to the dataset (Supplementary Materials Figure S1). We had four users who submitted 10 or more observations of rattlesnakes with the top observer submitting 21 observations. We redid all analyses above with data from these “super observers” removed and found the same qualitative results presented below.
3. Results
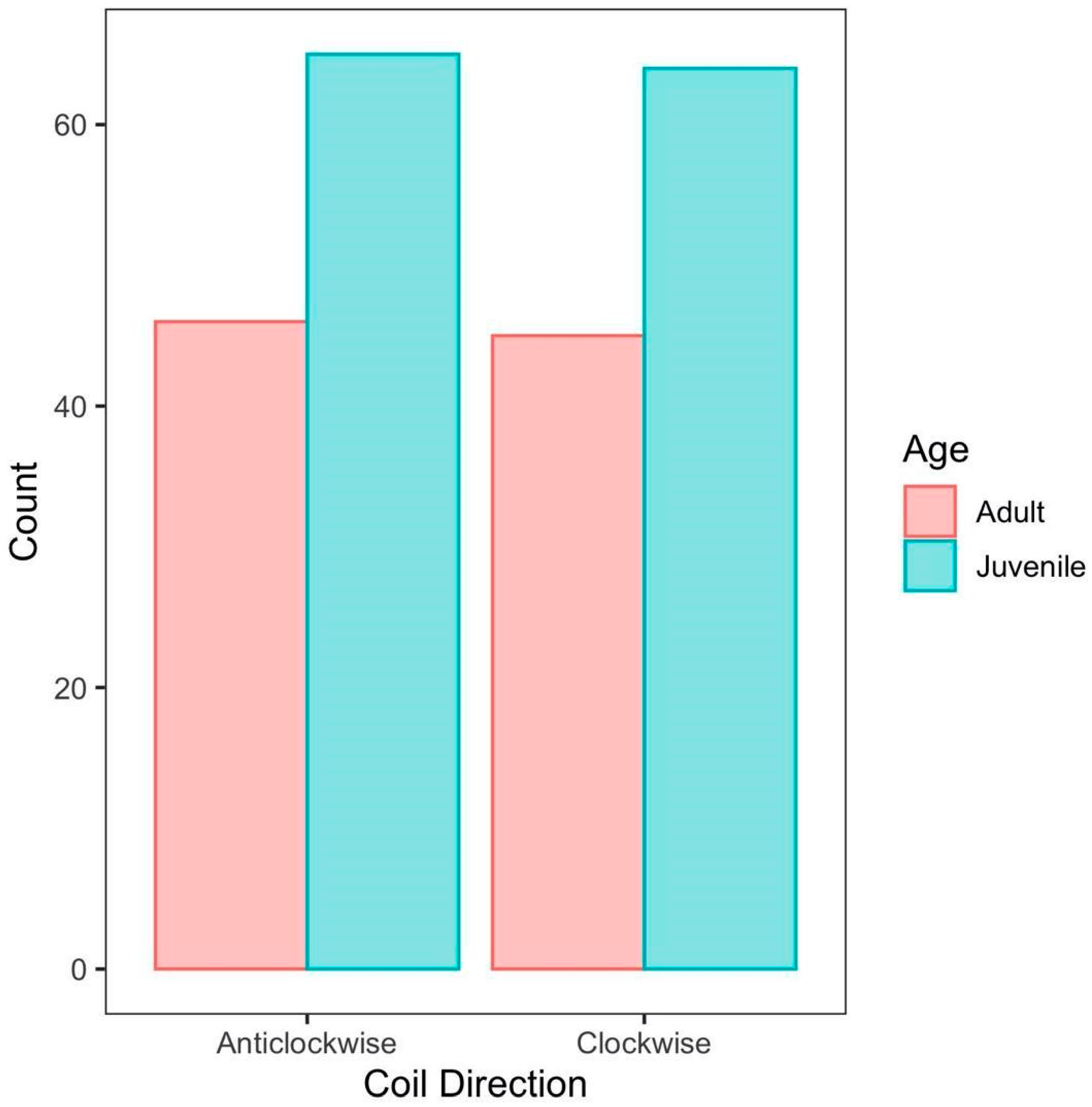
We found that juveniles (X2 = 0.008, df = 1, p-value = 0.930) and adults (X2 = 0.011, df = 1, p = 0.917) were both equally likely to coil clockwise or anticlockwise, and age had no effect on coil direction. (X2 = 0.177, df = 1, p = 0.674, Figure 2). We further found that microhabitat did not affect coil direction (X2 = 7.009, df = 4, p = 0.135), and there was no interaction between age and microhabitat on coil direction (X2 = 4.000, df = 4, p = 0.406).
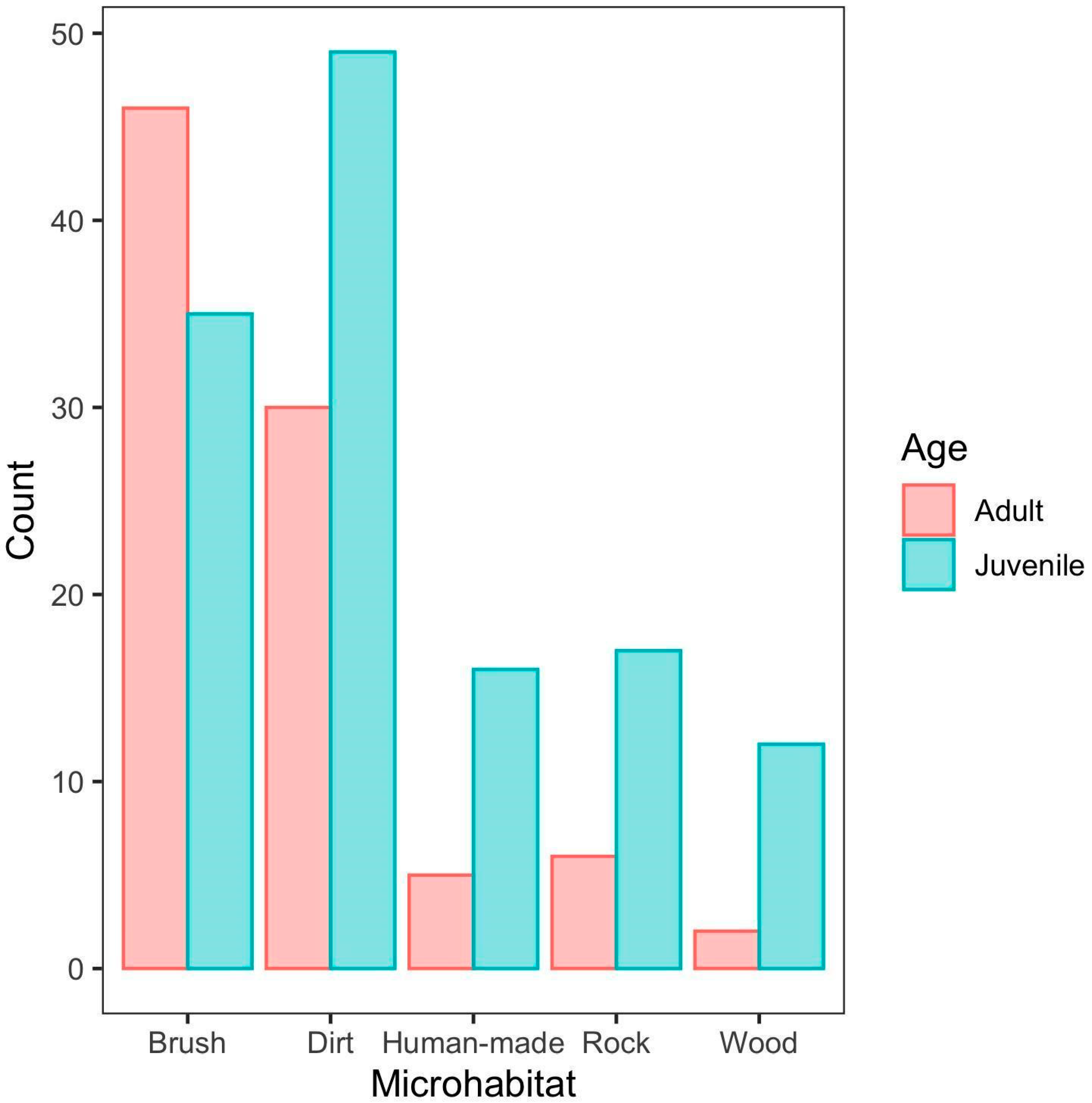
We found that age had an effect on microhabitat utilization (X2 = 17.478, df = 4, p = 0.002). Adults were shown to utilize brush microhabitats in 52% of all observations, while 73% of juvenile observations were recorded in all other microhabitats other than brush (Figure 3). Juveniles were most often observed using dirt (38% of all observations). Both age groups were least likely to be found on wood compared to all other microhabitat categories, especially for adults which only had 2 observations on wood (Figure 3). Juveniles’ microhabitat utilization was more evenly distributed amongst the various categories (brush: 27%, dirt: 38%, human-made: 12%, rock: 13%, wood: 9%), while the brush and dirt microhabitats contributed to 85% of all adult observations (Figure 3).
We could only assess 78 out of the 220 observations for ground level hunting (hunting on ground vs. on an elevated perch) due to most photographs not containing enough background imagery around the snake. We found that the effect of age on ground level hunting was marginally non-significant (X2 = 3.674, df = 1, p = 0.052). Juveniles were more likely to be found on an elevated perch (25% of the time), while adults were almost exclusively found at ground level, and were only found on elevated perches 6% of the time.
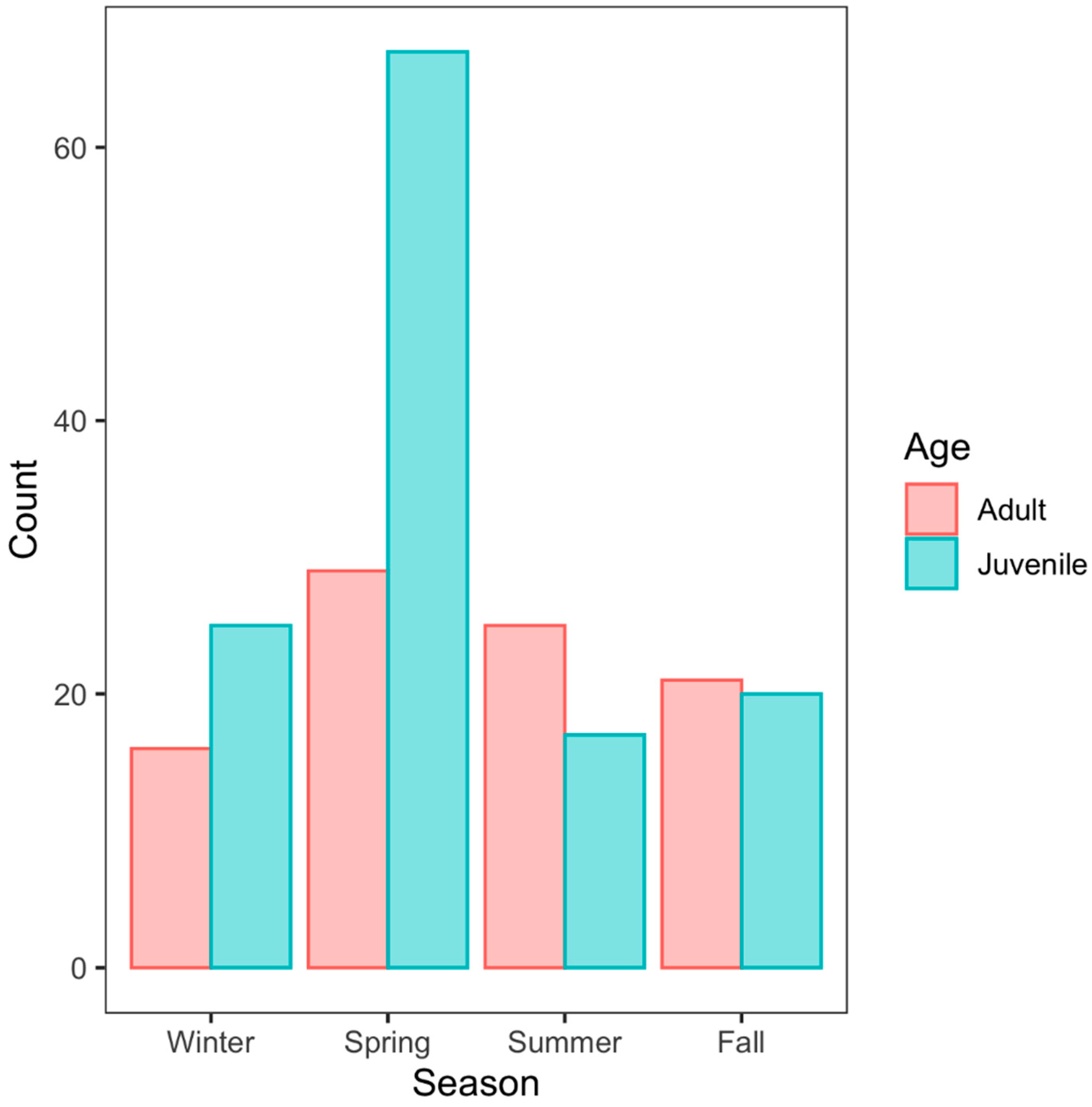
We found that age had an effect on seasonal activity (X2 = 12.371, df = 3, p = 0.006). Juveniles were most commonly observed during spring (52% of all observations) while observations of adults were more evenly dispersed throughout all four seasons with a slight peak during spring (Figure 4).
We found that age did not have an effect on the timing of observations (W = 4615.5, p = 0.566). Juveniles were found hunting 7.65 h after sunrise on average, while adults were found hunting 7.97 h after sunrise on average (Figure 5).
4. Discussion
Our study utilized two community science platforms, iNaturalist and HerpMapper, to quantify aspects of rattlesnake behavior. We processed nearly 4000 observations of Southern Pacific Rattlesnakes (C. helleri), in just under two months, collecting one of the largest datasets on wild juvenile rattlesnake behavior that we know of. Notably, only 220 of these fit our criteria to be included in the study, demonstrating that most observations were not of coiled snakes. Using these community science data, we aimed to find (1) whether Southern Pacific Rattlesnakes exhibit behavioral lateralization when hunting, and (2) whether hunting behaviors change ontogenetically.
In our study, we found that neither juvenile nor adult Southern Pacific Rattlesnakes displayed handedness. Coil direction was also not affected by microhabitat of the ambush site. Snakes exhibit asymmetry in much of their anatomy, such as in lung size [58,59]. Studies have shown that lateralization has been observed in mandibular tooth numbers in snail eating snakes (Pareas iwasakii) [50]. Due to asymmetry being found throughout snake anatomy, it was hypothesized that snakes would exhibit coil asymmetry, or handedness. Other snakes such as Cottonmouths (Agkistrodon piscivorus) have been found to display handedness at the population level [49]. This shows that handedness may differ amongst species. This previous study was also done in the lab, so it could not determine whether coil direction was modified by microhabitat. Why snakes exhibit lateralization is still unknown, but it has been speculated that male snakes may exhibit handedness due to asymmetry of copulatory organs [48]. Our study importantly demonstrates that body postures and orientations, such as coil direction, are straight-forward behavioral traits that can be easily quantified from community science photographs. This is promising for future studies on rattlesnakes and other taxonomic groups for which body posture associates with specific behaviors.
We found that age classes differed in their microhabitat utilization. The difference in hunting tactics could be due to differences in diet, such as juveniles attempting to ambush lizards and adults attempting to ambush small mammals [21,22,23]. Few studies have focused on ontogenetic shifts in the behaviors of free-ranging snakes, likely because it is difficult to track small-bodied juveniles. However, many studies have focused on the diets of rattlesnakes, but these are commonly performed by examining stomach contents of dead specimens, fecal matter, or chemosensory preferences of individuals brought into a lab [21,24,60]. Timber Rattlesnakes (C. horridus) have been found to favor arboreal mammals, which is why they are often seen hunting near trees [61]. Banded Rock Rattlesnakes (C. klauberi), on the other hand, have a diet that mostly consists of lizards, so these snakes are hypothesized to utilize more rocky areas [24]. In our study, juveniles were found to utilize wood and other microhabitats other than brush more often than adults, which could be due to differences in diet. Juvenile rattlesnakes being found in various microhabitats may suggest that their diets may be broader than studies have shown, or rattlesnakes become more specialized in microhabitat use with experience. Juveniles were also more commonly found on human-made materials than adults. This could be due to juveniles attempting to ambush lizards with relaxed antipredator responses in more urban areas [62,63]. Alternatively, adults may avoid human-made microhabitats due to negative experiences with humans or humans might select for individuals that avoid human-made habitats over time (i.e., kill rattlesnakes found in their yards). As urbanization increases, more rattlesnakes may be forced to utilize human-made materials as microhabitats. This causes potential threats to the rattlesnakes, and increased human-rattlesnake interactions, which may lead to snake bites, or snake death [51]. Our data also suggest that age may have an effect on ground level hunting, although we failed to reach statistical significance at alpha = 0.05. Both age groups were most commonly seen hunting at ground level, but juveniles utilized off ground hunting sites more often than adults. Our results may have been insignificant due to a low sample size, as many of the pictures did not include sufficient capturing of the surrounding habitat.
We also found that juveniles were most commonly found during the spring season, while adults had a more evenly dispersed seasonal activity (in terms of observations submitted by community scientists). Some studies have shown that rattlesnake diets change throughout the seasons [23,64]. Many lizards exhibit a spring mating season, leading to higher levels of activity during this period, and this might explain the increase in juvenile rattlesnake observations in spring [65]. These springtime peaks in observations can also be viewed on the iNaturalist website for both juvenile Southern Pacific Rattlesnakes (although this does not distinguish between hunting and non-hunting snakes), and the most commonly observed lizard species, the Western Fence Lizard (Sceloporus occidentalis) (www.inaturalist.org accessed on 20 June 2021). Juveniles may also be commonly observed in spring because they are coming out of winter inactivity and need to gain sustenance in order to reach sexual maturity [45,51]. It is curious that observations of adults remain consistent throughout the year. Previous research suggests that adults have a bimodal mating system with breeding occurring in the spring and fall seasons [55]. This leaves the summer season for foraging [66], yet we did not see a peak in observations during this period. Our results also show that both adult and juvenile rattlesnakes are found coiled even during the winter (i.e., there is no substantial decline in observations) even though this is considered the inactive season. These observations are likely biased toward individuals that occur at low elevations.
Finally, we found that age does not affect the timing of observations, in terms of hours from sunrise. We excluded nighttime observations, due to there being very few observations of rattlesnakes at night. It has been found that snakes use sunrise and sunset as hunting cues [52]. Even though we did not find a significant difference in age and hunting time, it has been hypothesized that predators may change hunting times based on prey activity [25,67]. Even though we did not find differences between age classes, we collected valuable data on the timing of observations that can be plotted out to determine patterns of rattlesnake behavior (Figure 5). These methods can be used for other species of conservation concern or that are difficult to study.
We recognize that some of the data extracted from community science platforms will contain biases due to the areas where humans take their observations and to the times at which human observers are active (e.g., we have very few nighttime observations). Thus, our dataset will represent daytime behaviors of rattlesnakes, which could potentially change at night. Although observations of accessible or easily-observed individuals are likely to make up the majority of our dataset, we still find significant differences between adults and juveniles in microhabitat use and seasonality of observations. Overall, these types of biases are no different than those associated with studies using traditional field methods, and we feel the large sample sizes that can be attained through community science often provide a large benefit in comparison to these costs. As an example, our dataset is likely the largest one to date on juvenile rattlesnake hunting behaviors. Notably, we were able to account for potential repeated observations of the same individual snakes by taking advantage of the locational data associated with observations, and we also found that most people submit only 1–2 observations (Supplementary Materials Figure S1) so there is reduced bias in user identity affecting the results.
As community science continues to grow, scientists will be able to crowd-source large datasets from which they can address research questions that may have been previously impossible or logistically difficult to answer. Yet, there are still limitations with these data. For our study, we found that only 5% of all relevant observations submitted to iNaturalist or HerpMapper met our inclusion criteria. This is likely the result of the cryptic nature of rattlesnakes and the difficulty non-experts might have in detecting coiled snakes that are well-camouflaged with their surroundings. Second, most photographs submitted to community science platforms are of close-ups or cropped images of the focal animal, which are ideal for extracting morphological or behavioral data, but make the extraction of habitat data more difficult. For example, we were unable to fully analyze whether snakes were hunting on the ground or on a perch off the ground because most photographs did not have sufficient background imagery (at least one body length away from the focal animal) for us to quantify this. Although we recognize that many users of these community science platforms submit observations for various reasons other than to specifically contribute to scientific research, we advocate for better communication with users on how they might enhance the usefulness of their observations. For instance, users can be encouraged to provide a close-up photograph of the focal animal, along with another image showing the surrounding habitat. Providing comments to observations, similar to field notes, would also be helpful in giving more context to the photographs.
Through community science, we were able to avoid many of the obstacles that come with field research. Past studies utilizing traditional field methods encountered obstacles when studying predatory animals. For example, tracking predators can be costly, and can sometimes lead to difficulties during the implantation of spatial tracking devices [68]. Not only did we overcome time and logistical barriers to tracking predators, but our study was ultimately free of monetary cost. Furthermore, taking a community science approach can lead to additional outcomes beyond research results. Studies that utilize community science for data collection found that volunteers who contributed to community science gained a better understanding and appreciation of the ecosystems around them, leading to positive support for conservation efforts [69,70].
5. Conclusions
Our study shows the promise of using community science to study the hunting behaviors of cryptic predators that may be relatively abundant, but logistically difficult to study. We can extract basic behavioral data such as microhabitat use, body posture and orientation, and timing of events, and age class data to examine ontogeny of behaviors. We found an ontogenetic shift in microhabitat use and a difference in the timing of observations between juveniles and adults with juveniles experiencing a large peak in observations during the spring season. Behavioral lateralization in coil direction does not seem to be apparent in this species, although it could be present within individuals. We provide recommendations to community scientists and/or community science facilitators for improving the usefulness of their observations for scientific research. Overall, our study contributes to the growing literature on the use of community science for the study of animal behavior.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/d13080349/s1, Figure S1: Histogram showing density of number of observations per observer, Table S1: Final Data Sheet.
Author Contributions
Conceptualization, E.R.U. and B.J.P.; methodology, B.J.P.; data collection, E.R.U.; data analysis, E.R.U. and B.J.P.; writing—original draft preparation, E.R.U. and B.J.P.; writing—review and editing, E.R.U. and B.J.P.; visualization, E.R.U. and B.J.P.; supervision, B.J.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not Applicable.
Data Availability Statement
The data presented in this study are available in the attached Supplementary Files.
Acknowledgments
We thank Enjie Li for helping with R code for some of the analyses, Christopher Smith and the staff at HerpMapper for granting access to location data, and all the community scientists for contributing the observations used in this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Burkholder, D.A.; Heithaus, M.R.; Fourqurean, J.W.; Wirsing, A.; Dill, L.M. Patterns of top-down control in a seagrass ecosystem: Could a roving apex predator induce a behaviour-mediated trophic cascade? J. Anim. Ecol. 2013, 82, 1192–1202. [Google Scholar] [CrossRef]
- Holt, A.R.; Davies, Z.G.; Tyler, C.; Staddon, S. Meta-Analysis of the effects of predation on animal prey abundance: Evidence from uk vertebrates. PLoS ONE 2008, 3, e2400. [Google Scholar] [CrossRef]
- Brown, J.S.; Kotler, B.P.; Smith, R.J.; Wirtz II, W.O. The effects of owl predation on the foraging behavior of heteromyid rodents. Oecologia 1988, 76, 408–415. [Google Scholar] [CrossRef]
- Woo, K.J.; Elliott, K.H.; Davidson, M.; Gaston, A.J.; Davoren, G.K. Individual specialization in diet by a generalist marine predator reflects specialization in foraging behaviour. J. Anim. Ecol. 2008, 77, 1082–1091. [Google Scholar] [CrossRef]
- Dézerald, O.; Céréghino, R.; Corbara, B.; Dejean, A.; Leroy, C. Functional trait responses of aquatic macroinvertebrates to simulated drought in a Neotropical bromeliad ecosystem. Freshw. Biol. 2015, 60, 1917–1929. [Google Scholar] [CrossRef]
- Brossard, D.; Lewenstein, B.; Bonney, R. Scientific knowledge and attitude change: The impact of a citizen science project. Int. J. Sci. Educ. 2005, 27, 1099–1121. [Google Scholar] [CrossRef]
- Cooper, C.B.; Dickinson, J.; Phillips, T.; Bonney, R. Citizen science as a tool for conservation in residential ecosystems. Ecol. Soc. 2007, 12, 11. [Google Scholar] [CrossRef]
- Thomson, R.C.; Wright, A.N.; Shaffer, H.B. California Amphibian and Reptile Species of Special Concern; University of California Press: Oakland, CA, USA, 2016. [Google Scholar]
- Martin, W.H.; Brown, W.S.; Possardt, E.; Sealy, J.B. The Biology of Rattlesnakes; Hayes, W.K., Beaman, K.R., Cardwell, M.D., Bush, S.P., Eds.; Loma Linda University Press: Loma Linda, CA, USA, 2008. [Google Scholar]
- Waldron, J.L.; Lanham, J.D.; Bennett, S.H. Using behaviorally-based seasons to investigate canebrake rattlesnake (Crotalus horridus) movement patterns and habitat selection. Herpetologica 2006, 62, 389–398. [Google Scholar] [CrossRef]
- Lowe, C.H. The Venomous Reptiles of Arizona; Arizona Game and Fish Department: Phoenix, AZ, USA, 1986. [Google Scholar]
- Ernst, C.H. Venomous Reptiles of North America; Smithsonian Books: Washington, DC, USA, 1999. [Google Scholar]
- Szymanski, J.; Pollack, C.; Ragan, L.; REdmer, M.; Clemency, L.; Voorhies, K.; Jaka, J. Species Status Assessment for the Eastern Massasauga Rattlesnake (Sistrurus catenatus); U.S. Fish and Wildlife: Bloomington, IN, USA, 2016.
- Martin, W.H.; Means, B.D. Distribution and habitat relationships of the eastern diamondback rattlesnake (Crotalus adamanteus). Herpetol. Nat. Hist. 2000, 7, 9–34. [Google Scholar]
- Stohlgren, K.M.; Spear, S.F.; Stevenson, D.J. A Status and Distribution Model for the Eastern Diamondback Rattlesnake (Crotalus adamanteus) in Georgia; U.S. Fish and Wildlife: Tiger, GA, USA, 2015.
- Hect, K.A.; Nickerson, M.A.; Colclough, P.B. Hellbenders (Cryptobranchus alleganiensis) may exhibit an ontogenetic dietary shift. Southeast. Nat. 2017, 16, 157–162. [Google Scholar] [CrossRef]
- Subalusky, A.; Fitzgerald, L.A.; Smith, L.L. Ontogenetic niche shifts in the american alligator establish functional connectivity between aquatic systems. Biol. Conserv. 2009, 142, 1507–1514. [Google Scholar] [CrossRef]
- Cundall, D.; Greene, H. Feeding: Form, Function, and Evolution in Tetrapod Vertebrates; Schwenk, K., Ed.; Academic Press: San Diego, CA, USA, 2000. [Google Scholar]
- Clark, R.W.; Tangco, S.; Barbour, M.A. Field video recordings reveal factors influencing predatory strike success of free-ranging rattlesnakes (Crotalus spp.). Anim. Behav. 2012, 84, 183–190. [Google Scholar] [CrossRef]
- Harmel, M.V.; Crowell, H.L.; Whelan, J.M.; Taylor, E.N. Rattlesnake colouration affects detection by predators. J. Zool. 2020, 311, 260–268. [Google Scholar] [CrossRef]
- Taylor, E.N. Diet of the Baja California Rattlesnake, Crotalus enyo (Viperidae). Copeia 2001, 2001, 553–555. [Google Scholar] [CrossRef]
- Mociño-Deloya, E.; Setser, K.; Heacker, M.; Peurach, S. Diet of New Mexico ridge-nosed rattlesnake (Crotalus willardi obscurus) in the Sierra San Luis and Sierra Pan Duro, México. J. Herpetol. 2015, 48, 104–107. [Google Scholar] [CrossRef]
- Glaudas, X.; Jezkova, T.; Rodrıguez-Robles, J.A. Feeding ecology of the Great Basin Rattlesnake (Crotalus lutosus, Viperidae). Can. J. Zool. 2008, 86, 723–734. [Google Scholar] [CrossRef] [Green Version]
- Holycross, A.T.; Painter, C.W.; Prival, D.B.; Swann, D.E.; Schroff, M.J.; Edwards, T.; Schwalbe, C.R. Diet of Crotalus lepidus klauberi (Banded Rock Rattlesnake). J. Herpetol. 2002, 36, 589–597. [Google Scholar] [CrossRef]
- Clark, R.W.; Dorr, S.W.; Whitford, M.D.; Freymiller, G.A.; Putman, B.J. Activity cycles and foraging behaviors of free-ranging sidewinder rattlesnakes (Crotalus cerastes): The ontogeny of hunting in a precocial vertebrate. Zoology 2016, 119, 196–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avila-Villegas, H.; Martins, M.; Arnaud, G. Feeding ecology of the endemic rattleless rattlesnake, Crotalus catalinensis, of Santa Catalina Island, Gulf of California, Mexico. Copeia 2007, 1, 80–84. [Google Scholar] [CrossRef]
- Lomas, E.; Maida, J.R.; Bishop, C.A.; Larsen, K.W. Movement ecology of northern pacific rattlesnakes (Crotalus o. oreganus) in response to disturbance. Herpetologica 2019, 75, 153–161. [Google Scholar] [CrossRef]
- Harvey, D.S.; Weatherhead, P.J. Habitat selection as the mechanism for thermoregulation in a northern population of massasauga rattlesnakes (Sistrurus catenatus). Ecoscience 2010, 17, 411–419. [Google Scholar] [CrossRef]
- Lind, C.M.; Beaupre, S.J. Natural variation in steroid hormone profiles of male Timber Rattlesnakes, Crotalus horridus, in northwest Arkansas. Gen. Comp. Endocrinol. 2014, 206, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Capehart, G.D.; Escallón, C.; Vernasco, B.J.; Moore, I.T.; Taylor, E.N. No drought about it: Effects of supplemental hydration on the ecology, behavior, and physiology of free-ranging rattlesnakes. J. Arid Environ. 2016, 134, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Jaramillo-Alba, J.L.; Díaz de la Vega-Pérez, A.H.; Bucio-Jiménez, L.E.; Méndez-De la Cruz, F.R.; Pérez-Mendoza, H.A. Comparative thermal ecology parameters of the mexican dusky rattlesnake (Crotalus triseriatus). J. Therm. Biol. 2020, 92, 102695. [Google Scholar] [CrossRef] [PubMed]
- Hale, V.L.; Macgowan, B.; Corriveau, L.; Huse, D.C.; Currylow, A.F.T.; Thompson, S. Radio transmitter implantation and movement in the wild timber rattlesnake (Crotalus horridus). J. Wildl. Dis. 2017, 53, 591–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mata-Silva, V.; Desantis, D.L.; Wagler, A.E.; Johnson, J.D. Spatial ecology of Rock Rattlesnakes (Crotalus lepidus) in far West Texas. Herpetologica 2018, 74, 245–254. [Google Scholar] [CrossRef]
- Amarello, M.; Nowak, E.M.; Taylor, E.N.; Schuett, G.W.; Repp, R.A.; Rosen, P.C.; Hardy, D.L. Potential environmental influences on variation in body size and sexual size dimorphism among Arizona populations of the western diamond-backed rattlesnake (Crotalus atrox). J. Arid Environ. 2010, 74, 1443–1449. [Google Scholar] [CrossRef]
- Petersen, C.E.; Goetz, S.M.; Dreslik, M.J.; Kleopfer, J.D.; Savitzky, A.H. Sex, mass, and monitoring effort: Keys to understanding spatial ecology of timber rattlesnakes (Crotalus horridus). Herpetologica 2019, 75, 162–174. [Google Scholar] [CrossRef]
- Wallace, R.L.; Diller, L. V Feeding Ecology of the Rattlesnake, Crotalus viridis oreganus, in Northern Idaho. J. Herpetol. 1990, 24, 246–253. [Google Scholar] [CrossRef]
- Bonney, R.; Phillips, T.B.; Ballard, H.L.; Enck, J.W. Can citizen science enhance public understanding of science? Public Underst. Sci. 2016, 25, 2–16. [Google Scholar] [CrossRef]
- Newman, G.; Chandler, M.; Clyde, M.; McGreavy, B.; Haklay, M.; Ballard, H.; Gray, S.; Scarpino, R.; Hauptfeld, R.; Mellor, D.; et al. Leveraging the power of place in citizen science for effective conservation decision making. Biol. Conserv. 2016, 208, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Kobori, H.; Dickinson, J.L.; Washitani, I.; Sakurai, R.; Amano, T.; Komatsu, N.; Kitamura, W.; Takagawa, S.; Koyama, K.; Ogawara, T.; et al. Citizen science: A new approach to advance ecology, education, and conservation. Ecol. Res. 2016, 31, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Losey, J.E.; Perlman, J.E.; Hoebeke, E.R. Citizen scientist rediscovers rare nine-spotted lady beetle, Coccinella novemnotata, in eastern North America. J. Insect Conserv. 2007, 11, 415–417. [Google Scholar] [CrossRef]
- Ziter, C.D.; Herrick, B.M.; Johnston, M.R.; Turner, M.G. Ready, set, go: Community science field campaign reveals habitat preferences of nonnative asian earthworms in an urban landscape. Bioscience 2021, 71, 280–291. [Google Scholar] [CrossRef]
- Callaghan, C.T.; Bino, G.; Major, R.E.; Martin, J.M.; Lyons, M.B.; Kingsford, R.T. Heterogeneous urban green areas are bird diversity hotspots: Insights using continental-scale citizen science data. Landsc. Ecol. 2019, 34, 1231–1246. [Google Scholar] [CrossRef]
- Lye, G.C.; Osborne, J.L.; Park, K.J.; Goulson, D. Using citizen science to monitor Bombus populations in the UK: Nesting ecology and relative abundance in the urban environment. J. Insect Conserv. 2012, 16, 697–707. [Google Scholar] [CrossRef]
- Putman, B.J.; Williams, R.; Li, E.; Pauly, G.B. The power of community science to quantify ecological interactions in cities. Sci. Rep. 2021, 11, 3069. [Google Scholar] [CrossRef]
- Klauber, L.M. Rattlesnakes, Their Habits, Life Histories, and Influence on Mankind; Abridged; University of California Press: Berkeley, CA, USA, 1982; ISBN 0520040392, 9780520040397. [Google Scholar]
- Hughes, A.L.; Cauthen, J.; Driscoll, C. Testing for behavioral lateralization in observational data: A monte carlo approach applied to neck-looping in american flamingos. Wilson J. Orinthol. 2014, 126, 345–352. [Google Scholar] [CrossRef]
- Frasnelli, E. Brain and behavioral lateralization in invertebrates. Front. Psychol. 2013, 4, 939. [Google Scholar] [CrossRef] [Green Version]
- Shine, R.; Olsson, M.M.; LeMaster, M.P.; Moore, I.T.; Mason, R.T. Are snakes right-handed? Asymmetry in hemipenis size and usage in gartersnakes (Thamnophis sirtalis). Behav. Ecol. 2000, 11, 411–415. [Google Scholar] [CrossRef] [Green Version]
- Roth, E.D. “Handedness” in snakes? Lateralization of coiling behaviour in a cottonmouth, Agkistrodon piscivorus leucostoma, population. Anim. Behav. 2003, 66, 337–341. [Google Scholar] [CrossRef] [Green Version]
- Hoso, M.; Asami, T.; Hori, M. Right-handed snakes: Convergent evolution of asymmetry for functional specialization. Biol. Lett. 2007, 3, 169–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stebbins, R.C.; McGinnis, S.M. Amphibians and Reptiles of California: Revised Edition, 1st ed.; University of California Press: Berkely, CA, USA, 2012; ISBN 9780520270510. [Google Scholar]
- Putman, B.J.; Barbour, M.A.; Clark, R.W. The Foraging Behavior of Free-ranging Rattlesnakes (Crotalus oreganus) in California Ground Squirrel (Otospermophilus beecheyi) Colonies. Herpetologica 2016, 72, 55–63. [Google Scholar] [CrossRef]
- Wittenberg, R.D. Foraging ecology of the Timber Rattlesnake (Crotalus horridus) in a fragmented agricultural landscape. Herpetol. Conserv. Biol. 2012, 7, 449–461. [Google Scholar]
- Webber, M.M.; Glaudas, X.; Rodríguez-Robles, J.A. Do sidewinder rattlesnakes (Crotalus cerastes, Viperidae) cease feeding during the breeding season? Copeia 2012, 100–105. [Google Scholar] [CrossRef]
- Lind, C.M.; Husak, J.F.; Eikenaar, C.; Moore, I.T.; Taylor, E.N. The relationship between plasma steroid hormone concentrations and the reproductive cycle in the Northern Pacific rattlesnake, Crotalus oreganus. Gen. Comp. Endocrinol. 2010, 166, 590–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Putman, B.J.; Clark, R.W. Behavioral thermal tolerances of free-ranging rattlesnakes (Crotalus oreganus) during the summer foraging season. J. Therm. Biol. 2017, 65, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Putman, B.J.; Lind, C.; Taylor, E.N. Does size matter? Factors influencing the spatial ecology of northern pacific rattlesnakes (Crotalus oreganus oreganus) in Central California. Copeia 2013, 2013, 485–492. [Google Scholar] [CrossRef]
- Kardong, K.V. Morphology of the respiratory system and its musculature in different snake genera. II. Charina bottae. Gegenbaurs Morphol. Jahrb. 1972, 117, 364–376. [Google Scholar] [PubMed]
- van Soldt, B.J.; Metscher, B.D.; Poelmann, R.E.; Vervust, B.; Vonk, F.J.; Muller, G.B.; Richardson, M.K. Heterochrony and early left-right asymmetry in the development of the cardiorespiratory system of snakes. PLoS ONE 2015, 10, e116416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saviola, A.J.; Chiszar, D.; Mackessy, S.P. Ontogenetic shift in response to prey-derived chemical cues in prairie rattlesnakes Crotalus viridis viridis. Curr. Zool. 2012, 58, 549–555. [Google Scholar] [CrossRef]
- Goetz, S.M.; Petersen, C.E.; Rose, R.K.; Kleopfer, J.D.; Savitzky, A.H. Diet and foraging behaviors of Timber Rattlesnakes, Crotalus horridus, in Eastern Virginia. J. Herpetol. 2016, 50, 520–526. [Google Scholar] [CrossRef] [Green Version]
- Grolle, E.K.; Lopez, M.C.; Gerson, M.M. Flight initiation distance differs between populations of Western Fence Lizards (Sceloporus occidentalis) at a rural and an urban Site. Bull. South. Calif. Acad. Sci. 2014, 113, 42–46. [Google Scholar] [CrossRef] [Green Version]
- Samia, D.S.M.; Nakagawa, S.; Nomura, F.; Rangel, T.F.; Blumstein, D.T. Increased tolerance to humans among disturbed wildlife. Nat. Commun. 2015, 6, 9877. [Google Scholar] [CrossRef] [Green Version]
- Webber, M.M.; Jezkova, T.; Rodríguez-Robles, J.A. Feeding Ecology of Sidewinder Rattlesnakes, Crotalus cerastes (Viperidae). Herpetologica 2016, 72, 324–330. [Google Scholar] [CrossRef]
- Stebbins, R.C. A Field Guide to Western Reptiles and Amphibians: Field Marks of all Species in Western North America, 18th ed.; Houghton Mifflin: Boston, MA, USA, 1966. [Google Scholar]
- Clark, R.W. Fixed videography to study predation behavior of an ambush forgaing snake, Crotalus horridus. Copeia 2006, 2, 181–187. [Google Scholar] [CrossRef]
- Lang, S.D.J.; Mann, R.P.; Farine, D.R. Temporal activity patterns of predators and prey across broad geographic scales. Behav. Ecol. 2019, 30, 172–180. [Google Scholar] [CrossRef] [Green Version]
- Jungen, M.T. Eastern Diamondback Rattlesnake (Crotalus adamanteus) Telemetry Techniques and Translocation; Marshall University: Huntington, WV, USA, 2018. [Google Scholar]
- Bela, G.; Peltola, T.; Young, J.C.; Balázs, B.; Arpin, I.; Pataki, G.; Hauck, J.; Kelemen, E.; Kopperoinen, L.; Van Herzele, A.; et al. Learning and the transformative potential of citizen science. Conserv. Biol. 2016, 30, 990–999. [Google Scholar] [CrossRef] [Green Version]
- MacPhail, V.J.; Gibson, S.D.; Colla, S.R. Community science participants gain environmental awareness and contribute high quality data but improvements are needed: Insights from bumble bee watch. PeerJ 2020, 8, e9141. [Google Scholar] [CrossRef]
Figure 1.
Filtering Process for Observations. Our community science searches yielded ~4000 observations. We disregarded any observations containing snakes that were dead, loosely coiled, elongated, duplicates, or found under cover boards. After filtering the observations, we were left with 220 observations of tightly coiled, hunting Southern Pacific Rattlesnakes.
Figure 1.
Filtering Process for Observations. Our community science searches yielded ~4000 observations. We disregarded any observations containing snakes that were dead, loosely coiled, elongated, duplicates, or found under cover boards. After filtering the observations, we were left with 220 observations of tightly coiled, hunting Southern Pacific Rattlesnakes.

Figure 2.
Age and Coil Direction. Snakes were divided by age groups (juvenile and adult) and coil directions were quantified as either clockwise or anticlockwise.
Figure 2.
Age and Coil Direction. Snakes were divided by age groups (juvenile and adult) and coil directions were quantified as either clockwise or anticlockwise.

Figure 3.
Microhabitat Distribution. The relationship between microhabitat utilization and age group.
Figure 3.
Microhabitat Distribution. The relationship between microhabitat utilization and age group.

Figure 4.
Age and Season. The relationship between age and seasonal activity. Seasons were based on biological behaviors. Winter included November–February and has been observed as an inactive period. Spring included March–May which was considered the mating season. Summer was June–July which is when snakes are often found foraging. Fall included August–October which is when snakes give birth.
Figure 4.
Age and Season. The relationship between age and seasonal activity. Seasons were based on biological behaviors. Winter included November–February and has been observed as an inactive period. Spring included March–May which was considered the mating season. Summer was June–July which is when snakes are often found foraging. Fall included August–October which is when snakes give birth.

Figure 5.
Age and Hours from Sunrise. Histograms showing when rattlesnakes were observed in terms of hours from sunrise. Calculated average values of hours from sunrise for adults and juveniles are indicated by the dashed vertical lines.
Figure 5.
Age and Hours from Sunrise. Histograms showing when rattlesnakes were observed in terms of hours from sunrise. Calculated average values of hours from sunrise for adults and juveniles are indicated by the dashed vertical lines.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Urquidi, E.R.; Putman, B.J. Quantifying Southern Pacific Rattlesnake (Crotalus oreganus helleri) Hunting Behavior through Community Science. Diversity 2021, 13, 349. https://0-doi-org.brum.beds.ac.uk/10.3390/d13080349
AMA Style
Urquidi ER, Putman BJ. Quantifying Southern Pacific Rattlesnake (Crotalus oreganus helleri) Hunting Behavior through Community Science. Diversity. 2021; 13(8):349. https://0-doi-org.brum.beds.ac.uk/10.3390/d13080349
Chicago/Turabian StyleUrquidi, Emily R., and Breanna J. Putman. 2021. "Quantifying Southern Pacific Rattlesnake (Crotalus oreganus helleri) Hunting Behavior through Community Science" Diversity 13, no. 8: 349. https://0-doi-org.brum.beds.ac.uk/10.3390/d13080349
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.