
Exploring the Genetic Diversity among Weedy Rice Accessions Differing in Herbicide Tolerance and Allelopathic Potential

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Genomic DNA Extraction
2.2. Polymerase Chain Reaction (PCR) Using Simple Sequence Repeat (SSR) Markers
2.3. Data Analysis
3. Results
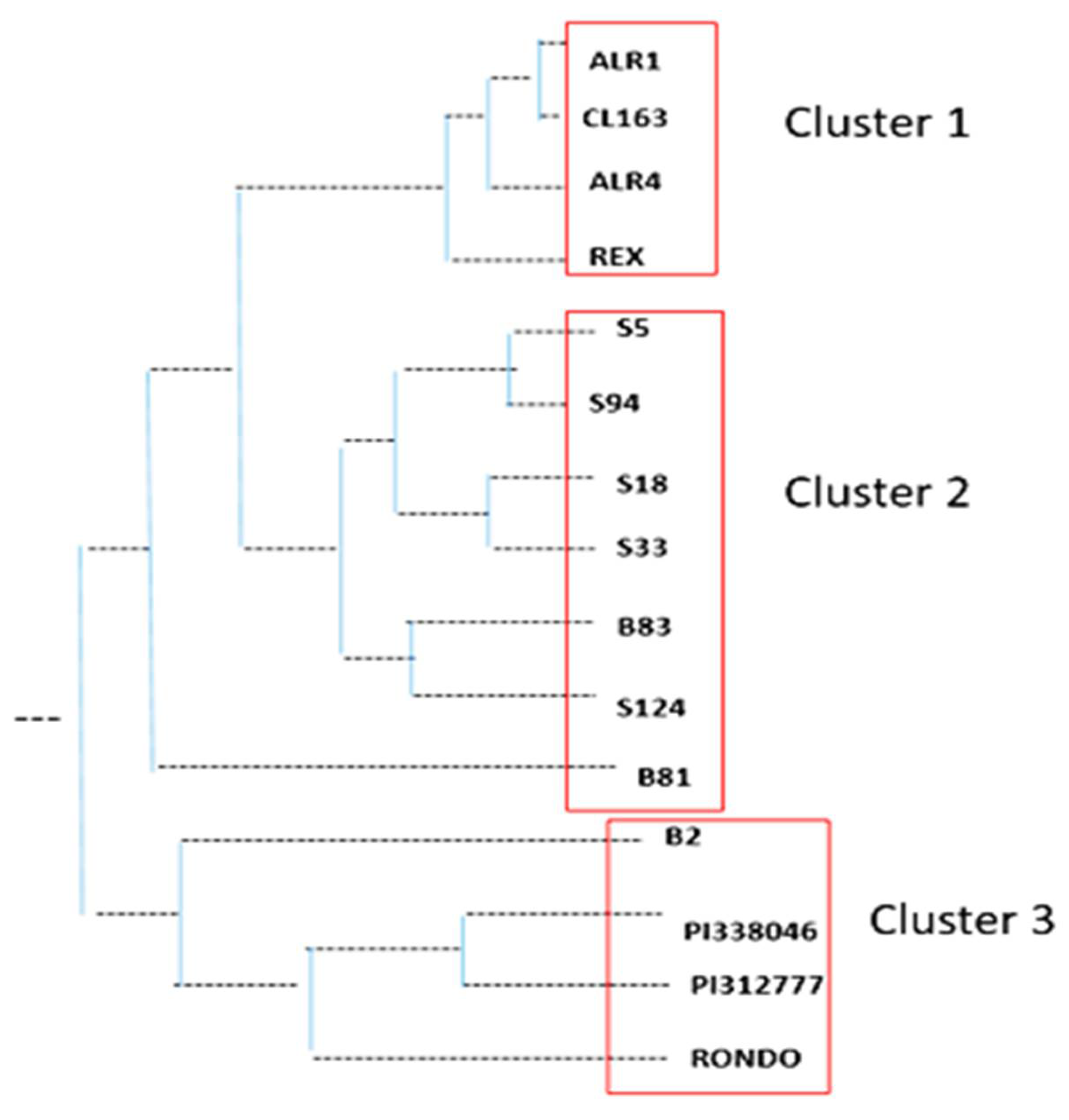
3.1. Genetic Diversity among Weedy Rice, Cultivated Rice, and Allelopathic Rice
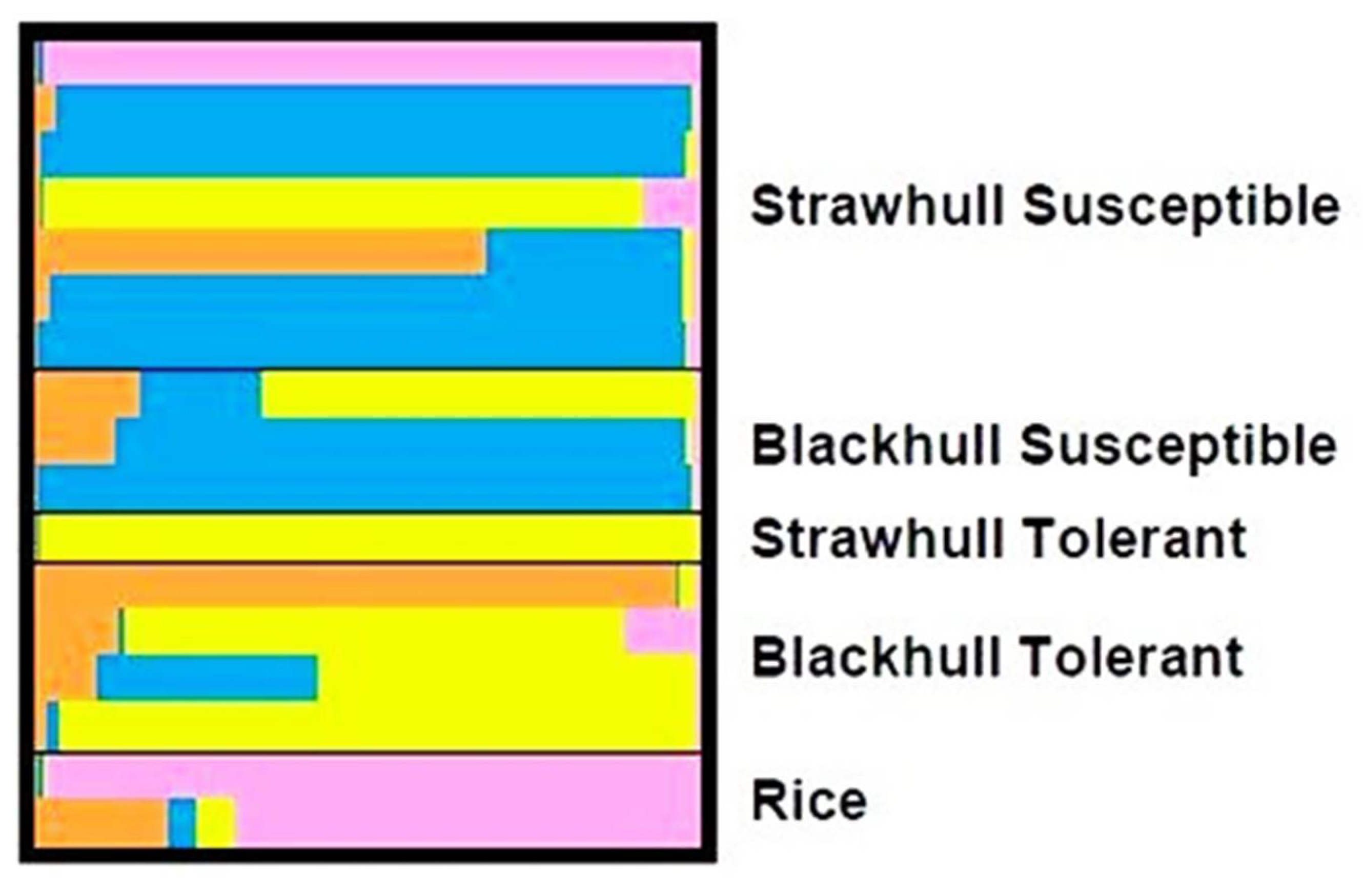
3.2. Genetic Relationship and Differentiation Based on Herbicide Tolerance
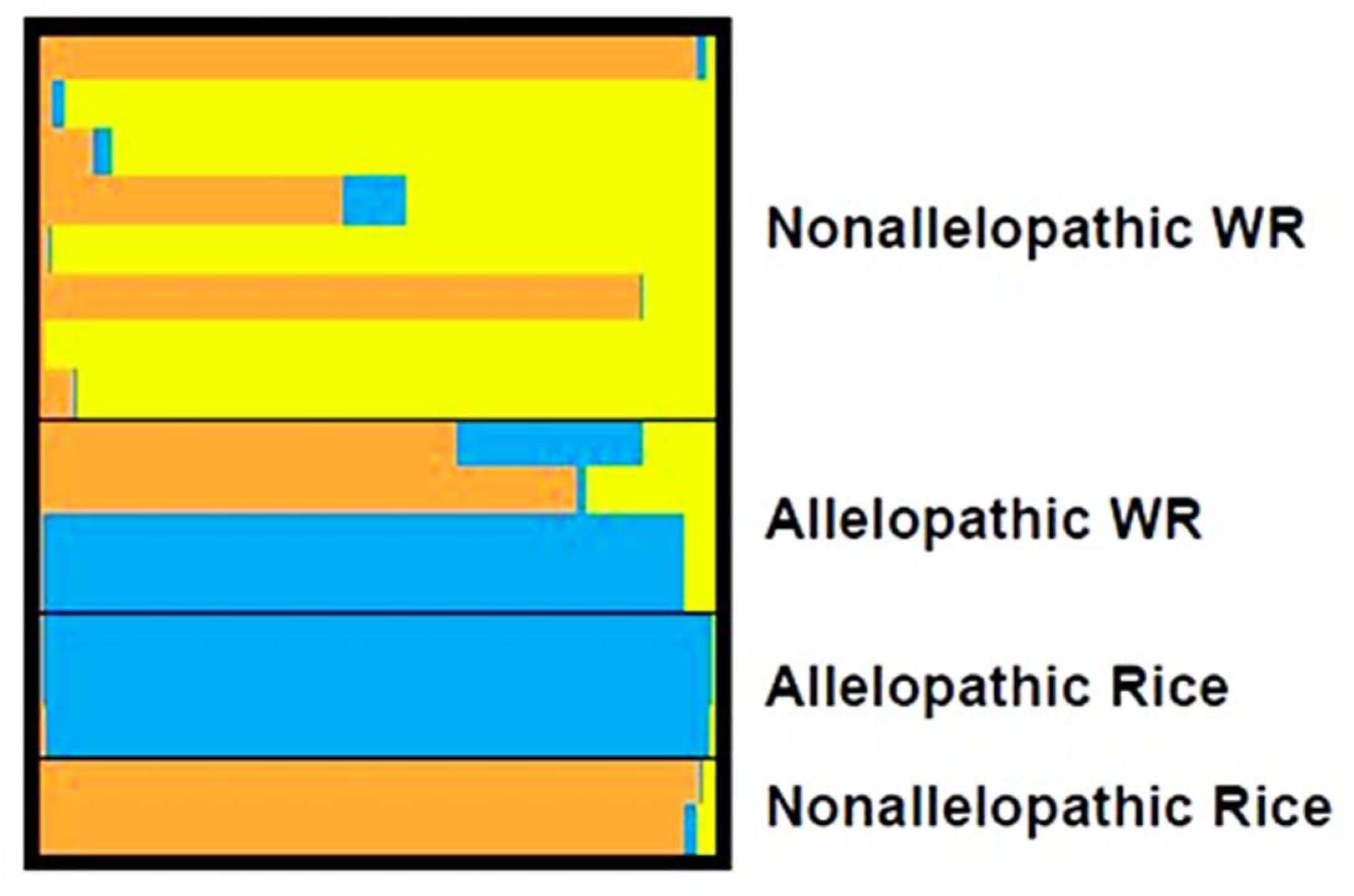
3.3. Genetic Diversity among the Accessions with Respect to Allelopathic Potential
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singh, V.; Zhou, S.; Ganie, Z.; Valverde, B.; Avila, L.; Marchesan, E.; Merotto, A.; Zorrilla, G.; Burgos, N.; Norsworthy, J.; et al. Rice Production in the Americas. In Rice Production Worldwide; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 137–168. [Google Scholar]
- Allston, R.F. The Rice Plant; University of Michigan, Humanities Text Initiative: Ann Arbor, MI, USA, 1846. [Google Scholar]
- Hoagland, R.E.; Paul, R.N. A Comparative SEM Study of Red Rice and Several Commerical Rice (Oryza sativa) Varieties. Weed Sci. 1978, 26, 619–625. [Google Scholar] [CrossRef]
- Burgos, N.R.; Norsworthy, J.; Scott, R.C.; Smith, K.L. Red Rice (Oryza sativa) Status after 5 Years of Imidazolinone-Resistant Rice Technology in Arkansas. Weed Technol. 2008, 22, 200–208. [Google Scholar] [CrossRef]
- Burgos, N.R.; Singh, V.; Tseng, T.-M.P.; Black, H.; Young, N.D.; Huang, Z.; Hyma, K.E.; Gealy, D.R.; Caicedo, A.L. The Impact of Herbicide-Resistant Rice Technology on Phenotypic Diversity and Population Structure of United States Weedy Rice. Plant Physiol. 2014, 166, 1208–1220. [Google Scholar] [CrossRef] [Green Version]
- Shivrain, V.K.; Burgos, N.R.; Gealy, D.R.; A Sales, M.; Smith, K.L. Gene flow from weedy red rice (Oryza sativa L.) to cultivated rice and fitness of hybrids. Pest Manag. Sci. 2009, 65, 1124–1129. [Google Scholar] [CrossRef]
- Gealy, D. Gene Movement between Rice (Oryza sativa) and Weedy Rice (Oryza sativa)—A U.S. Temperate Rice Perspective. Crop Feral. Volunt. 2005, 2, 323–354. [Google Scholar]
- Shivrain, V.K.; Burgos, N.R.; Agrama, H.A.; Lawton-Rauh, A.; Lu, B.; Sales, M.A.; Boyett, V.; Gealy, D.R.; Moldenhauer, K.A.K. Genetic diversity of weedy red rice (Oryza sativa) in Arkansas, USA. Weed Res. 2010, 50, 289–302. [Google Scholar] [CrossRef]
- Shivrain, V.K.; Burgos, N.R.; Scott, R.C.; Gbur, E.E., Jr.; Estorninos, L.E., Jr.; McClelland, M.R. Diversity of weedy red rice (Oryza sativa L.) in Arkansas, USA in relation to weed management. Crop Prot. 2010, 29, 721–730. [Google Scholar] [CrossRef]
- Tseng, T.-M.; Shivrain, V.K.; Lawton-Rauh, A.; Burgos, N.R. Dormancy-linked Population Structure of Weedy Rice (Oryza sp.). Weed Sci. 2018, 66, 331–339. [Google Scholar] [CrossRef]
- Noldin, J.A.; Chandler, J.M.; McCauley, G.N. Red Rice (Oryza sativa) Biology. I. Characterization of Red Rice Ecotypes. Weed Technol. 1999, 13, 12–18. [Google Scholar] [CrossRef]
- Cohn, M.A.; Hughes, J.A. Seed Dormancy in Red Rice (Oryza sativa) I. Effect of Temperature on Dry-Afterripening. Weed Sci. 1981, 29, 402–404. [Google Scholar] [CrossRef]
- Cao, Q.; Lu, B.-R.; Xia, H.; Rong, J.; Sala, F.; Spada, A.; Grassi, F. Genetic Diversity and Origin of Weedy Rice (Oryza sativa f. spontanea) Populations Found in North-eastern China Revealed by Simple Sequence Repeat (SSR) Markers. Ann. Bot. 2006, 98, 1241–1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Qi, X.; Gealy, D.R.; Olsen, K.M.; Caicedo, A.L.; Jia, Y. QTL Analysis for Resistance to Blast Disease in U.S. Weedy Rice. Mol. Plant-Microbe Interact. 2015, 28, 834–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziska, L.H.; McClung, A. Differential Response of Cultivated and Weedy (Red) Rice to Recent and Projected Increases in Atmospheric Carbon Dioxide. Agron. J. 2008, 100, 1259–1263. [Google Scholar] [CrossRef]
- Shrestha, S.; Sharma, G.; Burgos, N.R.; Tseng, T.-M. Competitive ability of weedy rice: Toward breeding weed-suppressive rice cultivars. J. Crop. Improv. 2020, 34, 455–469. [Google Scholar] [CrossRef]
- Khanh, T.D.; Chung, M.I.; Xuan, T.D.; Tawata, S. The Exploitation of Crop Allelopathy in Sustainable Agricultural Production. J. Agron. Crop. Sci. 2005, 191, 172–184. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, M.; Li, Y.; Shui, J.; Zhou, Y. Allelopathy of rice (Oryza sativa L.) root exudates and its relations with Orobanche cumana Wallr. and Orobanche minor Sm. germination. J. Plant Interact. 2014, 9, 722–730. [Google Scholar] [CrossRef]
- Shrestha, S.; Sharma, G.; Burgos, N.R.; Tseng, T.-M. Response of weedy rice (Oryza spp.) germplasm from Arkansas to glyphosate, glufosinate, and flumioxazin. Weed Sci. 2019, 67, 303–310. [Google Scholar] [CrossRef]
- Tseng, T.M. Genetic Diversity of Seed Dormancy and Molecular Evolution of Weedy Red Rice. Ph.D. Dissertation, University of Arkansas, Fayetteville, AR, USA, 2013. [Google Scholar]
- Doyle, J.J. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. Bot. Soc. Am. 1987, 19, 11–15. [Google Scholar]
- McCouch, S.R.; Teytelman, L.; Xu, Y.; Lobos, K.B.; Clare, K.; Walton, M.; Fu, B.; Maghirang, R.; Li, Z.; Xing, Y.; et al. Development and Mapping of 2240 New SSR Markers for Rice (Oryza sativa L.). DNA Res. 2002, 9, 199–207. [Google Scholar] [CrossRef]
- Yang, G.P.; Maroof, M.S.; Xu, C.G.; Zhang, Q.; Biyashev, R.M. Comparative analysis of microsatellite DNA polymorphism in landraces and cultivars of rice. Mol. Genet. Genom. 1994, 245, 187–194. [Google Scholar] [CrossRef]
- Buntjier, J.B. Cross Checker; Wageningen University: Wageningen, The Netherlands, 1999. [Google Scholar]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Londo, J.P.; Schaal, B.A. Origins and population genetics of weedy red rice in the USA. Mol. Ecol. 2007, 16, 4523–4535. [Google Scholar] [CrossRef] [PubMed]
- Redona ED CL163: A New High-Amylose Content Clearfield Rice Variety. Available online: http://drec.msstate.edu/sites/default/files/cl163%20a%20new%20high-amylose%20content%20clearfield-rice%20variety.pdf (accessed on 8 September 2018).
- Sun, C.Q.; Wang, X.K.; Li, Z.C.; Yoshimura, A.; Iwata, N. Comparison of the genetic diversity of common wild rice (Oryza rufipogon Griff.) and cultivated rice (O. sativa L.) using RFLP markers. Theor. Appl. Genet. 2001, 102, 157–162. [Google Scholar] [CrossRef]
- Lapitan, V.C.; Brar, D.S.; Abe, T.; Redoña, E.D. Assessment of Genetic Diversity of Philippine Rice Cultivars Carrying Good Quality Traits using SSR Markers. Breed. Sci. 2007, 57, 263–270. [Google Scholar] [CrossRef] [Green Version]
- Xia, H.-B.; Wang, W.; Xia, H.; Zhao, W.; Lu, B.-R. Conspecific Crop-Weed Introgression Influences Evolution of Weedy Rice (Oryza sativa f. spontanea) across a Geographical Range. PLoS ONE 2011, 6, e16189. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.J.; Lee, D.S.; Song, Z.P.; Suh, H.S.; LU, B.R. Gene flow from cultivated rice (Oryza sativa) to its weedy and wild relatives. Ann. Bot. 2004, 93, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Tseng, T.M.; Burgos, N.R.; Shivrain, V.K.; Alcober, E.A.; Mauromoustakos, A. Inter-and intrapopulation variation in dormancy of Oryza sativa (weedy red rice) and allelic variation in dormancy-linked loci. Weed Res. 2013, 53, 440–451. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sl. No. | Marker | Observed Alleles | Nei’s Gene Diversity | Shannon’s Index |
|---|---|---|---|---|
| 1 | RM495 | 3 | 0.3098 | 0.5837 |
| 2 | RM283 | 3 | 0.6431 | 1.0647 |
| 3 | RM237 | 3 | 0.4464 | 0.7578 |
| 4 | RM431 | 3 | 0.6464 | 1.0660 |
| 5 | RM154 | 2 | 0.4989 | 0.6921 |
| 6 | RM452 | 2 | 0.1896 | 0.3382 |
| 7 | OSR13 | 2 | 0.1896 | 0.3382 |
| 8 | RM338 | 3 | 0.6561 | 1.0820 |
| 9 | RM514 | 3 | 0.2690 | 0.4828 |
| 10 | RM124 | 3 | 0.5059 | 0.7461 |
| 11 | RM507 | 3 | 0.6358 | 1.0537 |
| 12 | RM413 | 3 | 0.1714 | 0.3666 |
| 13 | RM161 | 3 | 0.4464 | 0.7578 |
| 14 | RM133 | 3 | 0.6358 | 1.0537 |
| 15 | RM162 | 3 | 0.1470 | 0.3267 |
| 16 | RM125 | 3 | 0.4464 | 0.7578 |
| 17 | RM455 | 3 | 0.6445 | 1.0627 |
| 18 | RM118 | 3 | 0.1470 | 0.3267 |
| 19 | RM408 | 3 | 0.6259 | 1.0299 |
| 20 | RM152 | 3 | 0.4579 | 0.7810 |
| 21 | RM44 | 3 | 0.3098 | 0.5837 |
| 21 | RM284 | 3 | 0.3527 | 0.6383 |
| 23 | RM433 | 3 | 0.6358 | 1.0537 |
| 24 | RM447 | 3 | 0.3250 | 0.6035 |
| 25 | RM316 | 3 | 0.5240 | 0.8829 |
| 26 | RM215 | 3 | 0.6445 | 1.0627 |
| 27 | RM271 | 3 | 0.3415 | 0.6331 |
| 28 | RM484 | 3 | 0.5240 | 0.8829 |
| 29 | RM536 | 3 | 0.6445 | 1.0627 |
| 30 | RM277 | 3 | 0.3415 | 0.6331 |
| Mean | 2.9 | 0.44 | 0.75 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shrestha, S.; Sharma, G.; Stallworth, S.; Redona, E.D.; Tseng, T.M. Exploring the Genetic Diversity among Weedy Rice Accessions Differing in Herbicide Tolerance and Allelopathic Potential. Diversity 2022, 14, 44. https://0-doi-org.brum.beds.ac.uk/10.3390/d14010044
Shrestha S, Sharma G, Stallworth S, Redona ED, Tseng TM. Exploring the Genetic Diversity among Weedy Rice Accessions Differing in Herbicide Tolerance and Allelopathic Potential. Diversity. 2022; 14(1):44. https://0-doi-org.brum.beds.ac.uk/10.3390/d14010044
Chicago/Turabian StyleShrestha, Swati, Gourav Sharma, Shandrea Stallworth, Edilberto D. Redona, and Te Ming Tseng. 2022. "Exploring the Genetic Diversity among Weedy Rice Accessions Differing in Herbicide Tolerance and Allelopathic Potential" Diversity 14, no. 1: 44. https://0-doi-org.brum.beds.ac.uk/10.3390/d14010044