
The Enigmatic Avian Oogenus Psammornis: A Review of Stratigraphic Evidence


1
Centre National de la Recherche Scientifique (UMR 8538), Laboratoire de Géologie de l’Ecole Normale Supérieure, PSL Research University, 24 rue Lhomond, CEDEX 05, 75231 Paris, France
2
Palaeontological Research and Education Centre, Maha Sarakham University, Maha Sarakham 44150, Thailand
Diversity 2022, 14(2), 123; https://0-doi-org.brum.beds.ac.uk/10.3390/d14020123
Submission received: 31 December 2021
/
Revised: 31 January 2022
/
Accepted: 2 February 2022
/
Published: 8 February 2022
(This article belongs to the Special Issue Evolution and Palaeobiology of Flightless Birds)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Psammornis rothschildi is an avian taxon established by Andrews in 1911 on the basis of eggshell fragments surface-collected near the city of Touggourt, in the north-eastern part of the Algerian Sahara. Since the initial discovery, a number of Psammornis specimens have been reported from various localities in North Africa (Algeria, Tunisia, Libya, Mauritania) and the Middle East (Saudi Arabia, Iran). Most of the finds lack a stratigraphic context, which has resulted in considerable confusion about the geological age of Psammornis, with attributions ranging from the Eocene to the Holocene. A review of the available evidence shows that only two groups of localities provide reasonably reliable stratigraphic evidence: the Segui Formation of SW Tunisia, apparently of latest Miocene age, and the Aguerguerian (Middle Pleistocene) of NW Mauritania. This suggests a fairly long time range for Psammornis. Psammornis eggs are, in all likelihood, those of giant ostriches, although the lack of associated skeletal material makes it difficult to interpret the eggshell fragments in evolutionary terms. However, the oological record suggests that giant ostriches have been present in Africa since the late Miocene, which leads to the reconsideration of some hypotheses about the palaeobiogeographical history of the Struthionidae. The lack of Psammornis eggs transformed by humans suggests that this giant ostrich did not survive until Epipalaeolthic or Neolithic times.
1. Introduction
The genus Psammornis was established in 1911 by Andrews [1], with P. rothschildi as the type species, on the basis of eggshell fragments collected in southern Algeria in the course of one of Lord Rothschild’s expeditions to North Africa [2]. Since then, more eggshell material collected at a number of localities in Africa and the Middle East has been attributed to Psammornis; this rather enigmatic taxon has been the subject of much speculation, both because it is represented solely by fragmentary eggshell material not clearly associated with any skeletal remains, and because the stratigraphic provenance of the specimens is often very uncertain due to many of them being surface-collected rather than found in situ in sedimentary formations. Despite these uncertainties, Psammmornis is still often mentioned in works on ostrich evolution [3] as well as in papers on fossilisation [4], with different geological age estimates—Pleistocene, according to Mikhailov and Zelenkov [3], and Holocene, according to Wiemann et al. [4].
The systematic position of Psammornis has been the subject of much debate [5]. Although aepyornithid affinities have been suggested [6,7] and Dughi and Sirugue [8,9] thought that the microstructure of Psammornis eggshells was closer to that of rheas and moas, it is now widely accepted that the Psammornis eggs are probably those of giant ostriches [1,3,5,10]; according to Mikhailov and Zelenkov [3], they belong to their “non-specialised type S”, and are similar to both the thin-shelled eggs of the modern subspecies Struthio camelus camelus and the thick-shelled eggs of the Pliocene S. chersonensis from Europe. However, the purpose of the present paper is not to discuss systematic issues or morphological and microstructural characteristics, but to review whatever solid evidence may be available about the stratigraphic provenance of Psammornis eggshells, on the basis of a survey of the available literature; this includes papers, mainly in French, which have been overlooked by many authors dealing with the question. Bearing in mind that the geological age of the type material of Psammornis rothschildi can rightly be considered as uncertain [11], widely different opinions have been expressed about the geological age of Psammornis specimens from various localities; this, in turn, has resulted in divergent interpretations of ratite evolutionary history in Africa and other continents.
2. The Discovery of Psammornis and the Beginning of the Stratigraphic Conundrum
In 1909, during a visit to southern Algeria, Lord Rothschild and Ernst Hartert explored the area around the city of Touggourt [2], in the north-eastern part of the Algerian Sahara. They found abundant fragments of ostrich eggshell on the ground in various places. While picking up some of them about 22 miles east of Touggourt, Hartert found “three pieces of a very much thicker egg-shell of a much browner colour” ([2], p. 550). Rothschild immediately thought that they must belong to “an extinct large Struthionid bird”. The fragments were handed over for study to C.W. Andrews, a palaeontologist at the British Museum (Natural History), who described them as Psammornis rothschildi [1]. Andrews gave a detailed, but not illustrated, description of the microstructure and surface features of the eggshell, noting its considerable thickness (3.2 to 3.4 mm), much greater than that of eggs of the living ostrich (up to 2.10 mm) and second only to that of the eggshell of Aepyornis titan, from Madagascar. His conclusion was that the fragments were evidence of a “hitherto unknown bird which laid an egg considerably larger than the largest produced by any modern ostrich” and that, although there were some similarities with Aepyornis, “with Struthio the relationship was probably very close” ([1], p. 172).
Besides the lack of associated skeletal material, which made precise identification difficult, Andrews also noted uncertainties about the geological age of the specimens. Although they had been surface-collected and had been abraded by drifting sand, they were highly mineralised and had been found in the vicinity of a well. Andrews suggested that they might have been brought up from a considerable depth when digging the well. This gave no clue as to the exact antiquity of the eggshell fragments. Hartert [12], discussing Psammornis-like eggshell fragments found in the western Algerian Sahara, questioned Andrews’ suggestion, pointing out that the Psammornis fragments often had the same preservation as Struthio camelus eggshell fragments and, everywhere, were found together with the latter.
Since the initial discovery in Algeria, many eggshell fragments (but no complete eggs) attributed to Psammornis have been reported from various parts of North Africa (including Algeria, Tunisia and Mauritania; Figure 1) and the Middle East (Saudi Arabia, Iran), but many of them were surface-collected and lack details about their stratigraphic origin. In addition, some of these purported Psammornis specimens appear to be significantly different from the fragments described by Andrews [1], and their attribution to the genus seems dubious.
3. An Eocene Age for Psammornis?
Rothschild [13] (1911) listed Psammornis among what he called the Heterornithes, a loosely defined group of large birds which he considered as “fore-runners or ancestral forms of the Ratite section of the Palaeognathae” ([13], p. 148). This highly heterogenous group contained birds as different as gastornithids, phorusrhacoids and even the Jurassic Laopteryx (now considered a pterosaur). Psammornis was listed together with Eremopezus eocaneus as an Eocene representative of the Heterornithes from North Africa. Eremopezus eocaneus was described by Andrews [14] on the basis of the distal end of a tibiotarsus from the Upper Eocene Jebel Qatrani Formation of the Fayum, in Egypt. It was long believed to be an early ratite, but Rasmussen et al. [15] considered it a representative of an endemic group of large African birds.
Why Rothschild chose to associate Psammornis and Eremopezus is obscure. In his description of Psammornis, Andrews [1] mentioned Eremopezus, mainly to remark that it was only the size of a small ostrich (and, therefore, unlikely to have produced eggs the size of Psammonis eggs). Moreover, there was no solid reason to believe that the type material of Psammornis from Algeria came from an Eocene deposit, despite the fact that Andrews had suggested that the fragments might have been brought up to the surface from a considerable depth by a well. However, as mentioned above, as early as 1913, Hartert [12] had doubted that the Algerian Psammornis fragments could have been brought up to the surface by a well, and in 1918, Geyr von Schweppenburg [16] had noted that it was geologically unlikely that they were Eocene in age. The hypothesis that the Algerian specimens may have originated from Eocene deposits was dismissed on geological evidence by Dughi and Sirugue [8,9]. Sauer [5] reviewed, at some length, the question of the possible relationships between Psammornis and Eremopezus and concluded that there was no reason to believe they were related.
Nonetheless, Rothschild’s idea of an Eocene age for Psammornis was taken up first by Abel [17] and then by Lambrecht [6], who placed it together with Eremopezus in the family Eremopezinae within the Aepyornithformes. In 1933, Lambrecht [7] still placed Psammornis close to Eremopezus but as a genus incertae sedis, and gave it an Eocene age with a question mark. More recently, Piveteau [18] noted that Psammornis had been discovered in the Eocene of Touggourt. Brodkorb [19] still considered Psammornis as a possible synonym of Eremopezus, and also attributed it to the Eocene, with a question mark. Dementiev [20] used question marks concerning the placement of Psammornis among the “Eremopezidae” and its attribution to the Eocene. As late as 1965, Swinton [21] wrote that Psammornis had been found in the Eocene of southern Algeria. Obviously, Rothschild’s ill-founded assertion that Psammornis was an Eocene taxon has had long-lasting consequences. There is, in fact, no convincing stratigraphic evidence anywhere suggesting that Psammornis-type eggshells have been found in Eocene deposits.
4. Psammornis Outside Africa
4.1. Saudi Arabia
In 1933, Lowe [22] attributed to Psammornis a thick eggshell fragment collected by the explorer and secret agent St John Philby at Shuqqat al Khalfat in Saudi Arabia. He compared it with Andrews’ original material from Algeria and found close similarities in thickness and microstructure. The specimen had been surface-collected and lacked accurate information about its geological origin.
In this connection it should be noted that Bibi et al. [23] attributed eggshell fragments from the Upper Miocene Baynunah Formation of the United Arab Emirates not to Psammornis, but to Diamantornis, previously described from Namibia and Kenya. The pore complexes of the specimens from the United Arab Emirates are clearly different from what has been described in Psammornis, and Philby’s specimen from Saudi Arabia, whatever its age, may indicate a bird different from that from the Baynunah Formation.
4.2. Iran
Dughi and Sirugue [8] reported on eggshell fragments collected at several localities in the Lut desert of eastern Iran by the geographer Jean Dresch and the naturalist Théodore Monod in 1969 and 1970. Although these fragments were significantly thinner (1.8 to 2.3 mm) than the Psammornis material described by Andrews, Dughi and Sirugue attributed them to Psammornis on microstructural evidence, considering that they belonged to an early, less advanced form than that from North Africa. However, the Iranian specimens seem to have been collected in a sand dune environment without a well-defined stratigraphic context. Caution should be urged when trying to use the putative Psammornis specimens from Iran for evolutionary interpretations. In connection with these specimens, Dughi and Sirugue mentioned the large ostrich eggs known from the Neogene and Quaternary periods of China [3,24,25,26], which may indeed be more relevant than the African Psammornis.
5. African Records of Psammornis of Doubtful Identification or Stratigraphic Position
5.1. Algeria
In addition to the type material of Psammornis rothschildi from the Touggourt region in the north-eastern part of the Algerian Sahara, Rothschild and Hartert [2] reported that in 1911, Hilgert had surface-collected a number of large eggshell fragments, rather different from the type of P. rothschildi, near Biskra, north of Touggourt. Schönwetter [27] discussed the fragments found by Hilgert and found them similar to the type material from Touggourt. He also mentioned eggshell fragments collected at Ouargla and El Golea (SW of Touggourt) by Hilgert and Hartert and at Temassin (S of Touggourt) by Fromholz and found them different from the type material. In 1960, Schönwetter [28] came back to the topic of Psammornis and mentioned additional material from the Iguidi region, in the southern part of the Algerian Sahara, collected by Colonel Le Pivain.
Le Pivain also collected thick eggshell fragments at various points during a trip across the western Algerian Sahara in 1930; they were referred to Psammornis by Heim de Balsac [29].
Dughi and Sirugue [8] mentioned additional Psammornis eggshell fragments collected at various localities in the north-eastern Algerian Sahara (Souf region). No indications about the geological context were provided.
Apparently, all of the above-mentioned Algerian specimens were surface-collected and do not provide any reliable stratigraphic evidence about the age of Psammornis.
5.2. Libya
Psammornis libycus was described by Moltoni [30] on the basis of brownish-red eggshell fragments found in the dunes south of Giarabub (al-Jaghbub), a town in the eastern Libyan desert. No stratigraphic information was available. Moltoni established a new species because the fragments were significantly thinner (2.1 mm) than those of Psammornis rothschildi eggs. Schönwetter [28] concluded that these fragments had nothing to do with Psammornis and were reminiscent of specimens from Algeria possibly belonging to Struthio camelus. Sauer [5] described the type specimens of Psammornis libycus in detail and found similarities with Struthio camelus, wondering whether they could be the earliest S. camelus (or a link between Psammornis and Struthio). However, nothing is known about the geological age of P. libycus. Although this species was mentioned by some authors (e.g., [7,19]), there seems to be no reason to attribute it to Psammornis. The fragments described by Moltoni are more likely to belong to Struthio.
Psammornis has also been reported from the important early Miocene Jebel Zelten vertebrate locality in north-central Libya. However, this record is not based on oological evidence, and therefore, is completely unreliable. In a preliminary report, Arambourg and Magnier [31] initially mentioned a giant bird belonging to the Aepyornithidae, without specifying what kind of material this identification was based on. In 1961, this was clarified by Arambourg [32], who briefly attributed to an aepyornithid an incomplete large tibia (more properly, a tibiotarsus) lacking both ends, which in terms of size, shape and proportions was supposed to be reminiscent of Aepyornis. Arambourg then mentioned that large eggshell fragments from “Mio-Pliocene” deposits in the northern Sahara had been described as Psammornis rothschildi, but refrained from clearly attributing the Jebel Zelten giant bird to that genus. In 1967, however, in a list of the Jebel Zelten fauna, he mentioned: “Aepyornithide [sic] (Psammornis)”, without specifying what kind of material this was based on [33]. This mention obviously caused some confusion among subsequent authors who appear to have been unaware of Arambourg’s 1961 paper, in which it is clearly stated that the evidence for the presence of a giant bird at Jebel Zelten is based on a fossil bone. Curiously enough, Savage and Hamilton [34], who cite Arambourg’s 1961 paper, list ‘’? Eremopezus’’ among the fossil vertebrates from Jebel Zelten; this is probably a consequence of the above-mentioned confusion, initiated by Lord Rothschild, between Eremopezus and Psammornis. Vickers-Rich [35], not being aware of Arambourg’s 1961 paper, was not sure on what material the mention of Psammornis was based. Mlíkovsky [36], who apparently was not aware of Arambourg’s 1961 paper either, misquoted Arambourg and Magnier (his quote clearly refers to Arambourg’s 1967 paper, not cited in his list of references) and noted that the material on which a giant bird was identified at Jebel Zelten was “unknown” but probably consisted of eggshell fragments, since it was attributed to Psammornis. This, of course, is erroneous, since Arambourg’s mention of a giant bird was based on a tibiotarsus.
To sum up, there is no evidence of Psammornis-type eggshell material from Jebel Zelten. The large tibiotarsus from Jebel Zelten mentioned by Arambourg [32] was never described in detail or illustrated. It is of potentially considerable importance for our understanding of the evolution of giant birds in Africa, but it cannot provide much reliable evidence concerning the question of the geological age of Psammornis, and cannot be used as evidence for the presence of this egg-based taxon in the early Miocene of Libya.
6. Psammornis from Tunisia: A Late Miocene Record?
As early as 1911, Rothschild and Hartert [2] reported that the German naturalists Erlanger and Hilgert had found many fragments of large eggshells in the South Tunisian desert. Bédé [37] noted that he had found eggshell fragments of a brownish colour and thicker than those of the “ordinary ostrich” at Mezzouna, 100 km SW of the city of Sfax; he thought they belonged to Psammornis.
A stratigraphically more significant discovery of Psammornis specimens was reported by Choumowitch (not ‘Choumwitch’ as erroneously printed in his paper) in 1951 [38]. The eggshell fragments mostly came from outcrops SW of the city of Moularès, in south-western Tunisia. They were found in abundance in red beds deeply dissected by erosion (Figure 2) in an area known as Chebket Safra (“yellow network”). The brown-coloured eggshell fragments were attributed to Psammornis because of their thickness (3 mm). Choumowitch also noted that they were less convex than ostrich eggshells, thus indicating larger eggs.
As emphasised by Choumowitch, contrary to previous Psammornis finds, the eggshell fragments from Chebket Safra came from well-dated sediments. They were found in palaeosols containing calcareous concretions and fossil helicid gastropods, notably Leucochroa tissoti, considered as indicating a Pontian age. Choumowitch’s investigations showed that the eggshell fragments did not come from Quaternary pebble deposits topping the hills. Digging into the red clays to a depth of one metre below the surface, he found an accumulation of eggshell fragments (weighing altogether more than 11 kg) arranged into small groups, each of which apparently corresponded to an egg. The whole accumulation was interpreted as a nest.
According to the geological map, the egg-bearing red beds were Pontian in age, overlying fluvial sands. Choumowitch noted that similar Psammornis eggshell fragments occurred at other localities in the area. Other records from Tunisia (Metlaoui, Gabès) were listed by Dughi and Sirugue [8] on the basis of information provided by Gobert.
More details about the geology of the localities were sent to Dughi and Sirugue by Gobert and published by them [8,9]. He mentioned that within the Pontian red marls, the eggshell fragments were concentrated in a well-defined zone about 50 cm in thickness, containing abundant helicid gastropods and calcareous concretions, which were interpreted as having formed around grass roots. The egg-bearing deposit was considered a paleosol formed in a dry steppe environment.
The observations made by Choumowitch and Gobert thus indicate that at Chebket Safra, the Psammornis eggshells were found in situ, and not reworked.
Dughi and Sirugue [8,9] gave detailed descriptions of the morphology and microstructure of the eggshells from Chebket Safra (Figure 3A). They had no hesitation about attributing the Tunisian specimens to Psammornis rothschildi. However, they noted that Andrews’ description of the pores and canals of the Psammornis eggshell had to be significantly modified. This raises the question of whether the Tunisian specimens can really be attributed to Psammornis rothschildi. The comparison between Andrews’ material from Algeria and the Tunisian fragments is made difficult by the fact that the former have suffered aeolian abrasion so that their surface features are somewhat obscured [1]. On the basis of the type material from Touggourt, Andrews [1], and especially Sauer [5], emphasised the similarities between the eggshells of Struthio and Psammornis. Dughi and Sirugue [8], mainly on the basis of the Tunisian specimens, came to a somewhat different conclusion, viz. that Psammornis was more closely related to Rheiformes and Dinornithiformes. In a later paper [9], they found similarities with Aepyornithiformes, Rheiformes, Casuariformes and Dinornithiformes, rather than with ostriches. A detailed comparison between eggshell fragments from Chebket Safra and the type material of Psammornis rothschildi from Touggourt would be useful to check whether they can really be attributed to the same ootaxon.
By contrast with the various specimens from Algeria, which lack a stratigraphic context, the eggshells from Chebket Safra are of considerable importance concerning the geological age of Psammornis. However, the age attributions proposed at the time of the discovery need to be discussed in light of current knowledge.
On the basis of previous work in the area, notably that of Solignac [39], Choumowitch attributed the fossil-bearing red beds to the Pontian. The Pontian is a local stage for the Peri-Tethyan regions around the Black Sea corresponding to deposits close to the Miocene–Pliocene boundary. According to Rybkina and Rostovtseva [40], the Pontian of the Black Sea area can be correlated with the Messinian Salinity Crisis of the upper part of the Messinian stage (latest Miocene). However, correlations of the so-called Pontian of south-western Tunisia with the standard stratigraphic scale is not obvious. Moreover, the red beds of the Chebket Safra lack useful biostratigraphic markers. The abundant helicid gastropods they contain, in particular Helix (Leucochroa) tissoti, were once considered valuable stratigraphic indicators for a Pontian age in North Africa [38,41]. However, this has been doubted [42].
The lithostratigraphic characteristics and the abundant helicid gastropods lead to the consideration that the Psammornis-bearing red beds belong to the Segui Formation. This is in agreement with the Metlaoui sheet of the geological map of Tunisia at 1/100,000 [43], which shows a great development of this formation (attributed to the Pliocene) SW of Moularès, in the area of the Chebket Safra. Although Robinson and Wiman [42] described the Segui Formation as a temporal enigma, its stratigraphic position is relatively clear. As early as 1910, Roux and Douvillé [41] placed the red clays containing Helix tissoti in the upper part of the Miocene series and observed that they overlie white and yellow sands yielding mammal remains. This sandy formation is now called the Beglia Formation and two fossiliferous levels within it have yielded a rich vertebrate assemblage indicative of an age close to the middle–late Miocene boundary ([44] and references therein). The Beglia Formation has yielded skeletal remains of an ostrich [45], Struthio sp., which is not larger than the living Struthio camelus, and therefore, unlikely to have produced the very large Psammornis eggs. The Segui Formation, which in the Moularès region directly overlies the Beglia Formation, was considered Messinian to Pliocene [46], or Mio-Pliocene to Villafranchian [47]. In her reviews of Miocene formations in Tunisia, Mannai-Tayech [48,49] suggested a late Tortonian–early Messinian age for the Segui Formation. Mannai-Tayech’s lithological description of the Segui Formation suggests that the level with Psammornis eggshells is in the upper part of the formation. It is worth noting that Robinson and Wiman [42] mentioned the occurrence of gypsum deposits in the Segui Formation as possible evidence for contemporaneity with the Messinian salinity crisis.
Although some uncertainties remain about the age of the upper boundary of the Segui Formation, the above-mentioned evidence strongly suggests a late Miocene, possibly Messinian age, for the Psammornis eggshells from the Chebket Safra.
It is worth noting that in 1952, Arambourg [50] mentioned that, according to Gobert, Psammornis eggshell fragments were relatively frequent in continental deposits, probably Pontian in age, in southern Tunisia. However, he did not cite Choumowitch’s paper and seems to have been unaware of it (or possibly Choumowitch’s paper was published after Arambourg wrote his paper). Vickers-Rich [10] noted Arambourg’s mention of Psammornis in southern Tunisia, but she was unaware of Choumowitch’s and Dughi and Sirugue’s papers, so she could not appreciate the real significance of the Tunisian finds.
7. Psammornis from Mauritania: A Pleistocene Record
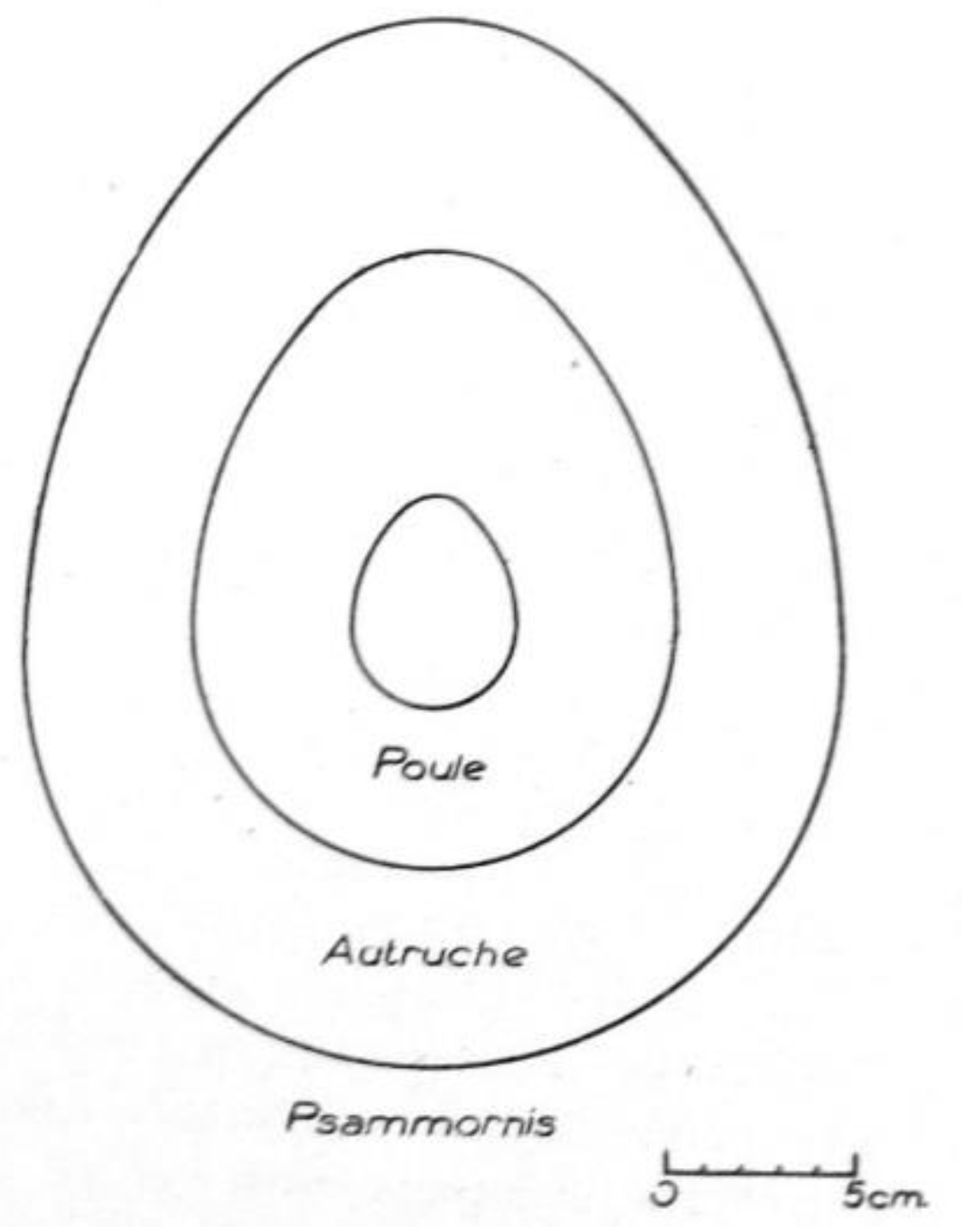
In 1939, Monod [51] was the first to report the occurrence of Psammornis on the Atlantic coast of Mauritania in a semi-popular short paper on eggshell fragments he had discovered at the “root” of the Cap Blanc (Râs Nouâdibhou) peninsula, in the northwestern corner of the country. The specimens were communicated to Schönwetter, who noted that they were 3 to 3,4 mm thick, and provided estimates for the dimensions of the complete shell (28 × 21 cm) and the weight of the egg (7 kg)—all of these figures being much larger than those for the living ostrich (as expressed by a comparative drawing). Monod gave no information about the geological provenance of the eggshell fragments but jokingly mentioned the omelets and water containers the Neolithic people of the Sahara could have made with such eggs, showing that he thought that they were of a comparatively recent date. In a more formal paper, including a better comparative drawing (Figure 4), Monod [52] quoted Schönwetter again; the German oologist had no doubt that the fragments belonged to Psammornis rothschildi, the thickness being the same, although he remarked that the disposition of the pores was not exactly similar.
The geological provenance of the Mauritanian Psammornis was made clear in 1971, when Tessier et al. [53] showed that the eggshells came from Aguerguerian deposits (the Aguerguerian is a local stage of the Pleistocene). They were found in calcareous and clayey sandstones, which, as in Tunisia, also yielded helicid gastropods. Seven localities (see map in [9]) were mentioned where Psammornis eggshell fragments were found in situ, with sharp, unworn edges (unlike the surface-collected specimens showing clear signs of aeolian erosion). Voisin [54] described the surface features (Figure 3B) and microstructure of eggshells collected by Hébrard at three Mauritanian localities, noting that they were extremely abundant at one of them, which seemed to correspond to a nesting site. She found a complete correspondence with Psammornis rothschildi and noted great similarities with ostrich eggs in the surface features of the shell. No association with human artefacts was noted. Dughi and Sirugue [9] also gave a description of Mauritanian eggshells, which they attributed to Psammornis, noting a certain variability in the eggshell surface.
Figure 3.
Pores on the outer surface of Psammornis eggshell fragments from Tunisia, (A) modified after Dughi and Sirugue [8]) and Mauritanaia (B), modified after Voisin [54].

Figure 4.
Outline reconstruction of a Psammornis egg by Monod on the basis of Schönwetter’s size estimates, with outlines of the eggs of an ostich and a hen for comparison. After Monod [51].
Figure 4.
Outline reconstruction of a Psammornis egg by Monod on the basis of Schönwetter’s size estimates, with outlines of the eggs of an ostich and a hen for comparison. After Monod [51].

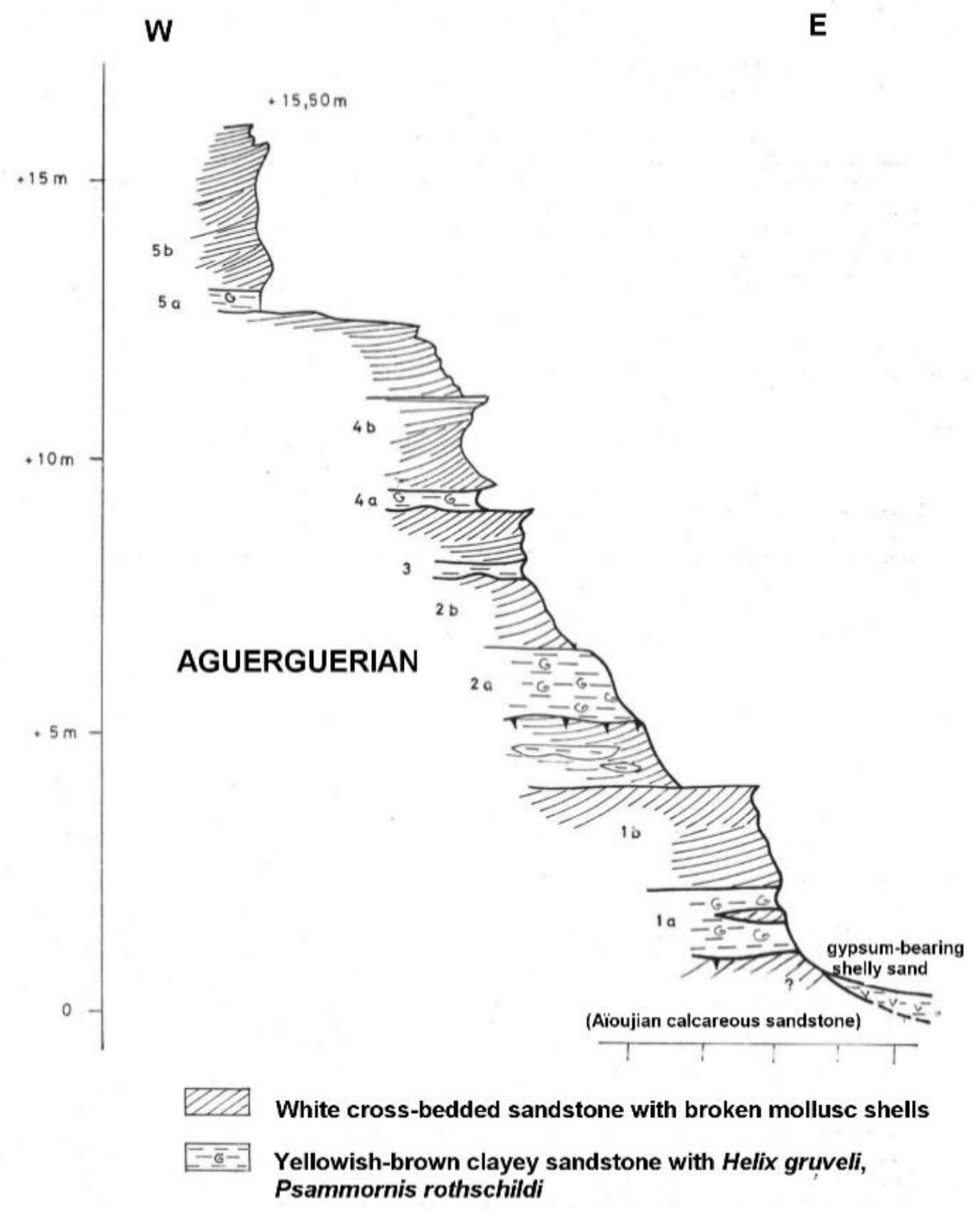
The specimens from Mauritania, like those from the Chebket Safra, are of especial importance because they were found in situ in a stratigraphic context, and therefore, provide evidence about the geological age of Psammornis. As noted by Tessier et al. [53], Ortlieb [55] and Hébrard [56], they occur in the Aguerguer sandstones, attributed to the Aguerguerian stage. A section of one of the localities was published by Hébrard [56] (Figure 5). As noted by Hébrard et al., [57], the Aguerguerian is, in fact, a regressive facies of the Middle Pleistocene Aioujian stage, and the clayey sandstones containing helicid gastropods and Psammornis correspond to paleosols.
Although the Aguerguerian has long been considered as Middle Pleistocene, its exact age remained uncertain until radiometric dates became available. C14 ages of more than 39,900 years were obtained for Psammornis eggshell fragments [55], but this was a minimum age. Giresse et al. [58] published U/Th dates from fossil mollusc shells from geological formations corresponding to several Pleistocene local stages along the Mauritanian coast. No ages were provided for the Aguerguerian; however, ages ranging from 241,000 to 258,000 years were obtained for the underlying Aioujian, and of up to 111,000 years for the overlying Inchirian. Since the Aguerguerian is considered a facies of the Aioujian, an age of about 200,000 years seems likely (Middle Pleistocene, Chibanian). As noted by Hébrard [56], Psammornis does not occur in the Inchirian deposits.
It is worth noting that engraved ostrich eggshells and ostrich eggshell beads are found in some abundance at prehistoric (Epipalaeolithic and Neolithic) sites in Mauritania [59]. Vernet et al. [59] mentioned that at the Cansado prehistoric site, many Psammornis eggshell fragments were collected in addition to fragments of Struthio camelus eggs; they noted that Psammornis had disappeared well before the occupation of the site by humans. More generally, there does not seem to be any record of the use of Psammornis eggs (for ornamentation or other purposes) by prehistoric humans, in Mauritania (R. Vernet, pers. com.) or elsewhere. Nevertheless, the presence of Psammornis eggs in the Middle Pleistocene Aguerguerian deposits of coastal Mauritania implies that this giant bird must have been contemporaneous with early humans, even though no early Palaeolithic industries have been found in Aïoujian and Aguerguerian sediments [56,60]. This absence may possibly be linked to unfavourable climatic conditions, since the Aguerguerian climate appears to show an evolution towards greater aridity [56,61].
8. The Stratigraphic Record of Psammornis and its Implications
(1) Most reported occurrences of Psammornis lack a reliable stratigraphic control. This applies to all records from the Algerian Sahara, including the type material from Touggourt. They correspond to surface finds, often in a sand dune environment, and the geological origin of the fragments could not be ascertained. This also applies to finds from Saudi Arabia (which may belong to Psammornis) and Iran (unlikely to be Psammornis in view of the thinness of the shell).
(2) The attribution to Psammornis of various eggshell fragments is dubious. This applies mainly to fragments which are significantly thinner than the original specimens from Touggourt and do not seem to be much thicker than normal Struthio camelus eggshells. A case in point is the material of “Psammornis libycus” from Libya which, in all likelihood, can be attributed to Struthio. As pointed out by Dughi and Sirugue [8], the unusual thickness of Psammornis eggshells was one of the main defining characteristics used by Andrews [1] to establish the taxon Psammornis rothschildi. However, among thick eggshells, differences in the pore system have been noted. This is reflected by the divergences between the interpretations of Andrews [1], Sauer [5] and Voisin [54], who find great similarities between Psammornis and Struthio, and that of Dughi and Sirugue [8,9], who conclude that Psammornis is related to Rheiformes, Casuariformes, Aepyornithiformes and Dinornithiformes. A revision of the material from Chebket Safra, on which Dughi and Sirugue’s conclusions were largely based, would help to clarify the question.
(3) Only two groups of localities seem to provide reliable stratigraphic evidence concerning the geological age of Psammornis:
- −
- The Segui Formation of south-western Tunisia, where Psammornis ‘nests’ have been discovered in situ in red sandy clays apparently containing paleosols. Although some uncertainties remain, the likeliest age for the Segui Formation seems to be latest Miocene (about 6 My ago);
- −
- The Aguerguerian of the Mauritanian coast, especially in its northern part, where what are apparently Psammornis nesting sites have been discovered in situ in sandstone formations showing evidence of paleosols. The Aguerguerian is placed in the Middle Pleistocene and may be about 200,000 years old.
These two relatively well dated Psammornis records are rather far apart in time, implying that, if the specimens from Chebket Safra do belong to this oogenus, Psammornis was present in North Africa over a long time span, covering at least 6 million years. This in itself is not unlikely for a genus of giant bird. The genus Struthio has a record extending from the Early Miocene (S. coppensi) to the present. It seems less likely that the Miocene and Pleistocene specimens of Psammornis should belong to a single species (P. rothschildi). It should be remembered, however, that Psammornis is an ootaxon, the evolution of which probably cannot be followed over time with the same precision as that of taxa based on skeletal material. In fact, the distribution of Psammornis eggshells mainly shows that very large ratite birds were present in North Africa over a fairly long period of time, from the Late Miocene to the Middle Pleistocene. There is no consensus about the systematic position of these birds, although the idea that they may have been related to the Aepyornithiformes of Madagascar has never been well-supported. Dughi and Sirugue [8,9], on the basis of microstructural characteristics, suggested relationships with rheas and moas. However, following Sauer’s revision [5], most authors [3,10,11,62] seem to agree that Psammornis was closely related to Struthio—if not, in fact, a member of that genus—and this would lead to the possible consideration of Psammornis as a junior synonym of the oogenus Struthiolithus. Further research is needed to decide whether the specimens from Chebket Safra can, indeed, safely be referred to Psammornis.
It may be worth noting that both in Tunisia and in Mauritania the deposits that yield Psammornis eggshells apparently show signs of aridity. The giant ostrich may have been adapted to dry environments, and this may, in turn, explain its large size. Size increase in the Australian dromornithids has been interpreted as an adaptation to an increasingly arid environment with decreasing food resources [63,64], and a similar process has been proposed for some of the giant ostriches of Eurasia [65].
(4) When Psammornis became extinct remains uncertain. The Mauritanian record indicates that it survived until the Middle Pleistocene, roughly 200,000 years ago; this implies that it was contemporaneous with early humans, although evidence of interactions between hominins and these giant birds has never been reported. Attributions of Psammornis to the Holocene (e.g., [4]) are not supported by the available evidence. A point worth noting, as mentioned above, is that apparently no engraved or otherwise transformed (for instance for bead-making) Psammornis eggshells have been reported, whereas decorated, cut or perforated Struthio eggshells are very common at Epipalaeolithic and Neolithic sites in North Africa [66,67]. This strongly suggests that Psammornis was no longer in existence when these cultures flourished in the Late Pleistocene and Holocene.
(5) Although very few Psammornis localities are well dated, the available reliable ages are important in terms of the evolution of giant ratites in North Africa. It should of course be borne in mind that ootaxa are more difficult to interpret in terms of evolutionary history than skeletal remains, and that no reliable association between Psammornis-type eggshells and fossil bones has been reported. Large ostriches have been reported on the basis of skeletal material from North Africa. They include Struthio barbarus, from the ‘Villafranchian’ (early Pleistocene) of Algeria [68], which is not associated with egg remains. At Ahl al Oughlam (late Pliocene or earliest Pleistocene of Morocco), a few bones of a large ostrich have been erroneously [3] assigned to Struthio asiaticus by Moure-Chauviré and Geraads [69]; they are accompanied by eggshell fragments described as being different from Psammornis. As long as no clear association between skeletal remains and Psammornis eggs is reported, it will remain difficult to interpret the Psammornis record in evolutionary terms. However, the stratigraphic evidence yielded by the Tunisian and Mauritanian records leads us to question some assumptions. For instance, Mikhailov and Zelenkov [3] assumed that Psammornis rothschildi is late Early Pleistocene to Middle Pleistocene in age and suggested that the Psammornis lineage evolved in more northern territories (perhaps from Struthio chersonensis from eastern Europe) and then dispersed to North Africa. However, if they actually belong to Psammornis, the latest Miocene specimens from Tunisia are not in agreement with this hypothesis. They indicate the presence in North Africa of a giant ostrich at a much earlier date and may even suggest an African origin for the large ostriches of late Neogene Europe, although the direction of dispersal remains uncertain. The whole picture is made even more complicated by the struthionid record from southern and eastern Africa, consisting of both bone and egg remains (but apparently not including Psammornis-type eggs), which is beyond the scope of the present paper and has led to speculations about the evolutionary history of the ostriches [70].
9. Conclusions
Ever since the initial description by Andrews in 1911, the egg-based taxon Psammornis has been the subject of widely divergent interpretations in terms of both zoological affinities and stratigraphic distribution. There now seems to be a measure of consensus about the fact that the large thick-shelled Psammornis eggs were produced by giant ostriches, although the lack of association with skeletal material makes an interpretation in evolutionary terms difficult. The stratigraphic conundrum which began at the time of the initial description, when an Eocene age was suggested without any solid evidence, has proved even more difficult to resolve, because many of the Psammornis finds (even when doubtful records are eliminated), consisting of surface finds, lack any solid stratigraphic context. However, a review of the literature, including various important papers which, for some reason, have been ignored by most authors dealing with the topic, shows that eggs attributed to Psammornis have been discovered in situ in relatively well-dated formations at two geographically widely separated groups of localities, in south-western Tunisia and on the Mauritanian coast. Interestingly, these sites are also quite distinct in geological age: the Tunisian finds are apparently latest Miocene in age, whereas the Mauritanian occurrences are in Middle Pleistocene deposits. Whether the Tunisian and Mauritanian Psammornis really belong to a single taxon is a moot point, but the occurrence in North Africa of large eggs probably produced by giant ostriches at two periods widely distant in time may suggest a long-lasting lineage of large ratites in that region. This should be taken into consideration when trying to reconstruct ostrich evolutionary history. In particular, it seems difficult to accept that giant ostriches dispersed from Eurasia to Africa as late as the Pleistocene, since they appear to have been present on the African continent as early as the late Miocene. The large eggs from Tunisia may even suggest dispersal in the opposite direction, from Africa to Eurasia. A detailed revision of the Tunisian specimens would, therefore, be very welcome.
Funding
This research received no external funding.
Acknowledgments
Thanks to Jihed Dridi and Beya Mannaï-Tayech for the information about the geology of south-western Tunisia, to Robert Vernet for the information about Psammornis in Mauritania, and to the three anonymous reviewers for their helpful comments.
Conflicts of Interest
The author declares no conflict of interest.
References
- Andrews, C.W. Note on some fragments of the fossil egg-shell of a large struthious bird from southern Algeria, with some remarks on some pieces of the egg-shell of an ostrich from northern India. In Verhandlungen des V. Internationalen Ornithologen-Kongresses in Berlin 1911; Schalow, H., Ed.; Deutsche Ornithologische Gesellschaft: Berlin, Germany, 1911; pp. 169–174. [Google Scholar]
- Rothschild, W.; Hartert, E. Ornithological explorations in Algeria. Novit. Zool. 1911, 18, 456–550. [Google Scholar] [CrossRef]
- Mikhailov, K.E.; Zelenkov, N. The late Cenozoic history of the ostriches (Aves: Struthionidae), as revealed by fossil eggshell and bone remains. Earth Sci. Rev. 2020, 208, 103270. [Google Scholar] [CrossRef]
- Wiemann, J.; Fabbri, M.; Yang, T.R.; Stein, K.; Sander, P.M.; Norell, M.A.; Brigg, D.E.G. Fossilization transforms vertebrate hard tissue proteins into N-heterocyclic polymers. Nat. Com. 2018, 9, 1234567890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauer, E.G.F. Taxonomic evidence and evolutionary interpretation of Psammornis. Bonn. Zool. Beitr. 1969, 20, 290–310. [Google Scholar]
- Lambrecht, K. Fossilium Catalogus I Animalia; Backhuys Publishers: Leiden, The Netherlands, 2007. [Google Scholar]
- Lambrecht, K. Handbuch der Palaeornithologie; Borntrager: Berlin, Germany, 1933. [Google Scholar]
- Dughi, R.; Sirugue, F. Sur la structure des œufs des Sauropsidés vivants ou fossiles; le genre Psammornis Andrews. Bull. Soc. géol. Fr. 1964, 6, 240–252. [Google Scholar] [CrossRef]
- Dughi, R.; Sirugue, F. Sur le Psammornis rotschildi [sic]Andrews. Bull. Inst. Fr. Afr.Noire 1978, 40, 6–27. [Google Scholar]
- Hartert, E. Die Vögel der Paläarktischen Fauna. Band III; Friedländer & Sohn: Berlin, Germany, 1922. [Google Scholar]
- Knox, A.G.; Walters, M.P. Extinct and endangered birds in the collections of the Natural History Museum. Brit. Ornith. Club Occas. Pub. 1994, 1, 1–292. [Google Scholar]
- Hartert, E. Expedition to the central western Sahara. IV. Birds. Novit. Zool. 1913, 20, 37–76. [Google Scholar]
- Rothschild, W. On the former and present distribution ot the so called Ratitae or ostrich-like birds with certain deductions and a description of a new form by C.W. Andrews. In Verhandlungen des V. Internationalen Ornithologen-Kongresses in Berlin 1911; Schalow, H., Ed.; Deutsche Ornithologische Gesellschaft: Berlin, Germany, 1911; pp. 144–169. [Google Scholar]
- Andrews, C.W. On the pelvis and hind-limb of Mullerornis betsilei M.-Edw. & Grand.; with a note on the occurrence of a ratite bird in the upper Eocene beds of the Fayum, Egypt. Proc. Zool. Soc.London 1904, 1, 163–171. [Google Scholar]
- Rasmussen, D.T.; Simons, E.L.; Hertel, F.; Judd, A. hindlimb of a giant terrestrial bird from the Upper Eocene, Fayum, Egypt. Palaeontology 2001, 44, 325–337. [Google Scholar] [CrossRef]
- Geyr von Schweppenburg, H. Ins Land der Tuareg. J. Ornithol. 1918, 66, 121–176. [Google Scholar] [CrossRef] [Green Version]
- Abel, O. Die Stämme der Wirbeltiere; Walter de Gruyter & Co.: Berlin, Germany; Leipzig, Germany, 1919. [Google Scholar]
- Piveteau, J. Oiseaux. Aves Linné. In Traité de Paléontologie; Tome, V., Piveteau, J., Eds.; Masson: Paris, France, 1955; pp. 994–1091. [Google Scholar]
- Brodkorb, P. Catalogue of fossil birds. Part I. (Archaeopterygiformes through Ardeiformes). Bull. Florida State Mus. 1963, 7, 179–293. [Google Scholar]
- Dementiev, G.P. Class Aves. Birds. In Fundamentals of Palaeontology, Amphibians, Reptiles and Birds; Rozhdestvenskii, A.K., Tatarinov, L.P., Eds.; Nauka: Moscow, Russia, 1964; pp. 660–699. (In Russian) [Google Scholar]
- Swinton, W.E. Fossil Birds; British Museum (Natural History): London, UK, 1965. [Google Scholar]
- Lowe, P.R. On Some Struthious Remains:-1. Description of some pelvic remains of a large fossil ostrich, Struthio oldawayi, sp. n., from the Lower Pleistocene of Oldaway (Tanganyika Territory); 2. Egg-shell fragments referable to Psammornis and other Struthiones collected by Mr. St. John Philby in southern Arabia. Ibis 1933, 75, 652–658. [Google Scholar]
- Bibi, F.; Shabel, A.B.; Kraatz, B.P.; Stidham, T.A. New fossil ratite (Aves: Palaeognathae) eggshell discoveries from the late Miocene Baynunah Formation of the United Arab Emirates, Arabian Peninsula. Palaeont. Electr. 2006, 9, 1–13. [Google Scholar]
- Lowe, P.R. Struthious remains from Northern China and Mongolia, with descriptions of Struthio wimani, Struthio anderssoni and Struthio mongolicus, spp. nov. Palaeont. Sin. C 1931, 6, 1–47. [Google Scholar]
- Buffetaut, E. ‘Dragon’s eggs’ from the ‘Yellow Earth’: The discovery of the fossil ostriches of China. Historia Natural 2021, 11, 47–63. [Google Scholar]
- Buffetaut, E.; Angst, D. A giant ostrich from the Lower Pleistocene Nihewan Formation of North China, with a review of the fossil ostriches of China. Diversity 2021, 13, 47. [Google Scholar] [CrossRef]
- Schönwetter, M. Fossile Vogelei-Schalen. Novit. Zool. 1929, 35, 192–203. [Google Scholar]
- Schönwetter, M. Handbuch der Oologie. Lieferung 1; Akademie Verlag: Berlin, Germany, 1960. [Google Scholar]
- Heim de Balsac, H. Premières données sur les oiseaux du Sahara occidental. Alauda 1930, 2, 451–463. [Google Scholar]
- Moltoni, E. Risultati zoologici della missione inviata dalla R. Società Geografica Italiana per l’esplorazione dell’oasi de Giarabub (1926–1927). Uccelli. Ann. Mus. Civ. Stor. Nat. G. Doria 1928, 52, 387–401. [Google Scholar]
- Arambourg, C. & Magnier, P. Gisements de vertébrés tertiaires dans le bassin de Syrte, Libye. C. R. Acad. Sci. Paris 1961, 252, 1181–1183. [Google Scholar]
- Arambourg, C. Note préliminaire sur quelques vertébrés nouveaux du Burdigalien de Libye. C. R. somm. Séances Soc. Géol. Fr. 1961, 4, 107–109. [Google Scholar]
- Arambourg, C. Continental vertebrate faunas of the Tertiary of North Africa. In African Ecology and Human Evolution; Howell, F.C., Bourlière, F., Eds.; Aldine Publishing Company: Chicago, IL, USA, 1967; pp. 55–64. [Google Scholar]
- Savage, R.J.G.; Hamilton, W.R. Introduction to the Miocene mammal faunas of Gebel Zelten, Libya. Bull. Brit. Mus. (Nat. Hist.) Geol. 1973, 22, 513–527. [Google Scholar]
- Vickers-Rich, P. Significance of the Tertiary avifaunas from Africa (with emphasis on a mid to late Miocene avifauna from southern Tunisia). Ann. Geol. Surv. Egypt 1974, 4, 167–210. [Google Scholar]
- Mlíkovky, J. Early Miocene birds of Djebel Zelten, Libya. Čas. Národ. Muz., Řad. Přírod. 2003, 172, 114–120. [Google Scholar]
- Bédé, P. Bericht über den v. intern orn. Kongress 1911. Psammornis Rothschildi, nov. Gen., nov. sp., by C.-W. Andrews. Rev. crit. Paléozool. 1919, 23, 56–57. [Google Scholar]
- Choumowitch, W. Sur l’Autruche géante de la Koudiat Safra. C.R. 70e Congr. Ass. Fr. Avanc. Sci. Tunis 1951, 1, 172–175. [Google Scholar]
- Solignac, M. Le Pontien dans le Sud tunisien. Ann. Univ. Lyon N.S. I. Sci. Med. 1931, 48, 1–29. [Google Scholar]
- Rybkina, A.I.; Rostovtseva, Y.V. New evidence of the age of the Black Sea Pontian substage. Russ. J. Earth Sci. 2017, 17, ES5004. [Google Scholar] [CrossRef] [Green Version]
- Roux, H.; Douvillé, H. La géologie des environs de Redeyef (Tunisie). Bull. Soc. géol. Fr. 1911, 10, 646–660. [Google Scholar]
- Robinson, P.; Wiman, S.K. A revision of the stratigraphic subdivision of the Miocene rocks of sub-dorsale Tunisia. Notes Serv. Géol. Tunisie 1976, 42, 71–86. [Google Scholar]
- Regaya, K.; Laatar, S.; Chaouachi, A. Metlaoui. Carte géologique de la Tunisie 1/100,000 1991, 65. [Google Scholar]
- Werdelin, L. Chronology of Neogene mammal localities. In Cenozoic Mammals of Africa; Werdelin, L., Sanders, W.J., Eds.; University of Californa Press: Berkeley, CA, USA; London, UK, 2010; pp. 27–43. [Google Scholar]
- Vickers-Rich, P. A fossil avifauna from the Upper Miocene Beglia Formation of Tunisia. Notes Serv. Géol. Tunisie 1972, 35, 29–66. [Google Scholar]
- Biely, A.; Rakus, M.; Robinson, P.; Salaj, J. Essai de corrélation des formations miocènes au Sud de la Dorsale tunisienne. Notes Serv. Géol. Tunisie 1972, 38, 73–92. [Google Scholar]
- Biely, A.; Rakus, M.; Robinson, P. Le Miocène de la Tunisie. Ann. Geol. Surv. Egypt 1974, 4, 307–318. [Google Scholar]
- Mannaï-Tayech, B. Les séries silico-clastiques miocènes du Nord-Est au Sud-Ouest de la Tunisie: Une mise au point. Geobios 2006, 39, 71–84. [Google Scholar] [CrossRef]
- Mannaï-Tayech, B. The lithostratigraphy of Miocene series from Tunisia, revisited. J. Afr; Earth Sci. 2009, 54, 53–61. [Google Scholar] [CrossRef]
- Arambourg, C. La Paléontologie des Vertébrés en Afrique du Nord Française; XIXe Congrès Géologique International, Monographies régionales: Alger, Algeria, 1952. [Google Scholar]
- Monod, T. Nous aussi. Notes afr. Instit. Fr. Afr. Noire 1939, 4, 47–48. [Google Scholar]
- Monod, T. Sur la présence d’un Psammornis en Afrique Occidentale Française. C.R. 1e Conf. Intern. Africanistes Ouest 1951, 1, 202–203. [Google Scholar]
- Tessier, F.; Hébrard, L.; Lappartient, J.R. Découverte de fragments d’oeufs de Psammornis et de Struthio dans le Quaternaire de la presqu’île du Cap Blanc (République Islamique de Mauritanie). C.R. Acad. Sci. Paris D 1971, 273, 2418–2421. [Google Scholar]
- Voisin, C. Etude de la structure de fragments de coquilles d’oeufs de Psammornis rothschildi Andrews provenant de Mauritanie. L’Oiseau Rev. Franç. Ornith. 1971, 41, 245–256. [Google Scholar]
- Ortlieb, L. Recherches sur les formations plio-quaternaires du littoral ouest-saharien (28°30′- 20°40′ lat. N). Trav. Doc. O.R.S.T.O.M. 1975, 48, 1–267. [Google Scholar]
- Hébrard, L. Contribution à l’étude géologique du Quaternaire du littoral mauritanien entre Nouakchott et Nouadhibou, 18°—21° latitude Nord. Participation à l’étude des désertifications du Sahara. Doc. Lab. Géol. Facult. Sci. Lyon 1978, 71, 1–210. [Google Scholar]
- Hébrard, L.; Elouard, P.; Faure, H. La synthèse stratigraphique du Quaternaire du littoral mauritanien entre Nouakchott et Nouadhibou. In Lexique Stratigraphique International. Nouvelle Série n°1. Afrique de l’Ouest. Introduction Géologque et Termes Stratigraphiques; Fabre, J., Ed.; Pergamon Press: Oxford, UK, 1983; pp. 158–170. [Google Scholar]
- Giresse, P.; Barrusseau, J.P.; Causse, C.; Diouf, M. Successions of sea-level changes during the Pleistocene in Mauritania and Senegal distinguished by sedimentary facies study and U/Th dating. Mar. Geol. 2000, 170, 123–139. [Google Scholar] [CrossRef]
- Vernet, R.; Rodrigue, A.; Tous, P. Les tests d’œuf d’autruche gravés du littoral atlantique saharien du nord du Banc d’Arguin à l’oued Draa. Sahara 2006, 17, 59–72. [Google Scholar]
- Vernet, R. Préhistoire de la Mauritanie; Centre Culturel Français de Nouakchott—Sépia: Nouakchott, Mauritania, 1993. [Google Scholar]
- Vernet, R. Le Golfe d’Arguin de la Préhistoire à l’Histoire: Littoral et plaines intérieures; Parc National du Banc d’Arguin: Nouakchott, Mauritania, 2007. [Google Scholar]
- Mikhailov, K.E. Fossil and recent eggshell in amniotic vertebrates: Fine structure, comparative morphology and classification. Spec. Pap. Palaeont. 1997, 56, 1–80. [Google Scholar]
- Murray, P.F.; Vickers-Rich, P. Magnificent Mihirungs: The Colossal Flightless Birds of the Australian Dreamtime; Indiana University Press: Bloomington, IN, USA, 2004. [Google Scholar]
- Angst, D.; Buffetaut, E. Paleobiology of Giant Flightless Birds. Elsevier & ISTE Press: Oxford, UK; London, UK, 2017. [Google Scholar]
- Zelenkov, N.V.; Lavrov, A.V.; Startsev, D.B.; Vislobokova, I.A.; Lopatin, A.V. A giant early Pleistocene bird from eastern Europe: Unexpected component of terrestrial faunas at the time of early Homo arrival. J. Vert. Paleont. 2019, 39, e1605521. [Google Scholar] [CrossRef]
- Camps-Fabrer, H. La disparition de l’autruche en Afrique du Nord; Institut Français des Sciences Humaines en Algérie: Alger, Algeria, 1963. [Google Scholar]
- Hodos, T.; Cartwright, C.R.; Montgomery, J.; Nowell, G.; Crowder, K.; Fletcher, A.C.; Gönster, Y. The origins of decorated ostrich eggs in the ancient Mediterranean and Middle East. Antiquity 2020, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Arambourg, C. Vertébrés villafranchiens d’Afrique du Nord (Artiodactyles, Carnivores, Primates, Reptiles, Oiseaux); Fondation Singer-Polignac: Paris, France, 1979. [Google Scholar]
- Mourer-Chauviré, C.; Geraads, D. The Struthionidae and Pelagornithidae (Aves: Struthioniformes, Odontopterygiformes) from the Late Pliocene of Ahl Al Oughlam, Morocco. Oryctos 2008, 7, 169–194. [Google Scholar]
- Mourer-Chauviré, C.; Senut, B.; Pickford, M.; Mein, P.; Dauphin, Y. Ostrich legs, eggs and phylogenies. S. Afr. J. Sci. 1996, 92, 492–495. [Google Scholar]
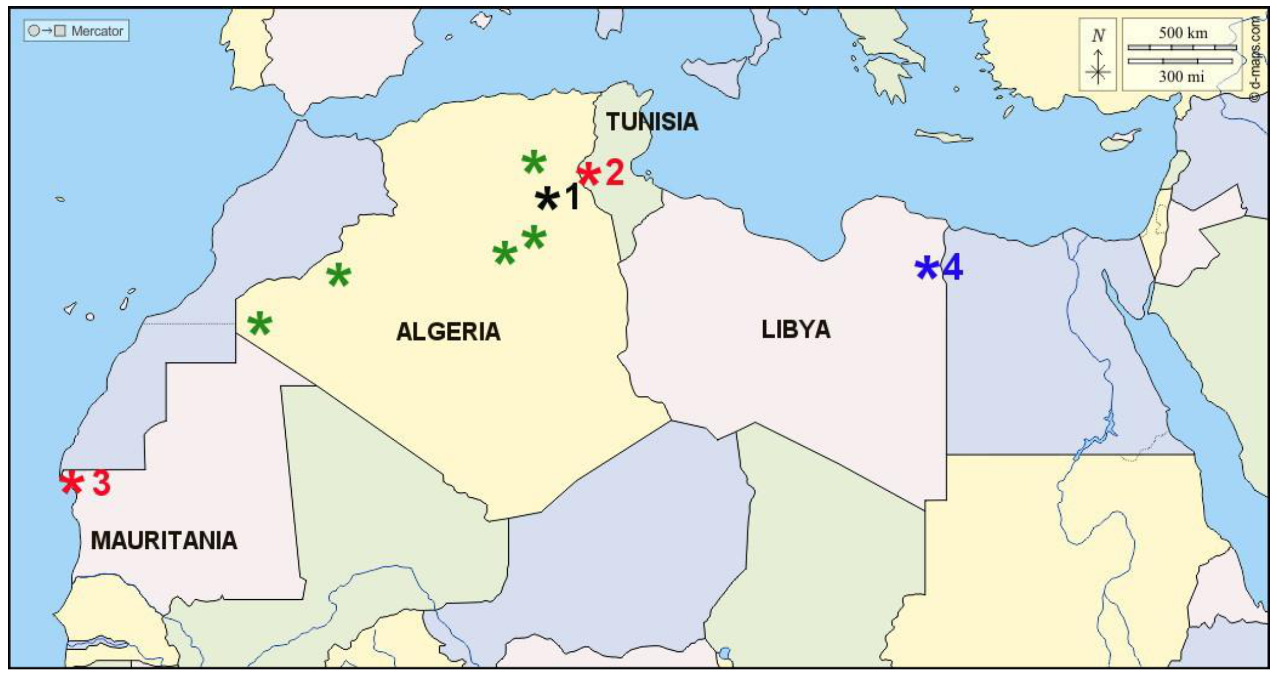
Figure 1.
Distribution map of Psammornis localities in North Africa: black star (1)—type locality of Psammornis rothschildi Andrews, near Touggourt, Algeria; red stars—localities with Psammornis eggshells in situ in a stratigraphic context: 2—Segui Formation (latest Miocene), Chebket Safra near Moulares, south-western Tunisia, 3—Aguerguerian (Middle Pleistocene), Lévrier Bay area, north-western Mauritania; blue star (4)—type locality of Psammornis libycus Moltoni (a probable synonym of Struthio camelus), near Giarabub, Libya; unnumbered green stars—Psammornis finds without a stratigraphic context in the Algerian Sahara.
Figure 1.
Distribution map of Psammornis localities in North Africa: black star (1)—type locality of Psammornis rothschildi Andrews, near Touggourt, Algeria; red stars—localities with Psammornis eggshells in situ in a stratigraphic context: 2—Segui Formation (latest Miocene), Chebket Safra near Moulares, south-western Tunisia, 3—Aguerguerian (Middle Pleistocene), Lévrier Bay area, north-western Mauritania; blue star (4)—type locality of Psammornis libycus Moltoni (a probable synonym of Struthio camelus), near Giarabub, Libya; unnumbered green stars—Psammornis finds without a stratigraphic context in the Algerian Sahara.

Figure 2.
The Psammornis-bearing red clays on the southern flank of the Chebket Safra, SW of the city of Moulares in south-western Tunisia. The level containing remains of broken eggs preserved in situ is shown by the yellow asterisk. Photo by E.G. Gobert, modified after Chomowitch (1951).
Figure 2.
The Psammornis-bearing red clays on the southern flank of the Chebket Safra, SW of the city of Moulares in south-western Tunisia. The level containing remains of broken eggs preserved in situ is shown by the yellow asterisk. Photo by E.G. Gobert, modified after Chomowitch (1951).

Figure 5.
Section showing Aguerguerian deposits at ‘point kilométrique 52′ at the north-eastern end of Lévrier Bay, north-western Mauritania. White cross-bedded sandstones alternate with yellowish-brown clayey sandstones containing the snail Helix gruveli and in situ Psammornis eggshells. The main Psammornis-bearing horizon is at the bottom of the section. Modified after Hébrard [56].
Figure 5.
Section showing Aguerguerian deposits at ‘point kilométrique 52′ at the north-eastern end of Lévrier Bay, north-western Mauritania. White cross-bedded sandstones alternate with yellowish-brown clayey sandstones containing the snail Helix gruveli and in situ Psammornis eggshells. The main Psammornis-bearing horizon is at the bottom of the section. Modified after Hébrard [56].

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Buffetaut, E. The Enigmatic Avian Oogenus Psammornis: A Review of Stratigraphic Evidence. Diversity 2022, 14, 123. https://0-doi-org.brum.beds.ac.uk/10.3390/d14020123
AMA Style
Buffetaut E. The Enigmatic Avian Oogenus Psammornis: A Review of Stratigraphic Evidence. Diversity. 2022; 14(2):123. https://0-doi-org.brum.beds.ac.uk/10.3390/d14020123
Chicago/Turabian StyleBuffetaut, Eric. 2022. "The Enigmatic Avian Oogenus Psammornis: A Review of Stratigraphic Evidence" Diversity 14, no. 2: 123. https://0-doi-org.brum.beds.ac.uk/10.3390/d14020123
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.