
Three New Species of Absidia (Mucoromycota) from China Based on Phylogeny, Morphology and Physiology


1
Institute of Microbiology, School of Ecology and Nature Conservation, Beijing Forestry University, Beijing 100083, China
2
College of Life Sciences, Shandong Normal University, Jinan 250014, China
3
School of Civil Engineering and Architecture, Anhui University of Technology, Ma’anshan 243002, China
4
Key Laboratory for Forest Resources Conservation and Utilization in the Southwest Mountains of China, Ministry of Education, Southwest Forestry University, Kunming 650224, China
*
Authors to whom correspondence should be addressed.
Diversity 2022, 14(2), 132; https://0-doi-org.brum.beds.ac.uk/10.3390/d14020132
Submission received: 28 December 2021
/
Revised: 9 February 2022
/
Accepted: 10 February 2022
/
Published: 13 February 2022
(This article belongs to the Special Issue Diversity and Evolution of Fungi)
Abstract
:Species of Absidia are distributed widely in the environment, while their diversity is insufficiently studied. Three new species, A. frigida, A. gemella and A. longissima, are proposed herein from Xinjiang and Yunnan in China based on phylogenetic, morphological and physiological evidence. According to maximum likelihood (ML), maximum parsimony (MP) and Bayesian inference (BI) analyses, the phylogenetical results suggest that A. frigida, A. gemella and A. longissima are closely related to A. psychrophilia, A. turgida and A. zonata and A. koreana, respectively, based on ITS and LSU rDNA sequences. Absidia frigida is characterized by a lower growth temperature, which does not grow above 24 °C. It differs from A. psychrophilia by sporangiophores, sporangia, columellae, collars and projections. Absidia gemella is distinguished from A. turgida by hypha, sporangiospores, sporangia, projections and sporangiophores. Absidia longissima is discriminated from A. zonata and A. koreana by sporangiophores, columellae and collars. The three new species are described and illustrated in this article.
1. Introduction
Absidia Tiegh. is a core genus of early-diverging fungi, belonging to Mucoromycota, Mucoromycotina, Mucoromycetes, Mucoromycetes, Mucorales and Cunninghamellaceae (www.indexfungorum.org, accessed on 27 December 2021). Absidia was erected in 1876 and typified by A. reflexa Tiegh. [1,2]. Members of Absidia are widely distributed in soils, plant residues, herbivorous dung, decaying substrates and air [2,3,4,5,6,7,8,9,10]. In particular, some species are found in the mycangia of ambrosia beetles (A. psychrophilia) [11,12], as well as the body surface of bats (A. stercoraria: https://bccm.belspo.be/content/remarkable-fungal-biodiversity-northern-belgium-bats, accessed on 27 December 2021). Species within Absidia possess important metabolites for industrial and medical applications, such as steroids, α-galactosidase, laccase, fatty acids and chitosan [13,14,15,16,17,18].
Species of Absidia produce stolons. Sporangiophores form on the middle part of the stolons, while rhizoids form at both ends. Sporangia are multi-spored, pyriform to globose, deliquescent-walled and apophysate [2,3,4,19]. Columellae are conical, suglobose to globose or applanate, commonly with one to several projections [2,4,20,21]. Zygospores are contained in zygosporangia, and their opposite suspensory cells are appendaged [13,20].
Some mycologists and taxonomists, such as Bainier, Hagemann, Lendner, Hesseltine, Ellis and Schipper, not only described and classified the genus Absidia s.l. but also inferred its phylogeny based on morphological and physiological features [13,22,23,24,25]. Incorporating molecular data, morphological traits and growth temperatures, the traditional classification of Absidia s.l. has been revised and divided into three genera, namely, Lentamyces (parasitic on other mucoralean fungi, optimum growth temperatures between 14 °C and 25 °C), Absidia s.s. (mesophilic, optimum growth temperatures between 25 °C and 34 °C) and Lichtheimia (thermotolerant, optimum growth temperatures between 37 °C and 45 °C) [13,20,21].
Currently, 43 species are described in Absidia (Table 1, www.indexfungorum.org, accessed on 13 December 2021), with type strains originating from 17 countries, including Australia, Brazil, Canada, China, Cuba, the Czech Republic, Egypt, France, Holland, India, Mexico, Pakistan, South Korea, Switzerland, Tanzania, Thailand and the USA. Until now, 14 species have been recorded in China [3,4,26], and more potential novel species are being illustrated [27]. In this paper, three new species, A. frigida, A. gemella and A. longissima, are described from soil in China according to phylogenetic, morphological and physiological evidence.
2. Materials and Methods
2.1. Sample Collection and Strain Isolation
Soil samples were collected from Yunnan and Xinjiang in China in September 2021. Then, strains were isolated according to the method in previous studies [3,4]. In brief, soil (1 g) was suspended in sterile water (100 mL), and then the suspension (100 uL) was spread on plates with potato dextrose agar (PDA: 200 g potato, 20 g glucose, 20 g agar and 1000 mL distilled water) supplied with streptomycin sulfate (100 mg/mL) and ampicillin (100 mg/mL). The plates were incubated in the dark at 20 °C and 25 °C. Colonies were purified and then deposited in the China General Microbiological Culture Collection Center, Beijing, China (CGMCC). Cultures were also dried and deposited in the Herbarium Mycologicum Academiae Sinicae, Beijing, China (HMAS).
2.2. Morphology and Maximum Growth Temperature
Morphological observations and maximum growth temperature tests followed the method by Zheng et al. [28,29,30,31,32,33]. In brief, malt extract agar medium [34] (MEA: malt extract 30 g, peptone 3 g, agar 20 g, 1000 mL distilled water), a stereomicroscope (SMZ1500, Nikon, Tokyo, Japan) and a microscope (Axio Imager A2, Carl Zeiss, Oberkochen, Germany) were used. For describing morphological characteristics, a range between the minimum and maximum sizes based on a statistic of more than 20 measurements was adopted. For maximum growth temperature tests, plates were firstly incubated at 20 °C for 2 d, and then the incubation temperature increased by a gradient of 1 °C until the colonies stopped growing.
2.3. DNA Extraction, Amplification and Sequencing
Colonies were grown at 20 °C and 25 °C on synthetic mucor agar medium (SMA: dextrose 20 g, asparagine 2 g, KH2PO4 0.5 g, MgSO4·H2O 0.25 g, thiamin chloride 0.5 mg, agar 20 g, 1000 mL distilled water, pH7) for a week. Total cell DNAs were extracted with a reagent kit (GO-GPLF-400, GeneOnBio Corporation, Changchun, China). A fragment covering the entire internal transcribed spacer (ITS) and a partial large subunit of ribosomal DNA (LSU rDNA) were amplified with the primer pair NS5M and LR5M (5′-GGC TTA ATT TGA CTC AAC ACG G-3′ and 5′-GCT ATC CTG AGG GAA ACT TCG-3′, respectively). The polymerase chain reaction (PCR) program followed Zhao et al. [3]. Sanger sequencing for PCR products was conducted by an external company (BGI Tech Solutions Beijing Liuhe Co., Limited, Beijing, China). ITS and LSU rDNA sequences were assembled and proofread with Geneious 9.0.2 (http://www.geneious.com, accessed on 1 May 2021) and then submitted to GenBank under the accession numbers in Table 2; top hits of the BLAST search for ITS sequences are provided in Supplementary Table S1.
2.4. Phylogenetic Analyses
For reconstructing phylogenetic trees, all the sequences were aligned with AliView (version 3.0) [35] and MAFFT (version 7, https://mafft.cbrc.jp/alignment/server/, accessed on 12 December 2021), and then manual proofreading was performed. Maximum likelihood (ML), maximum parsimony (MP) and Bayesian inference (BI) analyses were all adopted for phylogenetic analyses as described in Nie et al. [36,37], using RAxML (version 8) with the GTRGAMMA substitution model [38], PAUP (version 4.0b10) [39] and MrBayes (version 3.2.7a) [40], respectively. Finally, sequence alignments and phylogenetic trees were deposited in TreeBase (submission ID SS29123).
3. Results
3.1. Taxonomy
In this study, we propose three new species of Absidia from Xinjiang and Yunnan, China (Table 1 and Table 2; Figure 1, Figure 2, Figure 3 and Figure 4). All these novel taxa were demonstrated by molecular sequences, morphology and physiology.
3.1.1. Absidia frigida H. Zhao, Y.C. Dai & X.Y. Liu, sp. nov.
Fungal Names: FN570961. Figure 1.
Etymology: frigida (Lat.) referring to a relatively lower maximum growth temperature of no more than 24 °C.
Holotype: HMAS 351587.
Description: Colonies on MEA at 20 °C for 7 days, growing moderately slow, attaining 60 mm diameter, white at first, gradually becoming light brown, regularly at reverse. Hyphae branched, hyaline at first, sometimes brownish when mature, aseptate when juvenile, septate with age, 5.5–18.5 μm wide. Stolons branched, hyaline, brownish, smooth, with septa. Rhizoids finger-like, rarely present, always unbranched, rarely branched, few swollen at the top. Sporangiophores arising from rhizoids, or substrate hyphae, erect or slightly bent, 1–4 in whorls, unbranched or simple branched, while sympodial not observed, hyaline, sometimes with a septum 16.0–23.0 μm below apophyses, 21.0–300.0 μm or more in length and 3.0–7.0 μm in width. Sporangia subglobose to pyriform, deliquescent-walled, smooth, multi-spored, colorless when young, pigmented when elder, 13.0–33.0 μm long and 12.5–32.0 μm wide. Apophyses distinct, hyaline, subhyaline or slightly pigmented, 4.5–9.5 μm high, 3.5–7.0 μm wide at the base and 7.0–11.5 μm wide at the top. Collars absent. Columellae conical, ovoid, elliptical, subglobose to globose, hyaline or subhyaline, smooth, 16.0–23.0 μm long and 15.5–18.0 μm wide, or 13.0–20.5 μm in diameter. Projections present or absent, if present, always one only, rarely two, small, hyaline or subhyaline, 1.5–3.0 μm long. Sporangiospores cylindrical, hyaline to subhyaline, smooth, 2.5–4.5 μm long and 2.0–3.0 μm wide. Zygospores not observed. Chlamydospores absent.
Maximum growth temperature: 24 °C.
Material examined: China, Xinjiang, Ili Kazak, Zhaosu County, 43°13′58″ N, 81°10′45″ E, altitude: 2219 m, from soil sample, 31 October 2021, Heng Zhao (holotype HMAS 351587, living ex-holotype culture CGMCC 3.16201).
3.1.2. Absidia gemella H. Zhao, Y.C. Dai & X.Y. Liu, sp. nov.
Fungal Names: FN570962. Figure 2.
Etymology: gemella (Lat.) referring to producing two types of sporangiospores.
Holotype: HMAS 351588.
Description: Colonies on MEA at 27 °C for 7 days, growing slow, attaining 45 mm diameter, irregular zonate, white at first, become brown when older, irregular at reverse. Hyphae branched, hyaline at first, brownish when mature, aseptate when juvenile, septate with age, 4.5–17.0 μm wide, substrate hyphae well developed, always branched, sometimes formed elliptical, subglobose to globose swollen in the internal. Stolons branched, hyaline, brownish, smooth, septate. Rhizoids root-like, not well developed, always branched, rarely with a septum at the top. Sporangiophores arising from stolons, erect or slightly bent, often monopodial, simple branched, commonly sympodial, rarely 2–5 in whorls, sometimes a swelling beneath sporangium, hyaline, with one to several septa, 38.0 to more than 350.0 μm and 3.5–6.0 μm in width. Sporangia oval to pyriform, deliquescent-walled, smooth, multi-spored, colorless when young, pigmented when old, 16.0–25.5 μm long and 16.5–24.0 μm wide. Apophyses distinct, subhyaline to hyaline, 3.0–7.5 μm high, 3.5–7.0 μm wide at the base and 7.0–14.0 μm wide at the top. Collars present, small. Columellae globose, subglobose to elliptical, hyaline or pigmented, smooth or rough, 11.0–22.0 μm long and 12.5–17.5 μm wide. Projections absent. Sporangiospores two types, cylindrical, or subglobose to globose, hyaline or subhyaline, smooth, 3.0–4.5 μm long and 2.5–4.0 μm wide, or 2.5–3.5 μm in diameter. Zygospores not observed. Chlamydospores absent.
Maximum growth temperature: 29 °C.
Material examined: China, Xinjiang, Altay City, Burqin County, 28°37′1″ N, 87°2′58″ E, altitude: 1330 m, from soil sample, 31 October 2021, Heng Zhao (holotype HMAS 351588, living ex-holotype culture CGMCC 3.16202).
3.1.3. Absidia longissima H. Zhao, Y.C. Dai & X.Y. Liu, sp. nov.
Fungal Names: FN570963. Figure 3.
Etymology: longissima (Lat.) referring to producing very long rhizoids.
Holotype: HMAS 351589.
Description: Colonies on MEA at 27 °C for 7 days, growing moderately fast, attaining 70 mm diameter, irregular concentrically zonate with ring, white at first, gradually becoming gray, irregular at reverse. Hyphae branched, hyaline at first, brownish when mature, aseptate when juvenile, septate with age, 3.5–11.5 μm wide. Stolons branched, hyaline, brownish, smooth, with septa. Rhizoids root-like, well developed, always branched. Sporangiophores arising from stolons, erect or slightly bent, 2–5 in whorls, monopodial, unbranched or simple branched, while sympodial not observed, hyaline, with a septum 12.5–19.5 μm below apophyses, 40.0–190.0 μm or more in length and 2.5–4.5 μm in width. Sporangia globose to pyriform, deliquescent-walled, smooth, multi-spored, colorless when young, brownish when old, 9.5–28.0 μm long and 10.0–28.0 μm wide. Apophyses small, sometimes not distinct, subhyaline to hyaline, slightly pigmented, 2.5–7.5 μm high, 3.0–6.0 μm wide at the base and 4.5–17.5 μm wide at the top. Collars absent. Columellae subglobose to globose, hyaline, smooth, 7.0–11.5 μm in diameter. Projections present or absent, if present, one only. Sporangiospores cylindrical, hyaline, smooth, slightly contracts in center, 3.0–4.0 μm long and 1.5–2.0 μm wide. Zygospores not observed. Chlamydospores absent.
Maximum growth temperature: 36 °C.
Material examined: China, Yunnan Province, Qujing, Huize County, from soil sample, 15 November 2021, Heng Zhao (holotype HMAS 351589, living ex-holotype culture CGMCC 3.16203).
3.2. Phylogenetic Analyses
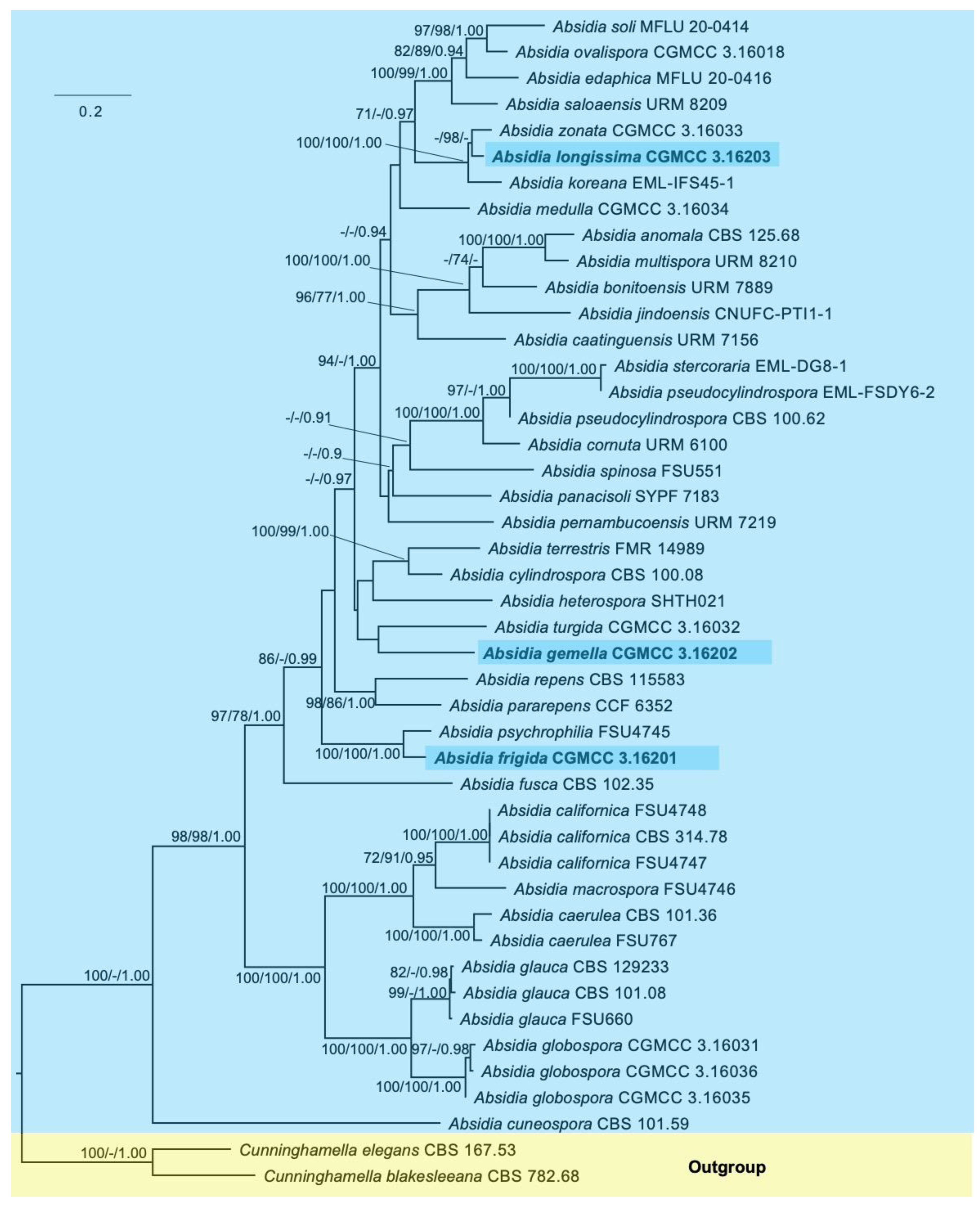
The phylogenetic trees of individual ITS and LSU rDNA are provided in Supplementary Figure S1a,b, respectively. The concatenated ITS and LSU rDNA sequence dataset consists of 45 taxa, including 36 species of Absidia and 2 species of the outgroup Cunninghamella. A total of 1652 sites are composed of 619 constant, 775 parsimony-informative and 258 parsimony-uninformative characters. The maximum parsimony (MP) tree result shows that the tree length (TL), consistency index (CI), homoplasy index (HI), retention index (RI) and rescaled consistency index (RC) are 5600, 0.3493, 0.6507, 0.5326 and 0.1860, respectively. The best model of Bayesian inference (BI) is GTR + I + G, and the average standard deviation of split frequencies is no more than 0.01. The topology of the maximum likelihood tree (ML) was chosen to represent the phylogenetic relationship (Figure 4), since ML, MP and BI resulted in similar topologies. The results suggest that Absidia frigida is closely related to A. psychrophilia (100/100/1.00); Absidia gemella is next to A. turgida; and, finally, A. longissima is closely related to A. zonata and A. koreana (100/100/1.00).
4. Discussion
The ITS and LSU rDNA tree (Figure 4) shows the phylogenetic positions of the three new species in the genus Absidia. In detail, A. frigida is closely related to A. psychrophilia with sufficient support values (100/100/1.00). However, A. frigida physiologically differs from A. psychrophilia by maximum growth temperature (25 °C vs. 28 °C) [11]. Additionally, morphologically, A. frigida is distinguished from A. psychrophilia by less whorls of sporangiophores (four vs. eight), smaller sporangia (13.0–33.0 μm long and 12.5–32.0 μm wide vs. 20.0–50.0 μm in diameter), smaller columellae (16.0–23.0 μm long and 15.5–18.0 μm wide, or 13.0–20.5 μm in diameter vs. 6.5–30.0 μm in diameter), no collars (absent vs. indistinct) and smaller projections (1.5–3.0 μm vs. 6.5 in length) [11].
Absidia gemella clusters with A. turgida, though their sibling relationship does not obtain a strong support. Physiologically, the maximum growth temperature of A. gemella is lower than that of A. turgida (30 °C vs. 33 °C). Morphologically, A. gemella is differentiated from A. turgida by narrower hyphae (4.5–17.0 μm wide vs. 9.0–23.0 μm wide), narrower sporangiospores (3.5–6.0 μm in width vs. 4.5–11.0 μm in width), different shape of sporangia (oval to pyriform vs. globose to pyriform), smaller sporangia (16.0–25.5 μm long and 16.5–24.0 μm wide vs. 20.5–42.5 μm long and 20.0–41.5(–46.0) μm wide) and no projections (absent vs. 9.5 μm in length) [4]. Moreover, sporangiophores in A. gemella are monopodial, simple branched, commonly sympodial and rarely 2–5 in whorls, while in A. turgida, sporangiophores are 1–4 in whorls, unbranched or sometimes simple [4].
Phylogenetically, Absidia longissima is most closely related to A. zonata (-/98/-) and A. koreana (100/100/1.00). Physiologically, A. longissima, A. zonata and A. koreana also possess similar maximum growth temperatures: no growth at 37 °C, no growth at 38 °C and restricted growth at 37 °C, respectively [4,41]. However, A. longissima morphologically differs from A. zonata and A. koreana by whorls of sporangiophores (2–5 in whorls vs. 2–5(–8) in whorls vs. 2–6 in whorls), shape of columellae (subglobose to globose vs. hemispherical vs. globose), smaller columellae (7.0–11.5 μm in diameter vs. 9.5–19.0 μm long and (6.0–)7.5–14.5(–16.5) μm wide vs. 11.4–19.0 μm long and 11.0–17.0 μm wide) and collars (absent vs. absent or present vs. present around each columella) [4,41].
Morphologically, sporangiospores and projections play essential roles in distinguishing Absidia from other genera [13,20,21]. However, as the numbers of species in Absidia gradually increased, some species were found to possess two or more different shapes of sporangiospores, such as A. gemella, A. multispora, A. pararepens, A. repens and A. turgida [4,5,10], which suggested more characters should be adopted. In addition, the majority of species form projections on the apex of columellae, except A. heterospora [11]. The new species A. gemella does not produce projections on MEA plates even after ten days. Therefore, we believe that the morphological delimitation of the genus Absidia should be revised, especially for the character of projections.
Since 2015, a total of 22 species of Absidia have been proposed, outnumbering those described in the last century (Table 1). Remarkably, in the past two years, seven and five new species have been described in Brazil and China, respectively. At present, Absidia is already the second largest genus in the phylum Mucormycota (www.indexfungorum.org, accessed on 27 December 2021). Taking into account the 3 new species reported herein, 46 species are accepted from all around the world, among which 16 are recorded in China.
Currently, an increasing number of studies focus on fungal diversity and ecological distribution [3,4,42,43,44,45,46,47,48], which provides a foundation to deeply understand the kingdom Fungi. Although a total of 46 species are described in Absidia, their ecological distribution has still not been unraveled. A few of these species were found in dung, insects, leaf litter, etc., while most species were collected from soil (Table 1), which suggested that they may have complex ecological habits or hosts. In this study, the maximum growth temperature of the new species A. frigida was 24 °C, implying that some species of Absidia have adapted to low-temperature environments, and that maybe more potential species will be hidden in extreme habitats.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/d14020132/s1, Figure S1: Maximum likelihood phylogenetic tree of Absidia based on ITS and LSU rDNA sequences, respectively, with Cunninghamella elegans and C. blakesleeana as outgroup, Table S1: Top hits for the new species based on BLAST search for ITS sequences from type materials.
Author Contributions
Data curation, H.Z.; formal analysis, Y.D.; funding acquisition, Y.D. and X.L.; investigation, H.Z.; methodology, Y.N. and T.Z.; project administration, X.L.; software, Y.N. and T.Z.; writing—original draft, H.Z.; writing—review and editing, Y.D. and X.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Second Tibetan Plateau Scientific Expedition and Research Program (STEP), Grant No. 2019QZKK0503, and the National Natural Science Foundation of China, Grant Nos. 31970009 and 32170012.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Sequences and trees have been deposited in GenBank (Table 2) and TreeBase (number: SS29123), respectively.
Acknowledgments
Many scholars are thanked for their help with sampling and depositing, and they are Bao-Kai Cui, Shun Liu, Chang-Ge Song and Tai-Min Xu (Beijing Forestry University); Ze-Fen Yu and Min Qiao (Yunnan University); and Ke Wang, Zhuo Du and You-Zhi Wang (Institute of Microbiology, Chinese Academy of Sciences).
Conflicts of Interest
All authors declare no conflict of interest.
References
- Tedersoo, L.; Sánchez-Ramírez, S.; Kõljalg, U.; Bahram, M.; Döring, M.; Schigel, D.; May, T.; Ryberg, M.; Abarenkov, K. High-level classification of the Fungi and a tool for evolutionary ecological analyses. Fungal Divers. 2018, 90, 135–159. [Google Scholar] [CrossRef] [Green Version]
- van Tieghem, P. Troisième mémoire sur les Mucorinées. Ann. Sci. Nat. Bot. 1878, 4, 312–399. [Google Scholar]
- Zhao, H.; Zhu, J.; Zong, T.K.; Liu, X.L.; Ren, L.Y.; Lin, Q.; Qiao, M.; Nie, Y.; Zhang, Z.D.; Liu, X.Y. Two new species in the family Cunninghamellaceae from China. Mycobiology 2021, 49, 142–150. [Google Scholar] [CrossRef]
- Zong, T.K.; Zhao, H.; Liu, X.L.; Ren, L.Y.; Zhao, C.L.; Liu, X.Y. Taxonomy and phylogeny of four new species in Absidia (Cunninghamellaceae, Mucorales) from China. Front. Microbiol. 2021, 12, 2181. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, T.R.L.; Nguyen, T.T.T.; Lima, D.X.; da Silva, S.B.G.; de Lima, C.F.; Leitão, J.D.; Gurgel, L.M.S.; Lee, H.B.; de Santiago, A.L.M.A. Two new species of the industrially relevant genus Absidia (Mucorales) from soil of the Brazilian Atlantic Forest. Acta Bot. Bras. 2020, 34, 549–558. [Google Scholar] [CrossRef]
- Zhang, T.Y.; Yu, Y.; Zhu, H.; Yang, S.Z.; Yang, T.M.; Zhang, M.Y.; Zhang, Y.X. Absidia panacisoli sp. nov. isolated from rhizosphere of Panax notoginseng. Int. J. Syst. Evol. Microbiol. 2018, 68, 2468–2472. [Google Scholar] [CrossRef] [PubMed]
- Lima, D.X.; Cordeiro, T.R.; de Souza, C.A.; de Oliveira, R.J.; Lee, H.B.; Souza–Motta, C.M.; de Santiago, A.L.A. Morphological and molecular evidence for two new species of Absidia from Neotropic soil. Phytotaxa 2020, 446, 61–71. [Google Scholar] [CrossRef]
- Urquhart, A.S.; Idnurm, A. Absidia healeyae: A new species of Absidia (Mucorales) isolated from Victoria, Australia. Mycoscience 2021, 62, 331–335. [Google Scholar] [CrossRef]
- Hurdeal, V.G.; Gentekaki, E.; Lee, H.B.; Jeewon, R.; Hyde, K.D.; Tibpromma, S.; Mortimer, P.E.; Xu, J. Mucoralean fungi in Thailand: Novel species of Absidia from tropical forest soil. Crypt. Mycol. 2021, 42, 39–61. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Chooi, Y.H.; Gilchrist, C.L.; Lacey, E.; Pitt, J.I.; Roets, F.; Swart, W.J.; Cano-Lira, J.F.; Valenzuela-Lopez, N.; et al. Fungal planet description sheets: 1042–1111. Persoonia 2020, 44, 301. [Google Scholar] [CrossRef]
- Hesseltine, C.; Ellis, J. The genus Absidia: Gongronella and cylindrical-spored species of Absidia. Mycologia 1964, 56, 568–601. [Google Scholar] [CrossRef]
- Kaitera, J.; Henttonen, H.M.; Müller, M.M. Fungal species associated with butt rot of mature Scots pine and Norway spruce in northern boreal forests of Northern Ostrobothnia and Kainuu in Finland. Eur. Plant Pathol. 2019, 154, 541–554. [Google Scholar] [CrossRef]
- Hoffmann, K. Identification of the genus Absidia (Mucorales, Zygomycetes): A comprehensive taxonomic revision. Mol. Identif. Fungi 2010, 439–460. [Google Scholar] [CrossRef]
- Chen, J.; Fan, F.; Qu, G.; Tang, J.; Xi, Y.; Bi, C.; Sun, Z.; Zhang, X. Identification of Absidia orchidis steroid 11β-hydroxylation system and its application in engineering Saccharomyces cerevisiae for one-step biotransformation to produce hydrocortisone. Metab. Eng. 2020, 57, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Kitahata, S.; Ishikawa, H.; Miyata, T.; Tanaka, O. Production of rubusoside derivatives by transgalactosylation of various α-galactosidases. Agric. Biol. Chem. 1989, 53, 2929–2934. [Google Scholar] [CrossRef]
- Kristanti, R.A.; Zubir, M.M.F.A.; Hadibarata, T. Biotransformation studies of cresol red by Absidia spinosa M15. J. Environ. Manag. 2016, 172, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Lv, M.L.; Liu, Z.; Zhang, M.Z.; Wang, Y.N.; Ju, X.; Song, Z.; Ren, L.Y.; Jia, B.S.; Qiao, M.; et al. High-yield oleaginous fungi and high-value microbial lipid resources from Mucoromycota. BioEnergy Res. 2021, 14, 1196–1206. [Google Scholar] [CrossRef]
- Davoust, N.; Persson, A. Effects of growth morphology and time of harvesting on the chitosan yield of Absidia repens. Appl. Microbiol. Biot. 1992, 37, 572–575. [Google Scholar] [CrossRef]
- Benny, G.L.; Humber, R.A.; Morton, J.B. Zygomycota: Zygomycetes. In The Mycota (A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research); McLaughlin, D.J., McLaughlin, E.G., Lemke, P.A., Eds.; Systematics and Evolution; Springer: Berlin/Heidelberg, Germany, 2001; Volume 7A, pp. 113–146. [Google Scholar] [CrossRef]
- Hoffmann, K.; Discher, S.; Voigt, K. Revision of the genus Absidia (Mucorales, Zygomycetes) based on physiological, phylogenetic, and morphological characters; thermotolerant Absidia spp. form a coherent group, Mycocladiaceae fam. nov. Mycol. Res. 2007, 111, 1169–1183. [Google Scholar] [CrossRef]
- Hoffmann, K.; Voigt, K. Absidia parricida plays a dominant role in biotrophic fusion parasitism among mucoralean fungi (Zygomycetes): Lentamyces, a new genus for A. parricida and A. zychae. Plant Biol. 2009, 11, 537–554. [Google Scholar] [CrossRef]
- Ellis, J.; Hesseltine, C. The genus Absidia: Globose-spored species. Mycologia 1965, 57, 222–235. [Google Scholar] [CrossRef]
- Hesseltine, C.; Ellis, J. Species of Absidia with ovoid sporangiospores. I. Mycologia 1966, 58, 761–785. [Google Scholar] [CrossRef]
- Ellis, J.; Hesseltine, C. Species of Absidia with ovoid sporangiospores. II. Sabouraudia 1967, 5, 59–77. [Google Scholar] [CrossRef]
- Schipper, M. Notes on Mucorales—I. Observations on Absidia. Persoonia 1990, 14, 133–149. [Google Scholar]
- Zheng, R.Y.; Liu, X.Y. Species Catalogue of China, Volume 3. Fungi: Chtrid, Zygomycotan, Glomeromycotan Fungi; Science Press: Beijing, China, 2018. [Google Scholar]
- Liu, X.Y. Taxonomy and Phylogeny of the Absidia (Cunninghamellaceae, Mucorales) Introducing Nine New Species and Two New Combinations from China; Research Square: Durham, NC, USA, 2021. [Google Scholar] [CrossRef]
- Zheng, R.Y.; Chen, G.Q.; Hu, F.M. Monosporus varieties of Syncephalastrum. Mycosystema 1988, 1, 35–52. [Google Scholar]
- Zheng, R.Y.; Liu, X.Y. Taxa of Pilaira (Mucorales, Zygomycota) from China. Nova Hedwig. 2009, 88, 255–267. [Google Scholar] [CrossRef]
- Zheng, R.Y.; Liu, X.; Wang, Y.N. Two taxa of the new record genus Backusella from China. Mycosystema 2013, 32, 330–341. [Google Scholar]
- Zheng, R.Y.; Chen, G.Q.; Huang, H.; Liu, X.Y. A monograph of Rhizopus. Sydowia 2007, 59, 273. [Google Scholar]
- Zheng, R.Y.; Liu, X.Y.; Li, R.Y. More Rhizomucor causing human mucormycosis from China: R. chlamydosporus sp. nov. Sydowia 2009, 61, 135–147. [Google Scholar]
- Zheng, R.Y.; Liu, X.Y.; Wang, Y.N. Circinella (Mucorales, Mucoromycotina) from China. Mycotaxon 2017, 132, 43–62. [Google Scholar] [CrossRef]
- Benny, G.L. Methods used by Dr. RK Benjamin, and other mycologists, to isolate zygomycetes. Aliso 2008, 26, 37–61. [Google Scholar] [CrossRef]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.; Yu, D.S.; Wang, C.F.; Liu, X.Y.; Huang, B. A taxonomic revision of the genus Conidiobolus (Ancylistaceae, Entomophthorales): Four clades including three new genera. MycoKeys 2020, 66, 55–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, Y.; Cai, Y.; Gao, Y.; Yu, D.S.; Wang, Z.M.; Liu, X.Y.; Huang, B. Three new species of Conidiobolus sensu stricto from plant debris in eastern China. MycoKeys 2020, 73, 133–149. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (* and Other Methods); Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Sys. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Ariyawansa, H.A.; Hyde, K.D.; Jayasiri, S.C.; Buyck, B.; Chethana, K.T.; Dai, D.Q.; Dai, Y.C.; Daranagama, D.A.; Jayawardena, R.S.; Luecking, R.; et al. Fungal diversity notes 111–252—Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2015, 75, 27–274. [Google Scholar] [CrossRef]
- Liu, X.F.; Tibpromma, S.; Xu, J.C.; Kumla, J.; Karunarathna, S.C.; Zhao, C.L. Taxonomy and phylogeny reveal two new potential edible ectomycorrhizal mushrooms of Thelephora from East Asia. Diversity 2021, 13, 646. [Google Scholar] [CrossRef]
- Cai, Y.; Nie, Y.; Gao, Y.; Huang, B. Natural populations of the entomopathogenic fungus Beauveria bassiana in Chinese forest ecosystems are diverse and reveal equal frequencies of mating types within phylogenetic species. Fungal Ecol. 2022, 56, 101139. [Google Scholar] [CrossRef]
- Guan, Q.X.; Li, Y.F.; Zhao, C.L. Morphological and phylogenetic evidence for recognition of two new species of Hyphoderma (Basidiomycota) from southern China, with a key to all Chinese Hyphoderma. MycoKeys 2021, 83, 145–160. [Google Scholar] [CrossRef]
- Wu, F.; Yuan, H.S.; Zhou, L.W.; Yuan, Y.; Cui, B.K.; Dai, Y.C. Polypore diversity in South China. Mycosystema 2020, 39, 653–682. [Google Scholar] [CrossRef]
- Zheng, H.D.; Zhuang, W.Y.; Wang, X.C.; Zeng, Z.Q.; Wei, S.L. Ascomycetes from the Qilian Mountains, China—Pezizomycetes and Leotiomycetes. Mycosystema 2020, 39, 1823–1845. [Google Scholar] [CrossRef]
- Dai, Y.C.; Yang, Z.L.; Cui, B.K.; Wu, G.; Yuan, H.S.; Zhou, L.W.; He, S.H.; Ge, Z.W.; Wu, F.; Wei, Y.L.; et al. Diversity and systematics of the important macrofungi in Chinese forests. Mycosystema 2021, 40, 770–805. [Google Scholar] [CrossRef]
- Wang, K.; Chen, S.L.; Dai, Y.C.; Jia, Z.F.; Li, T.H.; Liu, T.Z.; Phurbu, D.; Mamut, R.; Sun, G.Y.; Bau, T.; et al. Overview of China’s nomenclature novelties of fungi in the new century (2000–2020). Mycosystema 2021, 40, 822–833. [Google Scholar] [CrossRef]
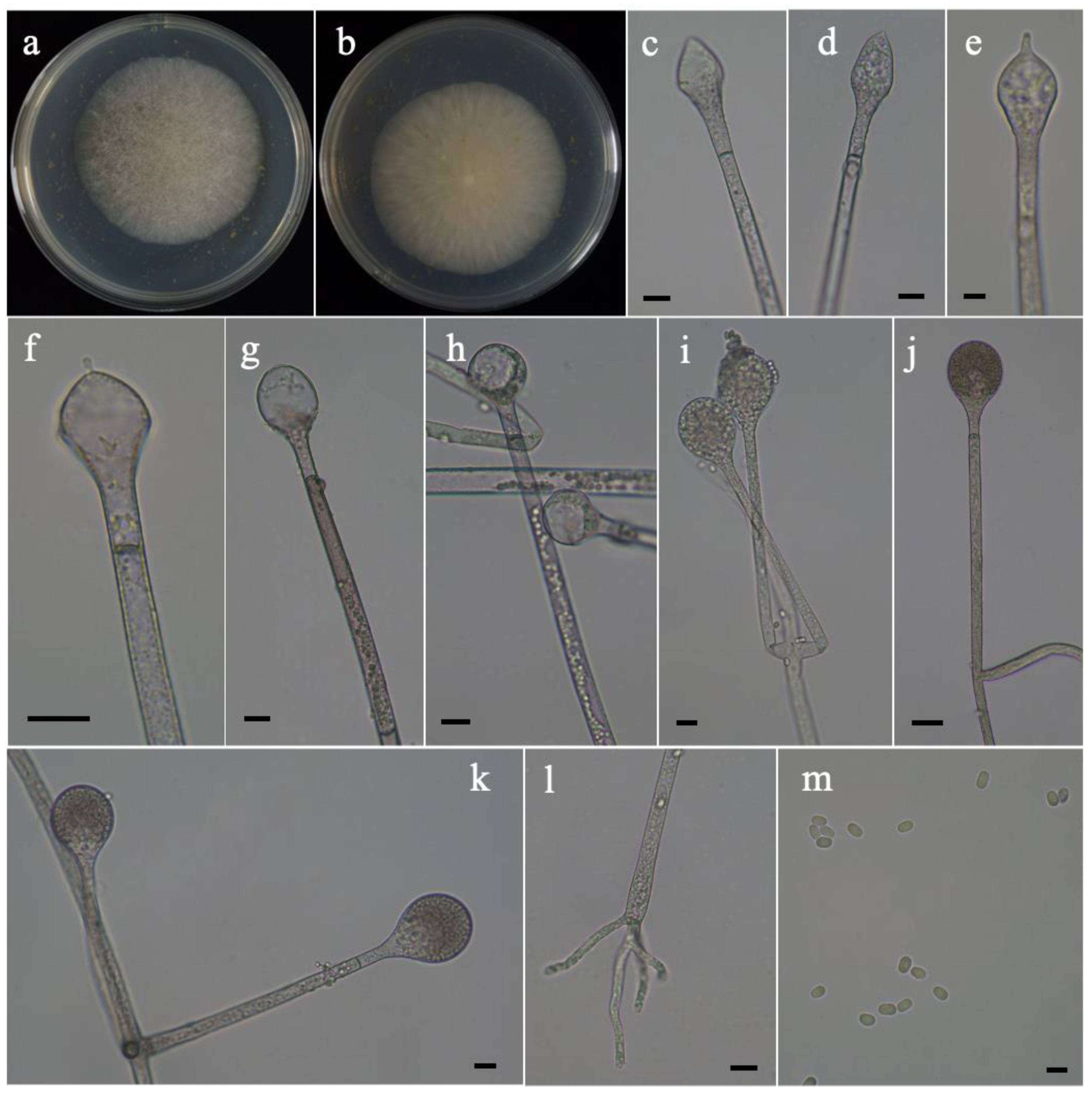
Figure 1.
Morphologies of Absidia frigida ex-holotype CGMCC 3.16201. (a,b) Colonies on MEA, (a) obverse, (b) reverse; (c–h) columellae and projection; (i–k) sporangium; (l) rhizoids; (m) sporangiospores. Scale bars: (c–l) 10 μm, (m) 5 μm.
Figure 1.
Morphologies of Absidia frigida ex-holotype CGMCC 3.16201. (a,b) Colonies on MEA, (a) obverse, (b) reverse; (c–h) columellae and projection; (i–k) sporangium; (l) rhizoids; (m) sporangiospores. Scale bars: (c–l) 10 μm, (m) 5 μm.

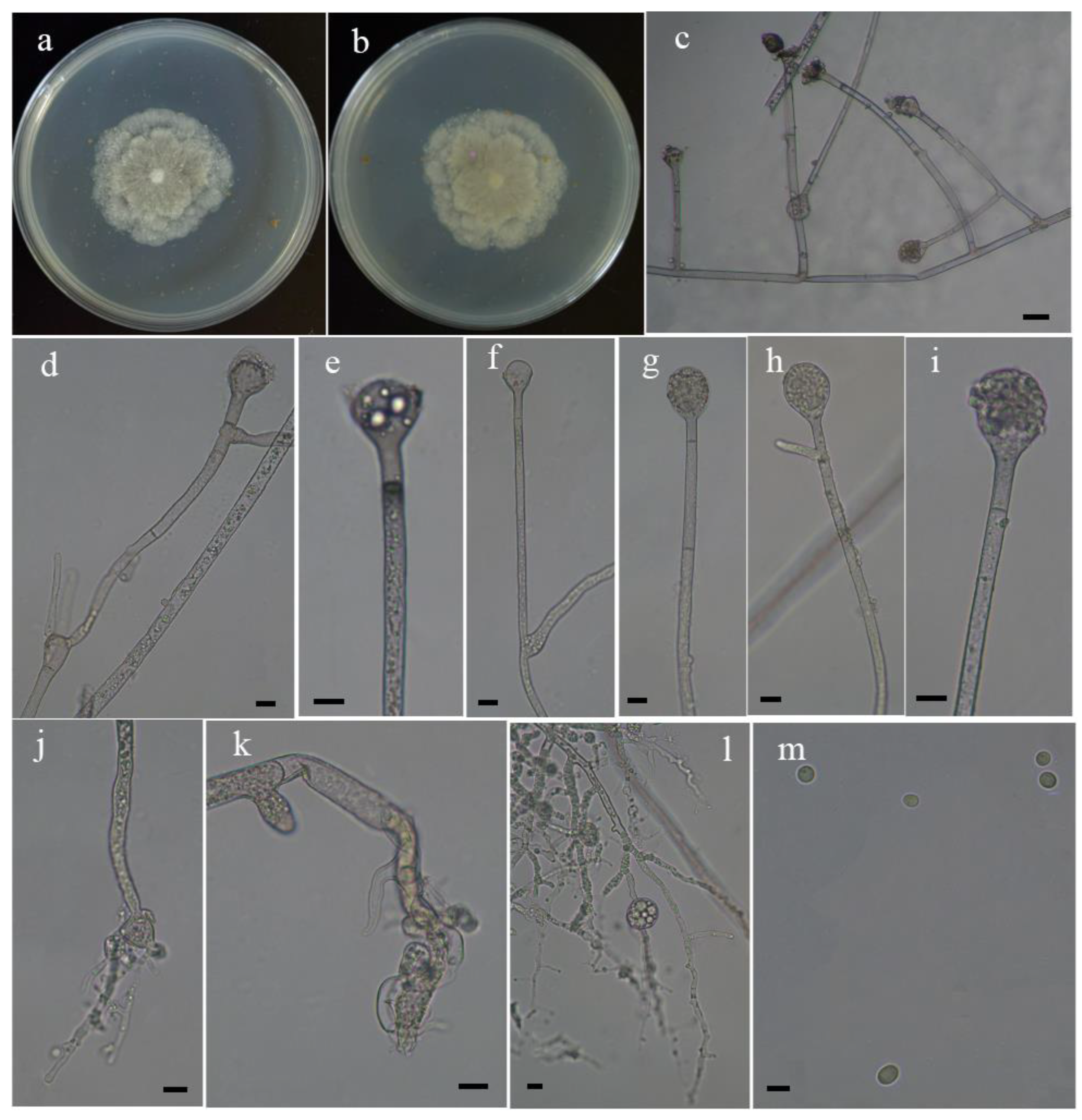
Figure 2.
Morphologies of Absidia gemella ex-holotype CGMCC 3.16202. (a,b) Colonies on MEA, (a) obverse, (b) reverse; (c) monopodial sporangiophores; (d) sympodial sporangiophore; (e,f) columellae; (g–i) sporangium; (j,k) rhizoids; (l) substrate hyphae with swellings; (m) sporangiospores. Scale bars: (c) 20 μm, (d–l) 10 μm, (m) 5 μm.
Figure 2.
Morphologies of Absidia gemella ex-holotype CGMCC 3.16202. (a,b) Colonies on MEA, (a) obverse, (b) reverse; (c) monopodial sporangiophores; (d) sympodial sporangiophore; (e,f) columellae; (g–i) sporangium; (j,k) rhizoids; (l) substrate hyphae with swellings; (m) sporangiospores. Scale bars: (c) 20 μm, (d–l) 10 μm, (m) 5 μm.

Figure 3.
Morphologies of Absidia longissima ex-holotype CGMCC 3.16203. (a,b) Colonies on MEA,(a) obverse, (b) reverse; (c), (d) columellae and projection; (e) sporangiophores arising from stolons borne on rhizoids; (f–h) sporangium or sporangia; (i) sporangiospores; (j) rhizoids. Scale bars: (c,d,g,h) 10 μm, (e,f,j) 20 μm, (i) 5 μm.
Figure 3.
Morphologies of Absidia longissima ex-holotype CGMCC 3.16203. (a,b) Colonies on MEA,(a) obverse, (b) reverse; (c), (d) columellae and projection; (e) sporangiophores arising from stolons borne on rhizoids; (f–h) sporangium or sporangia; (i) sporangiospores; (j) rhizoids. Scale bars: (c,d,g,h) 10 μm, (e,f,j) 20 μm, (i) 5 μm.

Figure 4.
Maximum likelihood phylogenetic tree of Absidia based on ITS and LSU rDNA sequences, with Cunninghamella elegans and C. blakesleeana as outgroups. The three new species, A. frigida, A. gemella and A. longissimi, are shaded. Maximum likelihood (ML) bootstrap values (≥70%)/maximum parsimony (MP) bootstrap values (≥70%)/Bayesian inference (BI) posterior probabilities (≥0.9) of each clade are indicated along branches. Scale bar in the upper left indicates substitutions per site.
Figure 4.
Maximum likelihood phylogenetic tree of Absidia based on ITS and LSU rDNA sequences, with Cunninghamella elegans and C. blakesleeana as outgroups. The three new species, A. frigida, A. gemella and A. longissimi, are shaded. Maximum likelihood (ML) bootstrap values (≥70%)/maximum parsimony (MP) bootstrap values (≥70%)/Bayesian inference (BI) posterior probabilities (≥0.9) of each clade are indicated along branches. Scale bar in the upper left indicates substitutions per site.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The information of type strains of species in Absidia.
| Name | Authors | Record in China | Year of Publication | Host | Location of Type Strains |
|---|---|---|---|---|---|
| Absidia reflexa | Tiegh. | no | 1878 | unknown | France |
| A. repens | Tiegh. | yes | 1878 | unknown | France |
| A. dubia | Bainier | no | 1882 | unknown | Unknown |
| A. caerulea | Bainier | no | 1889 | on wet bread | France |
| A. spinosa | Lendn | yes | 1907 | soil | Switzerland |
| A. cylindrospora | Hagem | yes | 1908 | unknown | Unknown |
| A. glauca | Hagem | yes | 1908 | unknown | Unknown |
| A. heterospora | Y. Ling | yes | 1930 | pine forest soil | Holland |
| A. fusca | Linnem | no | 1936 | unknown | Unknown |
| A. egyptiaca | R. Sartory, J. Mey. & Tawfic | no | 1939 | unknown | Egypt |
| A. cuneospora | G.F. Orr & Plunkett | no | 1959 | soil | USA |
| A. pseudocylindrospora | Hesselt. & J.J. Ellis | yes | 1962 | soil | Tanzania |
| A. psychrophilia | Hesselt. & J.J. Ellis | yes | 1964 | glands of Curculionidae | Canada |
| A. anomala | Hesselt. & J.J. Ellis | no | 1964 | soil | Cuba |
| A. californica | J.J. Ellis & Hesselt | no | 1965 | dung of Rattus | USA |
| A. clavata | B.S. Mehrotra & Nand | no | 1967 | dung of Bos taurus | India |
| A. macrospora | Váňová | no | 1968 | mountain forest soil | Czech Republic |
| A. fassatiae | Váňová | no | 1971 | soil | Czech Republic |
| A. inflata | J.H. Mirza, S.M. Khan, S. Begum & Shagufta | no | 1979 | soil | Pakistan |
| A. narayanae | Subrahm. | no | 1990 | on bat guano | India |
| A. idahoensis | Hesselt., M.K. Mahoney & S.W. Peterson | yes | 1990 | Nomia melanderi | USA |
| A. caatingaensis | D.X. Lima & A.L. Santiago | no | 2015 | soil | Brazil |
| A. koreana | Hyang B. Lee, Hye W. Lee & T.T. Nguyen | no | 2015 | soil | South Korea |
| A. stercoraria | Hyang B. Lee, H.S. Lee & T.T.T. Nguyen | no | 2016 | dung of rat | South Korea |
| A. panacisoli | T. Yuan Zhang, Ying Yu, He Zhu, S.Z. Yang, T.M. Yang, Meng Y. Zhang & Yi X. Zhang | yes | 2018 | rhizosphere of Panax notoginseng | China |
| A. terrestris | Rosas de Paz, Dania García, Guarro, Cano & Stchigel | no | 2018 | soil | Mexico |
| A. jindoensis | Hyang B. Lee & T.T.T. Nguyen | no | 2018 | rhizosphere soil of Coniferae | South Korea |
| A. cornuta | D.X. Lima, C.A. de Souza, H.B. Lee & A.L. Santiago | no | 2020 | soil | Brazil |
| A. pernambucoensis | D.X. Lima, Souza-Motta & A.L. Santiago | no | 2020 | soil | Brazil |
| A. multispora | T.R.L. Cordeiro, D.X Lima, Hyang B. Lee & A.L. Santiago | no | 2020 | soil | Brazil |
| A. saloaensis | T.R.L. Cordeiro, D.X Lima, Hyang B. Lee & A.L. Santiago | no | 2020 | soil | Brazil |
| A. pararepens | Jurjević, M. Kolařík & Hubka | no | 2020 | air | USA |
| A. healeyae | A.S. Urquhart & A. Idnurm | no | 2021 | leaf litter | Australia |
| A. aguabelensis | J.D. Leitão, T.R.L. Cordeiro, Hyang B. Lee & A.L. Santiago | no | 2021 | soil | Brazil |
| A. montepascoalis | L.W.S. Freitas, Hyang B. Lee, T.T.T. Nguyen | no | 2021 | soil | Brazil |
| A. bonitoensis | C.L. Lima, D.X. Lima, Hyang B. Lee & A.L. Santiago | no | 2021 | soil | Brazil |
| A. ovalispora | H. Zhao & X.Y. Liu | yes | 2021 | soil | China |
| A. globospora | T.K. Zong & X.Y. Liu | yes | 2021 | soil | China |
| A. medulla | T.K. Zong & X.Y. Liu | yes | 2021 | soil | China |
| A. turgida | T.K. Zong & X.Y. Liu | yes | 2021 | soil | China |
| A. zonata | T.K. Zong & X.Y. Liu | yes | 2021 | soil | China |
| A. edaphica | V.G. Hurdeal, E. Gentekaki, Hyang B. Lee & K.D. Hyde | no | 2021 | soil | Thailand |
| A. soli | V.G. Hurdeal, E. Gentekaki, Hyang B. Lee & K.D. Hyde | no | 2021 | soil | Thailand |
| A. frigida * | H. Zhao, Y.C. Dai & X.Y. Liu | yes | 2021 | soil | China |
| A. gemella | H. Zhao, Y.C. Dai & X.Y. Liu | yes | 2021 | soil | China |
| A. longissima | H. Zhao, Y.C. Dai & X.Y. Liu | yes | 2021 | soil | China |
* Species proposed herein are shown in bold.
Table 2.
The GenBank accession numbers of sequences used in this study.
| Species * | Strains ** | GenBank Accession nos. | |
|---|---|---|---|
| ITS | LSU | ||
| Absidia anomala | CBS 125.68 T | NR_103626 | NG_058562 |
| A. bonitoensis | URM 7889 T | MN977786 | MN977805 |
| A.caatinguensis | URM 7156 T | KT308169 | KT308171 |
| A. californica | CBS 314.78 | MH861141 | MH872902 |
| A. californica | FSU 4747 | AY944872 | EU736300 |
| A. californica | FSU 4748 | AY944873 | EU736301 |
| A. coerulea | CBS 101.36 | MH855718 | MH867230 |
| A. coerulea | FSU 767 | AY944870 | AF113443 |
| A. cornuta | URM 6100 | MN625256 | MN625255 |
| A. cuneospora | CBS 101.59 T | NR_159602 | NG058559 |
| A. cylindrospora | CBS 100.08 | JN205822 | JN206588 |
| A. edaphica | MFLU 20–0416 | MT396372 | MT393987 |
| A. frigida | CGMCC 3.16201 T | OM108487 | OM030223 |
| A. fusca | CBS 102.35 | NR103625 | NG058552 |
| A. gemella | CGMCC 3.16202 T | OM108488 | OM030224 |
| A. glauca | CBS 129233 | MH865253 | MH876693 |
| A. glauca | CBS 101.08 T | NR_111658 | NG_058550 |
| A. glauca | FSU 660 | AY944879 | EU736302 |
| A. globospora | CGMCC 3.16031 T | MW671537 | MW671544 |
| A. globospora | CGMCC 3.16035 | MW671538 | MW671545 |
| A. globospora | CGMCC 3.16036 | MW671539 | MW671546 |
| A. heterospora | SHTH021 | JN942683 | JN982936 |
| A. jindoensis | CNUFC-PTI1-1 T | MF926622 | MF926616 |
| A. koreana | EML-IFS45-1 T | KR030062 | KR030056 |
| A. longissima | CGMCC 3.16203 T | OM108489 | OM030225 |
| A. macrospora | FSU 4746 | AY944882 | EU736303 |
| A. medulla | CGMCC 3.16034 T | MW671542 | MW671549 |
| A. medulla | CGMCC 3.16037 | MW671543 | MW671550 |
| A. multispora | URM 8210 T | MN953780 | MN953782 |
| A. ovalispora | CGMCC 3.16018 T | MW264071 | MW264130 |
| A. panacisoli | SYPF 7183 T | MF522181 | MF522180 |
| A. pararepens | CCF 6352 | MT193669 | MT192308 |
| A. pernambucoensis | URM 7219 T | MN635568 | MN635569 |
| A. pseudocylindrospora | CBS 100.62 T | NR_145276 | NG_058561 |
| A. pseudocylindrospora | EML-FSDY6-2 | KU923817 | KU923814 |
| A. psychrophilia | FSU 4745 | AY944874 | EU736306 |
| A. repens | CBS 115583 T | NR103624 | HM849706 |
| A. saloaensis | URM 8209 T | MN953781 | MN953783 |
| A. soli | MFLU 20-0414 | MT396373 | MT393988 |
| A. spinosa | FSU 551 | AY944887 | EU736307 |
| A. stercoraria | EML-DG8-1 T | NR_148090 | KT921998 |
| A. terrestris | FMR 14989 T | LT795003 | LT795005 |
| A. turgida | CGMCC 3.16032 T | MW671540 | MW671547 |
| A. zonata | CGMCC 3.16033 T | MW671541 | MW671548 |
| Cunninghamella blakesleeana | CBS 782.68 | JN205869 | MH870950 |
| C. elegans | CBS 167.53 | JN205882 | HM849700 |
* Sequences obtained herein are shown in bold. ** The “T” represents type strains.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhao, H.; Nie, Y.; Zong, T.; Dai, Y.; Liu, X. Three New Species of Absidia (Mucoromycota) from China Based on Phylogeny, Morphology and Physiology. Diversity 2022, 14, 132. https://0-doi-org.brum.beds.ac.uk/10.3390/d14020132
AMA Style
Zhao H, Nie Y, Zong T, Dai Y, Liu X. Three New Species of Absidia (Mucoromycota) from China Based on Phylogeny, Morphology and Physiology. Diversity. 2022; 14(2):132. https://0-doi-org.brum.beds.ac.uk/10.3390/d14020132
Chicago/Turabian StyleZhao, Heng, Yong Nie, Tongkai Zong, Yucheng Dai, and Xiaoyong Liu. 2022. "Three New Species of Absidia (Mucoromycota) from China Based on Phylogeny, Morphology and Physiology" Diversity 14, no. 2: 132. https://0-doi-org.brum.beds.ac.uk/10.3390/d14020132
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.