
Diversity of Rotifers in Small Rivers Affected by Human Activity


1
Institute of Biology, Biotechnology and Environmental Protection, Faculty of Natural Sciences, University of Silesia in Katowice, Bankowa 9, 40-007 Katowice, Poland
2
Department of Invertebrate Zoology and Hydrobiology, Faculty of Biology and Environmental Protection, University of Lodz, Banacha 12/16, 90-237 Lodz, Poland
*
Author to whom correspondence should be addressed.
Diversity 2022, 14(2), 127; https://0-doi-org.brum.beds.ac.uk/10.3390/d14020127
Submission received: 28 December 2021
/
Revised: 7 February 2022
/
Accepted: 8 February 2022
/
Published: 10 February 2022
(This article belongs to the Special Issue Biodiversity of Rotifers)
Abstract
:The rivers flowing through Upper Silesia and the adjacent areas (Southern Poland) are affected by various anthropogenic pressures including urbanisation, agriculture and animal husbandry, as well as industry (e.g., mining), which are reflected in the measured physical and chemical water parameters. The species composition of rotifers relative to a variety of microhabitats was studied in eight small rivers of this region in 2017. Our research is a comprehensive and up-to-date analysis that focuses on the rotifers in small rivers and shows the diversity of rotifers relative to the microhabitats and environmental variables. The diversity of rotifers ranged from 0 to 23 taxa in individual samples. In the studied rivers, 129 taxa of rotifers were found. Notommata groenlandica, a species that has not been recorded in the country for 100 years, was found in two rivers. The Kruskal–Wallis one-way ANOVA and Dunn’s multiple comparison post hoc tests revealed statistically significant differences in the median number of rotifer taxa between the abiotic types of rivers, rivers, sampling sites, microhabitats and seasons. A multiple regression analysis revealed a significant relationship (correlation) between the number of rotifer taxa, and the concentration of nitrites, total dissolved solids and dissolved oxygen in the water.
Keywords:
anthropogenic pressure; human disturbance; salinisation; running waters; habitat; Rotifera1. Introduction
To date, more than 2030 species that belong to the Phylum Rotifera are known, which are classified into three main groups: the exclusively parthenogenetic subdivision Bdelloidea (about 461 clonal species), the largest subdivision Monogononta (about 1570 species), and the marine subdivision Seisonida (4 species) [1]. However, studies regarding their integration approaches indicate that the diversity of rotifers is much higher than is currently estimated [2,3]. Rotifers are considered to be a valuable tool in environmental assessments, mainly because they are quite abundant, and thus, are an important part of most non-marine food webs [4,5]. In addition, rotifers are generally cosmopolitan, and their distribution is generally limited by environmental conditions, but may also be limited by biogeographic barriers [6,7]. It is also known that because of their evolutionary adaptations, rotifers segregate according to the specificity of habitats [8,9]. For example, planktonic rotifers are used to monitor the water of dam reservoirs and lakes [10,11,12]. Surprisingly, despite the great diversity of rotifers and their habitat preferences, little is known about the habitat preferences of the periphytic rotifers [13]. Macrophytes can shape the diversity of rotifers by providing a food source and a suitable habitat for their life. On the other hand, periphytic rotifers can also cause the growth of macrophytes, and thus provide food for other animals [13,14,15]. Therefore, research that takes into account the approach to the habitat selectivity of rotifers is important and can be used to monitor aquatic ecosystems [16].
According to Limnofauna Europaea [17], five zoogeographical regions converge in Poland. Four of them (the Central Highlands, the Carpathians, the Central Plains and the Eastern Plains) converge in Upper Silesia, which is one of the largest coal basins in the world. Mining activity causes the discharge of saline mine waters into rivers (even after mines are closed for economic reasons), mainly through the smaller rivers that carry their waters to the Odra River and the Vistula River [18,19]. In addition, rivers flowing through agricultural areas are characterised by high concentrations of nutrients in the water [20,21]. Other threats to the stability of river ecosystems include changes in land use, toxic and domestic waste and climate change [22,23]. These individual changes in aquatic ecosystems could impair the natural functioning, and could also modify the structure and function of biotic communities [24,25]. The decline in biodiversity is also caused by overexploitation, urban development, invasion and disease, system modification, human disturbance, transport and energy production [26]. To summarise, the rivers of the Upper Silesia region and adjacent areas are affected by most of these “big killers”. The territory of Southern Poland includes both areas with significant anthropogenic transformations as a result of strong industrialisation and urbanisation, as well as less transformed and legally protected areas. Therefore, the rivers flowing through Southern Poland that have different land uses in their catchment areas differ in terms of the degree of anthropogenic pressure. Consequently, this region is an excellent area for ecological research into various aquatic ecosystems, especially in rivers.
The data on rotifers that was found in the scientific literature using the term “rotifer” via a bibliometric search in the popular scientific databases include over 22 000 documents. Using the same method, it was found that the term “rotifer” and the term “river” reveal six times fewer documents related to this type of water body. Meanwhile, when a term related to stagnant waters was used, four times as many documents were found than for the term “river”. This finding is in opposition to the trend of scientific interest in rivers as opposed to other types of inland waters [27]. This simple method revealed disproportions in the interest in the research on rotifers in lotic and lentic environments. Therefore, this reflects the insufficient state of study concerning the diversity of rotifers in small rivers. This issue perfectly fits in our research, in which we attempted to determine the habitat preferences of the rotifers that occur in small rivers subjected to different kinds of anthropogenic pressure.
The objectives of the research were to determine the diversity of the rotifers in small rivers subjected to various types of anthropogenic pressure and to reveal the habitat (namely microhabitats and environmental conditions) preferences for the identified rotifer species. In addition, we indicate further directions for research on rotifers in future ecological studies.
2. Materials and Methods
The research was conducted in the rivers that flow through one of the highly industrialised and urbanised regions in Europe, i.e., Upper Silesia and adjacent areas (Southern Poland) from spring to autumn 2017. Eight rivers of four abiotic types in the catchments of the Vistula and Odra rivers within two ecoregions (the Central Plains and the Carpathians) were selected according to the European Union Water Framework Directive (EU WFD) [28]. Depending on the degree of anthropogenic pressure, two sampling sites for each river were selected, i.e., one reference site and the other under significant anthropogenic pressure (Figure 1 and Figure S1). The general characteristics of the study sites and the main anthropogenic pressures are presented in Table 1.
Water samples for the physical and chemical analyses were collected before the biological sampling. The electrical conductivity (EC), total dissolved solids (TDS), temperature and dissolved oxygen were measured in the field using a Multi 3410 WTW meter. The salinity was measured in the field as the EC and then converted, according to Piscart et al. [29]. The concentrations of selected ions, total hardness and alkalinity were analysed in the laboratory according to Hermanowicz et al. [30].
The samples of rotifers were collected from various microhabitats: open water, stones, bottom sediments, macrophytes and diatom aggregations. The planktonic samples of rotifers were collected using the standard methods by pouring 20 dm3 of water through a plankton net with a mesh size of 50 µm. The periphytic samples of rotifers were collected from different substrata (macrophytes and diatom aggregations) by cutting different fragments (a total of 25 cm2 for each surface) of each macrophyte and diatom using a soft toothbrush [31,32]. The rotifer samples from the stones were collected from the same surface using a soft toothbrush. Biological samples from bottom sediments were collected using a sharp-edged cylinder with a surface area of 20 cm3, according to Bielańska-Grajner et al. [33]. Rotifer species were classified according to Segers [1] and identified according to citing publications [33,34,35,36,37,38].
The significance of the differences in the median values of the environmental variables between the abiotic types of rivers, the rivers and the sampling sites, as well as in the median values of the number of rotifer taxa between the sampling sites, rivers, abiotic types of rivers and different microhabitats [different substratum, types of plant growth and leaf morpha groups including diatoms, which were grouped (cumulated) according to study seasons] and different seasons were calculated using the Kruskal–Wallis one-way ANOVA and Dunn’s multiple comparison post hoc tests. Multiple regression techniques (including multilinearity checking) were used to elucidate the relationship between the species richness (number of rotifer taxa) and selected environmental variables, and then to assess the influence of an anthropogenic transformation on the rotifers in the studied rivers. Therefore, the data were first log-transformed to approximately conform to normality. The statistical analyses were performed using Statistica version 13.1.
3. Results
The conducted research indicated a large amount of diversity among the abiotic types of rivers (also the rivers and sampling sites) relative to the abiotic parameters. The physical and chemical parameters of the water in most sampling sites were influenced by the geological substratum of the catchment area of the rivers (calcareous, flysch, siliceous). However, the impact of anthropogenic pressure was also reflected, e.g., in the relatively high concentrations of nitrates (up to 79.74 mg dm−3), nitrites (up to 9.96 mg dm−3) and phosphates (up to 19.20 mg dm−3) in the water and in modifications of the riverbed at the sampling sites. In addition, very high values of EC (up to 46 600 µS cm−1), TDS (23 300 mg dm−3), total hardness (4857.92 mg CaCO3 dm−3), the concentrations of chlorides (up to 17 028 mg dm−3) and temperature (up to 29.1°C), were recorded in the lower course of the Bolina River (abiotic type 5) (Table 2, Tables S1 and S2).
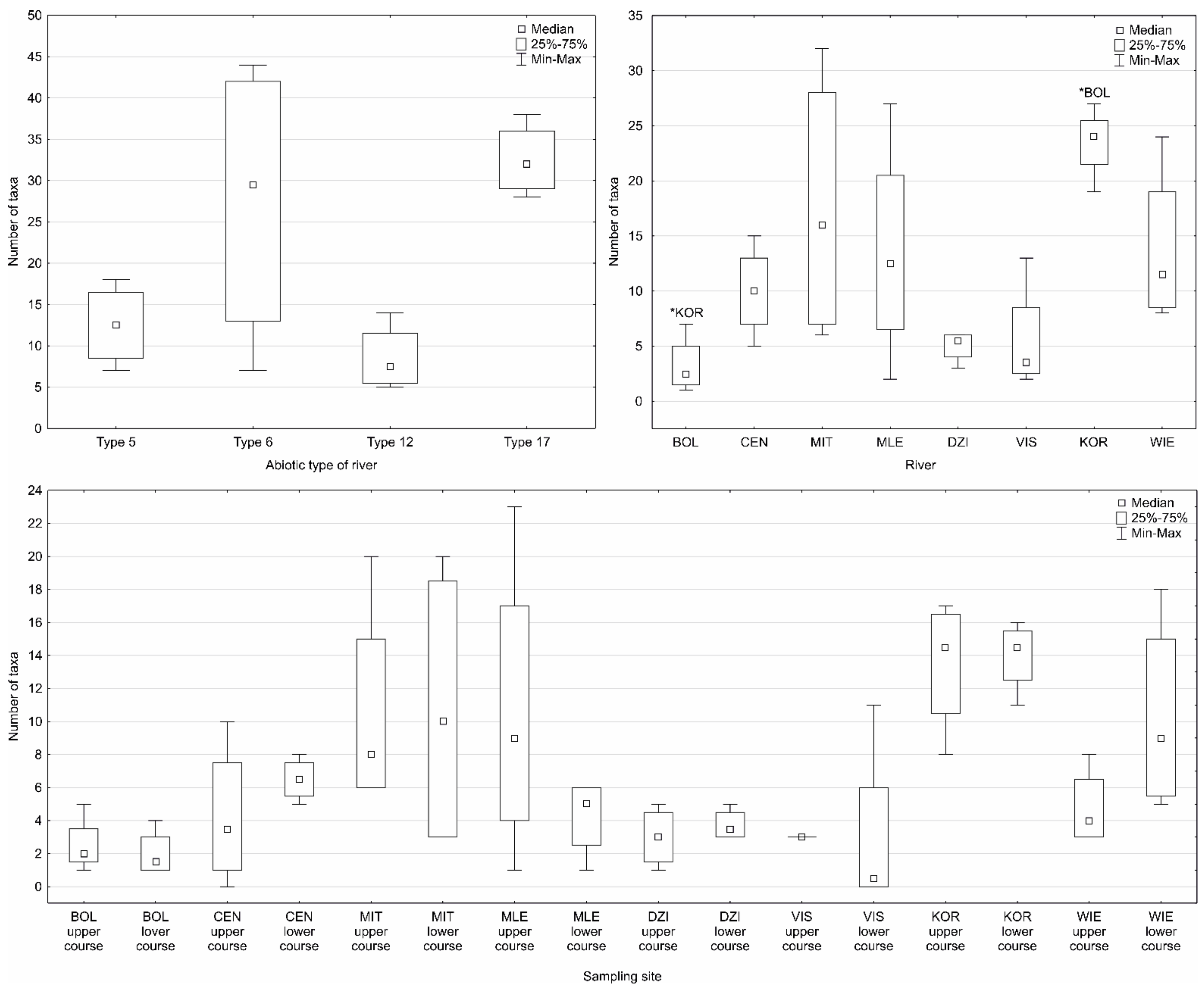
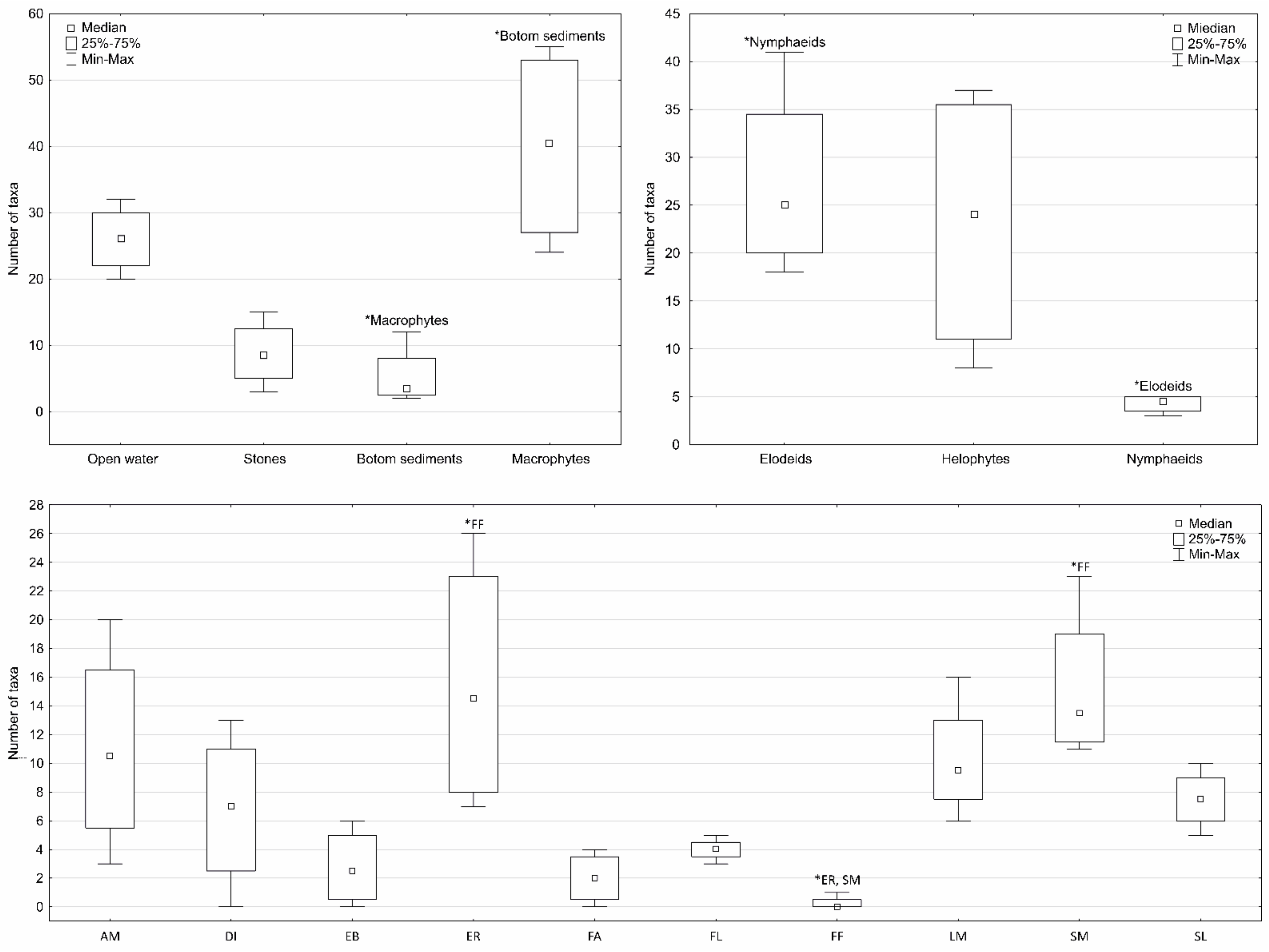
During the entire study period, 129 taxa (including 104 species and one subspecies) of rotifers were identified in all of the sampling sites (Table 3). The halophilic rotifer species Brachionus plicatilis was found in the Bolina River (only in the planktonic samples). In contrast, Notommata groenlandica was found in the upper course of the Centuria River and the lower course of the Wiercica River. N. groenlandica was rediscovered after more than 100 years in the inland waters of the territory of Poland. This species was only found in the periphytic samples that had been collected from Glyceria nemoralis (upper course of the Centuria River) and Sparganium erectum (lower course of the Wiercica River). Species diversity was the highest in the lowland sandy streams (79 taxa, including 69 species), while the lowest in the flysch streams (21 taxa, including 18 species). However, the lowest number of taxa was recorded in the most degraded river (the most anthropogenically salinised), i.e., in the Bolina River (ten taxa, including eight species) (Figure 2). For most of the samples, the number of Monogononta taxa dominated. The reverse trend was observed only in the sampling sites of flysch streams in which Bdelloidea taxa dominated. When the seasons were considered, a higher number of rotifer taxa were recorded in autumn. In contrast, a higher number of taxa were found in summer in the upper course of the Korzenica River (Table 3 and Table 4). The highest number of rotifer taxa was recorded in autumn in the periphyton samples, while the lowest was recorded in the bottom sediment samples. Among the periphyton samples, the highest number of taxa was recorded in the samples taken from the elodeids and the lowest from the nymphaeids. When analysing the leaf morph groups, the highest number of taxa was recorded on emergent reeds, sedges, while the lowest was recorded on the filamentous algae and floating-leaves (rooted) (Figure 2).
The differences in the median values of most of the physical and chemical parameters of the waters and the morphological features between the sampling sites were significant (p < 0.01) (the Kruskal–Wallis one-way ANOVA and the Dunn’s multiple comparison post hoc tests) (Table 2, Tables S1 and S2). The Kruskal–Wallis one-way ANOVA and the Dunn’s multiple comparison post hoc tests revealed statistically significant differences (p < 0.05) in the median number of rotifer taxa between the sampling sites, rivers, various microhabitats and seasons (Figure 2 and Figure 3, Table 4).
A multiple regression analysis revealed a significant relationship between the dependent variable, the species richness and at least three of the studied environmental variables. Table 5 presents the results of this analysis and includes information about the β-coefficients, indicating that species richness was significantly correlated with the concentration of nitrites, TDS and dissolved oxygen in the water. A regression analysis showed that 12.2% of the variance in species richness was explained by these variables (R2 = 0.548, p < 0.001, SE = 0.287). Significant relationships existed between the species richness and the following environmental variables: the concentration of nitrites, TDS and dissolved oxygen in the water. The species richness increased with an increase in the concentration of nitrites, whereas species richness decreased with an increase in the concentration TDS and dissolved oxygen in the water (Table 5).
4. Discussion
It is well documented that the number of aquatic invertebrate taxa increases with a river’s course, which is consistent with the River Continuum Concept (RCC) [39]. However, our results indicate otherwise. The more human-induced disturbances and the degree of their intensity, the more likely it is that this trend will be reversed [40,41]. For example, Afanasyev et al. [42] found a higher diversity of rotifers in flowing water bodies than in the waters of the Vita River (estuarine region of the Vita River). We especially observed this phenomenon in the anthropogenically salinised rivers, i.e., in the Bolina and Mleczna rivers in which we recorded a lower number of rotifer taxa in the lower courses compared to the upper courses of these rivers. A broader analysis of the impact of anthropogenic salinity on rotifer communities in one of the most anthropogenically salinised rivers was presented in our previous studies [43,44]. However, the impact of the anthropogenic salinisation of flowing waters on rotifer communities is still insufficiently documented. Thus, in this work, we add another contribution to this topic. In addition, we found rare halophilic rotifers in the anthropogenically salinised rivers, namely Brachionus plicatilis in the Bolina River and Testudinella clypeata in the Mleczna River. On the other hand, we also observed a decrease in the number of rotifer taxa along with an increase in the concentration of dissolved oxygen in the water in the reference sampling sites. These sampling sites were characterised by a relatively high concentration of dissolved oxygen and a low concentration of nutrients. Therefore, they provided favourable conditions for the occurrence of only a few oligotrophic species. Another regularity that is observed in natural river ecosystems is that the nutrient concentrations increase with the course of the river [31]. We observed a statistically significant positive correlation between the species richness and the concentration of nitrites in the water. It is known that the availability of nutrients, e.g., phosphates, is crucial for the development of rotifers [43,45]. Moreover, the different species of rotifers can consume different sources of food, namely, the diet of rotifers can consist of detritus, diatoms, algae or protozoans [46,47]. The importance of phosphates is reflected in the fact that in water, they occur in the dissolved form as orthophosphates (PO43−), which are attached to suspended inorganic particles and dissolved organic particles, mainly in bacteria and detrital particles. Other forms of phosphorus must be transformed into orthophosphates, which can only then be directly absorbed by algae and used by other organisms [40]. Hence, a higher nutrient concentration in the water promotes a greater diversity of rotifers. Therefore, our results indicate (especially in the Korzenica, Mleczna and Centuria rivers) that a greater diversity of rotifers was observed in the sampling sites richer in nutrients (contaminated by agriculture and fishponds, domestic sewage or even at reference ones with a higher concentration of nutrients) than in the sampling sites characterised by a greater depth and width of the riverbed.
Our observations are consistent with the focus on local conditions when analysing microinvertebrates, especially in small rivers [48,49]. When the local pollution is strong, it can significantly affect the shaping of rotifer communities. Therefore, this indicates that the rotifer communities in small rivers are a valuable tool that can be used in assessing human pressure on the aquatic environment. Therefore, further studies are needed, including, in particular, a quantitative analysis of the rotifer communities in different types of rivers under varying anthropogenic pressures (and including reference sites).
The planktonic animal organisms that constitute part of zooplankton (Cladocera, Copepoda, Rotifera) are indicators of anthropogenic changes in running aquatic environments [42,50,51,52,53,54,55,56,57,58,59]. However, in both small lowland and highland rivers, the retention time that is required to develop planktonic organisms and to maintain a large abundance of planktonic organisms may be too short. As a result, in small rivers, the zooplankton can only be represented by tychoplankton. In addition, a slower flow velocity and less intense turbulence can help to reduce the diversity and abundance of planktonic organisms through the phenomena of sedimentation and fish predation [43,60,61]. This results in poor knowledge about the zooplankton communities in small lowland and upland rivers, particularly in mountain rivers [55]. For example, the first study recently showed spatial changes in the zooplankton composition in a small mountain river relative to environmental changes in the catchment area and riverbed transformations [62]. However, studies of the species richness of rotifers in small rivers in the Ukraine (tributaries of the Dnieper River) showed a high species richness of rotifers in natural rivers and in rivers with a periodic alteration of the direction and velocity of the flow. High numbers of rotifer taxa were related to the location of rivers in natural flood land because the species richness of rotifers in the rivers located in flood land that was partially or totally drained (regulated or canalised rivers) was visibly lower [53]. In some of the studied rivers, we identified rotifers typical for pelagic zooplankton, for example, Asplanchna priodonta, Brachionus angularis, Filinia longiseta, Keratella cochlearis, K. quadrata, K tecta, Polyarthra vulgaris and Pompholyx sulcata. However, their presence can be explained by the connectivity of these rivers with water reservoirs. Therefore, the planktonic samples from most of the rivers were mainly represented by tychoplankton. Thus, our research is a comprehensive analysis and up-to-date survey that focuses on rotifers in small lowland and upland rivers, showing the diversity of rotifers relative to various microhabitats and environmental variables. The methods we used were different because of the various types of microhabitats, such as open waters, stones, macrophytes and bottom sediments. A high level of the contamination of the samples of the bottom sediments and macrophytes (including diatoms) collected from some of the sampling sites, which was caused, among others, by the presence of coal silt in rivers under the influence of mine waters, made it necessary to analyse live samples. This situation meant that preserving the periphyton samples would make it impossible to identify the individuals of most species from the Bdelloidea subdivision because they were abundant in such samples. Due to the long duration of analysing such quantitative samples, this is impossible to realise in a small team. Therefore, we suggest that each microhabitat be analysed separately. This will enable a proper quantitative analysis of the periphyton samples despite the lack of a uniform sampling methodology, which makes it difficult to compare studies. The importance of this problem was indicated more than 20 years ago [13], and it has not been solved as yet.
Until recently, research on rotifers has focused almost entirely on their role in the environment in areas of zooplankton research [63,64]. However, recently increased attention has been paid to the functional role of rotifers with particular reference to the functional feeding groups [41,65]. It is known that macrophytes create extremely complicated habitats that determine the formation of many ecological niches, as well as the possibility for the coexistence of various organisms [16,66,67]. In addition, the role of habitat heterogeneity is assessing the diversity of other groups of organisms (rotifers in this case) as well as the potential of the sampling taken approach (especially rotifers) from a different habitat, which can not only be used in assessing the quality of the environment in large water bodies [16,68,69,70]. Studies concerning small water reservoirs have shown that the diversity of rotifers depends on the water reservoir zone and its characteristics. For example, a recently published study showed that a littoral zone with elodeids had a greater diversity of rotifers than the same zone with helophytes or the pelagic [16]. Our studies, which focused on both planktonic and periphytic samples, were compatible with these results. Moreover, we took it a step further and showed that the taxonomic diversity of rotifers depends on the leaf morpha groups, including diatoms. However, such an approach requires further detailed research that is mainly based on seasonal quantitative studies. This is important because it has been documented that, in shallow ecosystems, plant morphology can play a crucial role in the ecological assessment and protection of small water bodies [71,72]. Research that is based on rotifers in small rivers has not focused on this aspect. Future studies on rotifers in small rivers should not only include the analysis of planktonic, but also periphytic samples. This would offer more possibilities for analyses and could be the basis for their use in river monitoring. In addition to the arguments presented above, this is supported by the common occurrence of macrophytes in the aquatic environment, as well as the ease of collecting periphyton samples.
Detailed taxonomic studies of the rivers of Upper Silesia and adjacent areas revealed a large diversity of rotifers, which were represented by almost 130 taxa, including some rare species, mainly for Poland, in particular: Brachionus plicatilis, Cephalodella delicata, C. globata, C. misgurnus, Dicranophorus rostratus, Encentrum marinum, E. lupus, E. tyrphos, Lepadella elliptica, Limnias melicerta, Lindia torulosa, L. truncata, Notommata groenlandica, Proales theodora, Proalinopsis squamipes and Testudinella clypeata. One of these, namely N. groenlandica, is known from several countries in Europe and in this study, it was rediscovered in the present-day territory of Poland after more than 100 years. We found this species in the periphytic samples (on Glyceria nemoralis and Sparganium erectum) in two rivers (the Centuria and Wiercica rivers). Its current closest known place of occurrence is at a distance of at least 400 km [73,74]. Our results provide a great deal of new data on the ecology of many species of rotifers. For example, to date, Notommata groenlandica has been known to occur only in acidic environments such as peat bogs, ponds and has been found only in moss and acid silt samples [33,75]. These observations indicate that these species, which are considered to be rare, are likely to be spread more widely and that the habitat preferences of rotifers might be much broader than those that are currently known. Therefore, more research is required on the rotifers in small rivers.
5. Conclusions
The present study revealed that rotifers inhabit rivers subjected to various types of anthropogenic pressure that have a wide range of physical and chemical parameters of the water. They were found in extremely degraded rivers with a salinity of up to 33.55‰ (the Bolina and Mleczna rivers), which is comparable to that of the seas (e.g., the salinity of the North Sea ranges at about 35.0‰), as well as a relatively high conductivity, and the concentrations of nutrients, i.e., ammonium, nitrites, nitrates, phosphates. The maximum number of rotifer taxa was recorded in the mid-altitude calcareous streams, with a fine particulate substratum on loess (type 6) and very high concentrations of phosphates up to 19.20 mg dm−3. Our study showed that the diversity of the rotifer communities in small rivers affected by various types of anthropogenic pressure is influenced by several (statistically important) environmental factors, including TDS, the concentration of dissolved oxygen in the water, nitrites, and also by the seasons. The research also revealed a large taxonomic diversity of rotifers relative to the different microhabitats. The highest taxonomic richness of rotifers was observed in the rivers characterised by a high concentration of nutrients in the water. At the same time, the highest number of rotifer taxa was recorded on macrophytes (elodeids) compared to the other microhabitats (open waters, stones, bottom sediments or other forms of plant growth). Periphytic rotifers and tychoplankton mainly represented the rotifer communities in these rivers. Among the identified species, the occurrence of Notommata groenlandica in Poland was rediscovered after more than 100 years. In addition, in this paper, new data on the ecology of some species of rotifers are provided. Therefore, small rivers, mainly those with a large diversity of aquatic vegetation, provide a suitable habitat for the development of rotifer communities. Since planktonic rotifers have successfully been used in monitoring stagnant waters, the presented results can be used as baseline study for the use of rotifers in monitoring small rivers. The finding related to the type of samples, i.e., bottom sediments, stones, should be considered since we proved that not all types of samples can be suitable for this kind of research. Finally, the research has shown that rotifers can be used as a valuable tool in assessing human pressure on small rivers, although more research is required, especially when comparing them to the reference sampling sites.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/d14020127/s1, Figure S1: Study area and sampling sites, Table S1: The physical and chemical parameters of the waters of the studied rivers (ranges) and the results of the Kruskal–Wallis one-way ANOVA and Dunn’s multiple comparison post hoc tests (superscript a, b, c, d, e, f, g, h denote significant differences between the rivers), Table S2: The physical and chemical parameters of the waters of the sampling sites (ranges) and the results of the Kruskal–Wallis one-way ANOVA and Dunn’s multiple comparison post hoc tests (superscript a, b, c, d, e, f, g, h denote significant differences among the rivers). Abbreviations: UC – upper course, LC – lower course.
Author Contributions
Conceptualisation, D.H., I.B.-G. and I.L.; methodology, D.H., I.B.-G. and I.L.; formal analysis, D.H.; investigation, D.H., I.B.-G., I.L. and A.S.; resources, D.H.; data curation, D.H.; writing—original draft preparation, D.H.; writing—review and editing, D.H., I.B.-G., I.L. and A.S.; visualization, D.H.; supervision, I.L. and I.B-G.; project administration, D.H.; funding acquisition, D.H. and I.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available upon reasonable request from the corresponding author.
Acknowledgments
The authors are deeply indebted to Szymon Jusik, University of Life Science in Poznań, Poland, for confirming the taxonomic identification of the bryophytes. The authors are also grateful to the Editors and the anonymous reviewers for their valuable suggestions and comments on this manuscript, and to Michele L. Simmons, the University of Silesia, Katowice for improving the English style.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Segers, H. Annotated checklist of the rotifers (Phylum Rotifera), with notes on nomenclature, taxonomy and distribution. Zootaxa 2007, 1564, 1–104. [Google Scholar] [CrossRef]
- Mills, S.; Alcántara-Rodríguez, J.A.; Ciros-Pérez, J.; Gómez, A.; Hagiwara, A.; Galindo, K.H.; Jersabek, C.D.; Malekzadeh-Viayeh, R.; Leasi, F.; Lee, J.-S.; et al. Fifteen species in one: Deciphering the Brachionus plicatilis species complex (Rotifera, Monogononta) through DNA taxonomy. Hydrobiologia 2017, 796, 39–58. [Google Scholar] [CrossRef] [Green Version]
- Schenk, J.; Fontaneto, D. Biodiversity analyses in freshwater meiofauna through DNA sequence data. Hydrobiologia 2019, 847, 2597–2611. [Google Scholar] [CrossRef]
- Ejsmont-Karabin, J.; Karpowicz, M. Rotifera in lake subhabitats. Aquat. Ecol. 2021, 55, 1285–1296. [Google Scholar] [CrossRef]
- Wallace, R.L.; Walsh, E.J.; Nandini, S.; Sarma, S.S.S. A meta-analysis of benthic rotifer community structure as a function of lake trophic state. Aquat. Ecol. 2021, 55, 1297–1304. [Google Scholar] [CrossRef]
- Ejsmont-Karabin, J. Does the world need faunists? Based on rotifer (Rotifera) occurrence reflections on the role of faunistic research in ecology. Int. Rev. Hydrobiol. 2019, 104, 49–56. [Google Scholar] [CrossRef]
- Iakovenko, N.; Smykla, J.; Convey, P.; Kašparová, E.; Kozeretska, I.; Trokhymets, V.; Dykyy, I.; Plewka, M.; Devetter, M.; Duriš, Z.; et al. Antarctic bdelloid rotifers: Diversity, endemism and evolution. Hydrobiologia 2015, 761, 5–43. [Google Scholar] [CrossRef]
- Kuczyńska-Kippen, N. The distribution of rotifers (Rotifera) within a single Myriophyllum bed. Hydrobiologia 2003, 506-509, 327–331. [Google Scholar] [CrossRef]
- Špoljar, M.; Dražina, T.; Šargač, J.; Borojević, K.K.; Žutinić, P. Submerged macrophytes as a habitat for zooplankton development in two reservoirs of a flow-through system (Papuk Nature Park, Croatia). Ann. de Limnol.-Int. J. Limnol. 2012, 48, 161–175. [Google Scholar] [CrossRef] [Green Version]
- Ejsmont-Karabin, J. The usefulness of zooplankton as lake ecosystem indicators: Rotifer trophic state index. Pol. J. Ecol. 2012, 60, 339–350. [Google Scholar]
- Dembowska, E.; Napiórkowski, P.; Mieszczankin, T.; Józefowicz, S. Planktonic indices in the evaluation of the ecological status and the trophic state of the longest lake in Poland. Ecol. Indic. 2015, 56, 15–22. [Google Scholar] [CrossRef]
- Stamou, G.; Katsiapi, M.; Moustaka-Gouni, M.; Michaloudi, E. Trophic state assessment based on zooplankton communities in Mediterranean lakes. Hydrobiologia 2019, 844, 83–103. [Google Scholar] [CrossRef]
- Duggan, I. The ecology of periphytic rotifers. Hydrobiologia 2001, 446/447, 139–148. [Google Scholar] [CrossRef]
- Rodgers, K.H.; Breen, C.M. An investigation of macrophyte, epiphyte and grazer interactions. In Periphyton of Freshwater Ecosystems; Wetzel, R.G., Ed.; Dr W. Junk Publishers: The Hague, The Netherlands, 1983; pp. 217–226. [Google Scholar]
- Jones, J.I.; Young, J.O.; Haynes, G.M.; Moss, B.; Eaton, J.W.; Hardwick, K.J. Do submerged aquatic plants influence their periphyton to enhance the growth and reproduction of invertebrate mutualists? Oecologia 1999, 120, 463–474. [Google Scholar] [CrossRef]
- Kuczyńska-Kippen, N.; Špoljar, M.; Mleczek, M.; Zhang, C. Elodeids, but not helophytes, increase community diversity and reduce trophic state: Case study with rotifer indices in field ponds. Ecol. Indic. 2021, 128, 107829. [Google Scholar] [CrossRef]
- Illies, J. Limnofauna Europaea. A Checklist of the Animals Inhabiting European Inland Waters, with Account of Their Distribution and Ecology; Second revised and enlarged edition; G. Fischer, Stuttgart Swets en Zeitlinger: Stuttgart, Germany; New York, NY, USA; Amsterdam, The Netherlands, 1978. [Google Scholar]
- Ericsson, B.; Hallmans, B. Treatment and disposal of saline wastewater from coal mines in Poland. Desalination 1994, 98, 239–248. [Google Scholar] [CrossRef]
- Harat, A.; Grmela, A. Impact of mine water from the Upper Silesian Coal Basin areas on change quality of water in Olza River in years 2000–2007. Nat. Environ. Monit. 2008, 9, 57–62. [Google Scholar]
- Harding, J.S.; Young, R.G.; Hayes, J.W.; Shearer, K.A.; Stark, J.D. Changes in agricultural intensity and river health along a river continuum. Freshw. Biol. 1999, 42, 345–357. [Google Scholar] [CrossRef]
- Lewin, I. The gastropod communities in the lowland rivers of agricultural areas—Their biodiversity and bioindicative value in the Ciechanowska Upland, Central Poland. Malacologia 2006, 49, 7–23. [Google Scholar] [CrossRef]
- Baldwin, A.H.; Jensen, K.; Schönfeldt, M. Warming increases plant biomass and reduces diversity across continents, latitudes, and species migration scenarios in experimental wetland communities. Glob. Chang. Biol. 2013, 20, 835–850. [Google Scholar] [CrossRef]
- Drake, B.G. Rising sea level, temperature, and precipitation impact plant and ecosystem responses to elevated CO2 on a Chesapeake Bay wetland: Review of a 28-year study. Glob. Chang. Biol. 2014, 20, 3329–3343. [Google Scholar] [CrossRef]
- Liu, Q.P.; Giesy, J.P.; Li, Z.H. Spatio-Temporal Distributions and Environmental Safety Threshold of Cropland Fertilization of Jiangsu Province, China. Adv. Mater. Res. 2014, 962-965, 2110–2115. [Google Scholar] [CrossRef]
- Hautier, Y.; Seabloom, E.; Borer, E.; Adler, P.B.; Harpole, W.S.; Hillebrand, H.; Lind, E.; MacDougall, A.S.; Stevens, C.; Bakker, J.D.; et al. Eutrophication weakens stabilizing effects of diversity in natural grasslands. Nature 2014, 508, 521–525. [Google Scholar] [CrossRef]
- Maxwell, S.L.; Fuller, R.A.; Brooks, T.M.; Watson, J.E.M. Biodiversity: The ravages of guns, nets and bulldozers. Nature 2016, 546, 143–145. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.J.; Greaves, H.M.; Sayer, C.D.; Hassall, C.; Milin, M.; Milner, V.S.; Marazzi, L.; Hall, R.; Harper, L.R.; Thornhill, I.; et al. Pond ecology and conservation: Research priorities and knowledge gaps. Ecosphere 2021, 12, e03853. [Google Scholar] [CrossRef]
- Directive, Water Framework. Directive 2000/60/EC of the European Parliament and of the Council of 23 October 2000 establishing a framework for Community action in the field of water policy. Off. J. Eur. Communities 2000, L327, 1–72. [Google Scholar]
- Piscart, C.; Moreteau, J.-C.; Beisel, J.-N. Monitoring Changes in Freshwater Macroinvertebrate Communities Along a Salinity Gradient Using Artificial Substrates. Environ. Monit. Assess. 2006, 116, 529–542. [Google Scholar] [CrossRef]
- Hermanowicz, W.; Dojlido, J.; Dożańska, W.; Koziorowski, B.; Zerbe, J. Fizyczno-Chemiczne Badanie Wody i Ścieków; Arkady: Warszawa, Poland, 1999. [Google Scholar]
- Chindah, A.C. Responses of periphyton community to salinity gradient in tropical estuary, Niger Delta. Pol. J. Ecol. 2004, 52, 83–89. [Google Scholar]
- Tarkowska-Kukuryk, M. Periphytic algae as food source for grazing chironomids in a shallow phytoplankton-dominated lake. Limnologica 2013, 43, 254–264. [Google Scholar] [CrossRef]
- Bielańska-Grajner, I.; Ejsmont-Karabin, J.; Radwan, S. Rotifers. In Rotifera Monogononta; Łódź University Press & Jagiellonian University Press: Łódź–Kraków, Poland, 2015. [Google Scholar]
- Bielańska-Grajner, I.; Ejsmont-Karabin, J.; Iakovenko, N. Wrotki. In Rotifera Bdelloidea; Wydawnictwo Uniwersytetu Łódzkiego: Łódź, Poland, 2014. [Google Scholar]
- Donner, P.J. Ordnung Bdelloidea (Rotatoria, Rädertiere); Bestimmungsbücher zur Bodenfauna Europas, 6; Akademie: Berlin, Germany, 1965. [Google Scholar]
- Nogrady, T.; Pourriot, R.; Segers, H. Rotifera. In Vol. 3: The Notommatidae and the Scaridiidae. Guides to the Identification of the Microinvertebrates of the Continental Waters of the World. 8 SPB; Academic Publishing: Amsterdam, The Netherlands, 1995. [Google Scholar]
- Segers, H. Rotifera Volume 2. The Lecanidae. (Monogononta). Guides to the Identification of the Microinvertebrates of the Continental Waters of the World 6; Academic Publishing: Amsterdam, The Netherlands, 1995. [Google Scholar]
- Kutikova, L.A. The Bdelloid Rotifers of the Fauna of Russia. Proc. Zool. Inst. 2005, 305, 314. [Google Scholar]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The River Continuum Concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Allan, J.D.; Castillo, M.M. Stream Ecology; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Shen, J.; Qin, G.; Yu, R.; Zhao, Y.; Yang, J.; An, S.; Liu, R.; Leng, X.; Wan, Y. Urbanization has changed the distribution pattern of zooplankton species diversity and the structure of functional groups. Ecol. Indic. 2021, 120, 106944. [Google Scholar] [CrossRef]
- Gromova, Y.F. The Structure and Composition of Pelagic Communities of the Estuarine Region of the Vita River. Hydrobiol. J. 2001, 37, 11. [Google Scholar] [CrossRef]
- Halabowski, D.; Bielańska-Grajner, I.; Lewin, I. Effect of underground salty mine water on the rotifer communities in the Bolina River (Upper Silesia, Southern Poland). Knowl. Manag. Aquat. Ecosyst. 2019, 420, 31. [Google Scholar] [CrossRef]
- Halabowski, D.; Lewin, I.; Buczyński, P.; Krodkiewska, M.; Płaska, W.; Sowa, A.; Buczyńska, E. Impact of the Discharge of Salinised Coal Mine Waters on the Structure of the Macroinvertebrate Communities in an Urban River (Central Europe). Water Air Soil Pollut. 2020, 231, 5. [Google Scholar] [CrossRef]
- Neschuk, N.; Claps, M.; Gabellone, N. Planktonic rotifers of a saline-lowland river: The Salado River (Argentina). Ann. de Limnol.-Int. J. Limnol. 2002, 38, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Korstad, J.; Olsen, Y.; Vadstein, O. Life history characteristics of Brachionus plicatilis (rotifera) fed different algae. Hydrobiologia 1989, 186-187, 43–50. [Google Scholar] [CrossRef]
- Sarma, S.S.S.; Nandini, S. Comparative population dynamics of six brachionid rotifers (Rotifera) fed seston from a hypertrophic, high altitude shallow waterbody from Mexico. Hydrobiologia 2019, 844, 55–65. [Google Scholar] [CrossRef]
- Zhao, J.; Ren, W.; Dai, Y.; Liu, L.; Wang, Z.; Yu, X.; Zhang, J.; Wang, X.; Xing, B. Uptake, Distribution, and Transformation of CuO NPs in a Floating Plant Eichhornia crassipes and Related Stomatal Responses. Environ. Sci. Technol. 2017, 51, 7686–7695. [Google Scholar] [CrossRef]
- Le Coz, M.; Chambord, S.; Souissi, S.; Meire, P.; Ovaert, J.; Buffan-Dubau, E.; Prygiel, J.; Azémar, F.; Sossou, A.; Lamothe, S.; et al. Are zooplankton communities structured by taxa ecological niches or by hydrological features? Ecohydrology 2018, 11, e1956. [Google Scholar] [CrossRef]
- Cimdinš, P.; Druvietis, I.; Liepa, R.; Parele, E.; Urtane, L.; Urtans, A. Latvian catalogue of indicator species of freshwater saprobity. Proc. Latv. Acad. Sci. 1995, 1, 122–133. [Google Scholar]
- Marneffe, Y.; Comblin, S.; Thomé, J.-P. Ecological water quality assessment of the Bütgenbach lake (Belgium) and its impact on the River Warche using rotifers as bioindicators. In Rotifera VIII: A Comparative Approach; Springer: Berlin/Heidelberg, Germany, 1998; Volume 134, pp. 459–467. [Google Scholar]
- Zhou, S.; Tang, T.; Wu, N.; Fu, X.; Cai, Q. Impacts of a Small Dam on Riverine Zooplankton. Int. Rev. Hydrobiol. 2008, 93, 297–311. [Google Scholar] [CrossRef]
- Gromova, Y.F.; Afanasyev, S.A.; Shevtsova, L.V. Structural Organization of Zooplankton in Transformed Small Rivers. Hydrobiol. J. 2013, 49, 21–29. [Google Scholar] [CrossRef]
- Xiong, W.; Li, J.; Chen, Y.; Shan, B.; Wang, W.; Zhan, A. Determinants of community structure of zooplankton in heavily polluted river ecosystems. Sci. Rep. 2016, 6, 22043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czerniawski, R.; Sługocki, Ł. A comparison of the effect of beaver and human-made impoundments on stream zooplankton. Ecohydrology 2018, 11, e1963. [Google Scholar] [CrossRef]
- Senkevich, V.A.; Stojko, T.G.; Tsyganov, A. Species Composition and Community Structure of Zooplankton in Small Rivers of the Forest-Steppe Zone. Inland Water Biol. 2018, 11, 448–455. [Google Scholar] [CrossRef]
- Nandini, S.; Ramírez-García, P.; Sarma, S.S.S.; Gutierrez-Ochoa, R.A. Planktonic indicators of water quality: A case study in the Amacuzac River Basin (State of Morelos, Mexico). River Res. Appl. 2019, 35, 268–279. [Google Scholar] [CrossRef]
- Xiong, W.; Ni, P.; Chen, Y.; Gao, Y.; Li, S.; Zhan, A. Biological consequences of environmental pollution in running water ecosystems: A case study in zooplankton. Environ. Pollut. 2019, 252, 1483–1490. [Google Scholar] [CrossRef]
- Sindt, A.R.; Wolf, M.C. Spatial and temporal trends of Minnesota River phytoplankton and zooplankton. River Res. Appl. 2021, 37, 776–795. [Google Scholar] [CrossRef]
- Chang, K.-H.; Doi, H.; Imai, H.; Gunji, F.; Nakano, S.-I. Longitudinal changes in zooplankton distribution below a reservoir outfall with reference to river planktivory. Limnology 2008, 9, 125–133. [Google Scholar] [CrossRef]
- Czerniawski, R.; Sługocki, Ł.; Kowalska-Góralska, M. Diurnal Changes of Zooplankton Community Reduction Rate at Lake Outlets and Related Environmental Factors. PLoS ONE 2016, 11, e0158837. [Google Scholar] [CrossRef] [Green Version]
- Sługocki, Ł.; Czerniawski, R.; Kowalska-Góralska, M.; Senze, M.; Reis, A.; Carrola, J.S.; Teixeira, C.A. The Impact of Land Use Transformations on Zooplankton Communities in a Small Mountain River (The Corgo River, Northern Portugal). Int. J. Environ. Res. Public Health 2018, 16, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duggan, I.C.; Green, J.D.; Shiel, R.J. Distribution of rotifers in North Island, New Zealand, and their potential use as bioindicators of lake trophic state. Rotifera IX 2001, 446, 155–164. [Google Scholar] [CrossRef]
- Kuczyńska-Kippen, N. Seasonal changes of the rotifer community in the littoral of a polymictic lake. Int. Ver. Theor. Angew. Limnol. Verh. 2000, 27, 2964–2967. [Google Scholar] [CrossRef]
- Liang, D.; Wang, Q.; Wei, N.; Tang, C.; Sun, X.; Yang, Y. Biological indicators of ecological quality in typical urban river-lake ecosystems: The planktonic rotifer community and its response to environmental factors. Ecol. Indic. 2020, 112, 106127. [Google Scholar] [CrossRef]
- Pakulnicka, J.; Zawal, A. Effect of changes in the fractal structure of a littoral zone in the course of lake succession on the abundance, body size sequence and biomass of beetles. PeerJ 2018, 6, e5662. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.-Y.; Kim, S.-K. Responses of Rotifer Community to Microhabitat Changes Caused by Summer-Concentrated Rainfall in a Shallow Reservoir, South Korea. Diversity 2020, 12, 113. [Google Scholar] [CrossRef] [Green Version]
- Kuczyńska-Kippen, N. Habitat choice in rotifera communities of three shallow lakes: Impact of macrophyte substratum and season. Hydrobiologia 2007, 593, 27–37. [Google Scholar] [CrossRef]
- Bolduc, P.; Bertolo, A.; Hudon, C.; Pinel-Alloul, B. Submerged aquatic vegetation cover and complexity drive crustacean zooplankton community structure in a large fluvial lake: An in situ approach. J. Great Lakes Res. 2020, 46, 767–779. [Google Scholar] [CrossRef]
- Deosti, S.; Bomfim, F.D.F.; Lansac-Tôha, F.M.; Quirino, B.A.; Bonecker, C.C.; Lansac-Tôha, F.A. Zooplankton taxonomic and functional structure is determined by macrophytes and fish predation in a Neotropical river. Hydrobiologia 2021, 848, 1475–1490. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, H.; Gao, X.; Zhang, H. Modeling nutrients, oxygen and critical phosphorus loading in a shallow reservoir in China with a coupled water quality—Macrophytes model. Ecol. Indic. 2016, 66, 212–219. [Google Scholar] [CrossRef]
- Špoljar, M.; Zhang, C.; Dražina, T.; Zhao, G.; Lajtner, J.; Radonić, G. Development of submerged macrophyte and epiphyton in a flow-through system: Assessment and modelling predictions in interconnected reservoirs. Ecol. Indic. 2017, 75, 145–154. [Google Scholar] [CrossRef]
- Lucks, R. Über ein neues Rädertier. Zool. Anz. 1911, 38, 568–571. [Google Scholar]
- Lucks, R. Zur Rotatorienfauna Westpreußen; Verlag und Offsetdruck von A. W. Kafemann G.m.b.H.: Gdańsk, Poland, 1912. [Google Scholar]
- Rotifer World Catalog. Available online: http://www.rotifera.hausdernatur.at/Species/Index/1451#TabStripSpecies-7 (accessed on 19 December 2021).
Figure 1.
Photos of the sampling sites. (a) upper course of the Bolina River, (b) lower course of the Bolina River, (c) upper course of the Centuria River, (d) lower course of the Centuria River, (e) upper course of the Mitręga River, (f) lower course of the Mitręga River, (g) upper course of the Mleczna River, (h) lower course of the Mleczna River, (i) upper course of the Dziechcinka River, (j) lower course of the Dziechcinka River, (k) upper course of the Vistula River, (l) lower course of the Vistula River, (m) upper course of the Korzenica River, (n) lower course of the Korzenica River, (o) upper course of the Wiercica River, (p) lower course of the Wiercica River.
Figure 1.
Photos of the sampling sites. (a) upper course of the Bolina River, (b) lower course of the Bolina River, (c) upper course of the Centuria River, (d) lower course of the Centuria River, (e) upper course of the Mitręga River, (f) lower course of the Mitręga River, (g) upper course of the Mleczna River, (h) lower course of the Mleczna River, (i) upper course of the Dziechcinka River, (j) lower course of the Dziechcinka River, (k) upper course of the Vistula River, (l) lower course of the Vistula River, (m) upper course of the Korzenica River, (n) lower course of the Korzenica River, (o) upper course of the Wiercica River, (p) lower course of the Wiercica River.

Figure 2.
Box-and-whisker plot showing the number of rotifer taxa in the abiotic types of rivers, in the rivers and at the sampling sites (asterisks above a whisker denote significant differences between the rivers, p < 0.05). Abbreviations as in Table 1.
Figure 2.
Box-and-whisker plot showing the number of rotifer taxa in the abiotic types of rivers, in the rivers and at the sampling sites (asterisks above a whisker denote significant differences between the rivers, p < 0.05). Abbreviations as in Table 1.

Figure 3.
Box-and-whisker plot showing species richness in the different microhabitats (asterisks above a whisker denote significant differences among the rivers, p < 0.05). Abbreviations: AM—Amphibious; DI—Diatoms; EB—Emergent broad-leaved; ER—Emergent reeds, sedges; FA—Filamentous algae; FL—Floating-leaved (rooted); FF—Free-floating; LM—Liverworts and mosses; SM—Submerged broad-leaved; SL—Submerged linear-leave.
Figure 3.
Box-and-whisker plot showing species richness in the different microhabitats (asterisks above a whisker denote significant differences among the rivers, p < 0.05). Abbreviations: AM—Amphibious; DI—Diatoms; EB—Emergent broad-leaved; ER—Emergent reeds, sedges; FA—Filamentous algae; FL—Floating-leaved (rooted); FF—Free-floating; LM—Liverworts and mosses; SM—Submerged broad-leaved; SL—Submerged linear-leave.


{kind=link}
{kind=link}
{kind=link}
Table 1.
General characteristics of the studied sampling sites. Abbreviations: BOL—the Bolina River, CEN—the Centuria River, MIT—the Mitręga River, MLE—the Mleczna River, DZI—the Dziechcinka River, VIS—the Vistula River, KOR—the Korzenica River, WIE—the Wiercica River.
Table 1.
General characteristics of the studied sampling sites. Abbreviations: BOL—the Bolina River, CEN—the Centuria River, MIT—the Mitręga River, MLE—the Mleczna River, DZI—the Dziechcinka River, VIS—the Vistula River, KOR—the Korzenica River, WIE—the Wiercica River.
| River | Sampling Site | Geographical Coordinates | Ecoregion | Type of River/Geology | Catchment Land Use | Main Anthropogenic Pressure | Bottom Sediments |
|---|---|---|---|---|---|---|---|
| BOL | Upper | N: 50°13.793; E: 19°05.142 | Ecoregion 14—Central Plains | Type 5/mid-altitude siliceous streams with a fine particulate substratum | Industrial and urban, grassland | Salinisation (coal mine), industrial and communal sewage, regulation of the riverbed | Silty |
| Lower | N: 50°14.742; E: 19°06.078 | Silty-sandy | |||||
| CEN | Upper | N: 50°24.879; E: 19°29.190 | Natural monument and Natura 2000; Woodland | None | Sandy-silty | ||
| Lower | N: 50°21.920; E: 19°29.682 | Natura 2000; Woodland and grassland | Organic pollution (agriculture, animal grazing), fishponds | Sandy-stony | |||
| MIT | Upper | N: 50°24.797; E: 19°22.779 | Ecoregion 14—Central Plains | Type 6/mid-altitude calcareous streams with a fine particulate substratum on loess | Built-up area and grassland | Dam reservoir, communal sewage | Sandy-silty |
| Lower | N: 50°26.070; E: 19°17.956 | Dam reservoir, communal sewage, regulation of the riverbed | Sandy-silty | ||||
| MLE | Upper | N: 52°09.754; E: 19°00.213 | Industrial and urban, grassland | Industrial and communal sewage, regulation of the riverbed | Sandy-silty | ||
| Lower | N: 50°07.018; E: 19°04.487 | Salinisation (coal mine), industrial and communal sewage, regulation of the riverbed | Silty-sandy | ||||
| DZI | Upper | N: 49°38.021; E: 18°50.828 | Ecoregion 10—Carpathians | Type 12/flysch streams | Woodland, road | None | Stony-gravel |
| Lower | N: 49°38.789; E: 18°52.025 | Built-up area, woodland | Regulation of the riverbed | Stony-concrete | |||
| VIS | Upper | N: 49°37.190; E: 18°59.160 | Nature reserve and Natura 2000, woodlands, built-up area | None | Stony-gravel | ||
| Lower | N: 49°38.728; E: 18°51.167 | Built-up area, woodland | Dam reservoir, communal sewage, regulation of the riverbed | Stony-gravel | |||
| KOR | Upper | N: 50°03.509; E:18°56.804 | Ecoregion 14—Central Plains | Type 17/lowland sandy streams | Built-up area and grassland | Fishponds and agriculture, communal sewage | Silty-sandy |
| Lower | N: 50°01.850; E: 19°05.839 | Built-up area and grassland, protected areas: Natura 2000 | Fishponds and agriculture | Sandy-stony | |||
| WIE | Upper | N: 50°41.117; E: 19°24.472 | Nature reserve and Natura 2000, woodlands and grassland | None | Sandy-stony | ||
| Lower | N: 50°52.471; E: 19°26.133 | Woodlands, grassland and built-up area | Agriculture, animal grazing, dam reservoirs | Sandy-silty |
Table 2.
The physical and chemical parameters of the waters of the abiotic type of rivers (ranges) and the results of the Kruskal–Wallis one-way ANOVA and Dunn’s multiple comparison post hoc tests (superscript a, b, c, d denote significant differences between the rivers).
Table 2.
The physical and chemical parameters of the waters of the abiotic type of rivers (ranges) and the results of the Kruskal–Wallis one-way ANOVA and Dunn’s multiple comparison post hoc tests (superscript a, b, c, d denote significant differences between the rivers).
| Parameter | Type 5 | Type 6 | Type 12 | Type 17 | H Value | p Value |
|---|---|---|---|---|---|---|
| Altitude [m a.s.l.] | 257–343 c,d | 236–317 c | 415–748 a,b,d | 215–309 a,c | 44.007 | <0.001 |
| Width of the riverbed [m] | 3.30–7.78 | 2.87–9.36 | 3.47–19.80 | 1.85–12.05 | 0.500 | 0.919 |
| Depth of the riverbed [cm] | 9.75–58.60 b | 36.80–109.17 a,c | 19.30–57.60 b | 6.70–98.33 | 18.904 | <0.001 |
| Flow velocity [m s−1] | 0.060–0.790 b | 0.007–0.384 a,c | 0.107–0.939 b,d | 0.057–0.706 c | 22.633 | <0.001 |
| Dissolved oxygen [mg dm−3] | 4.24–9.69 b | 0.69–6.78 a,c | 4.88–5.90 b | 2.98–6.49 | 11.609 | 0.009 |
| Temperature [°C] | 7.5–29.1 | 9.4–25.1 | 9.1–23.8 | 9.7–23.5 | 7.797 | 0.050 |
| Salinity [PSU] | 0.19–33.55 | 0.28–5.16 | 0.02–0.06 | 0.17–0.28 | 45.479 | <0.001 |
| EC [μS cm−1] | 250–46 600 c | 360–7160 c,d | 30–90 a,c,d | 220–370 b,c | 45.479 | <0.001 |
| TDS [mg dm−3] | 110–23 300 c | 170–3570 c,d | 10–30 a,b,d | 100–170 b,c | 45.881 | <0.001 |
| Chlorides [mg dm−3] | 8–17 028 c | 15–1970 c | 4–9 a,b,d | 4–25 c | 36.478 | <0.001 |
| Sulphates [mg dm−3] | 35–770 c,d | 22–272 c,d | 8–18 a,b | 10–64 a,b | 39.919 | <0.001 |
| Total hardness [mg CaCO3 dm−3] | 160.00–4857.92 c,d | 160.00–560.00 c,d | 28.00–68.00 a,b,d | 110–320 a,b,c | 47.971 | <0.001 |
| Magnesium [mg dm−3] | 1.94–670.00 c,d | 0.06–62.53 c | 0.04–5.14 a,b | 0.00–13.80 a | 32.260 | <0.001 |
| Calcium [mg dm−3] | 55–1310 c,d | 40–158 c | 10–21 a,b,d | 24–82 a,c | 45.340 | <0.001 |
| Alkalinity [mg CaCO3 dm−3] | 75.0–380.0 c | 125.0–275.0 c | 2.5–50.0 a,b,d | 20.0–180.0 c | 39.516 | <0.001 |
| pH | 7.2–7.9 | 6.8–8.1 | 6.5–8.4 | 6.2–8.2 | 2.006 | 0.571 |
| Nitrates [mg dm−3] | 0.00–79.74 | 0.89–15.95 c | 0.00–9.30 b,d | 0.44–18.61 c | 13.726 | <0.001 |
| Nitrites [mg dm−3] | 0.00–9.96 c | 0.03–093 c | 0.00–0.01 a,b | 0.00–0.59 | 30.897 | <0.001 |
| Ammonium [mg dm−3] | 0.00–12.12 c | 0.23–1.42 c | 0.13–0.45 a,b | 0.00–0.63 | 18.045 | <0.001 |
| Phosphates [mg dm−3] | 0.00–0.14 b,d | 0.08–19.20 a,c | 0.00–1.52 b | 0.00–0.87 a | 39.919 | <0.001 |
| Iron [mg dm−3] | 0.03–0.88 b | 0.25–1.46 a,c | 0.03–0.34 b,d | 0.03–3.11 c | 27.324 | <0.001 |
a Type 5, b Type 6, c Type 12, d Type 17.
Table 3.
Summary of the identified rotifers relative to the various microhabitats and seasons. Abbreviations: BOLU—upper course of the Bolina River, BOLL—lower course of the Bolina River, CENU—upper course of the Centuria River, CENL—lower course of the Centuria River, MITU—upper course of the Mitręga River, MITL—lower course of the Mitręga River, MLEU—upper course of the Mleczna River, MLEL—lower course of the Mleczna River, DZIU—upper course of the Dziechcinka River, DZIL—lower course of the Dziechcinka River, VISU—upper course of the Vistula River, VISL—lower course of the Vistula River, KORU—upper course of the Korzenica River, KORL—lower course of the Korzenica River, WIEU—upper course of the Wiercica River, WIEL—lower course of the Wiercica River, SP—Spring, SU—Summer, AU—Autumn, s.l.—sensu lato.
Table 3.
Summary of the identified rotifers relative to the various microhabitats and seasons. Abbreviations: BOLU—upper course of the Bolina River, BOLL—lower course of the Bolina River, CENU—upper course of the Centuria River, CENL—lower course of the Centuria River, MITU—upper course of the Mitręga River, MITL—lower course of the Mitręga River, MLEU—upper course of the Mleczna River, MLEL—lower course of the Mleczna River, DZIU—upper course of the Dziechcinka River, DZIL—lower course of the Dziechcinka River, VISU—upper course of the Vistula River, VISL—lower course of the Vistula River, KORU—upper course of the Korzenica River, KORL—lower course of the Korzenica River, WIEU—upper course of the Wiercica River, WIEL—lower course of the Wiercica River, SP—Spring, SU—Summer, AU—Autumn, s.l.—sensu lato.
| Taxon | Sampling Site | Microhabitat | Season |
|---|---|---|---|
| Adineta gracilis Janson, 1893 | DZIL, VISU | Chiloscyphus polyanthos, Fontinalis antipyretica, Platyhypnidium riparioides, Scapania undulata | SP, AU |
| Adineta vaga (Davis, 1873) | VISU | Hygrohypnum luridum, Scapania undulata | SU |
| Adineta vaga major Bryce, 1893 | WIEL | Phalaris arundinacea | SU |
| Anuraeopsis fissa Gosse, 1851 | MITU | open water | SU |
| Asplanchna priodonta Gosse, 1850 | MITU | open water | SU |
| Bdelloidea non determinata | CENL, DZIL, DZIL, DZIU, KORL, KORU, MITL, MITU, MLEU, WIEL, WIEU, VISU | open water, stones, bottom sediments, diatom aggregation, Berula erecta, Callitriche sp., Chiloscyphus polyanthos, Elodea canadensis, Fontinalis antipyretica, Glyceria maxima, Myosotis palustris, Phalaris arundinacea, Platyhypnidium riparioides, Potamogeton crispus, P. natans, Ranunculus aquatile, R. circinatum, Sagittaria sagittifolia, Scrophularia umbrosa, Sparganium erectum, Thamnobryum alopecurum, Veronica beccabunga | SP, SU, AU |
| Brachionus angularis Gosse, 1851 | BOLL, MITU | open water | SU |
| Brachionus plicatilis s.l. Müller, 1786 | BOLL, BOLU | open water | SU, AU |
| Brachionus quadridentatus Hermann, 1783 | MITL | diatom aggregation | SU |
| Brachionus rubens (Ehrenberg, 1838) | BOLL, BOLU | open water, Phragmites australis | SP |
| Brachionus species non determinata | BOLL | diatom aggregation | AU |
| Cephalodella auriculata (Müller, 1773) | CENU, KORL, KORU, MITL, MITU, MLEU, WIEL, VISL | open water, stones, bottom sediments, diatom aggregation, Berula erecta, Glyceria nemoralis, Phalaris arundinacea, Polygoum hydropiper, Potamogeton crispus, P. pectinatus, Rorippa amphibia, Sagittaria sagittifolia | SP, SU, AU |
| Cephalodella catellina (Müller, 1786) | KORU, MITL | open water, diatom aggregation, Callitriche sp., Potamogeton crispus | SP, SU, AU |
| Cephalodella delicata Wulfert, 1937 | CENU | Carex rostrata | SU |
| Cephalodella eva (Gosse, 1887) | KORL, KORU, MITU, WIEL, VISL | Callitriche sp., Elodea canadensis, Potamogeton crispus, Rorippa amphibia, Spraganium erectum | SU, AU |
| Cephalodella forficula (Ehrenberg, 1830) | KORL, MITU | Berula erecta, Fontinalis antipyretica | SU, AU |
| Cephalodella gibba (Ehrenberg, 1830) | KORL, KORU, MITL, MITU, MLEU, WIEL, WIEU, VISL | open water, stones, bottom sediments, diatom aggregation, Callitriche sp., Fontinalis antipyretica, Glyceria maxima, Lemna minor, Phalaris arundinacea, Polygonum hydropiper, Potamogeton crispus, Sparganium emersum, S. erectum | SP, SU, AU |
| Cephalodella globata (Gosse, 1887) | MITL, WIEU | open water, bottom sediments | SU, AU |
| Cephalodella gracilis (Ehrenberg, 1830) | CENL, CENU, KORU, MITU, MLEL, MLEU, WIEL | open water, Berula erecta, Carex rostrata, Phalaris arundinacea, Potamogeton natans, P. pectinatus, Rorippa amphibia, Sparganium erectum | SP, SU, AU |
| Cephalodella hoodii (Gosse, 1886) | MLEU | stones | AU |
| Cephalodella megalocephala (Glascott, 1893) | VISL | Elodea canadensis | AU |
| Cephalodella megalotrocha Wiszniewski, 1934 | CENU | Carex rostrata | SU |
| Cephalodella misgurnus Wulfert, 1937 | KORL | Sparganium erectum | AU |
| Cephalodella nana Myers, 1924 | KORL | open water | AU |
| Cephalodella species non determinata | KOLR, WIEL | open water, Potamogeton natans | SP, SU |
| Cephalodella stenroosi Wulfert, 1937 | MITU | Spraganium erectum | SU |
| Cephalodella ventripes (Dixon-Nuttall, 1901) | KORU | open water | SP |
| Collotheca species non determinata | KORL, MITU, VISL | Elodea canadensis, Fontinalis antipyretica, Phalaris arundinacea | SP, SU, AU |
| Colurella adriatica Ehrenberg, 1831 | BOLL, BOLU, CENL, CENU, DZIL, DZIU, KORL, MITL, MLEL, MLEU, WIEU, VISL, VISU | open water, stones, bottom sediments, diatom aggregation, Carex rostrata, Elodea canadensis, Enteromorpha sp., Fontinalis antipyretica, Glyceria nemoralis, Mougeotia sp., Phalaris arundinacea, Phragmites australis, Platyhypnidium riparioides, Veronica beccabunga | SP, SU, AU |
| Colurella colurus (Ehrenberg, 1830) | BOLL, CENU, DZIL, KORL, KORU, MLEL, WIEL, VISL, VISU | open water, Callitriche sp., Elodea canadensis, Enteromorpha sp., Glyceria maxima, G. nemoralis, Nuphar lutea, Phalaris arundinacea, Scirpus sylvaticus | SU, AU |
| Colurella species non determinata | CENL | open water | SU |
| Colurella uncinata (Müller, 1773) | KORL, KORU, MITL, MITU, MLEL, MLEU | open water, diatom aggregation, Fontinalis antipyretica, Phalaris arundinacea, Potamogeton pectinatus, Ranunculus aquatile, Sparganium emersum, S. erectum | SP, SU, AU |
| Dicranophorus forcipatus (Müller, 1786) | MITU, WIEL, WIEU | stones, Elodea canadensis, Rorippa amphibia, Thamnobryum alopecurum | AU |
| Dicranophorus grandis (Ehrenberg, 1832) | KORU | Potamogeton crispus | SU |
| Dicranophorus hercules Wiszniewski, 1932 | DZIU, KORU, MITU, WIEU | open water, stones, bottom sediments, Sparganium erectum | SP, SU, AU |
| Dicranophorus rostratus (Dixon-Nuttall & Freeman, 1902) | DZIU | bottom sediments | SU |
| Dicranophorus secretus Donner, 1951 | MITU | Spraganium erectum | SU |
| Dicranophorus species non determinata | KORU, MITU | open water, Berula erecta, Sparganium erectum | SU |
| Dissotrocha macrostyla (Ehrenberg, 1838) | KORL, MLEU | stones, Callitriche sp. | AU |
| Dissotrocha species non determinata | CENU | bottom sediments | AU |
| Encentrum diglandula (Zawadovsky, 1926) | BOLU | Enteromorpha sp. | AU |
| Encentrum lupus Wulfert, 1936 | MITL | Fontinalis antipyretica | AU |
| Encentrum marinum (Dujardin, 1841) | BOLL, BOLU, DZIU, KORL, KORU, MLEL | open water, stones, bottom sediments, diatom aggregation, Enteromorpha sp., Phragmites australis | SP, SU, AU |
| Encentrum mustela (Milne, 1885) | WIEL | Glyceria maxima | SP |
| Encentrum saundersiae (Hudson, 1885) | KORU | open water | SP |
| Encentrum species non determinata | CENL | Berula erecta | SU |
| Encentrum tyrphos Wulfert, 1936 | KORL, WIEL | open water, Phalaris arundinacea | SU, AU |
| Erignatha clastopis (Gosse, 1886) | KORL | Fontinalis antipyretica | SU |
| Erignatha species non determinata | KORL | open water | SU |
| Euchlanis deflexa (Gosse, 1851) | CENU, MITL, MLEU | Glyceria nemoralis, Phalaris arundinacea, Polygonum hydropiper | AU |
| Euchlanis dilatata Ehrenberg, 1832 | KORL, KORU, MITL, MLEU, WIEL | open water, stones, diatom aggregation, Callitriche sp., Glyceria maxima, Mougeotia sp., Phalaris arundinacea, Potamogeton crispus, P. natans, Ranunculus aquatile, Sparganium erectum | SP, SU, AU |
| Euchlanis species non determinata | KORL, KORU, WIEL, WIEU, VISL | open water, stones, Callitriche sp., Fontinalis antipyretica, Nuphar lutea, Phalaris arundinacea, Potamogeton crispus, Ranunculus aquatile, Thamnobryum alopecurum | SP, SU, AU |
| Filinia longiseta (Ehrenberg, 1834) | MITU, WIEL | open water | SU, AU |
| Floscularia ringens (Linnaeus, 1758) | MLEU | Callitriche sp. | AU |
| Habrotrocha roeperi (Milne, 1889) | DZIL, MITL | Fontinalis antipyretica, Platyhypnidium riparioides, Scrophularia umbrosa | SU, AU |
| Habrotrocha species non determinata | CENL, CENU, DZIL, DZIU, KORL, MITL, MLEU, WIEL, WIEU, VISL, VISU | stones, bottom sediments, Berula erecta, Callitriche sp., Carex rostrata, Fontinalis antipyretica, Hygrohypnum luridum, Nuphar lutea, Phalaris arundinacea, Platyhypnidium riparioides, Ranunculus aquatile, Sparganium erectum, Thamnobryum alopecurum | SP, SU, AU |
| Itura aurita (Ehrenberg, 1830) | KORU | open water | SU |
| Keratella cochlearis (Gosse, 1851) | CENL, KORU, MITL, MITU | open water | SP, SU, AU |
| Keratella quadrata (Müller, 1786) | CENL | open water | SP |
| Keratella tecta (Gosse, 1851) | CENL, MITL | open water | SP, SU |
| Lecane bulla (Gosse, 1851) | MLEU | open water | SU |
| Lecane closterocerca (Schmarda, 1859) | CENU, KORL, KORU, MITL, MITU, MLEL, MLEU, WIEL | open water, stones, diatom aggregation, Callitriche sp., Carex rostrata, Fontinalis antipyretica, Glyceria maxima, Mougeotia sp., Potamogeton crispus, P. natans, P. pectinatus, Ranunculus aquatile, Rorippa amphibia, Sparganium erectum | SP, SU, AU |
| Lecane hamata (Stokes, 1896) | WIEU | Thamnobryum alopecurum | AU |
| Lecane inermis (Bryce, 1892) | KORU, MITU, MLEL, MLEU, VISL | open water, stones, Potamogeton crispus, Spraganium erectum | SU, AU |
| Lecane luna (Müller, 1776) | KORL, MITL | open water, Callitriche sp., Phalaris arundinacea | SP, SU, AU |
| Lecane lunaris (Ehrenberg, 1832) | MITL, MLEL, MLEU | open water, bottom sediments, Fontinalis antipyretica, Phragmites australis, Polygonum hydropiper | SU, AU |
| Lecane scutata (Harring & Myers, 1926) | KORL | diatom aggregation | SU |
| Lecane species non detereminata | KORL | Sparganium emersum | SU |
| Lepadella (Lepadella) acuminata (Ehrenberg, 1834) | MITU, MLEU, WIEL | open water, Sparganium erectum | SU, AU |
| Lepadella (Lepadella) elliptica Wulfert, 1939 | WIEL | Glyceria maxima | SU |
| Lepadella (Lepadella) ovalis (Müller, 1786) | CENL, DZIL, KORU, MITU, MLEU | open water, Platyhypnidium riparioides, Ranunculus aquatile, R. circinatum | SP, SU |
| Lepadella (Lepadella) patella (Müller, 1773) | CENL, KORL, KORU, MITL, MITU, MLEL, MLEU, WIEL, WIEU | open water, diatom aggregation, Berula erecta, Elodea canadensis, Glyceria maxima, Myosotis palustris, Phalaris arundinacea, Potamogeton pectinatus, Ranunculus aquatile, Rorippa amphibia, Sparganium emersum, Sparganium erectum, Thamnobryum alopecurum | SP, SU, AU |
| Lepadella species non determinata | CENL, MITU | Berula erecta | SU |
| Limnias melicerta Weisse, 1848 | MLU | Callitriche sp. | AU |
| Lindia species non determinata | KORL, MITU, WIEL | Phalaris arundinacea, Sagittaria sagittifolia, Sparganium erectum | SU |
| Lindia (Lindia) torulosa Dujardin, 1841 | KORL, MLEU | open water | SP, AU |
| Lindia (Lindia) truncata (Jennings, 1894) | CENU | Glyceria nemoralis | SU |
| Macrotrachela species non determinata | KORL | Phalaris arundinacea | AU |
| Monogononta species non determinata | BOLU, CENU, KORU, MITL, WIEL | open water, bottom sediments, Glyceria nemoralis, Phragmites australis, Potamogeton crispus | SU |
| Monommata species non determinata | KORU, MITL | open water, Potamogeton crispus | SP, SU |
| Mytilina mucronata (Müller, 1773) | MITL | Fontinalis antipyretica | AU |
| Mytilina species non determinata | WIEL | Rorippa amphibia | SU |
| Mytilina ventralis (Ehrenberg, 1830) | WIEL | Phalaris arundinacea | SU, AU |
| Notommata cerberus (Gosse, 1886) | MLEU | Phalaris arundinacea | SU |
| Notommata cyrtopus Gosse, 1886 | CENL, MITL, MITU, WIEL | stones, diatom aggregation, Phalaris arundinacea, Ranunculus circinatus, Sparganium erectum | SU, AU |
| Notommata glyphura Wulfert, 1935 | BOLU, MITU, MELU | open water, stones, Sparganium erectum | SU, AU |
| Notommata groenlandica Bergendal, 1892 | CENU, WIEL | Glyceria nemoralis, Sparganium erectum | AU |
| Notommata species non determinata | MITL | open water, Sparganium erectum | SU |
| Otostephanos donneri Bartoš, 1959 | MLEU | stones | SU |
| Philodina acuticornis Murray, 1902 | CENL, CENU, DZIL, DZIU, KORL, KORU, MITL, MITU, MLEU, WIEL, WIEU, VISD, VISU | stones, diatom aggregation, Berula erecta, Cllitriche sp., Chiloscyphus polyanthos, Elodea canadensis, Fontinalis antipyretica, Glyceria maxima, Hygrohypnum luridum, Myosotis palustris, Phalaris arundinacea, Platyhypnidium riparioides, Polygonum hydropiper, Potamogeton crispus, P. natans, Ranunculus aquatile, R. circinatus, Rorippa amphibia, Scirpus sylvaticus, Sparganium emersum, S. erectum, Thamnobyryum alopecurum, Veronica anagalis-aquatica, V. beccabunga | SP, SU, AU |
| Philodina citrina Ehrenberg, 1832 | MITL | diatom aggregation | AU |
| Philodina flaviceps Bryce, 1906 | WIEU | Veronica beccabunga | SU |
| Philodina species non determinata | KORU | Phalaris arundinacea | SU |
| Philodinavus paradoxus (Murray, 1905) | KORL | stones | AU |
| Pleurotrocha petromyzon (Ehrenberg, 1830) | CENU, KORU, MITL | open water, Fontinalis antipyretica, Glyceria nemoralis, Phalaris arundinacea | SU, AU |
| Polyarthra species non determinata | MITU | open water | SU |
| Polyarthra vulgaris Carlin, 1943 | MITU, WIEL | open water | AU |
| Pompholyx sulcata Hudson, 1885 | KORU, MITU | open water | SU, AU |
| Proales daphnicola Thompson, 1892 | KORU, WIEL | open water, Rorippa amphibia | SP, AU |
| Proales sordida Gosse, 1886 | MITL, MITU, MLEU, VISL | bottom sediments, diatom aggregation, Berula erecta, Callitriche sp., Sparganium erectum | SU, AU |
| Proales species non determinata | WIEL | Glyceria maxima | SU |
| Proales theodora (Gosse, 1887) | KORU, WIEL | open water, stones, Glyceria maxima, Sparganium erectum | SP, SU, AU |
| Proalinopsis squamipes Hauer, 1935 | KORL | open water | AU |
| Rotaria citrina (Ehrenberg, 1838) | WIEL | stones, Potamogeton natans | SU |
| Rotaria macrura (Ehrenberg, 1832) | KORU | stones, Batrachium aquatice | SU |
| Rotaria magnacalcarata (Parsons, 1892) | KORL, MITU | Berula erecta, Fontinalis antipyretica | SP, SU |
| Rotaria rotatoria (Pallas, 1766) | BOLU, CENL, KORL, KORU, MITL, MITU, MLEL, MLEU, WIEU, | open water, stones, bottom sediments, diatom aggregation, Berula erecta, Callitriche sp., Elodea canadensis, Myosotis palustris, Phalaris arundinacea, Phragmites australis, Potamogeton pectinatus, Ranunculus circinatus, Sparganium erectum | SP, SU, AU |
| Rotaria species non determinata | KORL, MITU | Callitriche sp., Elodea canadensis | SP, SU |
| Rotaria tardigrada (Ehrenberg, 1830) | CENL, MLEL, WIEU | bottom sediments, Elodea canadensis, Phragmites australis, Veronica beccabunga | SP, SU, AU |
| Squatinella rostrum (Schmarda, 1846) | MITL | open water | SU |
| Synchaeta oblonga Ehrenberg, 1832 | VISL | open water | SU |
| Synchaeta stylata Wierzejski, 1893 | CENL | open water | SU |
| Synchaeta tremula (Müller, 1786) | KORU | open water | SP |
| Taphrocampa selenura Gosse, 1887 | MITL | Fontinalis antipyretica | SU |
| Testudinella clypeata (Müller, 1786) | MLEL, MLEU | open water, stones, bottom sediments, Phragmites australis, Potamogeton pectinatus | SU, AU |
| Testudinella patina (Hermann, 1783) | KORL, KORU, MLEU | open water, Callitriche sp., Fontinalis antipyretica | SU, AU |
| Trichocerca collaris (Rousselet, 1896) | CENU | bottom sediments | SU |
| Trichocerca cylindrica (Imhof, 1891) | MITU | open water | SU |
| Trichocerca intermedia (Stenroos, 1898) | WIEU | Veronica beccabunga | AU |
| Trichocerca rattus (Müller, 1776) | MITL | open water | SU |
| Trichocerca similis (Wierzejski, 1893) | CENL, KORU, MTIU | open water Potamogeton crispus | SP, SU, AU |
| Trichocerca species non determinata | CENL, KORL, MITL, WIEU | open water, bottom sediments, Sparganium erectum | SU, AU |
| Trichocerca taurocephala (Hauer, 1931) | MITU | bottom sediments | SU |
| Trichocerca tenuior (Gosse, 1886) | KORU | open water | SU |
| Trichocerca weberi (Jennings, 1903) | KORU | open water | SU |
| Trichotria pocillum (Müller, 1776) | WIEL | open water, Rorippa amphibia | AU |
| Trichotria tetractis (Ehrenberg, 1830) | MITL | diatom aggregation | SU |
| Wierzejskiella velox (Wiszniewski, 1932) | DZIU | bottom sediments | AU |
| Wulfertia ornata Donner, 1943 | VISL | Elodea canadensis | AU |
Table 4.
The number of rotifer taxa (ranges) in specific seasons of the year (superscript a, b, c denotes significant differences between the seasons).
Table 4.
The number of rotifer taxa (ranges) in specific seasons of the year (superscript a, b, c denotes significant differences between the seasons).
| Variable | Spring | Summer | Autumn | H Value | p Value |
|---|---|---|---|---|---|
| Number of taxa (ranges) | 0–15 b,c | 1–20 a,c | 2–23 a,b | 12.408 | 0.002 |
a Spring, b Summer, c Autumn.
Table 5.
The results of multiple linear regression analysis of the influence of the selected variables on species richness. Abbreviations: β—regression coefficient, SE—standard error of β.
Table 5.
The results of multiple linear regression analysis of the influence of the selected variables on species richness. Abbreviations: β—regression coefficient, SE—standard error of β.
| Variable | β | SE | t Value | p Value |
|---|---|---|---|---|
| Width of the riverbed | 0.1397 | 0.1820 | 0.7675 | 0.4506 |
| Flow velocity | −0.1104 | 0.1552 | −0.7111 | 0.4842 |
| Dissolved oxygen | −0.3332 | 0.1487 | −2.2408 | 0.0345 |
| Temperature | −0.0981 | 0.1594 | −0.6155 | 0.5443 |
| TDS | −0.9488 | 0.4505 | −2.1063 | 0.0463 |
| Calcium | −0.2322 | 0.5140 | −0.4520 | 0.6555 |
| pH | −0.0774 | 0.1358 | −0.5698 | 0.5743 |
| Nitrates | 0.0087 | 0.1286 | 0.0675 | 0.9468 |
| Nitrites | 0.6085 | 0.2652 | 2.2940 | 0.0313 |
| Ammonium | −0.0855 | 0.1862 | −0.4592 | 0.6504 |
| Phosphates | −0.2075 | 0.1797 | −1.1550 | 0.2600 |
| Iron | −0.0293 | 0.1717 | −0.1709 | 0.8658 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Halabowski, D.; Bielańska-Grajner, I.; Lewin, I.; Sowa, A. Diversity of Rotifers in Small Rivers Affected by Human Activity. Diversity 2022, 14, 127. https://0-doi-org.brum.beds.ac.uk/10.3390/d14020127
AMA Style
Halabowski D, Bielańska-Grajner I, Lewin I, Sowa A. Diversity of Rotifers in Small Rivers Affected by Human Activity. Diversity. 2022; 14(2):127. https://0-doi-org.brum.beds.ac.uk/10.3390/d14020127
Chicago/Turabian StyleHalabowski, Dariusz, Irena Bielańska-Grajner, Iga Lewin, and Agnieszka Sowa. 2022. "Diversity of Rotifers in Small Rivers Affected by Human Activity" Diversity 14, no. 2: 127. https://0-doi-org.brum.beds.ac.uk/10.3390/d14020127
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.