
The Hesperornithiformes: A Review of the Diversity, Distribution, and Ecology of the Earliest Diving Birds

Dinosaur Institute, Natural History Museum of Los Angeles County, 900 Exposition Boulevard, Los Angeles, CA 90007, USA
*
Author to whom correspondence should be addressed.
Diversity 2022, 14(4), 267; https://0-doi-org.brum.beds.ac.uk/10.3390/d14040267
Submission received: 23 February 2022
/
Revised: 24 March 2022
/
Accepted: 25 March 2022
/
Published: 1 April 2022
(This article belongs to the Special Issue Evolution and Palaeobiology of Flightless Birds)
Abstract
:The Hesperornithiformes (sometimes referred to as Hesperornithes) are the first known birds to have adapted to a fully aquatic lifestyle, appearing in the fossil record as flightless, foot-propelled divers in the early Late Cretaceous. Their known fossil record—broadly distributed across the Northern Hemisphere—shows a relatively rapid diversification into a wide range of body sizes and degrees of adaptation to the water, from the small Enaliornis and Pasquiaornis with lesser degrees of diving specialization to the large Hesperornis with extreme morphological specializations. Paleontologists have been studying these birds for over 150 years, dating back to the “Bone Wars” between Marsh and Cope, and as such have a long history of naming, and renaming, taxa. More recent work has focused to varying degrees on the evolutionary relationships, functional morphology, and histology of the group, but there are many opportunities remaining for better understanding these birds. Broad-scale taxonomic evaluations of the more than 20 known species, additional histological work, and the incorporation of digital visualization tools such as computed tomography scans can all add significantly to our understanding of these birds.
1. Introduction
In the winter of 1870, Othniel Charles Marsh discovered the distal-most end of a tibiotarsus of a large bird in Cretaceous (Coniacian-early Campanian) marine sediments near the Smoky Hill River in western Kansas (specimen 1205 at the Yale Peabody Museum [YPM]) [1]. This unremarkable specimen was the first look at a remarkable group of extinct animals, the first dinosaurs to adapt to a fully aquatic lifestyle and the earliest group of birds to swim away from the ability to fly. On second and third expeditions to western Kansas, in June of 1871 and the fall of 1872, Marsh discovered a more complete specimen (YPM 1200) of the same species as well as fossils of other ancient birds, one of which was nearly complete (YPM 1207) [1]. In 1872, as the infamous “Bone Wars”—an ignominious chapter in American paleontology—were just beginning, Marsh published his first work on these specimens [2]. Marsh designated the material as Hesperornis regalis, a large swimming bird that he interpreted as being most closely related to modern loons, albeit with significant differences from “all other known birds, recent and extinct” [3] (p. 361), and later assigned it to the Natatores [4], a paraphyletic group used at the time to unite modern swimming birds that has since been abandoned. Over subsequent years, Marsh sent numerous expeditions back to the Smoky Hill River in Kansas, resulting in the collection of hundreds of specimens of birds belonging to a group termed the Odontornithes, which Marsh erected for Ichthyornis and Apatornis [5]. Later, Marsh added the coeval Hesperornis to the Odontornithes on the basis of the presence of teeth in the jaws [6]. Marsh would go on to describe a second species of Hesperornis, H. gracilus, and three other related genera, Baptornis advenus, Coniornis altus, and Lestornis crassipes [7], although the latter two are now assigned to Hesperornis.
At the same time that Marsh was working on the North American toothed birds, Harry Seeley [8] was describing a group of small fossil birds from the Upper Cretaceous (Cenomanian) Cambridge Greensand in England from material discovered by Lucas Barrett in 1858 and briefly discussed by Lyell a year later [9]. Unlike Marsh’s larger birds that included well-preserved, articulated specimens, the two species identified by Seeley—Enaliornis barretti and E. sedgewicki—were entirely disarticulated and heavily eroded, as part of a reworked deposit [8]. However, like the fossils Marsh was discovering, these British fossils were also abundant, with dozens of isolated bones available for study in the Woodwardian Museum (now the Woodwardian Collection of the Sedgewick Museum) in Cambridge [8]. Seeley did not recover any specimens with teeth. Furthermore, he deemed Marsh’s reliance on teeth for designation of the Odontornithes to be unsupported, in light of the variability of teeth across modern mammals and reptiles [8], a view that was upheld by Furbringer [9] in 1888 when he established the order Hesperornithiformes (now phylogenetically defined as all taxa more closely related to Hesperornis regalis than to Neornithes or modern birds), removing Hesperornis and Baptornis from Odontornithes. Seeley noted numerous similarities between Enaliornis and modern loons, and so referred Enaliornis to the Natatores [8], as Marsh had originally done with Hesperornis. The placement of Enaliornis within the Hesperornithiformes was first proposed by Lydekker [10] a few years after the erection of this clade, which was later supported by Wetmore [11], Storer [12], Martin and Tate [13], and others. Our modern understanding of hesperornithiform phylogenetics places them within the Ornithurae and very close to the divergence of modern birds, Neornithes (Figure 1). Thus, by the end of the 19th century the Hesperornithiformes were the most diverse lineage of Cretaceous birds known, with a wide geographic and stratigraphic distribution and ranging in size from a bird the size of a grebe to birds as much as 1.5 m long.
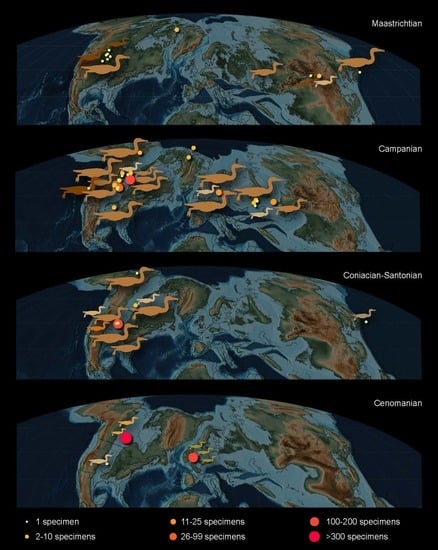
From these early studies, our modern understanding of the Hesperornithiformes has expanded to over 20 species from across Laurasia, identified from marine, transitional, and continental deposits (Figure 2). Interestingly, the fossil collections amassed at the Peabody Museum in Yale University by Marsh and at the Sedgewick Museum in Cambridge University remain the most abundant in terms of number of specimens, rivaled only by the Carrot River material of Pasquiaornis collected in the latter part of the twentieth century [14,15,16].
Studies over the past 150+ years have explored the evolutionary relationships and trajectories, biomechanics, ecology, life history, and biogeography of these incredible birds, as well as continually identifying new families, genera, and species. This review will provide an overview of this body of research, summarizing both where we are today in our understanding of the evolution and biology of these birds and how we got there, and highlight areas for potential future research.
2. General Anatomy
From the first studies of specimens by both Marsh and Seeley, the highly modified bauplan of these birds was recognized as a significant chapter in avian adaptation. This consists of a streamlined body with an elongated skull and neck, heavily reduced forelimb, and dramatically robust hindlimb (Figure 3). In superficial form, it is easy to see how early researchers, and some not-so-early researchers, identified the similarities to modern foot-propelled diving birds such as loons and concluded that these birds were part of the modern diving lineage (e.g., [3,8,17,18,19]). However, this view fails to account for the strong convergence found among modern diving bird lineages, as we now recognize foot-propelled diving to have evolved independently at least four times among modern birds (i.e., loons, grebes, diving ducks, and cormorants), and that even such morphologically similar birds as loons and grebes are not closely related at all [20,21]. In fact, the results of a comprehensive morphometric analysis of hesperornithiforms and modern diving birds showed that the former rarely share morphospace with loons and grebes, and that instead, they overlap more in morphospace with cormorants and diving ducks [22].
Figure 1.
Phylogenetic tree of birds, after Bell and Chiappe [23] and Tanaka et al. [24]. Arrows show the alternative placement of Enaliornis and Pasquiaornis recovered by Tanaka et al. [24].

The skull of hesperornithiforms is elongate, with a long rostrum similar to that seen in modern foot-propelled diving birds. This elongation is due primarily to the length of the premaxilla, as in modern birds, which makes up nearly half the length of the rostrum in Hesperornis [25]. This is unlike more stemward, longirostrine Mesozoic birds (e.g., Longipteryx, Rapaxavis, Dingavis), where elongation of the rostrum is due in part to an extended maxilla [26]. Within hesperornithiforms, there appears to be variation in the degree of elongation of the skull. Enaliornis, the most basal hesperornithiform currently known [23], has a proportionally shorter skull than that of hesperornithids (Parahesperornis and Hesperornis). This is seen in three different regions of the skull: the parietals and temporal fenestrae, the frontals, and the portion of the rostrum rostral to the nares [25]. Furthermore, these regions of the skull are proportionally shorter in Parahesperornis than in Hesperornis, implying potential ecological specializations (i.e., niche partioning) among these likely coeval and sympatric birds [25].
The dentary and maxillae of hesperornithiforms bear small recurved teeth set in a groove (Figure 4). While today we recognize a wide diversity of tooth retention patterns across Mesozoic birds [26,27], when first discovered by Marsh, this feature was striking. The retention of teeth in birds is a conserved character with similar molecular and developmental mechanisms inherited from their nonavian reptilian ancestors [28].
Figure 2.
Distribution of hesperornithiform specimens across the Northern Hemisphere, mapped on paleogeographic reconstructions of the Cretaceous (after [29]): (A) Maastrichtian; (B) Campanian; (C) Coniacian to Santonian; (D) Cenomanian [19]. After Bell and Chiappe [25].

Marsh’s [3] first description of the teeth of Hesperornis noted that they were not set in true sockets (i.e., thecodont implantation), but were instead separated by slight projections from the sides of the groove in which they were set in the jaws. While dentary fragments assigned to Pasquiaornis have been described as having similar tooth implantation in a groove with incomplete sockets [15], published images of some specimens appear to show much more extensive socket development than the slight projections found in hesperornithids (Figure 4).
More recent work exploring the nature of the teeth in Hesperornis via synchrotron imaging found that they have fully thecodont-style root attachments but that secondary loss of periodontal ligaments led to the implantation of the teeth in a groove [30]. While the retention of teeth in the jaw is plesiomorphic in Hesperornis, the emplacement of the teeth is an autapomorphy, uniquely evolved in hesperornithiforms and not seen in other toothed birds such as Archaeopteryx [31], Ichthyornis [32], other toothed ornithuromorphs [33], or toothed enantiornithines [26,34]. The enamel on Hesperornis teeth is thin and simple in structure, with fine fluted ornamentation [35] formed by thickened ridges of enamel [30]. The teeth have a relatively high extension rate (a measure of how fast the tooth growths in height) in the dentine compared to that of nonavian dinosaurs, as calculated from dentine increment lines preserved in the teeth [30]. Tooth replacement involved a resorption pit in the root of the functional tooth, leading to lingual replacement with a calculated mean frequency of 66 days [30]. The teeth of hesperornithiforms are unicuspid and highly recurved, with a hooked shape in side view [20,30]. The teeth exhibit a gradient in curvature, with the mesial teeth more recurved than the distal teeth [1], more than is seen in other Mesozoic birds [36]. Teeth are absent in the premaxillae, as in Ichthyornis and some other early ornithuromorphs (e.g., Gansus, Iteravis), while the dentary and maxilla are toothed.
Figure 3.
The basic bauplan of a hesperornithiform bird, based off Hesperornis regalis. While the degree of reduction of the forelimb and hindlimb proportions vary across taxa, this overall morphology—characterized by an elongated skull and neck, abbreviated forelimbs, and robust hindlimb with a long pelvis—is typical of all hesperornithiforms. Forelimb girdle shown in blue and hindlimb girdle shown in green.
Figure 3.
The basic bauplan of a hesperornithiform bird, based off Hesperornis regalis. While the degree of reduction of the forelimb and hindlimb proportions vary across taxa, this overall morphology—characterized by an elongated skull and neck, abbreviated forelimbs, and robust hindlimb with a long pelvis—is typical of all hesperornithiforms. Forelimb girdle shown in blue and hindlimb girdle shown in green.

The hesperornithiform skull retains numerous additional ancestral characters that are only well-preserved in two taxa (Hesperornis and Parahesperornis). Smaller hesperornithiforms only preserve limited parts of the skull (i.e., Pasquiaornis, Enaliornis, Potamornis). Marsh noted a number of similarities of the skull of Hesperornis to those of modern ratites, including palatines and pterygoids that articulate with facets on the basipterygoid process present on the body of the basisphenoid rostrum as well as an undivided quadrate head [1]. Elzanowski and Galton [37] identified additional ancestral features in the skulls of hesperornithiforms, including open frontoparietal and intraparietal sutures, caudal origination of the pseudotemporalis muscle, and the lack of carotid canals, among others.
A key feature contributing to the streamlined body of hesperornithiforms comes from the elongation of the neck. The majority of hesperornithiform specimens do not preserve a complete vertebral column, with the exception of one specimen of Parahesperornis (KUVP 2287) that appears to preserve the entirety of the vertebral column, minus the atlas. This specimen was largely collected in articulation, and the fit of articulation between the separated vertebral sections indicates it is likely that only the atlas is missing [25]. Modern foot-propelled diving birds use their elongate necks to increase maneuverability underwater. For example, cormorants have been documented to move their head and neck independently of the body during pursuit diving, thus avoiding limitations imposed by the limited turning radius of the entire body [38]. This contrasts with penguins, wing-propelled diving birds that swim with their necks retracted, thus limiting their range of motion to the turning radius of the entire body [39].
Figure 4.
Hesperornithiform teeth and dentaries: (A) dentaries of Hesperornis regalis, showing the groove with implanted teeth in ventral view (1–KUVP 71012) and a broken specimen in medial view showing the internal projections separating individual teeth (2–YPM 1206) as well as an isolated tooth preserved with KUVP 71012 (3); (B) isolated teeth (1, 2) preserved in the roof of the articulated premaxillae of Parahesperornis alexi KUVP 2287; (C) dentary fragment assigned to Pasquiaornis tankei (RSM P2995.5) in lateral (1) and dorsal (2) views showing more extensive socketing (i.e., alveolar configuration) and isolated teeth attributed to Pasquiaornis (3, 4) (images in c from [15]).
Figure 4.
Hesperornithiform teeth and dentaries: (A) dentaries of Hesperornis regalis, showing the groove with implanted teeth in ventral view (1–KUVP 71012) and a broken specimen in medial view showing the internal projections separating individual teeth (2–YPM 1206) as well as an isolated tooth preserved with KUVP 71012 (3); (B) isolated teeth (1, 2) preserved in the roof of the articulated premaxillae of Parahesperornis alexi KUVP 2287; (C) dentary fragment assigned to Pasquiaornis tankei (RSM P2995.5) in lateral (1) and dorsal (2) views showing more extensive socketing (i.e., alveolar configuration) and isolated teeth attributed to Pasquiaornis (3, 4) (images in c from [15]).

A third key feature contributing to the overall bauplan of the hesperornithiforms comes from the pelvic girdle. The pelvis is highly elongated, with an expanded preacetabular ilium that varies in degree across hesperornithiforms but is found in all taxa (Figure 5). The pelvis is fused only around the acetabulum, a plesiomorphic feature also found in most other stem birds (e.g., Archaeopteryx, Sapeornis, Confuciusornis, basal ornithuromorphs, and some enantiornithines). Many enantiornithines have secondarily developed a contact between the ischium and the postacetabular wing of the ilium [40].
Figure 5.
Comparison of hesperornithiform pelves of: (A) Hesperornis YPM 1476; (B) Parahesperornis KUVP 2287; (C) Fumicollis UNSM 20030; and (D) Baptornis AMNH 5101 in left lateral view. Elements are scaled to be of a similar acetabular diameter and aligned at the acetabulum. Inset shows silhouettes of the same elements to scale. After Bell and Chiappe [25].
Figure 5.
Comparison of hesperornithiform pelves of: (A) Hesperornis YPM 1476; (B) Parahesperornis KUVP 2287; (C) Fumicollis UNSM 20030; and (D) Baptornis AMNH 5101 in left lateral view. Elements are scaled to be of a similar acetabular diameter and aligned at the acetabulum. Inset shows silhouettes of the same elements to scale. After Bell and Chiappe [25].

The proportion and, perhaps most importantly, the orientation of the hindlimbs constitute a significant suite of adaptations that underlie interpretations of these birds as foot-propelled divers. In proportion, the femur and tarsometatarsus are reduced in length, and the tibiotarsus is highly elongated, as in modern diving birds (Figure 6). The degree of the shortening of the femur and extension of the tibiotarsus appears to vary dramatically across hesperornithiforms, with more basal taxa such as Baptornis and Fumicollis having a proportionally longer femur and shorter tibiotarsus than in the more derived Hesperornis and Parahesperornis [22].
Of particular interest to the mechanics of swimming in these birds is the orientation of the hindlimb. The earliest observations of these birds included reference to the extreme rotation of the femur, orienting the hindlimb behind the body. Marsh noted that the hindlimb orientation was such that the birds must have had difficulty standing and walking on land [3]. The orientation of the hindlimb directly behind the body is important for two reasons. It places the point of propulsion directly in line with the body being propelled, and it reduces the overall surface area of the body in the direction of motion, where resistance from the water is greatest.
Figure 6.
Ternary diagram showing proportions of the tarsometatarsus, tibiotarsus, and femur of hesperornithiform birds and modern foot-propelled diving birds (inset), and other modern birds. After Bell et al. [22].
Figure 6.
Ternary diagram showing proportions of the tarsometatarsus, tibiotarsus, and femur of hesperornithiform birds and modern foot-propelled diving birds (inset), and other modern birds. After Bell et al. [22].

A final key feature of the hesperornithiform bauplan is the extreme reduction of the forelimb to the point of flightlessness. This is discussed in evolutionary terms in more detail below, but the reduction of the forelimb is seen to varying degrees across the taxa for which forelimb elements are known, with Pasquiaornis displaying the least reduction (but see the discussion in Section 4 regarding this problematic taxon) and Hesperornis and Parahesperornis displaying the most (Figure 7). While the articular ends of the humerus of Pasquiaornis retain easily identifiable morphological landmarks, such as a deltopectoral crest at the proximal end and distinct dorsal and ventral condyles at the distal end, in Hesperornis virtually all such detail is lost, with no discernable deltopectoral crest and only a faint subdivision of the distal end where the condyles would be. Martin and Tate [13] questioned whether the more distal forelimb elements (ulna, radius, carpometatacarpus, and manual digits) developed at all, and proposed that perhaps they had been completely lost in some taxa. The complete loss of flight in hesperornithiforms underscores the success of these birds as foot-propelled divers, the first birds we currently know of to follow this evolutionary path. Among modern foot-propelled divers, flightlessness has evolved occasionally, with one species of flightless cormorant and two of grebes. The loss of flight may indicate that these birds were so well adapted to their foot-propelled lifestyle that they no longer needed flight to be successful for hunting, avoiding predation, and other activities for which birds typically use flight.
3. Taxonomy
The past 150 years of research have resulted in numerous hesperornithiform taxa being named, some of which have been revised or rejected and many of which have never been revisited in light of more recent discoveries. This is undoubtedly one of the main areas for future work on the Hesperornithiformes, as while many taxonomic units are still recognized as valid, they have not undergone robust analysis and lack strong support.
The current taxonomic structure of the Hesperornithiformes recognizes four families: the Enaliornithidae and the Brodavidae, which are both monogeneric; the Baptornithidae, with two genera; and the Hesperornithidae, with five genera (Table 1). Additionally, Pasquiaornis, Potamornis, and Fumicollis are currently unassigned to a family, as discussed in detail below. This overall structure largely predates the use of phylogenetic analyses in developing taxonomic hypotheses, as typified by the initial assignment and later removal of Pasquiaornis to the Baptornithidae. These families and intermediate genera are introduced in this section.
3.1. Enaliornithidae
The Enaliornithidae from the Cambridge Greensand of England are the oldest hesperornithiform family currently known. The Cambridge Greensand is usually interpreted as dating from the early Cenomanian, with reworked late Albian material from the underlying Gault Formation [41], indicating Enaliornis is the oldest group of hesperornithiforms. As described above, they were first reported in 1859 by Lyell [42] and then more fully described by Seeley [8].
Originally, two species were identified on the basis of size but without quantification of those differences [8]. As the remains are highly fragmentary and consist entirely of disarticulated and unassociated remains, determining the exact taxonomic diversity of this group has proven difficult (e.g., [8,13,43,44]). Brodkorb [45] identified a lectotype and numerous paralectotypes for each species. Later, extensive evaluation of all known material by Galton and Martin [44] proposed diagnostic features for the genus as well as for each species. The features identified in combination as diagnostic of the genus included a transversely constricted centrum in the preacetabular synsacrum, the presence of an antitrochanter on the ilium, absence of a distinct neck on the femur, tarsometatarsus with a cranioproximal process originating proximally from metatarsal III, a caudomedial ridge leading to trochlea II distally, and the cranial edge of trochlea IV caudal to the cranial edge of trochlea III. They also identified a third species, E. seeleyi, as being intermediate in size between the larger E. barretti and the smaller E. sedgewicki, but it remains unclear how unassociated elements were combined into a single species [44].
3.2. Baptornithidae
The Baptornithidae was erected to place Baptornis advenus within the Hesperornithiformes as a unique monogeneric family [13]. Since the establishment of the family, several new genera and species have been added to the group. A small baptornithid, Judinornis nogontsavensis, was described from a single thoracic vertebra discovered in Maastrichtian-aged fluvial deposits of Mongolia [46]. The genus Pasquiaornis, consisting of two species from Cenomanian-aged marine strata in Canada, was also added to the Baptornithidae [47]. However, more recent phylogenetic analysis has determined this placement to be unsupported (as discussed in Section 4 below).
Baptornis is monotypic and known from several specimens, both partially complete and isolated elements, in North America, an isolated vertebra from Europe, and an isolated tibiotarsus fragment from Mongolia, while Judinornis remains known from the isolated vertebrae from Mongolia described above. These specimens all date from the Late Cretaceous, the youngest of which is from the Lincoln Limestone Member of the Greenhorn Formation of Kansas (upper middle Cenomanian) [48] and the latest of which is from the Campanian to Maastrichtian-aged Tsagan-Khusu locality in Mongolia [49].
These studies, along with research into baptornithid specimens from Canada [16] and Kansas [48], have added to the diagnostic features of the family. A suite of features has been used as diagnostic of the Baptornithidae, including a slender coracoid (as compared to hesperornithids [13]), elongate preacetabular illium [13], pyramidal patella [13], and dorsally inclined cotyla of the tarsometatarsus [48].
3.3. Brodavidae
The Brodavidae was erected to unite four fragmentary specimens into a single family [50]. The Brodavidae is the second most taxonomically diverse family of hesperornithiforms following the hesperornithids, with four species that range widely in size (Figure 8). Three of the four species are known from isolated tarsometatarsi. However, the holotype and sole specimen of B. varneri is partially complete, preserving a portion of the vertebral column, ribs, pelvis, and most of the hindlimb. This specimen was originally assigned to Baptornis [51] but later moved to the Brodavidae [50], an assignment that has since been supported by phylogenetic analysis [23,24].
The brodavids are known primarily from the Maastrichtian (the fluvial Nemegt Formation of Mongolia, the Frenchman Formation of Canada, and the coastal plain Hell Creek Formation of the USA), with the oldest specimen dating to the Campanian (the marine Pierre Shale, USA) [50].
The Brodavidae was defined using features of the tarsometatarsus, known for all taxa, and the rest of the postcranial skeleton, known only from B. varneri [50]. Two diagnostic features were identified as separating brodavids from other hesperornithiforms [50]: the shortness compared to breadth of the tarsometatarsus (this value was not quantified) and the proximal displacement of the facet for metatarsal I (described as almost in the middle of the tarsometatarsus).
Figure 8.
Comparison of body size among the brodavids: (A) Brodavis varneri SDSM 68430; (B) Brodavis americanus cast of RSM P2315.6; and (C) Brodavis baileyi UNSM 50665.
Figure 8.
Comparison of body size among the brodavids: (A) Brodavis varneri SDSM 68430; (B) Brodavis americanus cast of RSM P2315.6; and (C) Brodavis baileyi UNSM 50665.

3.4. The Hesperornithidae
The family Hesperornithidae was proposed by Marsh [2] as Hesperornidae, later used as Hesperornithidae [7], and at the time was monogeneric and monospecific. Much later, Clarke [32] provided the first cladistic definition for the group as a stem-based name encompassing all taxa more closely related to Hesperornis regalis than to Baptornis advenus. Bell and Chiappe [22] later revised this definition to a node-based clade encompassing all taxa descended from the common ancestor of Hesperornis regalis and Parahesperornis alexi. The Hesperornithidae are the most taxonomically diverse group of hesperornithiforms. In addition to Hesperornis and Parahesperornis, Asiahesperornis [52], Canadaga [53], and eight additional species of Hesperornis [7,54,55,56] have been added to the family.
The Hesperornithidae are known from North America, Europe, and Asia, ranging in age from the Coniacian/Santonian (Vermillion River Formation, Canada) to the Maastrichtian (Hell Creek Formation, USA and Zhuravlovskaya Svita, Russia). This range may in fact be much older, as a specimen has been reported from the marginal marine Mesa Verde Group, USA [57], but the stratigraphic position is poorly constrained. It is interesting to note that the highest hesperornithid diversity in terms of body size and species richness is known from the oldest deposits (such as the marine Niobrara Formation), implying much information on the evolution and diversification of this group remains unknown.
Unfortunately, Marsh did not diagnose the family separately from the genus Hesperornis, and subsequent work has focused on individual species or genera and not diagnostic features of the entire family. Phylogenetic work by Bell and Chiappe [22] identified 28 unambiguous synapomorphies uniting the monophyletic clade Hesperornithidae, which could be evaluated for expanding the current diagnosis. Typical hesperornithid features include the combination of a dramatically reduced forelimb; robust, blocky coracoid; robust femur with expanded trochanter; expanded proximal tibiotarsus with robust cnemial crests; and enlarged fourth trochlea of the tarsometatarsus and pedal phalanx IV. While often considered the largest of the Hesperornithiformes, it is important to remember that species of hesperornithids range widely in size, with H. macdonaldi, H. lumgairi, H. mengeli, and Parahesperornis much smaller than the larger species such as H. regalis and H. rossicus (Figure 9).
3.5. Taxa Outside of Recognized Families
3.5.1. Pasquiaornis
Pasquiaornis was erected to unite two species of small hesperornithiforms from the marine Belle Fourche Formation (Cenomanian) of Canada [47]. Pasquiaornis was originally assigned to the Baptornithidae [47]. However, subsequent phylogenetic analysis failed to return Pasquiaornis + Baptornis as a monophyletic clade, and it was suggested that Pasquiaornis should not be considered part of the Baptornithidae [22]. Additional analysis has since supported this phylogenetic topography [24]. These taxa are only known in the literature from unassociated and disarticulated elements found in a bone-bed deposit in the Belle Fouche Formation [13].
Tokaryk et al. [47] proposed that a combination of size and select morphological differences could be used to separate the disarticulated elements and assigned Pasquiaornis specimens into two species, P. hardei and P. tankei. While P. tankei was described as the larger of the two taxa, specific size differentials used to separate these species have never been quantified. Furthermore, as there are no associated elements known within the genus, it is unclear how different elements are assigned together in one of the two species. Pasquiaornis was diagnosed as having a less-expanded trochanter and proximal end of the femur than is seen in Baptornis, as well as having the anterior intercotylar eminence on the tarsometatarsus overhanging the shaft and trochlea II of the tarsometatarsus positioned posterior to and near the base of trochlea III [47].
3.5.2. Chupkaornis
Chupkaornis is a small hesperornithiform discovered in the Coniacian to Santonian-aged Kashima Formation in Hokkaido, Japan [24]. The holotype and sole specimen was published as a partial skeleton preserving six vertebrae, distal femora, and a fragment of fibula [24]. However, examination of the photographs published indicates what is described as the distal left femur is actually a distal tibiotarsus. Phylogenetic analysis returned Chupkaornis as basal to Brodavis within the Hesperornithiformes and derived from Enaliornis and Pasquiaornis.
Diagnostic features proposed as separating Chupkaornis from other hesperornithiforms include the combination of vertebrae that are fully heterocoelous (but see discussion in Section 4 below) with emarginated lateral excavations on the centra and sharp ventral margins; slender ventral process and laterally expanded fibular condyle of the femur with a finger-like projection on the tibiofibular crest [24].
3.5.3. Fumicollis
The holotype of Fumicollis was originally identified as Baptornis [13] but later recognized as possessing some characters typical of hesperornithids and some typical of Baptornis during the course of research for a phylogenetic study of the hesperornithiforms [23]. This specimen was therefore used to erect a new species—Fumicollis hoffmani—phylogenetically intermediate between the Baptornithidae and the Hesperornithidae [58]. This placement has since been supported by additional analysis [24]. Two additional specimens, both isolated femora, have also been proposed as belonging to Fumicollis. Both of these are known from museum collection studies but are currently unpublished (specimen numbers not available). The holotype and only published specimen preserves a partial vertebral column and nearly complete hindlimb, making it one of the more complete hesperornithiform specimens. Fumicollis is known from the marine Smoky Hill Member of the Niobrara Formation (upper Coniacian to lower Campanian [59]) of Kansas (USA) [58].
A combination of features from the vertebrae, pelvis, femur, tibiotarsus, and tarsometatarsus were used to diagnose the genus [58]. These include an elongate preacetabular pelvis, expanded lateral condyle on the femur (defined as midshaft width 75% of lateral condyle width), medial cnemial crest extended to midshaft of the femur, pyramidal patella, a distinct dorsal ridge of the tarsometatarsus formed by the entire length of metatarsal IV, and others. The presence of both baptornithid and hesperornithid characters can be seen in these traits. For example, the degree of expansion of the lateral condyle is also found in Baptornis, while the dorsal surface of metatarsal IV forming a prominent ridge along its entire length is typical of hesperornithids.
3.5.4. Potamornis
Potamornis skutchi was erected for an isolated quadrate discovered from the fluviodeltaic Lance Formation (late Maastrichtian) in Wyoming (USA) [60]. The element was assigned to the Hesperornithiformes on the basis of an undivided head and an elongate pterygoid condyle, features typical of hesperornithiform quadrates. A unique combination of characters was identified as diagnostic of Potamornis skutchi, including: a strongly asymmetrical quadrate head, rostrally open pit near the medial apex of the head, shallow caudomedial depression, small orbital process, a quadratojugal buttress on the lateral process, and medial and lateral mandibular condyles meeting at an angle of 115 degrees [60]. An isolated tarsometatarsus from the same formation was also tentatively assigned to the genus on the basis of size [60]. However, the specimen was not figured or formally described, and no additional work has been done.
3.6. Taxonomic Challenges
Our understanding of hesperornithiform taxonomy is plagued by a host of problems common to paleontology, such as the renaming of previously described taxa [59,61]; taxa described from highly fragmentary material [24,50,55,56,62]; elements misidentified [24]; and subjective, unspecific, or incorrect characters used for diagnosis [47,63]; reliance upon which may result in further confusing the assignment of fragmentary taxa [50,64]. The majority of hesperornithiform species have been described from fragmentary material (Figure 10). Of the 25 described species, only six include specimens preserving more than three elements, one is known from two elements, and 18 species were described and remain known from a single bone. Whether or not all of these species are valid taxa has rarely been rigorously examined. For example, debate over the synonymy of Coniornis altus [65], Hesperornis altus [62], and Hesperornis montana [62] has appeared in the literature, with both Coniornis and H. montana being invalidated without H. altus ever being resolved in the form of a concise diagnostic description and justification of the “valid” taxon.
Perhaps due to difficulties arising from the fragmentary nature of the fossil record, a number of taxa have been poorly or inaccurately described. An example of one such recurring error and source of much confusion is the presence or absence of the proximal foramina on the cranial surface of the tarsometatarsi of hesperornithiforms. In his monograph on hesperornithiforms, Marsh did not mention the presence of these foramina in Hesperornis or Baptornis [1]. More recent descriptive work has specifically pointed out the lack of these foramina in numerous species of hesperornithiforms [13,44,63]. This has led to the presence of these foramina to be used, in part, as justification for the Cretaceous of Chile from the Hesperornithiformes [64]. Furthermore, the relative degree of development of the foramina has been used as a diagnostic feature of two species of Brodavis [50]. However, closer examination of specimens of H. regalis, H. gracilis, H. crassipes, P. alexi, B. advenus, and other unidentified hesperornithiforms shows that in all cases proximal foramina are present on the cranial surface of the tarsometatarsi [32,48]. Incomplete preparation of the bones may be to blame for the foramina being overlooked by previous authors. Additionally, the appearance of these foramina appears to be closely tied to preservation quality.
As discussed by Bell [65], another problem that is commonly seen in hesperornithiform taxonomy is the reliance on qualitative language to describe quantitative traits. For example, the tarsometatarsus of numerous species has been described as being “slender”. This is essentially a qualitative way of describing the length to width ratio of the element. Using this sort of language in the diagnosis of numerous species instead of presenting morphometric data to precisely define important aspects of morphology creates uncertainty and confusion when making comparisons across dozens of species.
The examples of fraught taxonomy of hesperornithiforms described here illustrate opportunities for future research, as significant synthesis and revision may be achieved from broad-scale studies of currently known material, enhanced and improved with the addition of more recently discovered material that has not been previously published as well as future discoveries [65].
4. Phylogeny of the Hesperornithiformes
Early approaches to hesperornithiform phylogenetics relied on a limited number of taxa (usually under five) and characters (usually under ten) identified as synapomorphies a priori and then used as justification for a particular tree topology. One example of this approach is Cracraft’s [19] tree of diving birds that showed hesperornithiforms within modern birds, basal to a clade containing loons and grebes and derived from penguins, pelicans, and other seabirds (Procellariiformes). Another example is Martin’s [63] work in which he developed two trees showing: (1) Hesperornis and Parahesperornis as a monophyletic clade progressively more derived than the Baptornithidae and the Enaliornithidae; and (2) hesperornithiforms as basal to a monophyletic clade of ichthyornithiforms and modern birds but more derived than “Sauriurae”. A more modern approach to a phylogenetic analysis was taken by Elzanowski and Galton [37], who developed a matrix of 17 characters for seven taxonomic units, but did not conduct an analysis of this matrix and did not offer any phylogenetic hypotheses from these data.
The explosion of Mesozoic bird fossils in the 1990s from China, South America, and Europe initiated a new wave of research into early bird evolution, including the widespread application of modern phylogenetic methods. In these analyses, hesperornithiforms, usually represented by Hesperornis, consistently placed as fairly derived within Ornithuromorpha, usually as the sister taxon to a clade containing Ichthyornis and modern birds (e.g., [32,66,67,68]). More recent work involving multiple hesperornithiform taxa has returned the Hesperornithiformes as the sister taxon to Neornithes, or crown clade birds [22,69,70,71], while other studies retain the placement of the hesperornithiforms as sister to Ichthyornis + Neornithes [24,72,73].
Two things are important to note in comparing the different placement of hesperornithiforms among derived ornithuromorphs. First, the choice of taxa seems to play a role in the placement of hesperornithiforms, with studies that include numerous hesperornithiforms more likely to resolve them as closer to Neornithes (e.g., [24,71]) than studies that include only one or two of the typically more derived hesperornithiforms (i.e., Hesperornis) (e.g., [32,72,73] but see [24]). Second, many matrices have not updated specimen codings in response to the ever-evolving taxonomic changes and in light of new evidence identifying mistakes in previous descriptions of taxa. For example, some studies maintain specimens coded as Baptornis that have since been removed from that genus (e.g., [73]) or use characters that have been identified as erroneous such as features of the quadrate of Baptornis, which was incorrectly described by Martin and Tate [13] and is, in fact, unknown for that taxon (as described in Bell [65] and Bell and Chiappe [25] (e.g., [24,73]).
There are few studies that have examined phylogenetic relationships among the hesperornithiforms, but these studies generally agree in overall topography (Figure 11). The most derived clade consists of a monophyletic Hesperornis, with Parahesperornis as sister taxa, followed by the progressively more basal Fumicollis, Brodavis, and Baptornis [22,24]. There is disagreement at the base of the tree, with Bell and Chiappe [22] resolving Pasquiaornis as more basal than Enaliornis and Tanaka et al. [24] resolving the reverse relationship and a similar switch between Baptornis and Brodavis (Figure 12). This disagreement likely results from the incredibly fragmentary nature of the material for both these taxa and stems from coding discrepancies of features easily obscured by weathering. The details of this are reviewed in Bell and Chiappe [25] but are rooted in the very poor preservation of Enaliornis as part of a reworked deposit that has resulted in the smoothing of the bones, thus making the observation of many details difficult, if not impossible.
5. Evolutionary Trends
As the oldest known lineage of diving birds, hesperornithiforms allow us to study a remarkable evolutionary transition—a group of birds that gave up the ability to fly in favor of foot-propelled diving. This transition is evident in a unique suite of skeletal adaptations as well as in the size of these birds and the range of environments they occupied.
Perhaps one of the most interesting aspects of the evolutionary trends described below is the absence of a fossil record of early stages of these trends. It remains unclear who the predecessors of the Hesperornithformes were. Despite an abundance of Early Cretaceous lagerstätten preserving both freshwater and estuarine environments in China and Spain, there are no clear foot-propelled diving adaptations in the diverse avifauna. The oldest hesperornithiforms, the species of Enaliornis, appear in the earliest Late Cretaceous already equipped with numerous adaptations that support interpretations of a foot-propelled diving lifestyle, including an expanded lateral condyle on the femur and angled articular surface with greatly expanded cnemial expansion on the tibiotarsus, and stacked or shingled metatarsals in the tarsometatarsus. By the middle of the Late Cretaceous (late Coniacian to early Campanian) some of the most abundant deposits of hesperornithiform birds are known from the Smoky Hill Chalk (Kansas, USA) of the Western Interior Seaway, with many roughly coeval taxa ranging from the small Baptornis to the large, highly derived Hesperornis regalis. Furthermore, all of the hesperornithiforms from the early Late Cretaceous (pre-Campanian) are known from entirely marine deposits, while fewer hesperornithiforms are known from the Maastrichtian, these are primarily known from marginal marine to terrestrial deposits (with a small number of exceptions in the Campanian) (Table 2). It is tempting to interpret this as demonstrating an evolutionary diversification from entirely marine taxa to taxa constituting different species adapted to the marine realm, shallow waters of estuaries, and even freshwater. It should be noted that there may be an element of taphonomic bias at play, as the depositional environment of an animal is not necessarily that in which it lived, and the fact that climate and tectonic-driven transgressions of the late Early to early Late Cretaceous led to a disproportionate abundance of marine deposits for this time period [74].
5.1. Flightlessness
The most dramatic and obvious evolutionary trend in the Hesperornithiformes is the complete adaptation to a foot-propelled diving lifestyle. For the majority of hesperornithiforms that preserve forelimb elements, there is agreement that these birds were entirely flightless [1,8,13,25,63,65,93]. The only hesperornithiform for which a degree of flight capacity has been proposed as plausible is Enaliornis, on the basis of the small body size and extensive pneumatization of the braincase [37]. Indeed, the degree of pneumatization in the braincase of Enaliornis is much greater than in either Parahesperornis or Hesperornis [25]. However, additional data from the forelimb, which is entirely unknown in Enaliornis, is required to better evaluate these claims. As described below, some form of limited flight capabilities have also been tentatively suggested for Pasquiaornis [15], but again the dearth of fossil evidence for this taxon makes those claims speculative.
The forelimb and shoulder girdle are indicative of the evolutionary pathway leading to loss of flight in hesperornithiforms, with Pasquiaornis less derived than Baptornis and hesperornithids. The coracoids of hesperornithiforms are less developed as compared to flying birds, with reduced acrocoracoid processes and the absence of a procoracoid process. This trend culminates in hesperornithids, which also have an increasingly short coracoidal neck compared to Baptornis and even more than in Pasquiaornis. A similar trend is seen in the scapula and clavicle, with the articular surfaces only faintly developed in hesperornithiforms. Another factor associated with flightlessness is the complete absence of a ventral keel on the sternum. Hesperornis and Parahesperornis both preserve nearly complete sterna which show the complete absence of a keel.
The humerus is known for several hesperornithiform taxa and consists of an incredibly gracile bone with little to no development of articular surfaces and the deltopectoral and bicipital crests, in both large birds such as Hesperornis and small birds such as Baptornis (Figure 7). Pasquiaornis is the only hesperornithiform that preserves forelimb material for which rudimentary flight abilities have been suggested as tentatively possible [15], based largely on the less reduced state of flight-related features such as the development of the distal condyles and deltopectoral and bicipital crests on the humerus [15,47]. The ulna and radius are only known in Baptornis and Pasquiaornis, where both are reduced with faintly developed articular ends compared to flying birds [13,15,93]. The carpometacarpus is only known in Pasquiaornis, which is also consistent with flightlessness in the thickened compact bone and the distal placement of the extensor process [15]. As indicated above, only the discovery of more complete material in this basal-most hesperornithiform can provide a reliable interpretation of its potential aerial capabilities.
5.2. Foot-Propelled Diving
In concert with the reduction of the forelimb described above, a number of hesperornithiform skeletal features point to a highly derived foot-propelled diving lifestyle. These features have been assessed morphometrically [22,94] and discussed in detail by Bell and Chiappe [25]. In particular, the articulation of the leg in hesperornithiforms has the femur splayed laterally from the pelvis and possibly contained entirely within the body, with the lower limb extending linearly from the knee joint parallel with the mainline of the body. This orients the feet, the source of propulsion, directly behind the body. Adaptations associated with this in Hesperornis include a robust femoral trochanter that extends evenly to the femoral head, an exaggerated lateral femoral condyle roughly even with the medial condyle, a sharply angled proximal articular surface on the tibiotarsus, twisted shafts on the tibiotarsus and tarsometatarsus, and a dramatically enlarged third toe with expanded lateral condyle for rotation of the toe. These features are variably present, but often to a lesser degree, in more basally diverging hesperornithiform taxa.
In addition to the dramatic restructuring of the hindlimb for foot-propelled diving, hesperornithiforms have a suite of more subtle features that also indicate a diving lifestyle. The number, shape, and arrangement of teeth in the jaws of Hesperornis have trophic implications, with the increased number of teeth in the dentary having been related to a piscivorous diet [28], an interpretation well aligned with the environment these birds lived in and morphological interpretations discussed above. The distinct hooked cranial terminus of the premaxilla, which may have been emphasized by the shape of the keratinous beak, may have also been useful for the retention or capture of larger fish. The wide variation in tooth loss, reduction, and shape seen across Mesozoic birds highlights the trophic diversity present among these early birds [26] (in some cases supported by gut contents). However, specific correlations between a particular dental trait, such as the dramatic mesio-distal recurvature gradient in hesperornithiforms, and specific dietary specializations, have not been identified to date [30].
Several features of the skull of hesperornithiforms have been used to support interpretations of a diving lifestyle. Elzanowski and Galton identified the large size of the auricular fossae, the reduced dorsal pneumatic recess, and the flattened cerebellar fossa as traits shared with modern diving birds [37]. The latter two of these features was noted as possibly associated with the expansion of the dural sinuses [37], a convergent feature found in a wide range of diving birds and mammals [95,96,97]. Histological work has identified a thick compact bone wall and comparatively small medullary cavity in the femur of Hesperornis, a feature also found in penguins and interpreted as decreasing buoyancy as a diving adaptation [98].
5.3. Gigantism
One of the first things noted about Hesperornis was its very large body size [2,3], which could approach 1.5 m in length. The discovery of the much smaller Baptornis soon after, showed the dramatic size range present in hesperornithiforms [99]. Most interestingly, this range of sizes does not appear to be correlated to any particular evolutionary trend and is unrelated to the degree of diving specialization [22,25]. While large-bodied taxa are missing among the most basally diverging hesperornithiforms, Enaliornis and Pasquiaornis, there are more derived taxa that are also small. Within the brodavids, for example, the midshaft of the tarsometatarsus of the smallest species, B. mongoliensis, is less than half the diameter of that of the largest species, B. varneri (Figure 9). Similarly, there are several small species of Hesperornis, and while it is not possible to make direct comparisons between them due to lack of overlap in preserved elements, they can be compared to larger species such as H. regalis and H. rossicus (Figure 9). The tarsometatarsus of H. mengeli is half the size of H. rossicus, while the femur of H. macdonaldi is less than half the length of that of H. regalis. All of these species of Hesperornis show similar development of the features described above associated with diving specializations, thus decoupling the evolution of foot-propelled diving from changes in body size [22,25] which we see varying within lineages of hesperornithiforms, indicating the independent evolution of gigantism [22]. This occurred at least twice, once in the brodavids and at least once in several species of Hesperornis, with miniaturization possible in some of the smallest species of Hesperornis as well.
The topic of body size goes hand-in-hand with that of growth rates and ontogenetic patterns. Very little histological work has been done on hesperornithiforms, so the manner or timing in which gigantism (or lack thereof) was achieved in these birds is largely unknown. The first histological study of hesperornithiforms was conducted to address the discrepancies at the time regarding the treatment of hesperornithiforms as either ratites or neognaths within neornithines (modern birds) and found that the bone microsctructure of the hindlimb of Hesperornis was like that of neognaths [100]. This study did not address growth rates. The next histological study of hesperornithiforms characterized the microstructure of the bone from a femur of Hesperornis, identifying the individual as a subadult from the lack of peripheral lamellar bone [98]. Significantly, this study did not identify lines of arrested growth in Hesperornis, indicating there was no evidence for cyclical growth as seen in more basal Mesozoic birds (e.g., Archaeopteryx, Sapeornis, Confuciusornis, enantionithines, and many stem ornithuromorphs). Thus, continuous growth and the resulting absence of lines of arrested growth is a derived feature that Hesperornis shares with modern birds [98]. This has important implications for physiology and life history, as it may indicate a fully endothermic physiology consistent with the interpretation of hesperornithids as venturing far offshore into deep marine waters [98] and growth patterns comparable to those of modern birds (i.e., reaching full-grown size within the first year). The capability for rapid, sustained growth would also contribute to the gigantic body size attained in multiple lineages of these birds.
More recently, hindlimb bones from Hesperornis specimens discovered along a latitudinal gradient from Kansas to the Arctic were examined to investigate the effects of climate and possible migration on bone microstructure [101]. This study found continuous bone deposition and did not identify cyclic growth marks [101], supporting the previous results of Chinsamy et al. [98] and indicating that migratory patterns to different climates is either not recorded in bone microstructure or that these birds achieved skeletal maturity before migrating [101]. The lack of histological work may be in part complicated by taphonomic processes in some of the more prolific hesperornithiform sedimentary units. For example, several members of the Pierre Shale are characterized by calcite crystallization in the preserved bones, destroying histological information.
6. Paleoecology
The obvious and dramatic diving adaptations in hesperornithiforms described above, combined with their wide distribution across the Northern Hemisphere (Figure 2), have led to a large body of work involving the paleoecology of these birds, including the aquatic environments they occupied [22,94,101], modern ecological analogues [22,94], and niche partioning [22,24].
The diversity of hesperornithiform taxa in terms of size, morphological features that are interpreted as diving specializations, and the range of environments in which they are preserved all point to habitat or trophic specializations among hesperornithiforms. Interpretations of habitat preference for hesperornithiforms are limited to the depositional environment in which their fossils were discovered. While these interpretations may not precisely align with the environment in which these birds actually lived, some general conclusions can be drawn. Hesperornithiforms are predominantly known from marine environments, with some specimens known from continental and transitional environments (Figure 12 and Table 2). As mentioned above, only in the latter half of the Late Cretaceous (Campanian-Maastrichtian) do fossils occur in nonmarine sediments, thus suggesting a possible colonization of these environments later in their evolutionary history. Body size does not appear to correlate well with depositional environment, with large and small taxa reported from continental, transitional, and marine environments. There may be an underlying trend of large-bodied birds restricted to marine environments that is obscured by taphonomic processes, particularly in regards to preservation in the reworked marine Cambridge Greensand and Belle Fouche Formation.
Of particular interest is the overlap of multiple taxa in single geologic units such as that seen in the Niobrara Formation and the Pierre Shale of the United States. Both of these units are widely deposited deep-water marine sediments of the Western Interior Seaway, with the older Niobrara Formation grading into the Pierre Shale in some places [102]. While the number of taxa reported may be inflated (see Section 3 above), the range of body sizes preserved in both is striking, with the small Baptornis and Fumicollis, the large H. regalis, and taxa such as Parahesperornis of intermediate size, known from the Niobrara Formation, and some of the smallest Hesperornis species, H. lumgairi and H. macdonaldi, found with large species such as H. chowi and H. regalis in the Pierre Shale. This juxtaposition should not necessarily be interpreted as direct evidence of niche partioning, as it may result in full or part from taphonomic processes or time averaging. However, it does raise the possibility of ecological specializations to reduce interspecific competition. Ecologic niche segregation is common among modern diving birds in which sympatric species differentiate in either diet (prey type) or foraging range [103].
7. Summary and Future Directions
Hesperornithiforms became the first birds (and dinosaurs) to adapt to a fully aquatic lifestyle. A number of morphological features highlight this evolutionary pathway, resulting in a highly streamlined body optimized for diving through the water, propelled by powerful hindlimbs. Adaptations include an elongate neck that allowed for increased maneuverability of a skull with sharp teeth and an expanded rostrum, ideal of capturing fish and other mobile prey. The pelvis was also elongate, allowing for the attachment of larger muscles for powering the feet. The femur was reduced to varying degrees in the different species of hesperornithiform, but the shortened length and horizontal articulation with the pelvis allowed for the orientation of the feet in line with and directly behind the body, optimizing power production by reducing drag. As evidence of the degree to which these birds optimized foot-propelled diving, the forelimbs of all hesperornithiforms were reduced to the point of flightlessness, with the hesperornithids showing the most extreme reduction.
This suite of morphological adaptations are present in varying degrees among the different specimens assigned to the group, suggesting a progression of diving specializations and even the evolution of niche partitioning among these birds. This is supported by the many geologic units where multiple species showing ranges in size and interpreted diving capabilities have been discovered.
The broad morphological diversity present among hesperornithiform specimens has been interpreted as representing an increasing number of taxa over the years. These taxonomic interpretations are complicated by the highly fragmentary nature of the fossil record of these birds. While the addition of more specimens, particularly of the most basal taxa, through future fieldwork is something to look forward to, there remains much to be done with the existing global collection of specimens. Much of the taxonomic work published to date has been limited to the geographic area of the authors (i.e., [13,47,56]) or relied on photographs and loans of select specimens (i.e., [15,24,50]). Very few studies have incorporated direct observations of unpublished material from multiple continents, but even these studies were not able to access much of the Asian and Canadian material (i.e., [22,25,65]). Digitization and publication of a more complete record of current museum collections as measurements, photographs, written morphological descriptions, and three-dimensional datasets such as those from computed tomography or laser scans, would enable broad-scale studies not limited by geography (or travel funds). Such studies could test taxonomic hypotheses that have remained largely untested over the past 150 years of taxonomic work. The creation of digital specimens might also enable digital reconstructions of the skulls, which are disarticulated and often deformed to some degree. Such reconstructions might provide insight into the shape of the hesperornithiform brain and allow comparisons to Mesozoic and modern birds as well as inferences about the sensory capabilities of these birds.
While much of the modern work on these birds has focused on taxonomy (e.g., [15,24,50,58]) or ecology [22,94,101], very little has been done regarding the ontogeny of these birds [98]. Additionally, histological studies might better inform taxonomic studies involving specimens that range widely in size but have less variability in morphology, such as the species of Hesperornis. There are a surprisingly large number of isolated hesperornithiform bones, primarily from the hindlimb, in museum collections across North America, and additional histological work to characterize growth patterns and rates as well as life history as a whole seem to be a potentially fruitful line of inquiry, despite complications from poor preservation.
In conclusion, taxonomic, phylogenetic, and paleoecologic studies on the Hesperornithiformes for the past 150 years have led us to an understanding of these birds as a fascinating chapter in adaptive evolution. Hesperornithiforms are the first group of marine diving birds to evolve, and while the origins of this group remain elusive, a large body of work documents their spread across Laurasia and their expansion from marine to estuarine and even to freshwater environments by the Maastrichtian. While we have a large body of previous research on these birds, there is much to be done in the future.
Author Contributions
Conceptualization, literature review, and initial draft of the manuscript and figures were drafted by A.B. Conceptualization, contribution to the manuscript, and review was provided by L.M.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded in part by a generous donation from the Augustyn Family.
Acknowledgments
The authors would like to thank Stephanie Abramowicz for assistance with figures as well as the collections management staff at all of the museums for their assistance with accessing specimens.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
AMNH, American Museum of Natural History, New York, NY, USA; IZASK, Institute of Zoology, Ministry of Science, Almaty, Kazakhstan; KUVP, University of Kansas Museum of Natural History, Lawrence, KS, USA; LACM, Natural History Museum of Los Angeles County, Los Angeles, CA, USA; MCM, Mikasa City Museum, Mikasa, Japan; RSM, Royal Saskatchewan Museum, Regina, SK, Canada; SDSM, Museum of Geology, South Dakota School of Mines and Technology, Rapid City, SD, USA; UNSM, University of Nebraska State Museum, Lincoln, NE, USA; YPM, Yale Peabody Museum, New Haven, CT, USA; YPM PU, Princeton University (collections now housed in the Yale Peabody Museum).
References
- Marsh, O.C. Odontornithes: A Monograph on the Extinct Toothed Birds of North America; United States Government Printing Office: Washington, DC, USA, 1880; 312p.
- Marsh, O.C. Discovery of a remarkable fossil bird. Am. J. Sci. Ser. 1872, 3, 56–57. [Google Scholar] [CrossRef]
- Marsh, O.C. ART. XLVIII.—Preliminary Description of Hesperornis regalis, with Notices of Four Other New Species of Cretaceous Birds. Am. J. Sci. Arts 1872, 3, 360. [Google Scholar] [CrossRef]
- Marsh, O.C. Fossil birds from the Cretaceous of North America. Am. J. Sci. Arts 1872, 5, 229–231. [Google Scholar]
- Marsh, O.C. On a new subclass of fossil birds. Am. J. Sci. Arts 1873, 5, 161. [Google Scholar]
- Marsh, O.C. On the Odontornithes, or birds with teeth. Am. J. Sci. Arts 1873, 10, 403. [Google Scholar]
- Marsh, O.C. Notice of new Odontornithes. Am. J. Sci. Arts 1876, 11, 509–511. [Google Scholar] [CrossRef]
- Seeley, H.G. On the British Fossil Cretaceous Birds. Q. J. Geol. Soc. 1876, 32, 496–512. [Google Scholar] [CrossRef]
- Furbinger, M. Untersuchungen Zur Morphologie und Systematik der Vögel, Zugleich ein Beitrag Zur ANATOMIE der Stütz- und Bewegungsorgan; T. van Holkema: Amsterdam, The Netherlands, 1888. [Google Scholar]
- Lydekker, R. Catalogue of the Fossil Birds in the British Museum (Natural History) United Kingdom, Order of the Trustees; Longmans & Co.: London, UK, 1891. [Google Scholar]
- Wetmore, A. The fossil birds of the AOU Check-list. Condor 1930, 32, 12–14. [Google Scholar] [CrossRef]
- Storer, R.W. Evolution in the Diving Birds. In Proceedings of the XII International Ornithological Congress; Tilgmannin Kirjapaino: Helsinki, Finland, 1958; Volume 2, pp. 694–707. [Google Scholar]
- Martin, L.D.; Tate, J. The Skeleton of Baptornis advenus (Aves: Hesperornithiformes). Smithson. Contrib. Paleobiol. 1976, 27, 35–66. [Google Scholar]
- Cumbaa, S.L.; Schröder-Adams, C.; Day, R.G.; Phillips, A.J. Cenomanian Bonebed Faunas from the Northeastern Margin, Western Interior Seaway, Canada. In Late Cretaceous Vertebrates from the Western Interior. New Mexico Museum of Natural History and Science Bulletin 35; Lucas, S., Sullivan, R., Eds.; Kansas Academy of Science: Kansas, MO, USA, 2006; pp. 139–155. [Google Scholar]
- Sanchez, J. Late Cretaceous (Cenomanian) Hesperornithiformes from the Pasquia Hills, Saskatchewan, Canada. Master’s Thesis, Carleton University, Ottawa, ON, Canada, 2010. [Google Scholar]
- Tokaryk, T.T.; Harington, C.R. Baptornis sp. (Aves: Hesperornithiformes) from the Judith River Formation (Campanian) of Saskatchewan, Canada. J. Paleontol. 1992, 66, 1010–1012. [Google Scholar] [CrossRef]
- Lambrecht, K. Handbuch der Palaeomithologie; Gebruder Borntraeger: Berlin, Germany, 1933; 1022p. [Google Scholar]
- Brodkorb, P. Origin and Evolution of Birds; Farner, D.S., King, J.R., Eds.; Avian Biology: New York, NY, USA; Academic Press: London, UK, 1971; Volume 1. [Google Scholar]
- Cracraft, J. Phylogenetic Relationships and Monophyly of Loons, Grebes, and Hesperornithiform Birds, with Comments on the Early History of Birds. Syst. Zool. 1982, 31, 22. [Google Scholar] [CrossRef]
- Hackett, S.J.; Kimball, R.T.; Reddy, S.; Bowie, R.C.; Braun, E.L.; Braun, M.J.; Chojnowski, J.L.; Cox, W.A.; Han, K.; Harshman, J.; et al. A phylogenomic study of birds reveals their evolutionary history. Science 2008, 320, 1763–1768. [Google Scholar] [CrossRef]
- Jetz, W.; Thomas, G.H.; Joy, J.B.; Hartmann, K.; Mooers, A.O. The Global Diversity of Birds in Space and Time. Nature 2012, 491, 444–448. [Google Scholar] [CrossRef]
- Bell, A.; Wu, Y.-H.; Chiappe, L.M. Morphometric Comparison of the Hesperornithiformes and Modern Diving Birds. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 513, 196–207. [Google Scholar] [CrossRef]
- Bell, A.; Chiappe, L.M. A Species-Level Phylogeny of the Cretaceous Hesperornithiformes (Aves: Ornithuromorpha): Implications for Body Size Evolution amongst the Earliest Diving Birds. J. Syst. Palaeontol. 2016, 14, 239–251. [Google Scholar] [CrossRef]
- Tanaka, T.; Kobayashi, Y.; Kurihara, K.; Fiorillo, A.R.; Kano, M. The Oldest Asian Hesperornithiform from the Upper Cretaceous of Japan and the Phylogenetic Reassessment of Hesperornithiformes. J. Syst. Palaeontol. 2017, 16, 689–709. [Google Scholar] [CrossRef]
- Bell, A.; Chiappe, L. Anatomy of Parahesperornis: Evolutionary Mosaicism in the Cretaceous Hesperornithiformes (Aves). Life 2020, 10, 62. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, J.K. The trophic habits of early birds. Palaeogeo. Palaeoclim. Palaeoeco. 2019, 513, 178–195. [Google Scholar] [CrossRef]
- O’Connor, J.K.; Wang, M.; Hu, H. A new ornithuromorph (Aves) with an elongate rostrum from the Jehol Biota, and the early evolution of rostralization in birds. J. Syst. Palaeontol. 2016, 14, 939–948. [Google Scholar] [CrossRef]
- Wu, Y.H.; Chiappe, L.M.; Bottjer, D.J.; Nava, W.; Martinelli, A.G. Dental replacement in Mesozoic birds: Evidence from newly discovered Brazilian enantiornithines. Sci. Rep. 2021, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Scotese, C.R. Atlas of Earth History; PALEOMAP Project: Arlington, TX, USA, 2001; Volume 1. [Google Scholar]
- Dumont, M.; Tafforeau, P.; Bertin, T.; Bhullar, B.-A.; Field, D.; Schulp, A.; Strilisky, B.; Thivichon-Prince, B.; Viriot, L.; Louchart, A. Synchrotron Imaging of Dentition Provides Insights into the Biology of Hesperornis and Ichthyornis, the “Last” Toothed Birds. BMC Evol. Biol. 2016, 16, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howgate, M.E. The teeth of Archaeopteryx and a reinterpretation of the Eichstätt specimen. Zoo. J. Linn. Soc. 1984, 82, 159–175. [Google Scholar] [CrossRef]
- Clarke, J.A. Morphology, Phylogenetic Taxonomy, and Systematics of Ichthyornis and Apatornis (Avialae: Ornithurae). Bull. Am. Mus. Nat. Hist. 2004, 286, 1–179. [Google Scholar] [CrossRef]
- Clarke, J.A.; Zhou, Z.; Zhang, F. Insight into the evolution of avian flight from a new clade of Early Cretaceous ornithurines from China and the morphology of Yixianornis Grabaui. J. Anat. 2006, 208, 287–308. [Google Scholar] [CrossRef]
- O’Connor, J.K.; Chiappe, L.M. A revision of enantiornithine (Aves: Ornithothoraces) skull morphology. J. Syst. Palaeo. 2011, 9, 135–157. [Google Scholar] [CrossRef]
- Sander, P.M. The microstructure of reptilian tooth enamel: Terminology, function, and phylogeny. F. Pfeil 1999, 38, 1–102. [Google Scholar]
- Martin, L.D.; Stewart, J.D. Implantation and replacement of bird teeth. Smiths. Cont. Paleobio. 1999, 89, 295–300. [Google Scholar]
- Elzanowski, A.; Galton, P. Braincase of Enaliornis, an Early Cretaceous Bird from England. J. Vertebr. Paleontol. 1991, 11, 90–107. [Google Scholar] [CrossRef]
- Ribak, G.; Weihs, D.; Arad, Z. Consequences of Buoyancy to the Maneuvering Capabilities of a Foot-Propelled Aquatic Predator, the Great Cormorant (Phalacrocorax carbo sinensis). J. Exp. Biol. 2008, 211, 3009–3019. [Google Scholar] [CrossRef] [Green Version]
- Lovvorn, J.R. Mechanics of Underwater Swimming in Foot-Propelled Diving Birds. Proc. Int. Ornithol. Congr. 1991, 20, 1868–1874. [Google Scholar]
- Wang, M.; O’Connor, J.K.; Pan, Y.; Zhou, Z. A bizarre Early Cretaceous enantiornithine bird with unique crural feathers and an ornithuromorph plough-shaped pygostyle. Nat. Commun. 2017, 8, 14141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machalski, M. The Cenomanian ammonite Schloenbachia varians (J. Sowerby, 1817) from the Cambridge Greensand of eastern England: Possible sedimentological and taphonomic implications. Cret. Res. 2018, 87, 120–125. [Google Scholar] [CrossRef]
- Lyell, C. Manual of Elementary Geology, Supplement to the 5th Edn, 3rd ed.; Murray: London, UK, 1859. [Google Scholar]
- Storer, R.W. [Review of] P. Brodkorb, Catalogue of Fossil Birds, Part 1. Auk 1965, 82, 657–658. [Google Scholar] [CrossRef]
- Galton, P.M.; Martin, L.D. Postcranial Anatomy and Systematics of Enaliornis SEELEY, 1876, a Foot-Propelled Diving Bird (Aves: Ornithurae: Hesperornithiformes) from the Early Cretaceous of England. Rev. Paleobiol. 2002, 21, 489–538. [Google Scholar]
- Brodkorb, P. Catalogue of Fossil Birds. Bull. Fla. St. Mus. Bio. Sci. 1963, 7, 179–293. [Google Scholar]
- Kurochkin, E.N. Mesozoic Birds of Mongolia and the Former USSR. In The Age of Dinosaurs in Russia and Mongolia; Benton, M., Shishkin, M., Unwin, D., Kurochkin, E., Eds.; Cambridge University Press: Cambridge, UK, 2000; pp. 544–559. [Google Scholar]
- Tokaryk, T.T.; Cumbaa, S.L.; Storer, J.E. Early Late Cretaceous Birds from Saskatchewan, Canada: The Oldest Diverse Avifauna Known from North America. J. Vertebr. Paleontol. 1997, 17, 172–176. [Google Scholar] [CrossRef]
- Everhart, M.J.; Bell, A. A Hesperornithiform Limb Bone from the Basal Greenhorn Formation (Late Cretaceous; Middle Cenomanian) of North Central Kansas. J. Vertebr. Paleontol. 2009, 29, 952–956. [Google Scholar] [CrossRef]
- Zelenkov, N.V.; Panteleyev, A.V.; Yarkov, A.A. New Finds of Hesperornithids in the European Russia, with Comments on the Systematics of Eurasian Hesperornithidae. Paleontol. J. 2017, 51, 547–555. [Google Scholar] [CrossRef]
- Martin, L.D.; Kurochkin, E.N.; Tokaryk, T.T. A New Evolutionary Lineage of Diving Birds from the Late Cretaceous of North America and Asia. Palaeoworld 2012, 21, 59–63. [Google Scholar] [CrossRef]
- Martin, J.; Cordes-Person, A. A New Species of the Diving Bird Baptornis (Ornithurae: Hesperornithiformes) from the Lower Pierre Shale Group (Upper Cretaceous) of Southwestern South Dakota. Geol. Soc. Am. Spec. Pap. 2007, 427, 227–237. [Google Scholar]
- Nessov, L.A.; Prizemlin, B. A Large Advanced Flightless Marine Bird of the Order Hesperornithiformes of the Late Senonian of Turgai Strait-the First Finding of the Group in the USSR. Tr. Zool. Inst. SSSR 1991, 239, 85–107. [Google Scholar]
- Hou, L.-I. New Hesperornithid (Aves) from the Canadian Arctic. Gu Ji Zhui Dong Wu Xue Bao 1999, 37, 231–237. [Google Scholar]
- Nessov, L.A.; Yarkov., A.A. Hesperornis in Russia, Russk. Ornitol. Zh 1993, 2, 37–54. [Google Scholar]
- Martin, L.D.; Lim, J.-D. New Information on the Hesperornithiform Radiation. In Proceedings of the 5th Symposium of the Society of Avian Paleontology and Evolution, Beijing, China, 1–4 June 2000; Science Press: Beijing, China, 2002. [Google Scholar]
- Aotsuka, K.; Sato, T. Hesperornithiformes (Aves: Ornithurae) from the Upper Cretaceous Pierre Shale, Southern Manitoba, Canada. Cretac. Res. 2016, 63, 154–169. [Google Scholar] [CrossRef]
- Gill, J.; Cobban, W. Regional unconformity in late cretaceous, Wyoming. Am. Geol. Surv. Prof. Pap. 1966, 550-B, B20–B27. [Google Scholar]
- Bell, A.; Chiappe, L.M. Identification of a New Hesperornithiform from the Cretaceous Niobrara Chalk and Implications for Ecologic Diversity among Early Diving Birds. PLoS ONE 2015, 10, e0141690. [Google Scholar] [CrossRef] [Green Version]
- Lucas, F.A. Notes on the Osteology and Relationship of the Fossil Birds of the Genera Hesperornis, Hargeria, Baptornis, and Diatryma. Proc. U. S. Natl. Mus. 1903, 26, 545–556. [Google Scholar] [CrossRef]
- Elzanowski, A.; Paul, G.S.; Stidham, T.A. An Avian Quadrate from the Late Cretaceous Lance Formation of Wyoming. J. Vertebr. Paleontol. 2000, 20, 712–719. [Google Scholar] [CrossRef]
- Rees, J.; Lindgren, J. Aquatic Birds from the Upper Cretaceous (Lower Campanian) of Sweden and the Biology and Distribution of Hesperornithiforms: Cretaceous Aquatic Birds. Palaeontology 2005, 48, 1321–1329. [Google Scholar] [CrossRef]
- Shufeldt, R.W. The Fossil Remains of a Species of Hesperornis Found in Montana. Auk 1915, 32, 290–294. [Google Scholar] [CrossRef]
- Martin, L.D. A New Hesperornithid and the Relationships of the Mesozoic Birds. Trans. Kans. Acad. Sci. 1984, 87, 141–150. [Google Scholar] [CrossRef]
- Olson, S.L. Neogaeornis wetzeli Lambrecht, a cretaceous loon from Chile (Aves: Gaviidae). J. Vertebr. Paleontol. 1992, 12, 122–124. [Google Scholar] [CrossRef]
- Bell, A.K. Evolution and Ecology of Mesozoic Birds: A Case Study of the Derived Hesperornithiformes and the Use of Morphometric Data in Quantifying Avian Paleoecology. Master’s Thesis, The University of Southern California, Los Angeles, CA, USA, 2013. [Google Scholar]
- Chiappe, L.M. The first 85 million years of avian evolution. Nature 1995, 378, 349–355. [Google Scholar] [CrossRef]
- Chiappe, L.M.; Jorge, O.C. Neuquenornis volans, a new Late Cretaceous bird (Enantiornithes: Avisauridae) from Patagonia, Argentina. J. Vert. Paleont. 1994, 14, 230–246. [Google Scholar] [CrossRef]
- Padian, K.; Chiappe, L.M. The origin and early evolution of birds. Bio. Rev. 1998, 73, 1–42. [Google Scholar] [CrossRef]
- Liu, D.; Chiappe, L.M.; Zhang, Y.; Bell, A.; Meng, Q.; Ji, Q.; Wang, X. An advanced, new long-legged bird from the Early Cretaceous of the Jehol Group (northeastern China): Insights into the temporal divergence of modern birds. Zootaxa 2014, 3884, 253–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connor, J.K.; Zhou, Z. A redescription of Chaoyangia beishanensis (Aves) and a comprehensive phylogeny of Mesozoic birds. J. Syst. Palaeont. 2013, 11, 889–906. [Google Scholar] [CrossRef]
- O'Connor, J.K.; Chiappe, L.M.; Bell, A. Pre-modern birds: Avian divergences in the Mesozoic. In Living Dinosaurs: The Evolutionary History of Modern Birds; John Wiley & Sons: Hoboken, NJ, USA, 2011; pp. 39–114. [Google Scholar]
- Pittman, M.; O’Connor, J.; Tse, E.; Makovicky, P.; Field, D.J.; Ma, W.; Turner, A.H.; Norell, M.A.; Pei, R.; Xu, X. The Fossil Record of Mesozoic and Paleocene Pennaraptorans. Bull. Am. Mus. Nat. Hist. 2020, 440, 37–95. [Google Scholar]
- Wang, M.; O’Connor, J.K.; Zhou, Z. A new robust enantiornithine bird from the Lower Cretaceous of China with scansorial adaptations. J. Vert. Paleont. 2014, 34, 657–671. [Google Scholar] [CrossRef]
- Fluteau, F.; Ramstein, G.; Besse, J.; Giraud, R.; Masse, J.P. Impacts of palaeogeography and sea level change on Mid-Cretaceous climate. Palaeogeog. Palaeoclim. Palaeoeco. 2007, 247, 357–381. [Google Scholar] [CrossRef]
- Gradzinski, R.; Kielan-Jaworowska, Z.; Maryanska, T. Upper Cretaceous Djadokhta, Barun Goyot and Nemegt formations of Mongolia, including remarks on previous subdivisions. Acta Geol. Pol. 1977, 27, 281–326. [Google Scholar]
- Breithaupt, B. Paleontology and paleoecology of the Lance Formation (Maastrichtian), east flank of Rock Springs Uplift, Sweetwater County, Wyoming. Rocky Mt. Geol. 1982, 21, 123–151. [Google Scholar]
- Kupsch, W. Frenchman formation of eastern Cypress Hills, Saskatchewan, Canada. GSA Bull. 1957, 68, 413–420. [Google Scholar] [CrossRef]
- Ogunyomi, O.; Hills, L. Depositional environments, foremost formation (Late Cretaceous), Milk river area, southern Alberta. Bull. Can. Petrol. Geol. 1977, 25, 929–968. [Google Scholar]
- Wood, J.; Thomas, R.; Vasser, J. Fluvial processes and vertebrate taphonomy: The upper cretaceous Judith River formation, south-central dinosaur Provincial Park, Alberta, Canada. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1988, 66, 127–143. [Google Scholar] [CrossRef]
- Eberth, D.A. Stratigraphy and sedimentology of vertebrate microfossil sites in the uppermost Judith River formation (Campanian), Dinosaur Provincial Park, Alberta, Canada. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1990, 78, 1–36. [Google Scholar] [CrossRef]
- Johnson, K.R.; Nichols, D.J.; Hartman, J.H. Hell Creek formation: A 2001 synthesis. In The Hell Creek Formation and the Cretaceous-Tertiary Boundary in the Northern Great Plains: An Integrated Continental Record of the End of the Cretaceous; Geological Society of America Special Paper, 361; Hartman, J.H., Johnson, K.R., Nichols, D.J., Eds.; Geological Society of America: Boulder, CO, USA, 2002; pp. 503–510. [Google Scholar]
- Bardack, D. Fossil vertebrates from the marine Cretaceous of Manitoba. Can. J. Earth Sci. 1968, 5, 145–153. [Google Scholar] [CrossRef]
- Reifenstuhl, R.R. Gilead sandstone, northeastern Brooks Range, Alaska: An Albian to Cenomanian marine clastic succession. In Short Notes on Alaskan Geology; Professional Report; Reger, R.D., Ed.; Alaska Division of Geological & Geophysical Surveys: Fairbanks, AK, USA, 1991; Volume 111, pp. 69–76. [Google Scholar]
- Barlow, L.; Kauffman, E. Depositional cycles in the Niobrara formation, Colorado front range. In The Society of Economic Paleontologists and Mineralogists (SEPM) Fine-Grained Deposits and Biofacies of the Cretaceous Western Interior Seaway: Evidence of Cyclic Sedimentary Processes (FG4); AAPG: Tulsa, OK, USA, 1985. [Google Scholar]
- Russell, D. Cretaceous vertebrates from the Anderson River, N.W.T. Can. J. Earth Sci. 1967, 4, 21–43. [Google Scholar] [CrossRef]
- Panteleyev, A.V.; Popov, E.V.; Averianov, A.O. New record of Hesperornis rossicus (Aves, Hesperornithiformes) in the campanian of Saratov Province, Russia. Paleontol. Res. 2004, 8, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Hills, L.; Strong, W. Multivariate analysis of late cretaceous Kanguk formation (Arctic Canada) Palynomorph assemblages to identify nearshore to distal marine groupings. Bull. Can. Petrol. Geol. 2007, 55, 160–172. [Google Scholar] [CrossRef]
- Shultz, L.G.; Tourtelot, H.A.; Gill, J.R.; Boerngen, J.G. Composition and properties of the Pierre Shale and equivalent rocks, northern Great Plains region. Geol. Surv. Prof. Pap. 1980, 1064-B, 123. [Google Scholar]
- Hilton, R. Dinosaurs and Other Mesozoic Reptiles of California; University of California Press: Berkeley, CA, USA, 2003. [Google Scholar]
- Bell, A.; Irwin, K.J.; Davis, L.C. Hesperornithiform birds from the Late Cretaceous (Campanian) of Arkansas, USA. Trans. Kans. Acad. Sci. 2015, 118, 219–229. [Google Scholar] [CrossRef]
- Tanaka, T.; Kobayashi, Y.; Ikuno, K.; Ikeda, T.; Saegusa, H. A marine hesperornithiform (Avialae: Ornithuromorpha) from the Maastrichtian of Japan: Implications for the paleoecological diversity of the earliest diving birds in the end of the Cretaceous. Cretac. Res. 2020, 113, 104492. [Google Scholar] [CrossRef]
- Dyke, G.; Malakhov, D.; Chiappe, L.M. A re-analysis of the marine bird Asiahesperornis from northern Kazakhstan. Cretac. Res. 2006, 27, 947–953. [Google Scholar] [CrossRef]
- Marsh, O.C. A New Cretaceous Bird Allied to Hesperornis. Am. J. Sci. 1893, 45, 81–82. [Google Scholar] [CrossRef]
- Hinić-Frlog, S.; Motani, R. Relationship between osteology and aquatic locomotion in birds: Determining modes of locomotion in extinct Ornithurae. J. Evol. Bio. 2010, 23, 372–385. [Google Scholar] [CrossRef]
- Galantsev, V.P. Adaptational changes in the venous system of diving mammals. Can. J. Zool. 1991, 69, 414–419. [Google Scholar] [CrossRef]
- Girgis, S. Anatomical and functional adaptations in the venous system of a diving reptile, Trionyx triunquis. Proc. Zool. Soc. Lond. 1962, 138, 355–377. [Google Scholar] [CrossRef]
- Jessen, C. Selective brain cooling in mammals and birds. Jpn. J. Physiol. 2001, 51, 291–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinsamy, A.; Martin, L.D.; Dobson, P. Bone microstructure of the diving Hesperornis and the voltant Ichthyornis from the Niobrara Chalk of western Kansas. Cretac. Res. 1998, 19, 225–235. [Google Scholar] [CrossRef]
- Marsh, O.C. ART. XII. Characters of the Odontornithes, with notice of a new allied genus. Am. J. Sci. Arts 1877, 14, 79–84. [Google Scholar]
- Houde, P. Histological evidence for the systematic position of Hesperornis (Odontornithes: Hesperornithiformes). Auk 1987, 104, 125–129. [Google Scholar] [CrossRef]
- Wilson, L.E.; Chin, K.; Cumbaa, S.; Dyke, G. A High Latitude Hesperornithiform (Aves) from Devon Island: Palaeobiogeography and Size Distribution of North American Hesperornithiforms. J. Syst. Palaeontol. 2011, 9, 9–23. [Google Scholar] [CrossRef]
- Carpenter, K. Vertebrate biostratigraphy of the Smoky Hill Chalk (Niobrara Formation) and the Sharon Springs Member (Pierre Shale). In High-Resolution Approaches in Stratigraphic Paleontology; Springer: Dordrecht, The Netherlands, 2008; pp. 421–437. [Google Scholar]
- Petalas, C.; Lazarus, T.; Layoie, R.A.; Elliott, K.H.; Guigueno, M.F. Foraging niche partitioning in sympatric seabird populations. Sci. Res. 2021, 11, 2493. [Google Scholar] [CrossRef] [PubMed]
Figure 7.
Comparison of the humeri of hesperornithiform taxa: (A) Hesperornis FHSM VP-2293; (B) Parahesperornis KUVP 2287; (C) Baptornis KUVP 2290; and (D) Pasquiaornis RSM P2995.1 (from [15]); as well as the modern cormorants: (E) Phalacrocorax penicillatus (flighted); and (F) Nannopterum harrisi (flightless) in (1) dorsal; and (2) cranial views. Elements are scaled to be of a similar width across the distal condyles and aligned at the distal ends. Inset shows silhouettes of the same elements to scale. After Bell and Chiappe [25].
Figure 7.
Comparison of the humeri of hesperornithiform taxa: (A) Hesperornis FHSM VP-2293; (B) Parahesperornis KUVP 2287; (C) Baptornis KUVP 2290; and (D) Pasquiaornis RSM P2995.1 (from [15]); as well as the modern cormorants: (E) Phalacrocorax penicillatus (flighted); and (F) Nannopterum harrisi (flightless) in (1) dorsal; and (2) cranial views. Elements are scaled to be of a similar width across the distal condyles and aligned at the distal ends. Inset shows silhouettes of the same elements to scale. After Bell and Chiappe [25].

Figure 9.
Comparison of body size among Hesperornithidae taxa: (A) H. regalis YPM 1200; (B) H. chowi YPM PU 17208; (C) H. rossicus SGU 3442 Ve01; (D) H. gracilis, YPM 1473; (E) H. lumgairi CFDC B78.02.07; (F) H. bairdi YPM PU 17208a; (G) Asiahesperornis bazhanovi IZASK 5/287/86a; (H) H. mengeli CFDC 78.01.08; (I) Parahesperornis alexi KUVP 2287; (J) H. altus YPM 515; (K) H. macdonaldi LACM 9727. Silhouettes are approximately scaled to averages of measurements of the corresponding elements preserved in specimens of H. regalis.
Figure 9.
Comparison of body size among Hesperornithidae taxa: (A) H. regalis YPM 1200; (B) H. chowi YPM PU 17208; (C) H. rossicus SGU 3442 Ve01; (D) H. gracilis, YPM 1473; (E) H. lumgairi CFDC B78.02.07; (F) H. bairdi YPM PU 17208a; (G) Asiahesperornis bazhanovi IZASK 5/287/86a; (H) H. mengeli CFDC 78.01.08; (I) Parahesperornis alexi KUVP 2287; (J) H. altus YPM 515; (K) H. macdonaldi LACM 9727. Silhouettes are approximately scaled to averages of measurements of the corresponding elements preserved in specimens of H. regalis.

Figure 10.
Specimen completeness of hesperornithiform taxa. Insets show percentages of specimens that preserve: (A) elements belonging to different regions of the body; and (B) main elements of the hindlimb. After Bell and Chiappe [20].
Figure 10.
Specimen completeness of hesperornithiform taxa. Insets show percentages of specimens that preserve: (A) elements belonging to different regions of the body; and (B) main elements of the hindlimb. After Bell and Chiappe [20].

Figure 11.
Phylogenetic tree of hesperornithiform birds, after Bell and Chiappe [22], with the addition of Chupkaornis from Tanaka et al. [24]. Arrows indicate alternative positions for Enaliornis and Pasquiaornis and Baptornis and Brodavis in Tanaka et al. [24]. Diagrams indicate anatomical adaptations proposed to correlate with foot-propelled diving: (A) angled articular surface and cnemial expansion on the tibiotarsus; (B) reduced humerus; (C) expanded femoral trochanter; (D) fourth trochlea of the tarsometatarsus forms a sharp ridge; (E) enlarged, triangular patella; (F) expanded intercotylar eminence on the tarsometatarsus.
Figure 11.
Phylogenetic tree of hesperornithiform birds, after Bell and Chiappe [22], with the addition of Chupkaornis from Tanaka et al. [24]. Arrows indicate alternative positions for Enaliornis and Pasquiaornis and Baptornis and Brodavis in Tanaka et al. [24]. Diagrams indicate anatomical adaptations proposed to correlate with foot-propelled diving: (A) angled articular surface and cnemial expansion on the tibiotarsus; (B) reduced humerus; (C) expanded femoral trochanter; (D) fourth trochlea of the tarsometatarsus forms a sharp ridge; (E) enlarged, triangular patella; (F) expanded intercotylar eminence on the tarsometatarsus.

Figure 12.
Distribution of hesperornithiform specimens in geologic units and: (A) depositional environments; and (B) time periods within the Late Cretaceous. All formations from which fewer than five specimens are known are included in “other” for each depositional environment and time period. After Bell et al. [22].
Figure 12.
Distribution of hesperornithiform specimens in geologic units and: (A) depositional environments; and (B) time periods within the Late Cretaceous. All formations from which fewer than five specimens are known are included in “other” for each depositional environment and time period. After Bell et al. [22].


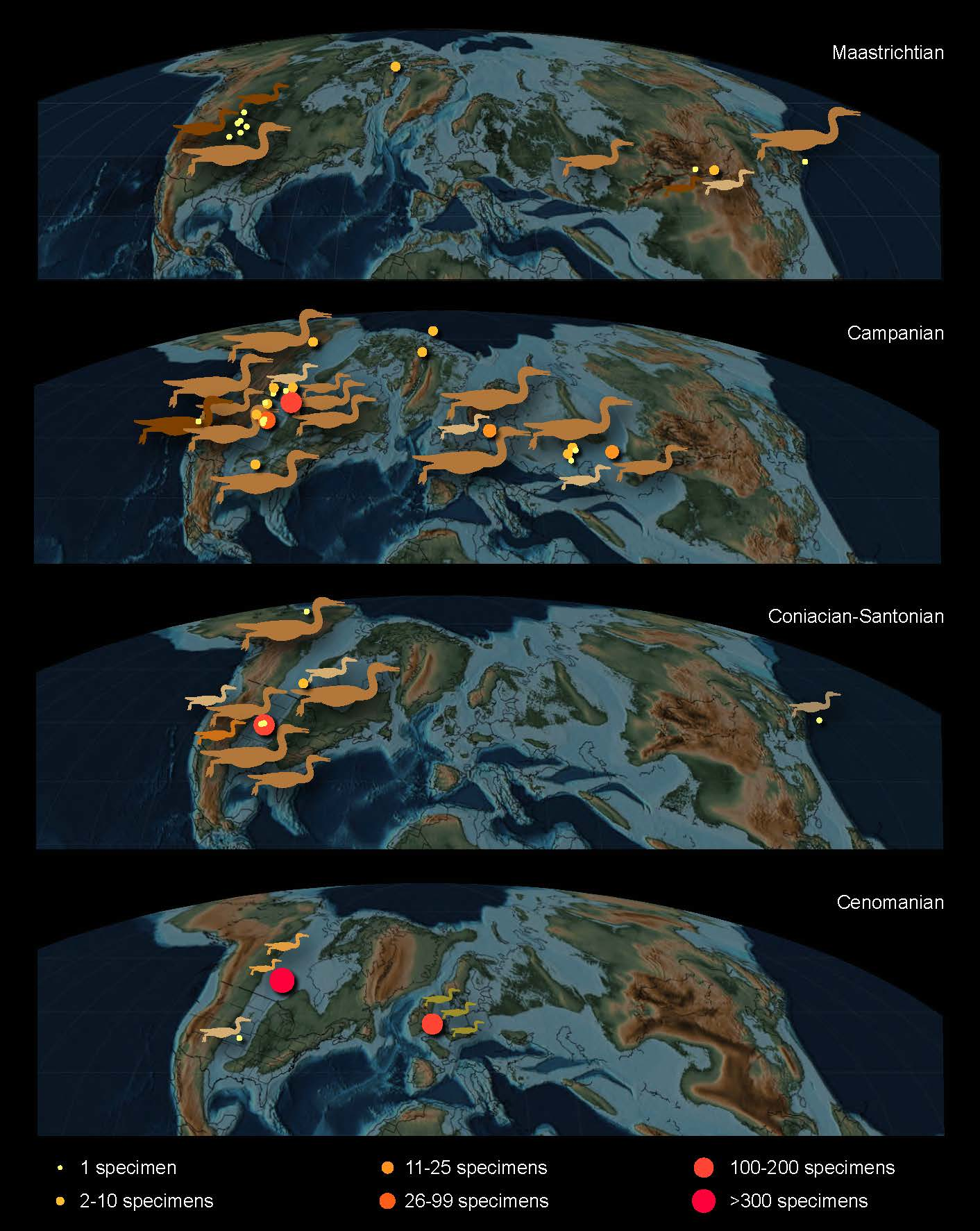{kind=link}
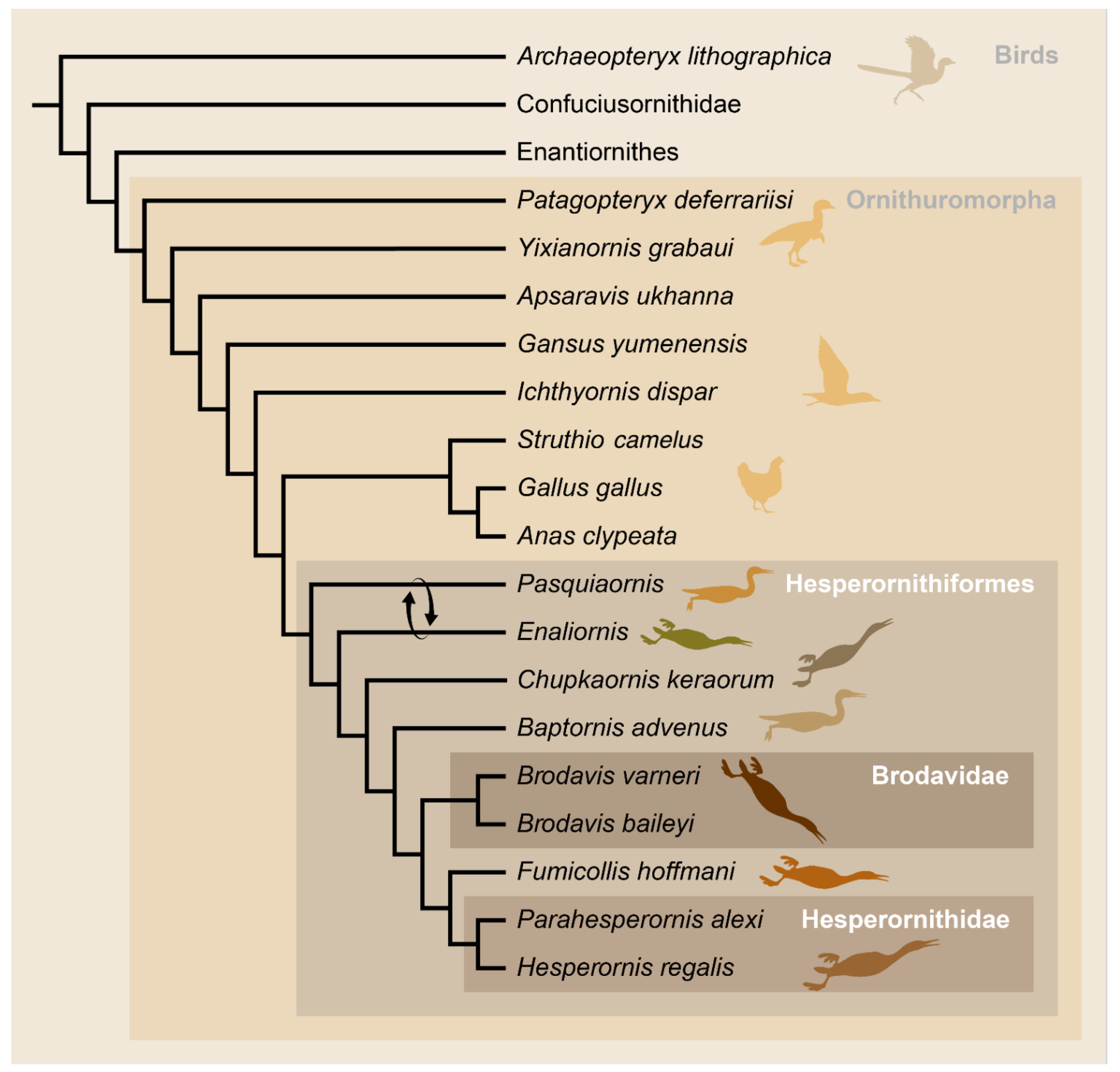{kind=link}
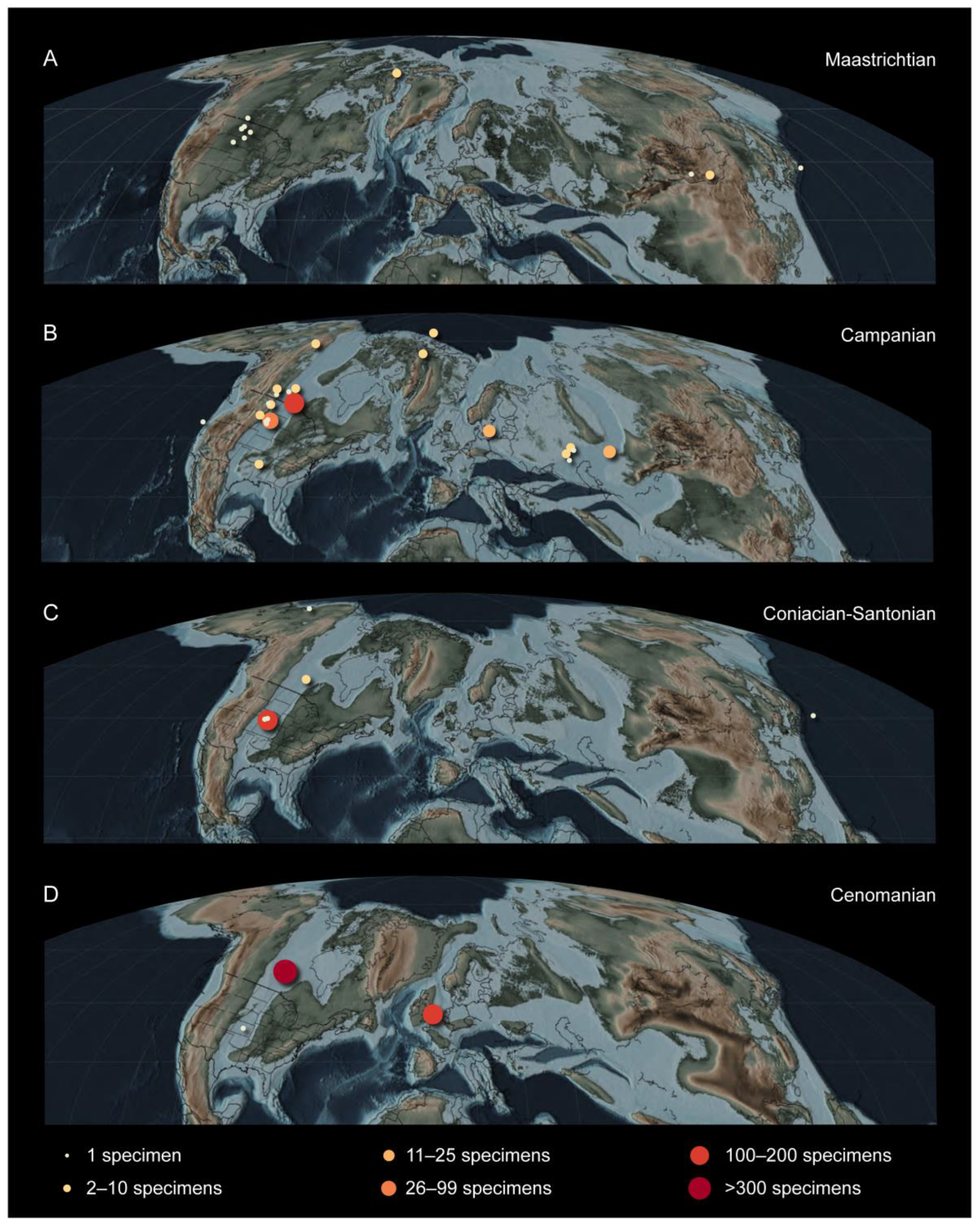{kind=link}
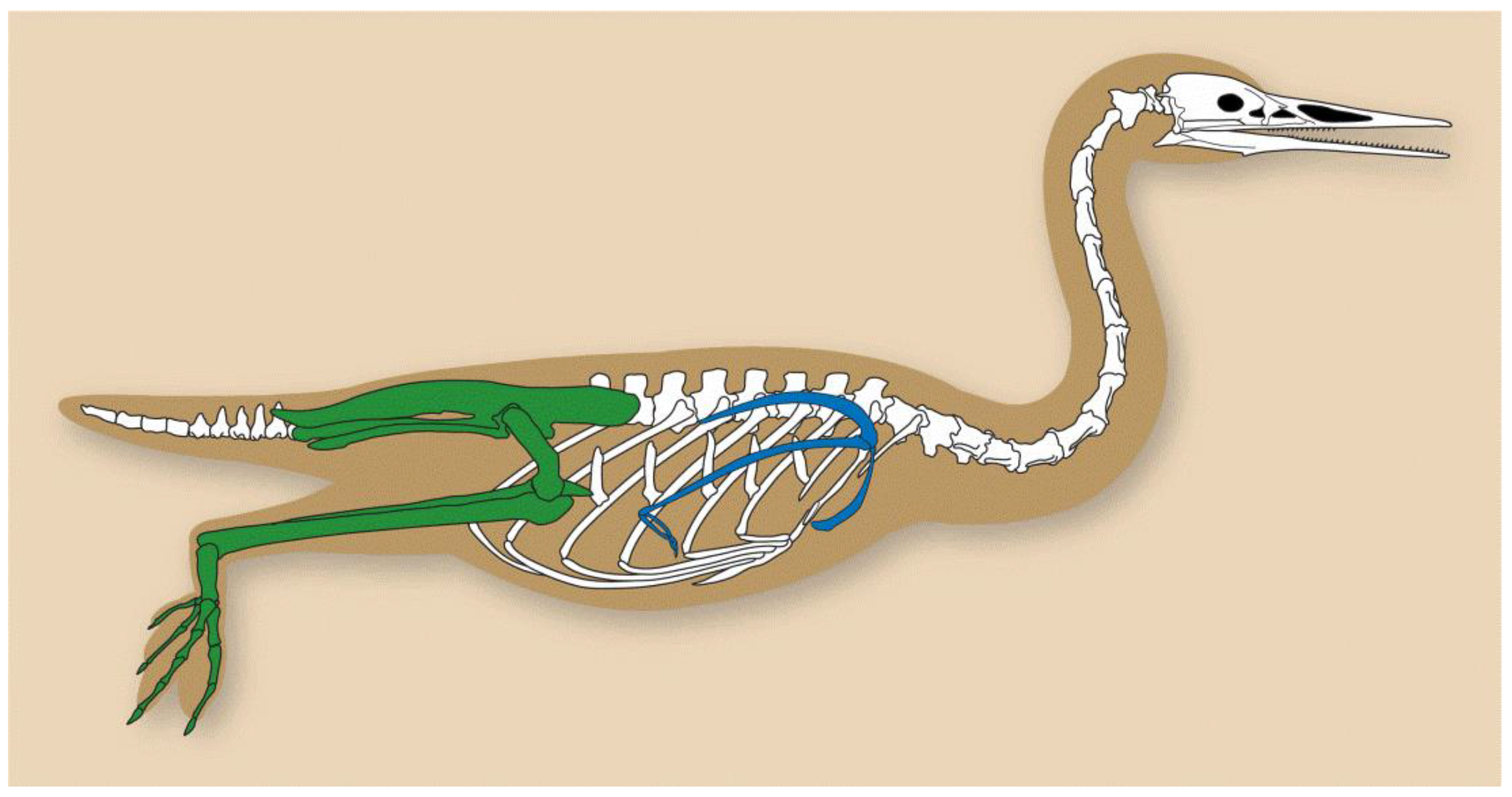{kind=link}
{kind=link}
{kind=link}
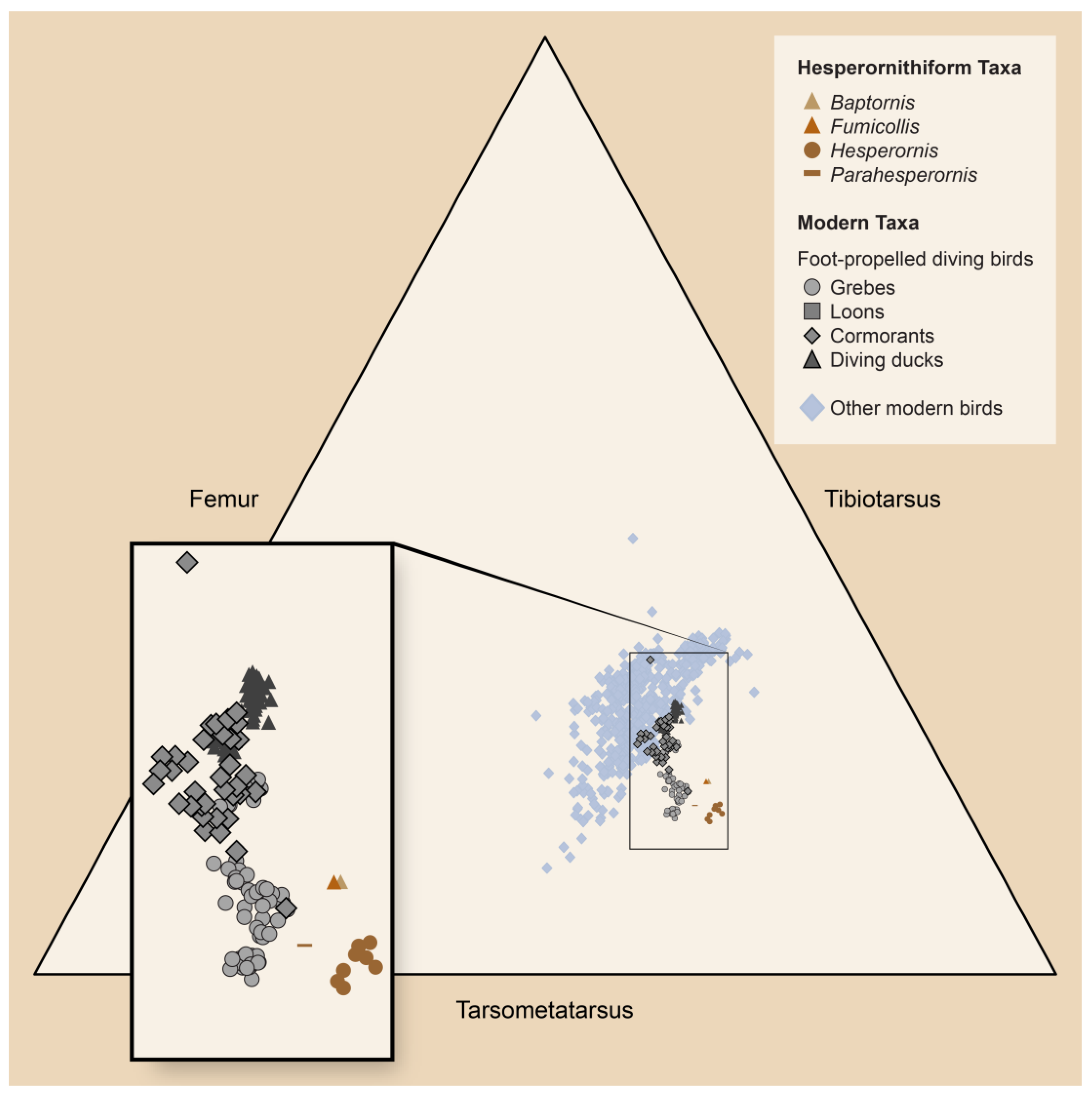{kind=link}
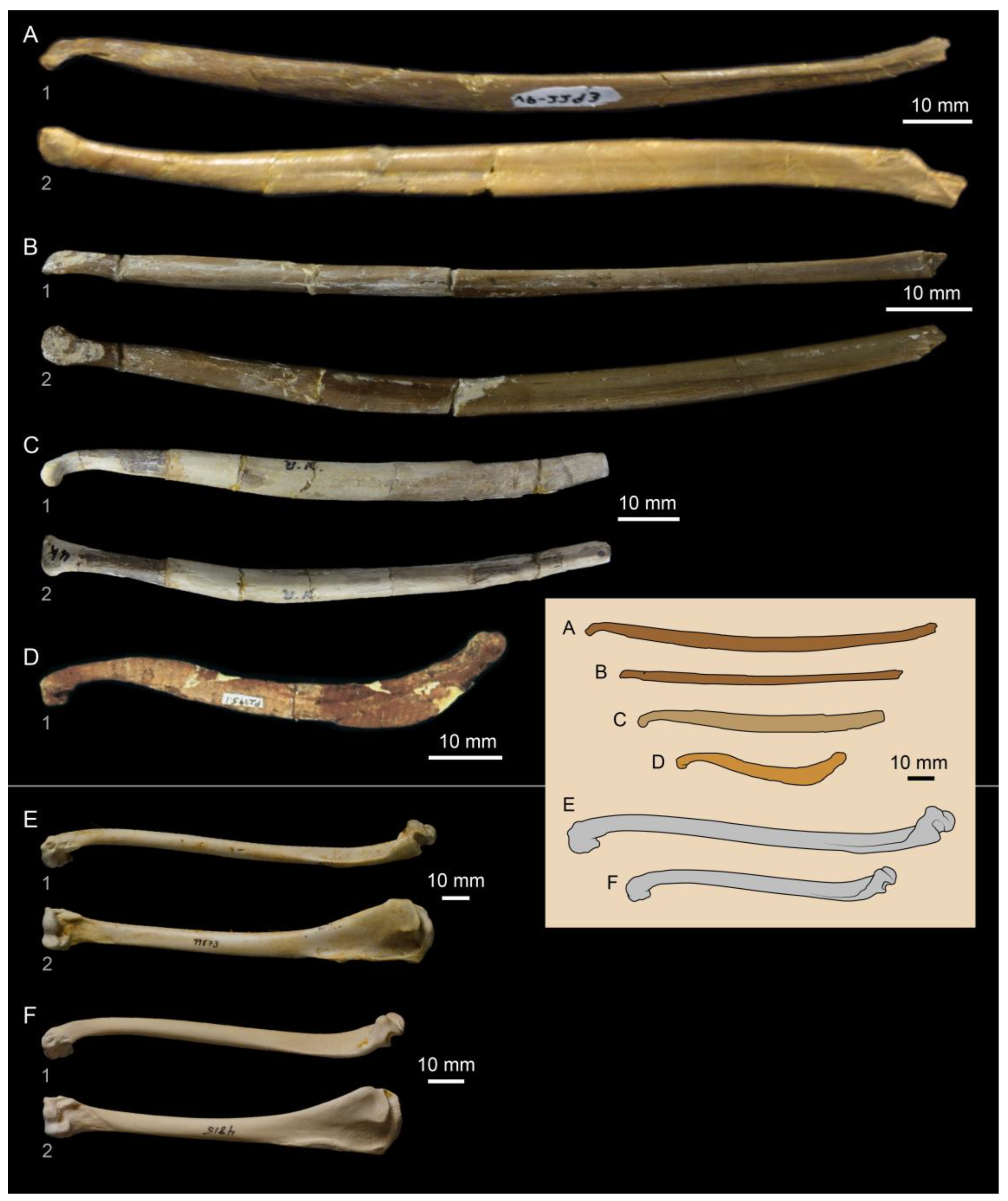{kind=link}
{kind=link}
{kind=link}
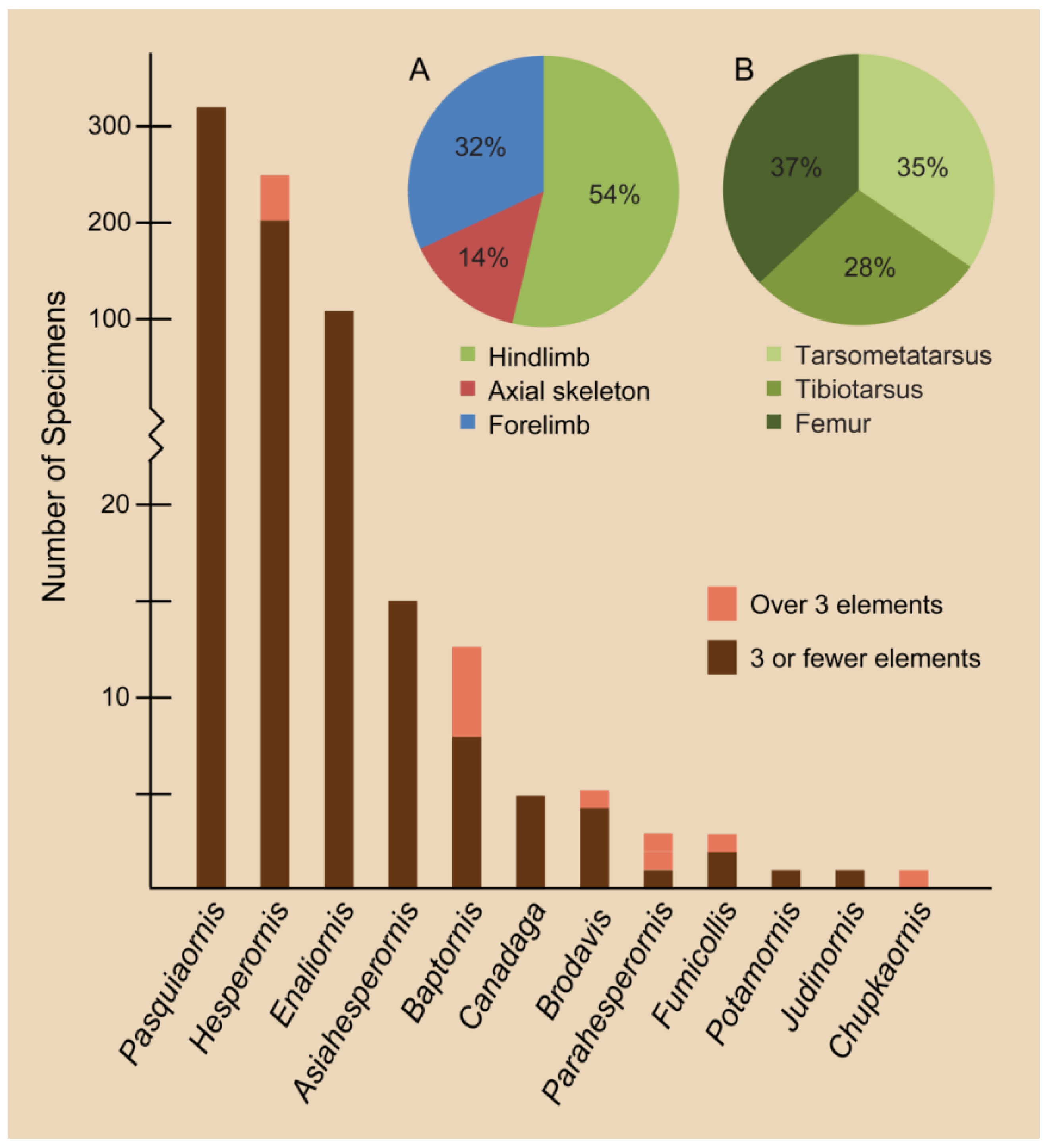{kind=link}
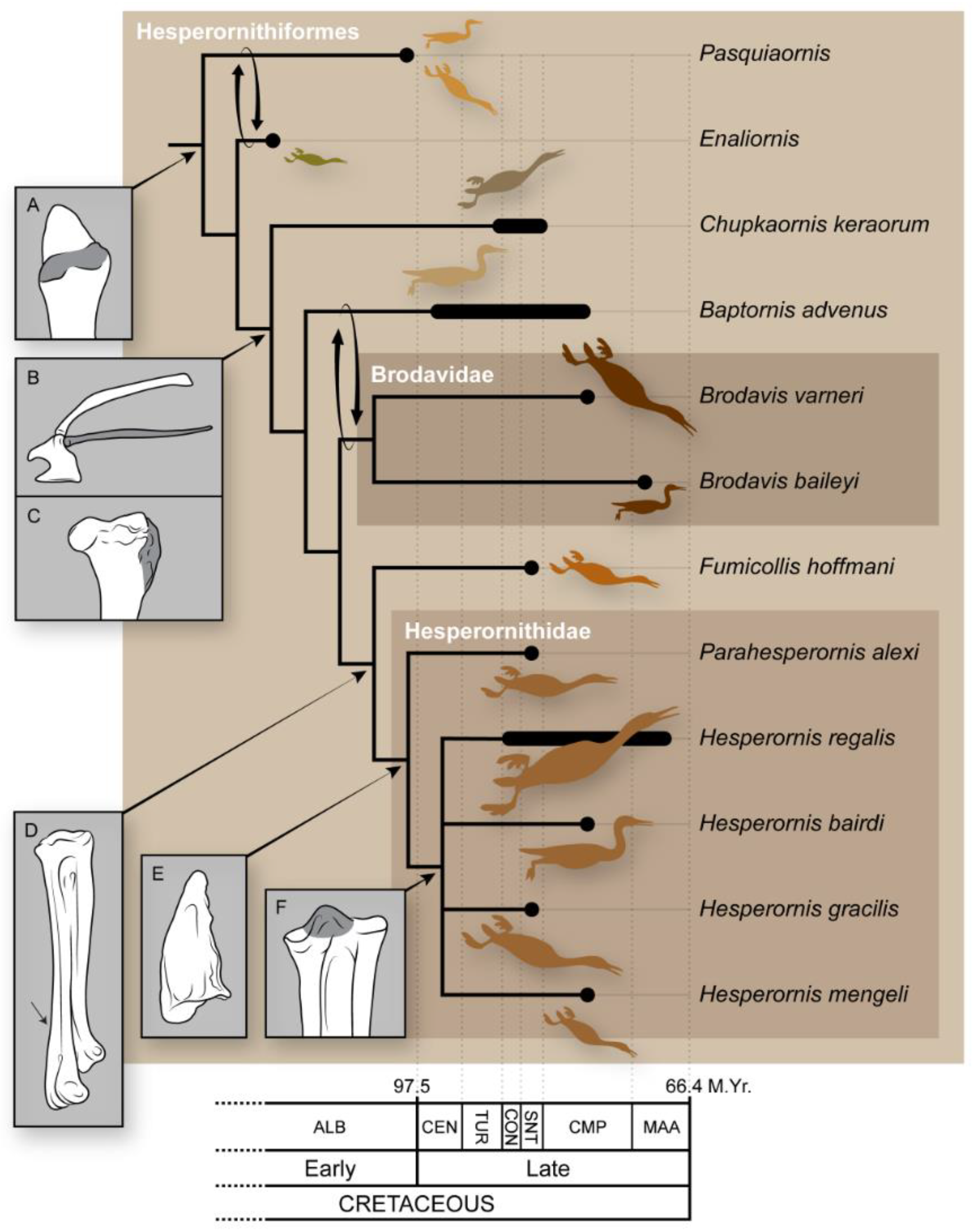{kind=link}
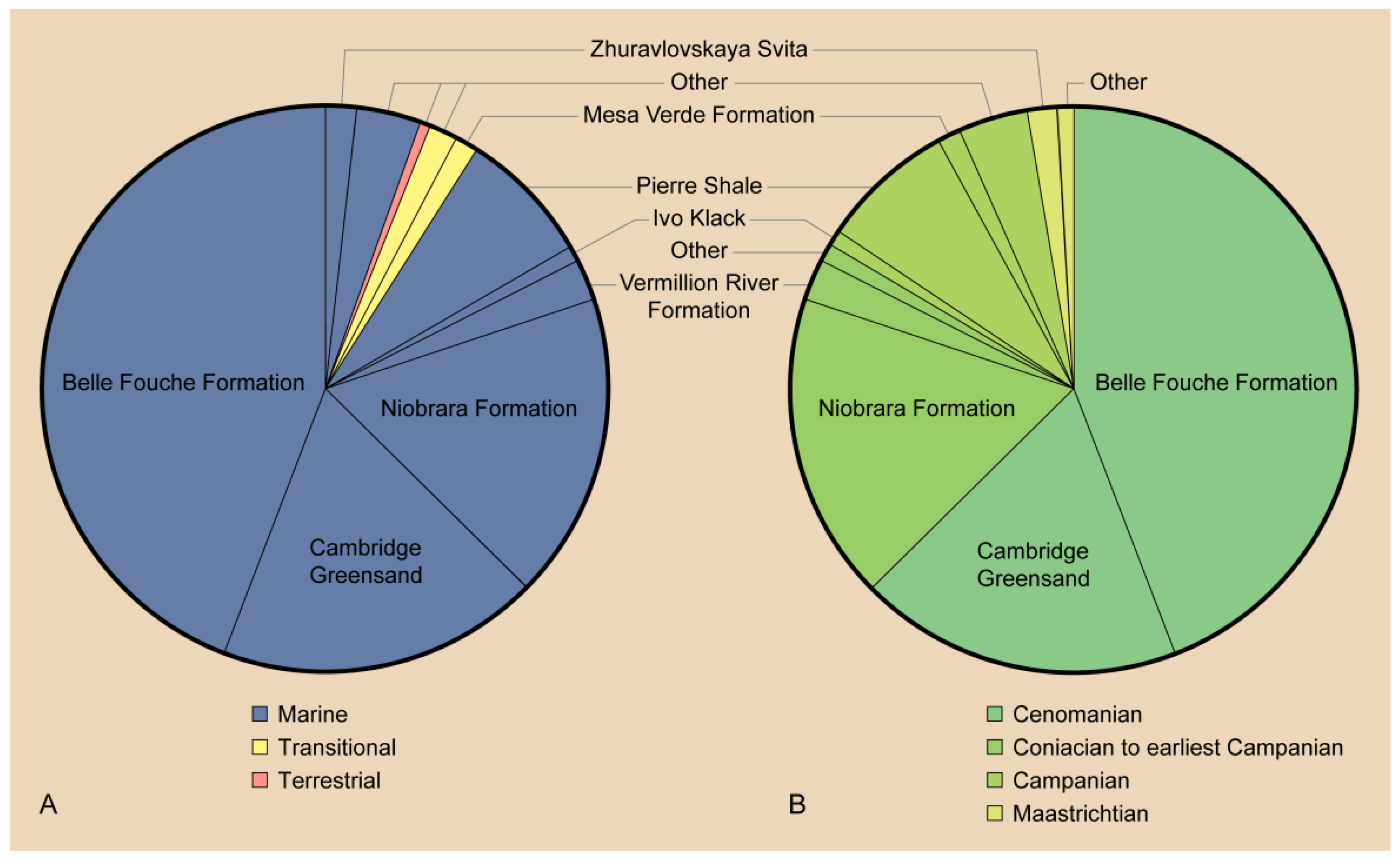{kind=link}
Table 1.
Current taxonomic framework of the Hesperornithiformes, with taxa that have been previously invalidated shown in red.
Table 1.
Current taxonomic framework of the Hesperornithiformes, with taxa that have been previously invalidated shown in red.
| Class | Family | Genus | Species |
|---|---|---|---|
| Hesperornithiformes | Enaliornithidae | Enaliornis | barretti |
| sedgewicki | |||
| seeleyi | |||
| Baptornithidae | Baptornis | advenus | |
| varneri | |||
| Judinornis | nogontsavensis | ||
| Parascaniornis | stensoei | ||
| Brodavidae | Brodavis | americanus | |
| baileyi | |||
| mongoliensis | |||
| varneriv | |||
| Hesperornithidae | Asiahesperornis | bazhanoviv | |
| Canadaga | arctica | ||
| Coniornis | altus | ||
| Hargeria | gracilis | ||
| Hesperornis | altus | ||
| bairdi | |||
| chowi | |||
| crassipes | |||
| gracilis | |||
| macdonaldi | |||
| mengeli | |||
| montana | |||
| regalis | |||
| rossicus | |||
| Lestornis | crassipes | ||
| Parahesperornis | alexi | ||
| NA | Pasquiaornis | hardei | |
| tankei | |||
| NA | Chupkaornis | keraorum | |
| NA | Fumicollis | hoffmani | |
| NA | Potamornis | skutchi |
Table 2.
Geologic units, divided by depositional environment, with published hesperornithiform taxa. After Bell et al. [22].
Table 2.
Geologic units, divided by depositional environment, with published hesperornithiform taxa. After Bell et al. [22].
| Continental | ||
|---|---|---|
| Mesa Verde Formation (Teapot Sandstone) [57] | Hesperornis regalis | Campanian |
| Hesperornis sp. | ||
| Nemegt Formation [75] | Brodavis mongoliensis | Maastrichtian |
| Judinornis nogontsavensis | ||
| Hesperornithidae indet. | ||
| Lance Formation [76] | Potamornis skutchi | Late Maastrichtian |
| Frenchman Formation [77] | Brodavis americanus | Late Maastrichtian |
| Transitional | ||
| Foremost Formation [78] | Hesperornis sp. | Campanian |
| Judith River Formation [79] | Baptornis sp. | Campanian |
| Hesperornis altus | ||
| Dinosaur Provincial Park Formation [80] | Baptornis sp. | Late Campanian |
| Hell Creek Formation [81] | Brodavis baileyi | Maastrichtian |
| Hesperornis sp. | ||
| Marine | ||
| Cambridge Greensand Member (West Melbury Chalk Formation) [44] | Enaliornis barretti | Early Cenomanian |
| Enaliornis seeleyi | ||
| Enaliornis sedgewicki | ||
| Belle Fouche Formation (formerly Ashville Formation) [14] | Pasquiaornis hardiei | Late Cenomanian |
| Pasquiaornis tankei | ||
| Greenhorn Formation [48] | Baptornis sp. | Cenomanian |
| Kashima Formation [24] | Chupkaornis keraorum | Coniacian to Santonian |
| Vermillion River Formation [82] | Hesperornis regalis | Coniacian to Santonian |
| Hesperornis sp. | ||
| Ignek Formation [83] | Hesperornis sp. | Late Coniacian to Campanian |
| Smoky Hill Chalk, Niobrara Formation [84] | Baptornis advenus | Late Coniacian to early Campanian |
| Hesperornis crassipes | ||
| Hesperornis gracilis | ||
| Hesperornis regalis | ||
| Hesperornis sp. | ||
| Fumicollis hoffmani | ||
| Parahesperornis alexi | ||
| Parahesperornis sp. | ||
| Smoking Hills Formation [85] | Hesperornis regalis | Middle Santonian to early late Campanian |
| Eginsaiskaya [49] | Baptornis advenus | Latest Santonian to Early Campanian |
| Asiahesperornis bazhanovi | ||
| Rybushka Formation [86] | Hesperornis rossicus | Early Campanian |
| Hesperornis sp. | ||
| Kristianstad Basin (unreported formation) [61] | Baptornis sp. | Latest early Campanian |
| Hesperornis rossicus | ||
| Hesperornis sp. | ||
| Kanguk Formation [87] | Canadaga arctica | Early to middle Campanian |
| Hesperornis sp. | ||
| Clagget Shale [57] | Hesperornis sp. | Campanian |
| Pierre Shale [88] | Baptornis advenus | Campanian |
| Brodavis varneri | ||
| Hesperornis bairdi | ||
| Hesperornis chowi | ||
| Hesperornis lumgairi | ||
| Hesperornis macdonaldi | ||
| Hesperornis mengeli | ||
| Hesperornis regalis | ||
| Hesperornis rossicus | ||
| Hesperornis sp. | ||
| Chico Formation [89] | Hesperornis sp. | Campanian |
| Ozan Formation [90] | Hesperornis sp. | Campanian |
| Kita-ama Formation [91] | Hesperornithiformes undet. | Early Maastrichtian |
| Zhuravlovskaya Svita [92] | Asiahesperornis bazhanovi | Maastrichtian |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bell, A.; Chiappe, L.M. The Hesperornithiformes: A Review of the Diversity, Distribution, and Ecology of the Earliest Diving Birds. Diversity 2022, 14, 267. https://0-doi-org.brum.beds.ac.uk/10.3390/d14040267
AMA Style
Bell A, Chiappe LM. The Hesperornithiformes: A Review of the Diversity, Distribution, and Ecology of the Earliest Diving Birds. Diversity. 2022; 14(4):267. https://0-doi-org.brum.beds.ac.uk/10.3390/d14040267
Chicago/Turabian StyleBell, Alyssa, and Luis M. Chiappe. 2022. "The Hesperornithiformes: A Review of the Diversity, Distribution, and Ecology of the Earliest Diving Birds" Diversity 14, no. 4: 267. https://0-doi-org.brum.beds.ac.uk/10.3390/d14040267
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.