
Similar Response of a Range Expanding Dragonfly to Low- and High-Elevation Predators


1
Laboratoire Biologie, Eau et Environment (LBEE), Université 8 Mai 1945, Guelma 24000, Algeria
2
Department of Evolutionary Biology and Environmental Studies, University of Zurich, Winterthurerstrasse 190, CH-8057 Zurich, Switzerland
3
Institute for Resources, Environment and Sustainability, University of British Columbia, Vancouver, 429-2202 Main Mall, Vancouver, BC V6T 1Z4, Canada
4
Department of Biological Sciences, Simon Fraser University, Burnaby, BC V5A 1S6, Canada
*
Author to whom correspondence should be addressed.
Diversity 2022, 14(4), 302; https://0-doi-org.brum.beds.ac.uk/10.3390/d14040302
Submission received: 16 February 2022
/
Revised: 11 April 2022
/
Accepted: 12 April 2022
/
Published: 15 April 2022
(This article belongs to the Special Issue Diversity, Ecology and Evolution of Odonata)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Recent range expansion of many species northward and upward in elevation suggests that the expanding species are able to cope with new biotic interactions in the leading edge. To test this hypothesis, we used a common garden experiment expanding the elevation range of an obligatorily univoltine dragonfly (Sympetrum striolatum) to investigate whether the growth, behavioral (food intake), and morphological (8th and 9th abdominal lateral spine) responses differed when confronted with dragonfly predators that dominate low-elevation (Aeshna cyanea) and high-elevation (A. juncea) lentic freshwater systems under two temperature treatments (20 °C and 24 °C). Growth rate and growth efficiency increased at higher temperature. Overall, low- and high-elevation predators induced a similar increase in growth rate and growth efficiency but a decrease in food intake at 24 °C. Lateral abdominal spines were longer only in low-elevation dragonflies at 18 °C. Our study suggests that range-expanding species may have been successful in colonizing new areas at higher elevations because they respond to dominant high-elevation predators in a similar way to the more familiar low-elevation predators.
1. Introduction
To comprehend and predict future climate impacts on freshwater communities, there is growing interest in the biotic response of species to new colonizer species and its consequences on life history traits [1,2,3,4,5]. The ability to recognize and respond to different types of predators is a key trait that contributes to the successful establishment of new populations [6,7]. The response of growth rate to predatory stress has been widely investigated because of its intimate relationship with fitness [8]. Because the magnitude of biotic interactions (e.g., predation) can change with temperature, recent studies have focused on the thermal dynamics of predator-prey interaction to understand the outcome of range shift events that occur across geographic gradients such as latitude and elevation [9,10,11]. Both consumptive (visual cues) and non-consumptive (chemical cues) effects of predation can affect growth rate and induce changes in fitness-related traits [3,12,13]; however, studies on non-consumptive effects of cold-adapted (unfamiliar) predators on warm-adapted prey that gradually expand its geographic range to cooler areas are still lacking [4].
There are different physiological and behavioral mechanisms that determine growth rate, and understanding these mechanisms unravels some underlying plastic responses [5,14]. Usually, growth rate is a function of the amount of food ingested (food intake) and the amount of energy allocated to growth (growth efficiency) [15,16]. Thus, although a constant growth rate across an environmental gradient may be interpreted as an absence of the adaptive response, it may hide dynamic plasticity in the behavioral and physiological components that contribute to the growth rate [17,18]. On one hand, an increased food intake likely increases individual activity and foraging behavior for prey acquisition, which often results in a higher mortality rate due to increased detectability by predators [7]. On the other hand, an accelerated growth efficiency leads to faster energy allocation and results in lower energy reserves and immune function [5,19]. Thus, it is important to understand whether behavioral and physiological components of the growth rate change in response to different types of predators and different temperatures to reveal the selective forces acting against the evolution of growth rate and predict the population dynamics of plasticity in new habitats after range expansion.
Apart from physiology and behavior, predators may induce plastic morphological responses such as defensive weaponry that reduces successful predation attempts [20,21]. As with growth efficiency, investment in defensive weaponry has survival costs and leads to a lower allocation of energy to other functional traits [22]. It has been shown that there is a negative relationship between behavioral and morphological defenses, so-called trait compensation [21,23]. Spending energy on defensive weaponry results in reduced vigilance while foraging. It is still not clear how species with time constraints respond to various types of predators, and which morphological, physiological, and/or behavioral mechanisms are prioritized.
While recent experimental studies have investigated the latitudinal patterns in predator–prey interactions [4,5], similar studies on the elevational gradient are still lacking. In the current study, we investigate the elevational range shift in Switzerland of Sympetrum striolatum, a widespread obligatorily univoltine species in North Africa, Europe, and Asia. To understand whether the species is able to cope with new biotic interactions at higher elevations, we assessed in a common garden experiment the response of growth rate, growth efficiency, food intake, and morphology (larval lateral spines) to temperature and different predation treatments. We exposed the species to two predators, a familiar (Aeshna cyanea: mainly at low elevations) dragonfly predator and an unfamiliar (A. juncea: mainly at high elevations) dragonfly predator. The low-high elevation dragonfly comparison reveals how the species would respond in a scenario of range shift to new habitats with a new predator, a phenomenon that has occurred repeatedly during the last decades [24,25]. Documenting the mechanisms controlling variation in growth rate across different treatments of predation and temperature is essential to understand the evolution of plasticity and predict future population dynamics in freshwater ecosystems [17].
2. Materials and Methods
2.1. Study Species and Distribution Data
Sympetrum striolatum is an obligatorily univoltine dragonfly that inhabits various types of wetlands, mostly stagnant freshwater, where it lays eggs from mid-summer to autumn and occupies an intermediate trophic level (as both predator and prey) in the freshwater food web. The species has shown a northward range expansion in the UK in recent decades [25], which makes it appropriate for the research question. In Switzerland, the species is historically considered a low-elevation species [26]. Long-term observations of adult S. striolatum carried out in Switzerland between 1990 and 2013 were obtained from CSCF (Centre Suisse de Cartographie de la Faune. The data included not only adult observations but also sex and reproductive states. To assess range shift across elevation, we included only records that showed a reproductive state such as copulation, oviposition, and emergence. The 95th percentile of the elevational distribution (leading edge) of S. striolatum and A. cyanea, and the 5th percentile (trailing edge) of the elevational distribution of A. juncea were calculated for each year to assess their historical changes during 1990–2013.
2.2. Study Site and Treatments
To investigate the effect of predation risk on the behaviour and physiology of dragonflies as well as the underlying proximate mechanisms, the response of larvae of Sympetrum striolatum to two different predators was assessed in a common garden experiment in the laboratory at the University of Zurich, Switzerland. Eggs of S. striolatum were collected from two sites in Zurich, Switzerland (Irchel pond: 47.3977° N, 8.5448° E, 480 m elevation; Chatzensee pond: 47.4264° N, 8.4878°, 440 m elevation). Both sites occurred within or near urban areas. We obtained the eggs from copulating females that were captured with a hand net in the middle of the day in early October 2015. The female abdomen was immersed in a vial with water, which led to the oviposition (about a few hundred eggs per female) [27]. Five different females from each site were sampled. Eggs were brought to the laboratory within two hours and put at room temperature (21 °C) and natural light conditions. Eggs of S. striolatum had a direct development and hatched within 2–3 weeks. After hatching, 20 larvae were placed in each of the six treatments (2 temperatures [18 °C, 24 °C] × 3 predation treatments [control, low-elevation, and high-elevation]) and raised individually in 180-mL plastic cups filled with dechlorinated tap water and floating on water tubs. Using polystyrene foam, the cups were able to remain floating on the surface of the water.
A randomized full factorial design using eggs from each of the two populations which were exposed to the combination of two predation treatments (low-elevation [A. cyanea] and high-elevation [A. juncea]) and two constant temperatures (20 and 24 °C). Temperatures were maintained by setting heaters at the bottom of water tubs. The low-elevation dragonfly was Aeshna cyanea and the high-elevation dragonfly was A. juncea, whose larvae were collected from Irchel forest (47.3893° N, 8.5611° E, 660 m) and Flumserberg (47.0661° N, 9.2546° E, 1904 m), respectively. Although the two species’ elevational distribution overlap, the largest populations of each do not coexist at a similar elevation [26]. The two temperatures used in the experiment are far below the thermal critical maximum of S. striolatum [28].
Larvae under predation treatments received only chemical (no visual) cues from predators. The predator medium containing chemical cues was acquired by individually setting the predators (four large dragonfly larvae [size > 4 cm] in containers filled with dechlorinated tap water [12 × 12 cm filled to the height of 4 cm]). All predators were fed every day with a single S. striolatum larva (body length: 0.5–1 cm). We mixed together the chemical cues of four individuals of the same predator species, then 1 mL of the solution was provided daily to the prey. All S. striolatum larvae were fed once a day with Artemia sp. nauplii (~80–110 individuals in 5 mL).
2.3. Growth Efficiency and Food Intake
During 60 days after hatching, growth rates were quantified by measuring the head width of larvae every four days, taking pictures with a stereoscope, and estimating the distance between the edges of the eyes to the nearest 0.01 mm using ImageJ v1.52. To estimate the growth efficiency, a 4-day experiment as described by Stoks, Swillen and De Block [5] was used on 60-day-old larvae. This experiment consists of weighing larvae from different treatments to the nearest 0.01 mg using an electronic balance at the start and the end of the 4-day period. Then growth rate was measured as [log(final mass)–log(initial mass)]/4 days, which accounts for differences in mass at the start of the experiment [18]. Given the widespread distribution of Daphnia in freshwater ecosystems worldwide, and its importance as prey to dragonflies [29], we provided S. striolatum larvae with 20 individual Daphnia of similar size each day. The number of Daphnia eaten per day was counted and replaced. The number of eaten daphnia was converted to total dry mass ingested using an equation [dry mass = a wet mass + b] established through weighing (to the nearest μg using an electronic balance) Daphnia before and after drought-treatment in the oven at 60 °C during 48 h. Dry mass of larvae was also estimated in the same way to obtain a conversion factor (dry mass = 0.114 × wet mass + 0.478).
2.4. Morphological Defense
Predation avoidance in freshwater prey might involve morphological modifications that reduce the probability of being eaten [1,20,21]. To test whether there is variability in the morphological response to different predator types, the 8th and 9th lateral abdominal spines of larvae were measured at the end of the experiments, 65 days after hatching. The size of lateral abdominal spines shows plasticity when exposed to predators [21]. The length of the spines was corrected for the larval size by dividing it by the head width of larvae; a widely used measure of body size which describes the developmental stage of larvae [30].
2.5. Statistical Analyses
All statistical analyses were carried out using R 3.5.1 [31]. We calculated the leading and trailing edge of the distribution of all three species using 95th and 5th percentile of the elevation of the occurrence data, respectively. Then, elevational range shift was assessed using simple linear regressions including our estimate of the elevational distribution edge (leading or trailing) as a response variable and years as an explanatory variable. To determine differences in growth rate among treatments, linear mixed-effects (LME) model including head width as a response variable and time (days), predation and temperature as explanatory variables, and individual identifier as a random effect, was carried out using lme4 package [32]. We used the lstrends function from lsmeans package [33] on our LME model to calculate for each treatment average growth rate with 95% confidence intervals. Since we had a single measure per individual (no repeated measures), we used a linear model to determine the difference in growth efficiency, food intake and lateral spine length between temperature and predation treatments.
3. Results
3.1. Elevational Range Shift
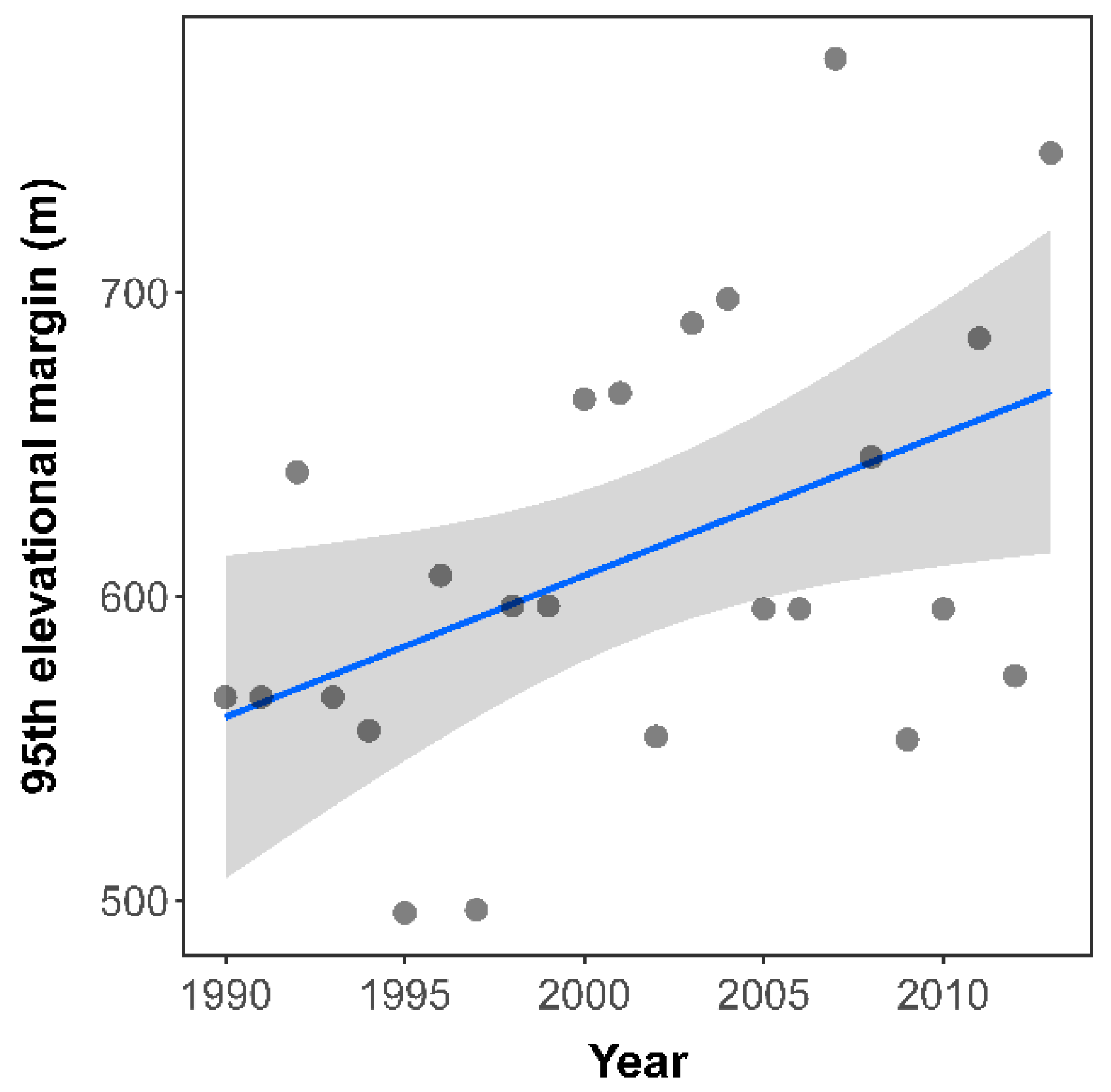
The 95th margin of the elevational distribution (leading edge) shifted by 4.6 m/year (Figure 1; linear model: slope = 4.66 m/yr, R2 = 0.21, p = 0.02). The trailing edge (5th percentile) of the high-elevation predator A. juncea did not show a significant shift (slope = −2.63 m/yr, R2 = 0.03, p = 0.40), nor the leading edge of the low-elevation predator A. cyanea (slope = −12.2 m/yr, R2 = 0.14, p = 0.06) during the same period.
3.2. Larval Growth
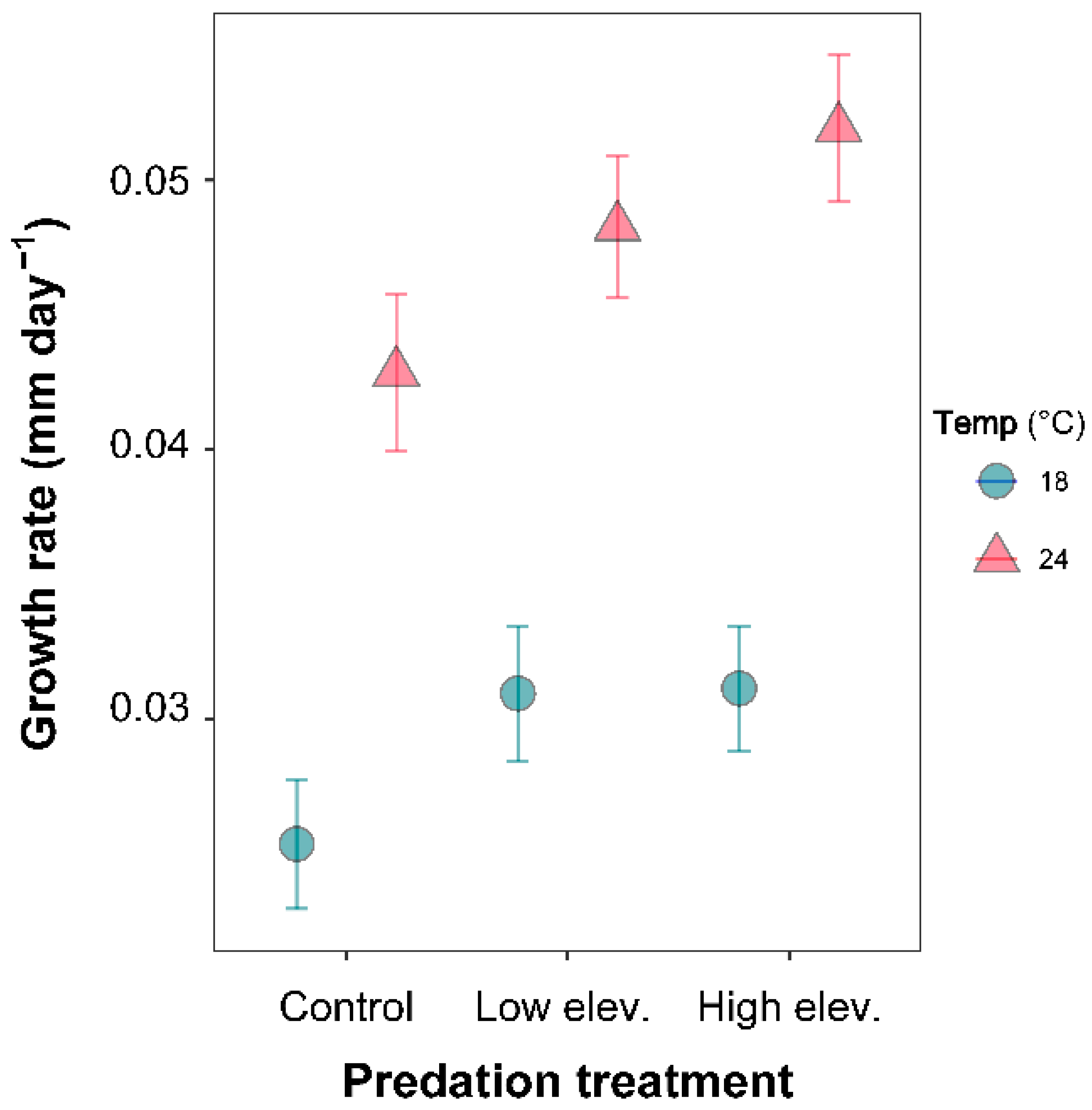
Larvae grew faster in water at 24 °C than at 18 °C (LME: p = 0.0001; Table S1). Growth rate was significantly different among predation treatments (LME: p = 0.002; Table S1), revealing a faster growth in predation treatment compared to the control treatment (Figure 2). The significant interaction between time and temperature and the non-significant three-way interaction between time, temperature, and predation indicate that larvae responded similarly to the different predators at both temperatures (p > 0.05; Table S1).
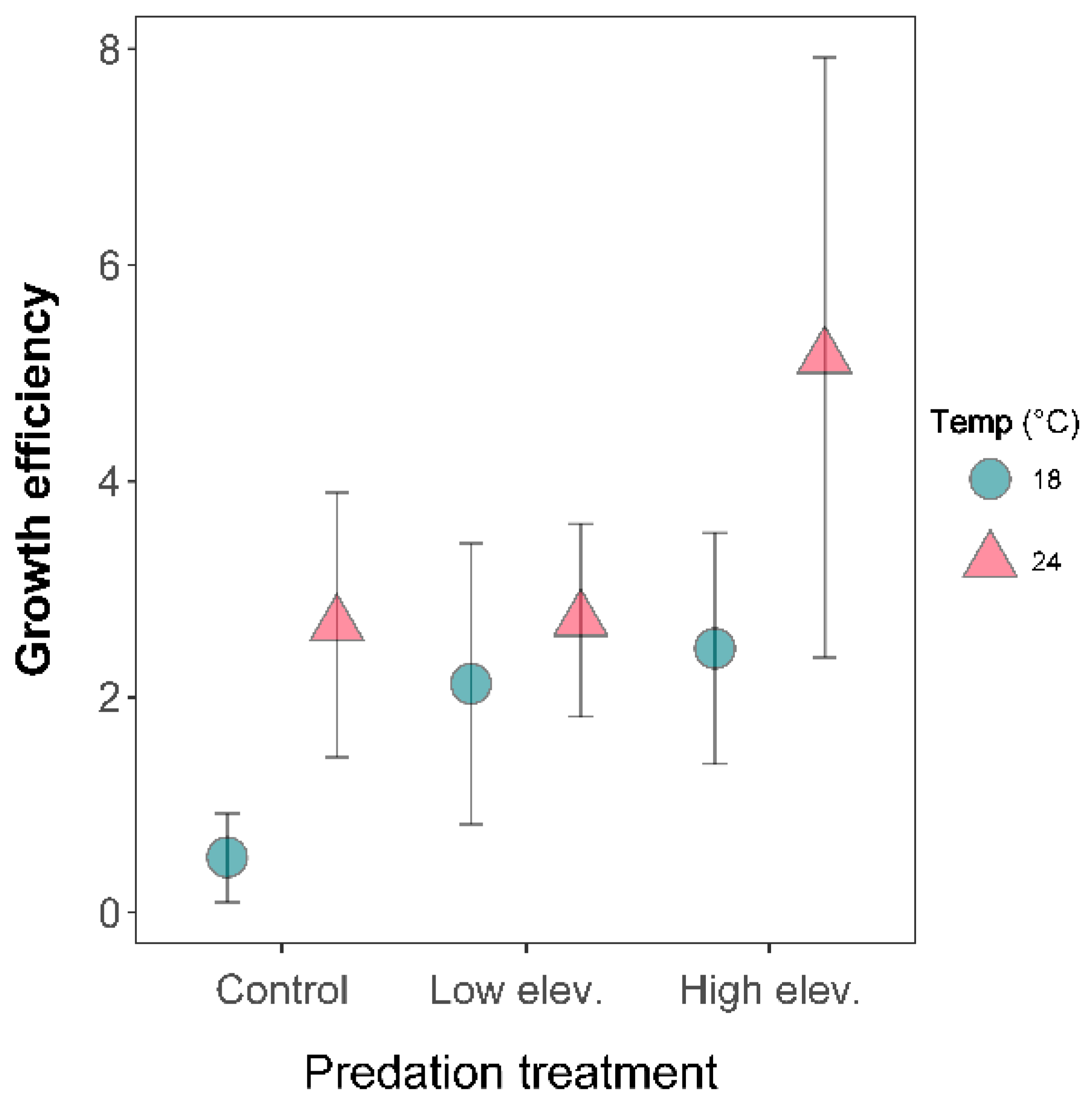
Growth efficiency was significantly different between temperature (LME: p = 0.0002) and predator treatments (LME: p < 0.002) (Figure 3). Growth efficiency was 81.6% greater at higher temperature than at lower temperature. Larvae in predation treatments showed a higher growth efficiency than in the control treatment, but there was no significant difference between low and high-elevation predator (Figure 3). There was a small significant interaction between temperature and predation treatment (Table S2).
3.3. Behavioral Response
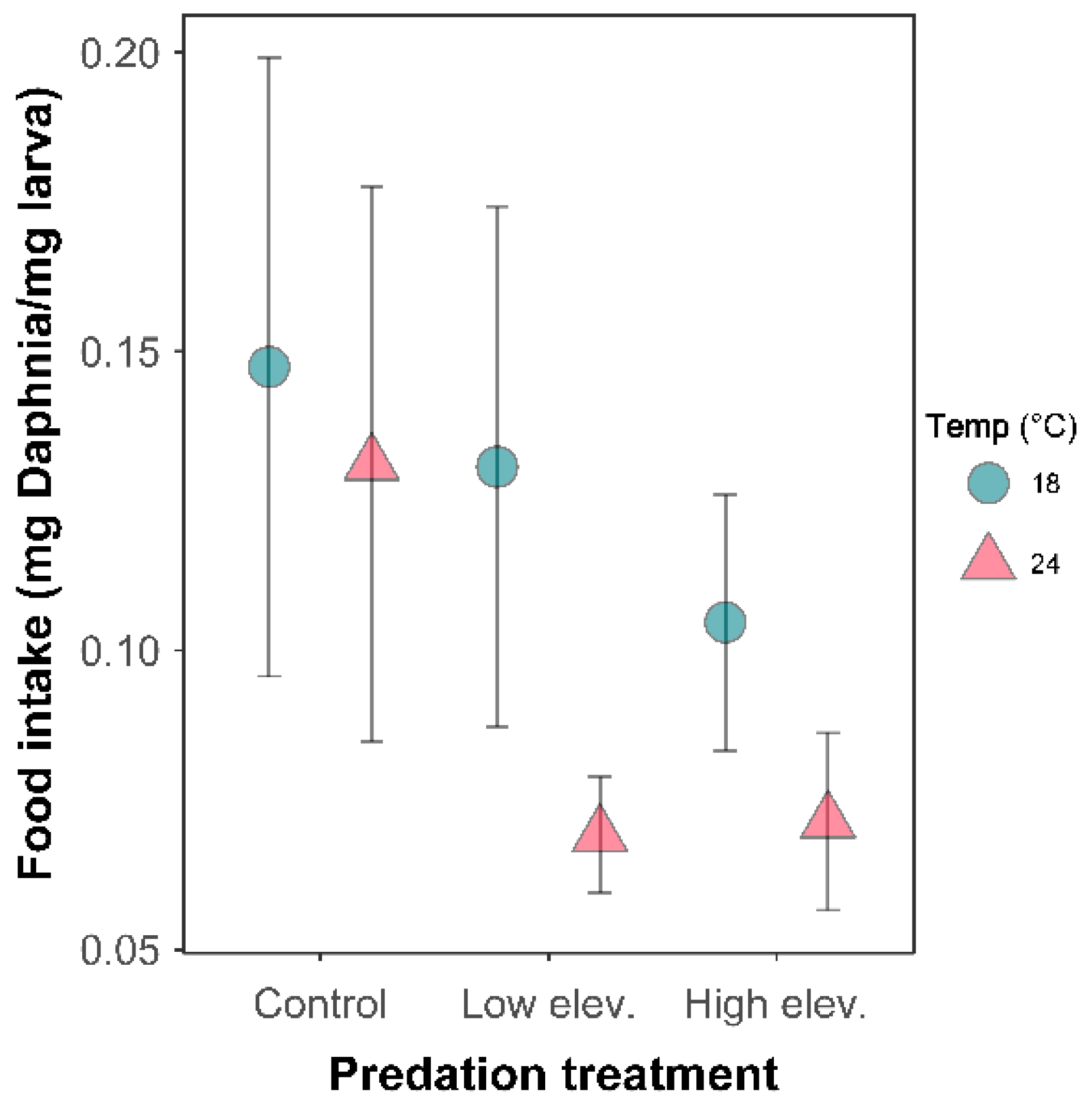
There was a significant interaction between temperature and predation (Figure 4, Table S3). At 18 °C, larvae showed a lower food intake in high-elevation predator treatment (p = 0.002). At 24 °C, larval food intake was lower in both low- and high-elevation predator treatment than in the control, although marginal for the high-elevation predator (Table S3, p = 0.01; p = 0.07, respectively).
3.4. Morphological Response
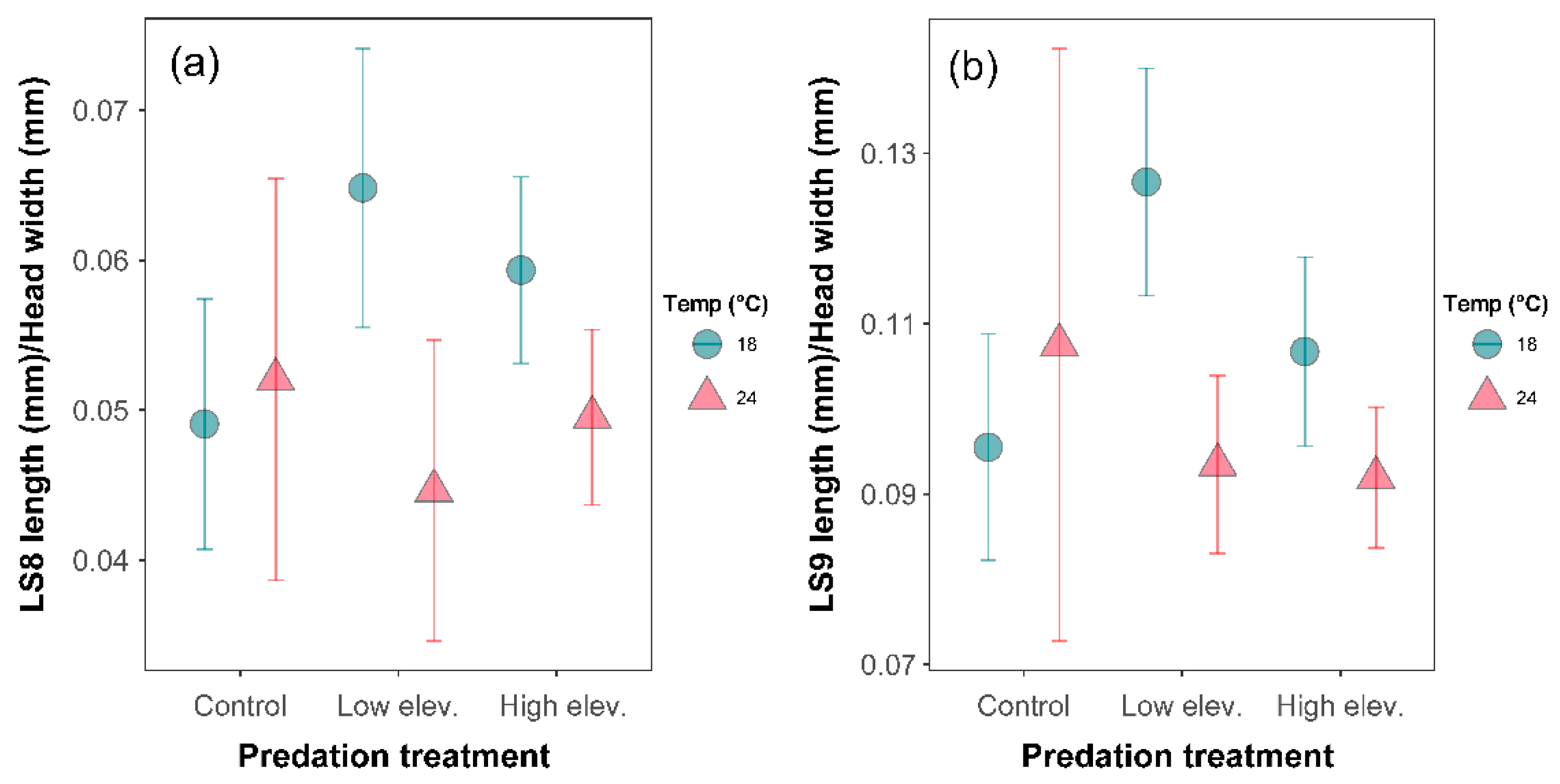
The length of the 8th lateral spine was longer in the low-elevation dragonfly treatment (p = 0.01; Table S4a, Figure 5a), but neither temperature nor the interaction of temperature with predation showed a significant effect (Table S4a). The length of the 8th lateral spine was 32% longer in the low-elevation predator treatment than in the control (Figure 5a, Table S4a). Similar to the 8th lateral spine, there was no significant effect of temperature on the length of the 9th lateral spine (Table S4b, p = 0.64). However, larvae in the low-elevation dragonfly treatment under 18 °C had significantly longer 9th spines than the other treatments (p = 0.001; Table S4b, Figure 5b). The average length of the 9th spine in the low-elevation predator treatment was 33% longer than that of the control.
4. Discussion
In this study, we showed that the low-elevation obligatorily univoltine S. striolatum expanded its range by 4.6 m/yr across elevation during 1990–2013, which suggests that the species was able to adapt to the new biotic environment. The common garden experiments assessing the non-consumptive response of growth (rate and efficiency), behavior (food intake), and morphology (abdominal spine) of S. striolatum to different predators including low- (familiar) and high-elevation (non-familiar) dragonflies showed that overall dragonfly larvae responded similarly in growth and food intake to both predators, but they responded distinctly in morphology to the familiar dragonfly predator.
4.1. Range Shift
Sympetrum striolatum has shown an elevational range shift of 4.6 m/year between 1990 and 2013. The same species has been reported to expand its northern range limit in the UK by 346 km between 1960 and 1995, which is equivalent to a rate of 8.6 km/year [25]. Given that this species is still flourishing at low elevations and in the southern range limit, it can be inferred that the range has expanded both in latitude and elevations. Discrepancies between latitudinal and elevational range shift are probably due to the steeper cline in environmental conditions across elevation, which restricts the speed at which new populations are established at higher elevations. However, the successful establishment of higher elevation populations is probably due to recent climate warming in the Swiss Alps, but it also suggests that the species was able to cope with new biotic conditions. Furthermore, the absence of elevational shift of the trailing distribution edge of A. juncea suggests that the encounter probability between S. striolatum and A. juncea has increased. We suggest that the overlap will increase even further in the following years.
4.2. Temperature Effects
Temperature had various effects on growth, food intake, and length of lateral spines of larvae. Higher temperatures led to an increased growth rate and growth efficiency. In many taxa, growth rate is expected to increase with temperature when temperatures are below the thermal optimum [34], which is typically higher than 24 °C in temperate Libellulidae [35]. However, unlike studies on odonates [5,36] and butterflies [37] which found that the increase in growth rate was associated with an increase in food intake, our study showed that it was associated with an increase in growth efficiency [16]. Thus, it is likely that this is an adaptive mechanism that allows obligatorily univoltine species such as S. striolatum to respond to an increase in temperature by increasing growth efficiency despite lowering food intake. Such a strong physiological response probably has considerable fitness trade-offs for adults [38]. In a climate warming scenario, temperature-induced changes in physiology and behavior might have a major impact on predator-prey interaction and community structure [4,39,40]. For example, an increase of 4 °C (difference in temperature treatments in this study) causes an increase in growth rate and a decrease in food intake which may lead to an earlier emergence (shorter exposure of prey to predators) and a decline in predation rates (less prey eaten during the short exposure time). These effects may have direct [41] and indirect consequences [42] on the freshwater food web. Interestingly, a positive response in anti-predatory morphological defenses was observed only at low temperatures where food intake was higher, suggesting that at higher temperature larvae invested in growth rather than anti-predatory weaponry.
4.3. Predation Effects
The analysis of growth rate showed a significant difference among predator treatments. Unlike the commonly reported pattern of growth reduction under predation treatments [17,43], we found a predatory-induced acceleration of growth which was also reported in odonates [5,44,45,46] and other vertebrates [47,48]. One explanation of the increased growth rate under predation is the ‘escape theory’ which is predicted by some optimal models when non-predatory costs of growth exist [49,50]. This theory is further supported by the fact that many Sympetrum species are obligatorily univoltine [51], and thus environmental limitations such as predation or time constraints result in a fastening of growth rates [5,52]. This finding highlights the fact that the recent range shift of many large species of odonates, invertebrates, and vertebrates might alter the life history of many prey taxa of freshwater ecosystems, which might result in major evolutionary and ecological changes.
There was a lower food intake under dragonfly predation (both low- and high-elevation) than in the control, which suggests that larvae detected the predator through chemical cues and subsequently lowered their food intake probably to avoid being detected and eaten [2]. A similar decline in food intake under dragonfly predation was detected in damselflies [5,17] and dragonflies [53]. Furthermore, there was a higher growth efficiency under dragonfly treatment than in the control treatment, which means that larvae grew fast with respect to the amount of food consumed. A similar increase in growth efficiency has been observed in damselflies in the presence of predation [5]. The lower food intake and higher growth efficiency for a species adapted to time constraints could be theoretically explained by optimality models [50]. Food intake involves at least the movement of mouthpieces (mentum), which makes prey larvae detectable by large dragonfly predators. Thus, decreasing food intake is a common anti-predatory behavioral mechanism that individuals adopt to increase their survival probability [54]. High growth efficiency is a physiological anti-predatory response that allows larvae to minimize their exposure to predation and reach the adult stage as early as possible [55]. This hypothesis is supported by previous studies on many taxa carried out in freshwater ecosystems on odonates [45,46], frogs [48], and fish [3,47]. The costs of growth acceleration have been documented and could involve a weakening of the immune system and reduction in energy reserves [15], and could even carry over until the adult stage where individuals encounter shorter longevity and reproductive success [56].
Morphological responses to predators have been well documented in odonates, but mostly against fish [20,21,57]. Interestingly, our study showed a morphological response of the lateral abdominal spines when exposed to chemical cues of dragonfly predators. Studies have shown that fish reject larval dragonflies which have long abdominal spines, thus allowing dragonflies to increase their survival probabilities [20]. It is unclear how longer spines benefit larvae from dragonfly predation. One likely hypothesis is that longer spines might be a by-product of physiological responses rather than an adaptive response to dragonfly predation [47]. In fact, higher growth efficiency might result in morphological changes that might include variation in the length or morphology of abdominal structures such as lateral spines. However, this hypothesis does not explain the observed longer lateral spines in low-elevation dragonfly treatment but not in high-elevation dragonfly treatment. This intriguing response to the more familiar predator warrants further investigations of the role of abdominal lateral spines against dragonfly predation.
While larvae used to estimate physiological and behavioral parameters had similar ages, there might be a difference in larval instars among individuals that might influence the response of larvae. Nevertheless, the observed physiological and behavioral responses were similar to other studies [5,45,48]. Since antipredator response can be costly in invertebrates in general [58], one would expect that selection should favor accuracy in detecting the predators that induce mortality or damage [59]. The fact that low- and high-elevation predators (A. cyanea and A. juncea) are closely related species with potentially similar diet could explain the similar behavioral and physiological response of S. striolatum larvae. Studies have linked the composition of chemical signals (kairomones) and predator diet [60,61,62]. The ability to detect a non-familiar predator is a dispersal asset for species, allowing larvae to develop, persist, and successfully establish a viable population in new habitats. For instance, many libellulids such as S. striolatum are invaders of ponds and lakes of northern and high-elevation areas [63], causing changes in species composition and probably competing with rare specialist species [35,64]. It could be that their success in invading new territories is due to their ability to respond similarly to new predators as their familiar predator. However, whether populations of S. striolatum at low elevations living in different types of landscapes (e.g., natural, suburban, or urban) respond similarly to different predators remains to be investigated. Our results suggest that species that successfully expanded their range to higher elevation habitats might be preadapted to the new biotic interactions. Future studies should investigate the implication of odonate range expansion on the dynamics of freshwater and terrestrial foodweb [65].
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/d14040302/s1, Table S1: Summary results of the linear mixed-effects model regressing head width of Sympetrum striolatum against time, temperature and predation treatments. Table S2: Summary results of the linear mixed-effects model regressing growth efficiency of Sympetrum striolatum against temperature and predation treatments. Table S3: Summary results of the linear mixed-effects model regressing food intake of Sympetrum striolatum against temperature and predation treatments. Table S4: Summary results of the linear mixed-effects model regressing the length of the 8th (a) and 9th (b) lateral spine corrected for body size of Sympetrum striolatum against temperature and predation treatments.
Author Contributions
Conceptualization, R.K.; methodology, R.K., R.Z. and H.M.; software, R.K., R.Z. and H.M.; validation, R.K.; formal analysis, R.Z.; investigation, R.K., R.Z. and H.M.; data curation, H.M.; writing—original draft preparation, R.K. and R.Z.; writing—review and editing, R.K., R.Z. and H.M.; visualization, H.M.; supervision, R.K.; funding acquisition, R.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Swiss National Science Foundation, grant number P2ZHP2_175028. The APC was funded by Swiss National Science Foundation.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We would like to thank the Centre suisse de cartographie de la faune and Christian Monnerat for providing the Swiss atlas data of odonates.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sih, A. Predators and prey life style. In Predation: Direct and Indirect Impacts on Aquatic Communities; Kerfoot, W.C., Sih, A., Eds.; University Press of New England: Hanover, UK, 1987. [Google Scholar]
- Werner, E.E.; Anholt, B.R. Ecological consequences of the trade-off between growth and mortality rates mediated by foraging activity. Am. Nat. 1993, 142, 242–272. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.; Dingemanse, N.; Hankison, S.; Langenhof, M.; Rollins, K. Early exposure to nonlethal predation risk by size-selective predators increases somatic growth and decreases size at adulthood in threespined sticklebacks. J. Evol. Biol. 2011, 24, 943–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Block, M.; Pauwels, K.; Van Den Broeck, M.; De Meester, L.; Stoks, R. Local genetic adaptation generates latitude-specific effects of warming on predator–prey interactions. Glob. Chang. Biol. 2013, 19, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Stoks, R.; Swillen, I.; De Block, M. Behaviour and physiology shape the growth accelerations associated with predation risk, high temperatures and southern latitudes in Ischnura damselfly larvae. J. Anim. Ecol. 2012, 81, 1034–1040. [Google Scholar] [CrossRef]
- Richardson, J.M. A comparative study of phenotypic traits related to resource utilization in anuran communities. Evol. Ecol. 2002, 16, 101–122. [Google Scholar] [CrossRef]
- Stoks, R.; McPeek, M.; Mitchell, J. Evolution of prey behavior in response to changes in predation regime: Damselflies in fish and dragonfly lakes. Evolution 2003, 57, 574–585. [Google Scholar] [CrossRef]
- Conover, D.O.; Schultz, E.T. Phenotypic similarity and the evolutionary significance of countergradient variation. Trends Ecol. Evol. 1995, 10, 248–252. [Google Scholar] [CrossRef]
- Wellborn, G.A.; Skelly, D.K.; Werner, E.E. Mechanisms creating community structure across a freshwater habitat gradient. Annu. Rev. Ecol. Evol. Syst. 1996, 27, 337–363. [Google Scholar] [CrossRef] [Green Version]
- Parmesan, C.; Ryrholm, N.; Stefanescu, C.; Hill, J.K.; Thomas, C.D.; Descimon, H.; Huntley, B.; Kaila, L.; Kullberg, J.; Tammaru, T. Poleward shifts in geographical ranges of butterfly species associated with regional warming. Nature 1999, 399, 579. [Google Scholar] [CrossRef]
- Hassall, C.; Thompson, D.J. Accounting for recorder effort in the detection of range shifts from historical data. Methods Ecol. Evol. 2010, 1, 343–350. [Google Scholar] [CrossRef]
- Lima, S.L.; Dill, L.M. Behavioral decisions made under the risk of predation: A review and prospectus. Can. J. Zool. 1990, 68, 619–640. [Google Scholar] [CrossRef]
- Antoł, A.; Sniegula, S. Damselfly eggs alter their development rate in the presence of an invasive alien cue but not a native predator cue. Ecol. Evol. 2021, 11, 9361–9369. [Google Scholar] [CrossRef] [PubMed]
- Blanckenhorn, W.; Demont, M. Bergmann and converse Bergmann latitudinal clines in arthropods: Two ends of a continuum? Integr. Comp. Biol. 2004, 44, 413–424. [Google Scholar] [CrossRef] [Green Version]
- Stoks, R.; Block, M.D.; McPeek, M.A. Physiological costs of compensatory growth in a damselfly. Ecology 2006, 87, 1566–1574. [Google Scholar] [CrossRef]
- Stoks, R.; Block, M.D.; Van De Meutter, F.; Johansson, F. Predation cost of rapid growth: Behavioural coupling and physiological decoupling. J. Anim. Ecol. 2005, 74, 708–715. [Google Scholar] [CrossRef]
- McPeek, M.A. The growth/predation risk trade-off: So what is the mechanism? Am. Nat. 2004, 163, E88–E111. [Google Scholar] [CrossRef]
- McPeek, M.A.; Grace, M.; Richardson, J.M. Physiological and behavioral responses to predators shape the growth/predation risk trade-off in damselflies. Ecology 2001, 82, 1535–1545. [Google Scholar] [CrossRef]
- Thaler, J.S.; McArt, S.H.; Kaplan, I. Compensatory mechanisms for ameliorating the fundamental trade-off between predator avoidance and foraging. Proc. Natl. Acad. Sci. USA 2012, 109, 12075–12080. [Google Scholar] [CrossRef] [Green Version]
- Mikolajewski, D.J.; Rolff, J. Benefits of morphological defence demonstrated by direct manipulation in larval dragonflies. Evol. Ecol. Res. 2004, 6, 619–626. [Google Scholar]
- Mikolajewski, D.J.; Johansson, F. Morphological and behavioral defenses in dragonfly larvae: Trait compensation and cospecialization. Behav. Ecol. 2004, 15, 614–620. [Google Scholar] [CrossRef]
- Kishida, O.; Trussell, G.C.; Mougi, A.; Nishimura, K. Evolutionary ecology of inducible morphological plasticity in predator–prey interaction: Toward the practical links with population ecology. Popul. Ecol. 2010, 52, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Dmitriew, C.; Rowe, L. Resource limitation, predation risk and compensatory growth in a damselfly. Oecologia 2005, 142, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Angert, A.L.; Crozier, L.G.; Rissler, L.J.; Gilman, S.E.; Tewksbury, J.J.; Chunco, A.J. Do species’ traits predict recent shifts at expanding range edges? Ecol. Lett. 2011, 14, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Hickling, R.; Roy, D.B.; Hill, J.K.; Thomas, C.D. A northward shift of range margins in British Odonata. Glob. Chang. Biol. 2005, 11, 502–506. [Google Scholar] [CrossRef]
- Wildermuth, H.; Gonseth, Y.; Maibach, A. Die Libellen der Schweiz. Fauna Helv. 2005, 12, 398. [Google Scholar]
- Khelifa, R.; Zebsa, R.; Kahalerras, A.; Mahdjoub, H. Clutch size and egg production in Orthetrum nitidinerve Selys, 1841 (Anisoptera: Libellulidae): Effect of body size and age. Int. J. Odonatol. 2012, 15, 51–58. [Google Scholar] [CrossRef]
- Khelifa, R. Spatiotemporal Pattern of Phenology across Geographic Gradients in Insects. Ph.D. Thesis, University of Zurich, Zurich, Switzerland, 2017. [Google Scholar]
- Thompson, D.J. Towards a realistic predator-prey model: The effect of temperature on the functional response and life history of larvae of the damselfly, Ischnura elegans. J. Anim. Ecol. 1978, 47, 757–767. [Google Scholar] [CrossRef]
- Corbet, P.S. Dragonflies: Behaviour and Ecology of Odonata; Harley Books: Colchester, UK, 1999. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Lenth, R. Least-Squares Means: The R package (lsmeans). J Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Angilletta, M.J. Thermal Adaptation: A Theoretical and Empirical Synthesis; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Suhling, F.; Suhling, I.; Richter, O. Temperature response of growth of larval dragonflies—An overview. Int. J. Odonatol. 2015, 18, 15–30. [Google Scholar] [CrossRef]
- Van Doorslaer, W.; Stoks, R. Growth rate plasticity to temperature in two damselfly species differing in latitude: Contributions of behaviour and physiology. Oikos 2005, 111, 599–605. [Google Scholar] [CrossRef]
- Karl, I.; Fischer, K. Why get big in the cold? Towards a solution to a life-history puzzle. Oecologia 2008, 155, 215–225. [Google Scholar] [CrossRef]
- De Block, M.; Stoks, R. Fitness effects from egg to reproduction: Bridging the life history transition. Ecology 2005, 86, 185–197. [Google Scholar] [CrossRef]
- Hunt, R.J.; Swift, M. Predation by larval damselflies on cladocerans. J. Freshw. Ecol. 2010, 25, 345–351. [Google Scholar] [CrossRef]
- Raczyński, M.; Stoks, R.; Johansson, F.; Sniegula, S. Size-mediated priority effects are trait-dependent and consistent across latitudes in a damselfly. Oikos 2021, 130, 1535–1547. [Google Scholar] [CrossRef]
- Traill, L.W.; Lim, M.L.; Sodhi, N.S.; Bradshaw, C.J. Mechanisms driving change: Altered species interactions and ecosystem function through global warming. J. Anim. Ecol. 2010, 79, 937–947. [Google Scholar] [CrossRef]
- Yang, L.H.; Rudolf, V. Phenology, ontogeny and the effects of climate change on the timing of species interactions. Ecol. Lett. 2010, 13, 1–10. [Google Scholar] [CrossRef]
- Altwegg, R. Predator-induced life-history plasticity under time constraints in pool frogs. Ecology 2002, 83, 2542–2551. [Google Scholar] [CrossRef]
- Slos, S.; Stoks, R. Behavioural correlations may cause partial support for the risk allocation hypothesis in damselfly larvae. Ethology 2006, 112, 143–151. [Google Scholar] [CrossRef]
- Wohlfahrt, B.; Mikolajewski, D.J.; Joop, G.; Vamosi, S.M. Ontogenetic changes in the association between antipredator responses and growth variables. Ecol. Entomol. 2007, 32, 567–574. [Google Scholar] [CrossRef]
- Mikolajewski, D.J.; Brodin, T.; Johansson, F.; Joop, G. Phenotypic plasticity in gender specific life-history: Effects of food availability and predation. Oikos 2005, 110, 91–100. [Google Scholar] [CrossRef]
- Johansson, F.; Andersson, J. Scared fish get lazy, and lazy fish get fat. J. Anim. Ecol. 2009, 78, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Benard, M.F. Predator-induced phenotypic plasticity in organisms with complex life histories. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 651–673. [Google Scholar] [CrossRef]
- Abrams, P.A.; Rowe, L. The effects of predation on the age and size of maturity of prey. Evolution 1996, 50, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- Higginson, A.; Ruxton, G. Adaptive changes in size and age at metamorphosis can qualitatively vary with predator type and available defenses. Ecology 2010, 91, 2756–2768. [Google Scholar] [CrossRef] [PubMed]
- Corbet, P.S.; Suhling, F.; Soendgerath, D. Voltinism of Odonata: A review. Int. J. Odonatol. 2006, 9, 1–44. [Google Scholar] [CrossRef]
- Johansson, F.; Stoks, R.; Rowe, L.; De Block, M. Life history plasticity in a damselfly: Effects of combined time and biotic constraints. Ecology 2001, 82, 1857–1869. [Google Scholar] [CrossRef]
- Pierce, C. Predator avoidance, microhabitat shift, and risk-sensitive foraging in larval dragonflies. Oecologia 1988, 77, 81–90. [Google Scholar] [CrossRef]
- Sih, A.; Bell, A.; Johnson, J.C. Behavioral syndromes: An ecological and evolutionary overview. Trends Ecol. Evol. 2004, 19, 372–378. [Google Scholar] [CrossRef] [Green Version]
- Dmitriew, C.M. The evolution of growth trajectories: What limits growth rate? Biol. Rev. 2011, 86, 97–116. [Google Scholar] [CrossRef]
- Metcalfe, N.B.; Monaghan, P. Growth versus lifespan: Perspectives from evolutionary ecology. Exp. Gerontol. 2003, 38, 935–940. [Google Scholar] [CrossRef]
- Johansson, F. Reaction norms and production costs of predator-induced morphological defences in a larval dragonfly (Leucorrhinia dubia: Odonata). Can. J. Zool. 2002, 80, 944–950. [Google Scholar] [CrossRef]
- Van Buskirk, J. Phenotypic lability and the evolution of predator-induced plasticity in tadpoles. Evolution 2002, 56, 361–370. [Google Scholar] [CrossRef]
- Belden, L.; Wildy, E.; Hatch, A.; Blaustein, A. Juvenile western toads, Bufo boreas, avoid chemical cues of snakes fed juvenile, but not larval, conspecifics. Anim. Behav. 2000, 59, 871–875. [Google Scholar] [CrossRef] [Green Version]
- Kiesecker, J.M.; Chivers, D.P.; Anderson, M.; Blaustein, A.R. Effect of predator diet on life history shifts of red-legged frogs, Rana aurora. J. Chem. Ecol. 2002, 28, 1007–1015. [Google Scholar] [CrossRef]
- Li, D.; Jackson, R.R. Influence of diet-related chemical cues from predators on the hatching of egg-carrying spiders. J. Chem. Ecol. 2005, 31, 333–342. [Google Scholar] [CrossRef]
- Ferland-Raymond, B.; Murray, D.L. Predator diet and prey adaptive responses: Can tadpoles distinguish between predators feeding on congeneric vs. conspecific prey? Can. J. Zool. 2008, 86, 1329–1336. [Google Scholar] [CrossRef]
- Ott, J. The expansion of Crocothemis erythraea (Brullé, 1832) in Germany-an indicator of climatic changes. In Odonata: Biology of Dragonflies; Scientific Publishers: Jodhpur, India, 2007; pp. 201–222. [Google Scholar]
- Korkeamäki, E.; Suhonen, J. Distribution and habitat specialization of species affect local extinction in dragonfly Odonata populations. Ecography 2002, 25, 459–465. [Google Scholar] [CrossRef]
- Trussell, G.C.; Ewanchuk, P.J.; Matassa, C.M. The fear of being eaten reduces energy transfer in a simple food chain. Ecology 2006, 87, 2979–2984. [Google Scholar] [CrossRef]
Figure 1.
Historical changes of the leading elevational edge (95th percentile) of Sympetrum striolatum in Switzerland between 1990 and 2013. The linear regression was positively significant (R2 = 0.21, p = 0.02), suggesting a historical shift towards higher elevations.
Figure 1.
Historical changes of the leading elevational edge (95th percentile) of Sympetrum striolatum in Switzerland between 1990 and 2013. The linear regression was positively significant (R2 = 0.21, p = 0.02), suggesting a historical shift towards higher elevations.

Figure 2.
Growth rate of Sympetrum striolatum larvae in the combination of two temperature and three predator treatments. Error bars are 95% confidence intervals.
Figure 2.
Growth rate of Sympetrum striolatum larvae in the combination of two temperature and three predator treatments. Error bars are 95% confidence intervals.

Figure 3.
Growth efficiency of Sympetrum striolatum larvae in the combination of two temperature and three predator treatments. Error bars are 95% confidence intervals.
Figure 3.
Growth efficiency of Sympetrum striolatum larvae in the combination of two temperature and three predator treatments. Error bars are 95% confidence intervals.

Figure 4.
Food intake (corrected for body mass) of Sympetrum striolatum larvae in the combination of two temperature and three predator treatments. Error bars are 95% confidence intervals.
Figure 4.
Food intake (corrected for body mass) of Sympetrum striolatum larvae in the combination of two temperature and three predator treatments. Error bars are 95% confidence intervals.

Figure 5.
Length of the 8th (a) and 9th (b) lateral spine of Sympetrum striolatum larvae in the combination of two temperature and three predator treatments. Error bars are 95% confidence intervals.
Figure 5.
Length of the 8th (a) and 9th (b) lateral spine of Sympetrum striolatum larvae in the combination of two temperature and three predator treatments. Error bars are 95% confidence intervals.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zebsa, R.; Mahdjoub, H.; Khelifa, R. Similar Response of a Range Expanding Dragonfly to Low- and High-Elevation Predators. Diversity 2022, 14, 302. https://0-doi-org.brum.beds.ac.uk/10.3390/d14040302
AMA Style
Zebsa R, Mahdjoub H, Khelifa R. Similar Response of a Range Expanding Dragonfly to Low- and High-Elevation Predators. Diversity. 2022; 14(4):302. https://0-doi-org.brum.beds.ac.uk/10.3390/d14040302
Chicago/Turabian StyleZebsa, Rabah, Hayat Mahdjoub, and Rassim Khelifa. 2022. "Similar Response of a Range Expanding Dragonfly to Low- and High-Elevation Predators" Diversity 14, no. 4: 302. https://0-doi-org.brum.beds.ac.uk/10.3390/d14040302
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.