
Phylogenetic Analyses of Cyprinid Species from the Rokel River Basin of Sierra Leone, West Africa: Taxonomic, Biogeographic, and Conservation Implications


Abstract
:1. Introduction
2. Material and Methods
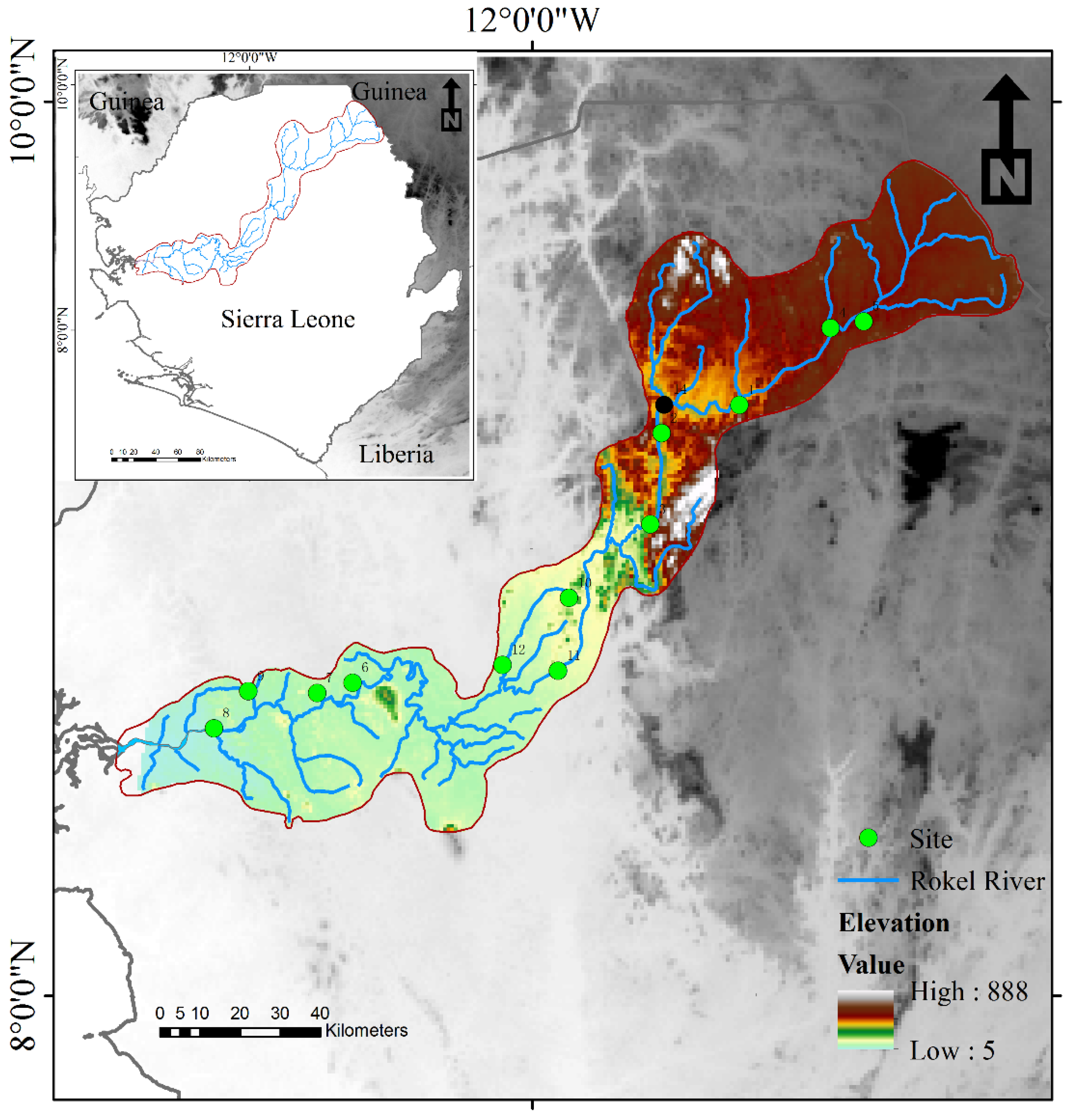
2.1. Study Area
2.2. Data Analysis
2.3. Phylogenetic Analysis
2.4. Species Delineation
3. Results
4. Discussion
4.1. Misidentification and Unrecognized Diversity
4.2. Genetic Placement of Prolabeo Batesi
4.3. Biogeographic Implication
4.4. Endemism and Conservation Implications
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| RR | Rokel River basin |
| IHB-CAS | Institute of Hydrobiology, Chinese Academy of Sciences |
| ASAP | Assemble Species by Automatic Partitioning |
| bPTP | Poisson Tree Processes |
References
- Payne, A.L. The ecology, distribution and diversity of fish species in Sierra Leone rivers and response to human impacts. Environ. Biol. Fishes 2018, 101, 843–864. [Google Scholar] [CrossRef]
- Wilson, C.; Liang, B.; Wilson, S.; Akiwumi, F. Land use, microclimate, and surface runoff linkages: Space-Time modeling from Rokel-Seli river basin, Sierra Leone. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2018, 42, 225–230. [Google Scholar] [CrossRef] [Green Version]
- Paugy, C.; Lévêque, D.; Teugels, G.G. The Fresh and Brackish Water Fishes of West Africa; Royal Museum for Central Africa: Tervuren, Belgium, 2003; Volume 148, pp. 148–162. [Google Scholar]
- Paugy, D.; Leveque, C.; Teugels, G.G.; Bigorne, R.; Romand, R. Freshwater Fishes of Sierra Leone and Liberia Annotated Checklist and Distribution. Rev. Hydrobiol. Trop. 1990, 23, 329–350. [Google Scholar]
- Roberts, T.R. Geographical distribution of African freshwater fishes. Zool. J. Linn. Soc. 1975, 4, 9–15. [Google Scholar] [CrossRef]
- Carr, J.; Adeleke, A.; Angu, K.A.; Belle, E.; Burgess, N.; Carrizo, S.; Choimes, A.; Coulthard, N.; Darwall, W.; Foden, W.; et al. Ecosystem Profile. Guinean Forests of West Africa Biodiversity Hotspot. Available online: https://www.cepf.net/sites/default/files/en_guinean_forests_ecosystem_profile.pdf (accessed on 16 December 2021).
- Schmidt, R.C.; Bart, H.L.; Pezold, F.; Friel, J.P. A Biodiversity Hotspot Heats Up: Nine New Species of Suckermouth Catfishes (Mochokidae: Chiloglanis) from Upper Guinean Forest Streams in West Africa. Copeia 2017, 105, 301–338. [Google Scholar] [CrossRef]
- Schmidt, R.C. Historical Biogeography of Fishes of the Fouta Djallon Highlands and Surrounding Areas. Ph.D Thesis, Tulane University, New Orleans, LA, USA, September 2014. [Google Scholar]
- Schmidt, R.C.; Dillon, M.N.; Kuhn, N.M.; Bart, H.L.; Pezold, F. Unrecognized and Imperilled Diversity in an Endemic Barb (Smiliogastrini, Enteromius) from the Fouta Djallon Highlands. Zool. Scr. 2019, 48, 605–613. [Google Scholar] [CrossRef]
- Schmidt, R.; Bart, H.; Pezold, F. High Levels of Endemism in Suckermouth Catfishes (Mochokidae: Chiloglanis) from the Upper Guinean Forests of West Africa. Mol. Phylogenet. Evol. 2016, 100, 199–205. [Google Scholar] [CrossRef]
- Skelton, P.H. New species of the amphiliid catfish genera Amphilius, Doumea and Phractura and the taxonomy of Paramphilius from West Central Africa (Siluriformes, Amphiliidae). Zootaxa 2007, 1578, 41–68. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, R.C.; Bart, H.L.; Nyingi, W.D. Integrative taxonomy of the red-finned barb, Enteromius apleurogramma (Cyprininae: Smiliogastrini) from Kenya, supports recognition of E. amboseli as a valid species. Zootaxa 2018, 4482, 566–578. [Google Scholar] [CrossRef]
- Decru, E.; Moelants, T.; De Gelas, K.; Vreven, E.; Verheyen, E.; Snoeks, J. Taxonomic Challenges in Freshwater Fishes: A Mismatch between Morphology and DNA Barcoding in Fish of the North-Eastern Part of the Congo Basin. Mol. Ecol. Resour. 2016, 16, 342–352. [Google Scholar] [CrossRef]
- Van Ginneken, M.; Decru, E.; Verheyen, E.; Snoeks, J. Morphometry and DNA Barcoding Reveal Cryptic Diversity in the Genus Enteromius (Cypriniformes: Cyprinidae) from the Congo Basin, Africa. Eur. J. Taxon. 2017, 310, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, R.C.; Pezold, F. Morphometric and molecular variation in mountain catfishes (Amphiliidae: Amphilius) in Guinea, West Africa. J. Nat. Hist. 2011, 45, 521–552. [Google Scholar] [CrossRef]
- Leveque, C. Fishes of African Continental Waters: Diversity, Ecology, Human Activities; IRD Editions: Paris, France, 1999; p. 521. [Google Scholar]
- IUCN Red List Committee. The IUCN Red List of Threatened Species™ Strategic Plan 2013–2020. Available online: https://www.iucn.org/sites/dev/files/import/downloads/red_list_strategic_plan_2013_2020.pdf (accessed on 25 November 2021).
- Magalhães FD, M.; Lyra, M.L.; De Carvalho, T.R.; Baldo, D.; Brusquetti, F.; Burella, P.; Colli, G.R.; Gehara, M.C.; Giaretta, A.A.; Haddad, C.F.B.; et al. Taxonomic Review of South American Butter Frogs: Phylogeny, Geographic Patterns, and Species Delimitation in the Leptodactylus latrans Species Group (Anura: Leptodactylidae). Herpetol. Monogr. 2020, 34, 131–177. [Google Scholar] [CrossRef]
- Yang, L.; Sado, T.; Hirt, M.V.; Pasco-Viel, E.; Arunachalam, M.; Li, J.; Wang, X.; Freyhof, J.; Saitoh, K.; Simons, A.M.; et al. Phylogeny and Polyploidy: Resolving the Classification of Cyprinine Fishes (Teleostei: Cypriniformes). Mol. Phylogenet. Evol. 2015, 85, 97–116. [Google Scholar] [CrossRef] [PubMed]
- Hayes, M.M.; Armbruster, J.W. The Taxonomy and Relationships of the African Small Barbs (Cypriniformes: Cyprinidae). Copeia 2017, 105, 348–362. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Ribeiro, N.; Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large data sets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Xia, X. DAMBE5: A comprehensive software package for data analysis in molecular biology and evolution. Mol. Biol. Evol. 2013, 30, 1720–1728. [Google Scholar] [CrossRef] [Green Version]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Bauldry, S. Structural Equation Modeling, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2015; Volume 22, ISBN 9780080970875. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. Partitionfinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian Inference of Phylogenetic Trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, R.D.; Hanner, R.; Hebert, P.D.N. The Campaign to DNA Barcode All Fishes, FISH-BOL. J. Fish Biol. 2009, 74, 329–356. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; DeWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Maetens, H.; Van Steenberge, M.; Snoeks, J.; Decru, E. Revalidation of Enteromius Alberti and Presence of Enteromius Cf. Mimus (Cypriniformes: Cyprinidae) in the Lake Edward System, East Africa. Eur. J. Taxon. 2020, 2020, 1–28. [Google Scholar] [CrossRef]
- Mayr, E.; O’Hara, R.J. The Biogeographic Evidence Supporting the Pleistocene Forest Refuge Hypothesis. Evolution 1986, 40, 55–67. [Google Scholar] [CrossRef]
- Brailsford, L.E. Evidence for Genetic Decline within Afromontane Forest Fragments on the Mambilla Plateau, Nigeria. Master’s Thesis, University of Canterbury, Christchurch, New Zealand, March 2018; pp. 1–114. [Google Scholar]
- Vences, M.; Marty, C.; Blanc, M.; Gemmell, N.J.; Fouquet, A. Underestimation of Species Richness in Neotropical Frogs Revealed by mtDNA Analyses. PLoS ONE 2007, 2, e1109. [Google Scholar] [CrossRef]
- Dankwa, H.; Entsua-Mensah, M. Enteromius liberiensis. The IUCN Red List of Threatened Species 2020: E.T182865A126383820. Available online: https://0-dx-doi-org.brum.beds.ac.uk/10.2305/IUCN.UK.2020-3.RLTS.T182865A126383820.en (accessed on 15 January 2020).
- Funk, W.C.; Caminer, M.; Ron, S.R. High levels of cryptic species diversity uncovered in Amazonian frogs. Proc. R. Soc. B Biol. Sci. 2012, 279, 1806–1814. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Species | Specimens ID | Countries | Locality/River | GenBank No. | Year Collected |
|---|---|---|---|---|---|
| Enteromius anema | AUF5493 | Guinea | Bafing | MF135226.1 | Hayes and Armbruster 2017 |
| Enteromius anema | AUF5494 | Guinea | Bafing | MF135225.1 | |
| Enteromius ablabes | Unknown | Ivory Coast | Agnebi | AF180835.1 | Tsigenopoulos et al., 1999 |
| Enteromius macrops | AUF 5524 | Guinea | Forécariah | MF135212.1 | |
| Enteromius macrops | Gui048 | Guinea | Forécariah | MF135201.1 | |
| Enteromius macrops | Gui0237 | Guinea | Forécariah | MF135200.1 | |
| Enteromius macrops | AUF5481 | Guinea | Safa-Khoure | MF135210.1 | |
| Enteromius macrops | Guinea | Koumba | MF135203.1 | ||
| Enteromius macrops | Guinea | Koumba | MF135202.1 | ||
| Enteromius macrops | AUF5454 | Guinea | Tinkisso | MF135204.1 | |
| Enteromius macrops | Guinea | Tinkisso | MF135205.1 | ||
| Enteromius macrops | Guinea | Kolenté | MF135208.1 | ||
| Enteromius macrops | AUF5476 | Guinea | Kolenté | MF135206.1 | |
| Enteromius macrops | Guinea | Kolenté | MF135209.1 | ||
| Enteromius macrops | Guinea | Kolenté | MF135207.1 | ||
| Enteromius macrops | Guinea | Konkouré | AF180832.1 | ||
| Enteromius macrops | Guinea | Doulou | MF135211.1 | ||
| Enteromius camptacanthus | Unknown | Ghana | Lake Volta | KF791270.1 | |
| Enteromius anema | Unknown | Nilo-Sudan | Blue Nile | KP712159.1 | |
| Enteromius guildi | AUF5505 | Guinea | Zie | MF135218.1 | |
| Enteromius cadenati | Unknown | Sierra Leone | Taia/Jong | AF180834.1 | |
| Enteromius cadenati | AUF5364 | Guinea | Dimmah | MF135224.1 | |
| Enteromius liberiensis | AUF5483 | Guinea | Safa-Khoure | MF135213.1 | |
| Enteromius bigornei | MNCN: 46CK | Guinea | Kaba, Kouloundela | AY004752.1 | Machordom and Doadrio 2001 |
| Enteromius aspilus | Unknown | Guinea | Konkouré | KF791275.1 | Schmidt et al., 2019 |
| Enteromius foutensis | Gui0858 | Guinea | Little Scarcies | MK329230.1 | |
| Enteromius foutensis | Gui0146 | Guinea | Little Scarcies | MK329231.1 | |
| Enteromius foutensis | Gui0167 | Guinea | Little Scarcies | MF135220.1 | |
| Enteromius foutensis | Gui0168 | Guinea | Little Scarcies | MF135219.1 | |
| Enteromius foutensis | Gui3435 | Guinea | Konkouré | MK329229.1 | |
| Enteromius foutensis | Gui3494 | Guinea | Konkouré | MK329228.1 | |
| Enteromius foutensis | Gui0145 | Guinea | Konkouré | MK329227.1 | |
| Enteromius foutensis | Gui_0206 | Guinea | Konkouré | MK329233.1 | |
| Enteromius foutensis | Gui0146 | Guinea | Konkouré | MK329226.1 | |
| Enteromius foutensis | Gui 0146 | Guinea | Konkouré | MK329225.1 | |
| Enteromius foutensis | Gui_0204 | Guinea | Konkouré | MK329232.1 | |
| Enteromius foutensis | Gui 0167 | Guinea | Konkouré | MK329224.1 | |
| Enteromius foutensis | Gui 0018 | Senegal | Gambie/Senegali | MK329241.1 | |
| Enteromius foutensis | Gui 0133 | Senegal | Gambie/Senegali | MK329240.1 | |
| Enteromius cf. guildi | AUF5443 | Guinea | Bafing | MF135223.1 | Hayes and Armbruster 2017 |
| Enteromius ablabes | AUF5431 | Guinea | Bafing | MF135227.1 | |
| Enteromius ablabes | AUF5441 | Guinea | Bafing | MF135228.1 | |
| Enteromius trispilos | AUF5496 | Guinea | Mia | MF135193.1 | Hayes and Armbruster 2017 |
| Enteromius trispilos | AUF5498 | Guinea | Mia | MF135194 | |
| Enteromius anema | AUF5493 | Guinea | Mia | MF135225.1 | |
| Enteromius anema | AUF5494 | Guinea | Mia | MF135226.1 | |
| Enteromius huguenyi | AUF5589 | Guinea | Masseni | MF135214.1 | |
| Enteromius punctitaeniatus | AUF5610 | Guinea | Mafou | MF135199.1 | |
| Enteromius profundus | DNG_PROF2_2 | Kenya | Kisumu | MH484558.1 | Ndeda, Mateos, and Hurtado 2018 |
| Enteromius profundus | DNG_PROF4_4 | Kenya | Kisumu | MH484556.1 | |
| Enteromius callipterus | Unknown | Gabon | Loa -Loa | AP009313.1 | Saitoh 2006 |
| Enteromius callipterus | CBM-ZF-11498 | Gabon | Loa -Loa | KP712230.1 | |
| Labeobarbus sacratus | MNCN 4CK | Guinea | Tangala | AF287445.1 | Tsigenopoulos, Naran, and Berrebi 1999 |
| Enteromius tiekoroi | UAIC14166.05 | Sierra Leone | Mao | KP659410.1 | Yang et al., 2015 |
| Labeobarbus sacratus | Guinea | Tangala | AF180868.1 | Tsigenopoulos, Naran, and Berrebi 1999 | |
| Labeobarbus sacratus | AF287445.1 | ||||
| Labeobarbus wurtzi | MNCN 92CK | Guinea | Kouloundela | AF287448.1 | |
| Labeobarbus wurtzi | MNCN 91CK | Guinea | Kaba, | AF180864.1 | |
| Labeo forskalii | CU 94562 | Ethiopia | Alwero | JX074287.1 | Yang and Mayden 2012 |
| Labeo forskalii | UAIC14744.4 | Ethiopia | Alwero | FJ196833.1 | Beshera and Phillip 2019 |
| Labeobarbus cyclorhynchus | CBM ZF 11452 | AP011359.1 | Miya 2009 | ||
| Labeo forskalii | AAU:0512009 | Ethiopia | Alwero | FJ196831.1 | Tang, Getahun, and Liu 2009 |
| Labeo lukulae | DRC | Lukula | JX097084.1 | Hirt 2012 | |
| Labeo parvus | BMNH:2006.3.7.1 | Benin | bei Malauville | JX074292.1 | Yang and Mayden 2012 |
| Labeo parvus | CBM: ZF: 12695 | Ethiopia | Alwero | AP013339.1 | Beshera and Phillip 2019 |
| Labeo parvus | Ethiopia | Baro | JX074285.1 | Yang and Mayden 2012 | |
| Labeo parvus | Ethiopia | Baro | JX074286.1 | ||
| Raiamas senegalensis | Benin | Iguidi | AP010780.1 | Saitoh et al., 2008 | |
| Raiamas senegalensis | HM224332.1 | ||||
| Labeo horie | Ethiopia | Alwero | JX074288.1 | ||
| Labeo nasus | DRC | Congo Basin | AP013333.1 | Miya 2013 | |
| Labeo lineatus | AP012154.1 | ||||
| Labeo altivelis | AP013322.1 | Miya 2011 | |||
| Labeo coubie | Nigeria | Niger Basin | AP012149.1 | ||
| Labeo coubie | JX074261.1 | ||||
| Labeo altivelis | JX074228.1 | ||||
| Raiamas steindachneri | N/A | AP012113.1 | |||
| Raiamas cf. steindachneri | IHB29666 | Sierra Leone | Rokel/Seli/upper | MW660585 | This study |
| Raiamas aff. scarciensis | IHB29555 | Sierra Leone | Rokel/lower | MW660586 | This study |
| Labeo aff. coubie | IHB29688 | Sierra Leone | Rokel/Seli/upper | MW660599 | This study |
| Labeo aff. parvus | IHB29699 | Sierra Leone | Rokel/Seli/upper | MW660600 | This study |
| Labeo aff. parvus | IHB29799 | Sierra Leone | Rokel/Seli/upper | MW660601 | This study |
| Labeobarbus wurtzi | IHB29999 | Sierra Leone | Rokel/Seli/lower | MW660597 | This study |
| Labeobarbus wurtzi | IHB29999B | Sierra Leone | Rokel/Seli/lower | MW660597B | This study |
| Labeobarbus sacratus | IHB29898 | Sierra Leone | Rokel/Seli/upper | MW660598 | This study |
| Enteromius aff. foutensis | IHB29317 | Sierra Leone | Rokel/Seli/upper | MW660590 | This study |
| Enteromius aff. foutensis | IHB29318 | Sierra Leone | Rokel/Seli/upper | MW660591 | This study |
| Enteromius aff. foutensis | IHB29319 | Sierra Leone | Rokel/Seli/upper | MW660592 | This study |
| Enteromius aff. foutensis | IHB29320 | Sierra Leone | Rokel/Seli/upper | MW660593 | This study |
| Enteromius aff. foutensis | IHB29444 | Sierra Leone | Rokel/Seli/upper | MW660594 | This study |
| Enteromius aff. liberiensis | IHB29241 | Sierra Leone | Rokel/Seli/upper | MW660587 | This study |
| Enteromius aff. liberiensis | IHB29362 | Sierra Leone | Rokel/Seli/upper | MW660589 | This study |
| Enteromius aff. liberiensis | IHB29242 | Sierra Leone | Rokel/Seli/upper | MW660588 | This study |
| Enteromius aff. macrops | IHB29355 | Sierra Leone | Rokel/Seli/upper | MW660595 | This study |
| Enteromius aff. macrops | IHB29610 | Sierra Leone | Rokel/Seli/upper | MW660596 | This study |
| Enteromius aff. ablabes | IHB29544 | Sierra Leone | Rokel/Seli/upper | MZ013921 | This study |
| Enteromius aff. ablabes | IHB29545 | Sierra Leone | Rokel/Seli/upper | MZ013922 | This study |
| Prolabeo batesi | IHB29377 | Sierra Leone | Rokel/lower | MZ013919 | This study |
| Prolabeo batesi | IHB29399 | Sierra Leone | Rokel/lower | MZ013920 | This study |
| Paracanthocobitis zonalternans | Asia | MK608087.1 | Slechtova and Dvorak 2019 | ||
| Paracanthocobitis mockenziei | MK608121.1 |
| No. | Species | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Labeobarbus wurtzi | ||||||||||||
| 2 | L. sacratus | 0.054 | |||||||||||
| 3 | Labeo forskallii | 0.182 | 0.157 | ||||||||||
| 4 | L aff. parvus | 0.180 | 0.159 | 0.080 | |||||||||
| 5 | L. parvus | 0.177 | 0.157 | 0.072 | 0.071 | ||||||||
| 6 | L. cyclorhynchus | 0.146 | 0.147 | 0.135 | 0.135 | 0.113 | |||||||
| 7 | L. longipinnis | 0.147 | 0.154 | 0.142 | 0.141 | 0.137 | 0.098 | ||||||
| 8 | L. aff. coubie | 0.161 | 0.168 | 0.135 | 0.143 | 0.130 | 0.097 | 0.068 | |||||
| 9 | L. coubie | 0.147 | 0.148 | 0.121 | 0.131 | 0.118 | 0.082 | 0.065 | 0.057 | ||||
| 10 | R. aff. steindachneri | 0.283 | 0.268 | 0.233 | 0.245 | 0.238 | 0.246 | 0.260 | 0.257 | 0.251 | |||
| 11 | R. steindachneri | 0.275 | 0.269 | 0.236 | 0.245 | 0.241 | 0.253 | 0.262 | 0.259 | 0.258 | 0.037 | ||
| 12 | R. scarciensis | 0.305 | 0.289 | 0.274 | 0.263 | 0.276 | 0.294 | 0.283 | 0.297 | 0.290 | 0.218 | 0.233 | |
| 13 | R. senegalensis | 0.292 | 0.284 | 0.268 | 0.259 | 0.263 | 0.249 | 0.281 | 0.260 | 0.260 | 0.216 | 0.229 | 0.204 |
| No. | Species | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Prolabeo batesi | ||||||||||||||||||||||
| 2 | Enteromius trispilos | 0.155 | |||||||||||||||||||||
| 3 | Enteromius cadenati | 0.161 | 0.157 | ||||||||||||||||||||
| 4 | Enteromius aff. cadenati | 0.183 | 0.147 | 0.165 | |||||||||||||||||||
| 5 | Enteromius aff. liberiensis RR | 0.178 | 0.154 | 0.182 | 0.046 | ||||||||||||||||||
| 6 | Enteromius liberiensis | 0.149 | 0.118 | 0.159 | 0.129 | 0.133 | |||||||||||||||||
| 7 | Enteromius tiekoroi | 0.161 | 0.132 | 0.171 | 0.145 | 0.160 | 0.090 | ||||||||||||||||
| 8 | Enteromius macrops s.str. | 0.156 | 0.147 | 0.166 | 0.173 | 0.179 | 0.130 | 0.134 | |||||||||||||||
| 9 | Enteromius aff. macrops1 | 0.150 | 0.137 | 0.162 | 0.158 | 0.166 | 0.119 | 0.132 | 0.035 | ||||||||||||||
| 10 | Enteromius aff. macrops2 | 0.148 | 0.143 | 0.159 | 0.157 | 0.162 | 0.120 | 0.138 | 0.027 | 0.027 | |||||||||||||
| 11 | Enteromius aff. macrops RR | 0.148 | 0.145 | 0.169 | 0.169 | 0.175 | 0.121 | 0.134 | 0.027 | 0.029 | 0.024 | ||||||||||||
| 12 | Enteromius anema | 0.150 | 0.134 | 0.170 | 0.145 | 0.163 | 0.131 | 0.139 | 0.132 | 0.128 | 0.136 | 0.139 | |||||||||||
| 13 | Enteromius guildi | 0.155 | 0.138 | 0.171 | 0.150 | 0.169 | 0.135 | 0.139 | 0.135 | 0.129 | 0.142 | 0.142 | 0.006 | ||||||||||
| 14 | Enteromius aff. foutensis RR | 0.153 | 0.148 | 0.169 | 0.139 | 0.156 | 0.132 | 0.123 | 0.126 | 0.124 | 0.123 | 0.123 | 0.101 | 0.104 | |||||||||
| 15 | Enteromius foutensis s.str. | 0.151 | 0.144 | 0.167 | 0.137 | 0.141 | 0.128 | 0.134 | 0.121 | 0.118 | 0.117 | 0.122 | 0.105 | 0.109 | 0.039 | ||||||||
| 16 | Enteromius aff. foutensis1 | 0.162 | 0.148 | 0.167 | 0.143 | 0.154 | 0.133 | 0.126 | 0.122 | 0.115 | 0.117 | 0.124 | 0.087 | 0.089 | 0.068 | 0.068 | |||||||
| 17 | Enteromius aff. foutensis2 | 0.155 | 0.149 | 0.166 | 0.157 | 0.164 | 0.137 | 0.125 | 0.120 | 0.118 | 0.117 | 0.121 | 0.087 | 0.089 | 0.083 | 0.080 | 0.051 | ||||||
| 18 | Enteromius ablabes | 0.160 | 0.151 | 0.163 | 0.161 | 0.165 | 0.137 | 0.125 | 0.121 | 0.120 | 0.120 | 0.124 | 0.094 | 0.097 | 0.087 | 0.080 | 0.052 | 0.009 | |||||
| 19 | Enteromius punctitaeniatus | 0.162 | 0.153 | 0.165 | 0.162 | 0.165 | 0.137 | 0.126 | 0.122 | 0.121 | 0.122 | 0.124 | 0.096 | 0.098 | 0.087 | 0.081 | 0.054 | 0.011 | 0.001 | ||||
| 20 | Enteromius aff. ablabes1 | 0.144 | 0.137 | 0.134 | 0.164 | 0.161 | 0.134 | 0.145 | 0.129 | 0.128 | 0.123 | 0.129 | 0.137 | 0.140 | 0.118 | 0.125 | 0.118 | 0.112 | 0.112 | 0.113 | |||
| 21 | Enteromius profundus | 0.147 | 0.139 | 0.162 | 0.170 | 0.190 | 0.131 | 0.134 | 0.135 | 0.131 | 0.129 | 0.133 | 0.127 | 0.127 | 0.128 | 0.128 | 0.130 | 0.118 | 0.125 | 0.127 | 0.138 | ||
| 22 | Enteromius aff. anema | 0.147 | 0.149 | 0.167 | 0.151 | 0.168 | 0.128 | 0.132 | 0.139 | 0.124 | 0.135 | 0.135 | 0.133 | 0.134 | 0.116 | 0.128 | 0.120 | 0.108 | 0.113 | 0.115 | 0.126 | 0.116 | |
| 23 | Enteromius aff. ablabes RR | 0.142 | 0.137 | 0.156 | 0.133 | 0.139 | 0.106 | 0.120 | 0.135 | 0.130 | 0.137 | 0.139 | 0.118 | 0.120 | 0.118 | 0.113 | 0.116 | 0.106 | 0.104 | 0.105 | 0.119 | 0.101 | 0.098 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kanu, U.C.; Liang, C.; Nwafor, C.C.; Shen, J.; Zhang, E. Phylogenetic Analyses of Cyprinid Species from the Rokel River Basin of Sierra Leone, West Africa: Taxonomic, Biogeographic, and Conservation Implications. Diversity 2022, 14, 299. https://0-doi-org.brum.beds.ac.uk/10.3390/d14040299
Kanu UC, Liang C, Nwafor CC, Shen J, Zhang E. Phylogenetic Analyses of Cyprinid Species from the Rokel River Basin of Sierra Leone, West Africa: Taxonomic, Biogeographic, and Conservation Implications. Diversity. 2022; 14(4):299. https://0-doi-org.brum.beds.ac.uk/10.3390/d14040299
Chicago/Turabian StyleKanu, Unisa Conteh, Cao Liang, Chinedu Charles Nwafor, Jianzhong Shen, and E Zhang. 2022. "Phylogenetic Analyses of Cyprinid Species from the Rokel River Basin of Sierra Leone, West Africa: Taxonomic, Biogeographic, and Conservation Implications" Diversity 14, no. 4: 299. https://0-doi-org.brum.beds.ac.uk/10.3390/d14040299