
Two New Species of Dacrymyces (Dacrymycetales, Basidiomycota) from Southwestern China


1
Institute of Microbiology, School of Ecology and Nature Conservation, Beijing Forestry University, Beijing 100083, China
2
College of Forestry, Beijing Forestry University, Beijing 100083, China
*
Author to whom correspondence should be addressed.
Diversity 2022, 14(5), 379; https://0-doi-org.brum.beds.ac.uk/10.3390/d14050379
Submission received: 24 March 2022
/
Revised: 25 April 2022
/
Accepted: 8 May 2022
/
Published: 11 May 2022
(This article belongs to the Special Issue Diversity and Evolution of Fungi)
Abstract
:Two new species of Dacrymyces, D. cerebriformis and D. sinostenosporus, are presented from southwestern China, based on morphological characteristics and phylogenetic analyses. Dacrymyces cerebriformis is characterized by obviously cerebriform basidiomata when mature, hyphae without clamp connections, and hyaline, thin-walled, allantoid, 0–7-septate basidiospores (18.4–23.1 × 5.5–7.7 μm). Dacrymyces sinostenosporus is characterized by discoid and applanate basidiomata when mature, hyphae without clamp connections, and hyaline, thin-walled, cylindrical to allantoid, 0–7-septate basidiospores (18.0–23.5 × 6.3–8.0 μm). Dacrymyces cerebriformis can be distinguished from D. sinostenosporus by the narrower thin- or thick-walled terminal cells in the marginal hyphae of the sterile surface (1.7–3.5 μm vs. 3.5–6.0 μm in diameter) and thinner basidiospores (Q = 3.14–3.28 vs. Q = 2.63–2.77). A phylogenetic analysis of Dacrymyces is performed by using a dataset composed of concatenated internal transcribed spacer regions (ITS) and a large subunit (nrLSU) of ribosomal DNA gene, and two new species nested in two distinct lineages with robust support. A full description and illustrations of the two new species are provided here.
1. Introduction
Dacrymyces Nees was established to accommodate a single species, D. stillatus Nees, which is characterized by sessile, pustulate to pulvinate, yellowish, gelatinous basidiomata when fresh, hyphae without clamp connections, bifurcate basidia, and septate basidiospores [1]. Plenty dacrymycetous fungi with bifurcate basidia and septate basidiospores have been found, some related to genera including Ditiola Fr., Femsjonia Fr., Cerinomyces (G.W. Martin), and Dacryopinax (G.W. Martin et al.), having been established and recognized [2,3,4,5,6,7,8,9]. Some new species were described in Dacrymyces, and its generic limit became broader [10,11,12,13]. Nowadays, a total of 114 Dacrymyces taxa has been accepted according to Index Fungorum, and they have been found to be widely distributed around the world, mostly in Europe, America, Asia, and Oceania [10,11,12,13,14,15,16,17,18]. Species in the genus usually inhabit dead trees, stumps, fallen trunks, or branches of angiosperms or gymnosperms, and play a key role in the wood decomposition process, causing a brown rot [14,17].
In addition, species of Dacrymyces, in the present sense of the genus, have diverse basidiomata, and several species with clamped hyphae have been described in the genus [14,15,16,17,18]. In the phylogenetic analysis, Dacrymyces was suggested to belong to a polyphyletic group through sequence analyses of the D1/D2 domains of the large subunit (nrLSU) of ribosomal DNA gene [19]. Most species in Dacrymyces are found nested in Dacrymycetaceae, and a few species nest in Cerinomycetaceae or form a separate clade in Dacrymycetes [14,19]. Those species that nest out of Dacrymycetaceae transfer to Cerinomyces or Unilacryma (Shirouzu et al. [15,18]). However, Dacrymyces is still of polyphyletic origin, scattering in Dacrymycetaceae in the phylogeny, and no phenotypic features can be used to classify Dacrymyces and its related genera [14,16,18,19]. Therefore, the genus is still ambiguous in taxonomy and phylogeny.
In the present study, five specimens with typical characteristics of Dacrymyces are collected from Yunnan province in southwestern China, which is rich in gelatinous wood-decaying fungi [20,21]. In the phylogenetic analyses of Dacrymyces, using a dataset composed of concatenated ITS and nrLSU genes, the sequences of four specimens collected by us form two distinct lineages with strong support. Although another specimen’s, Dai 19805, DNA sequence extractions failed, the five specimens were identified as two new species, viz. D. cerebriformis and D. sinostenosporus, based on their morphological characteristics and a phylogenetic analysis. The aim of this article is to provide detailed information on the phylogenetic position and morphological characteristics of the two new species.
2. Materials and Methods
2.1. Sample Collection
Samples of dacrymycetous fungi were collected from coniferous, deciduous broad-leaved, and evergreen broad-leaved forests in subtropical Yunnan province, southwestern China. When sampling, basidiomata were collected together with the substrate. When possible, the corresponding tree species to the substrate were identified in the forests. Collected samples were dried by a mushroom dryer at a temperature of approximately 35 °C, and the dry samples were placed in an envelope and transported to the laboratory.
2.2. Morphological Studies
A morphological description was performed following work by Shirouzu et al. [15]. Macro-morphological characteristics were observed and described based on the field-collected fresh specimens and photos, and the color names followed Petersen’s [22] work. Micro-morphological characteristics were obtained from dried specimens. Hand-cut sections were rehydrated with 5% KOH and stained with 2% Phloxine B (C20H2Br4Cl4Na2O5) reagent. Microscopic structures, such as basidia, basidiospores, and hyphae, were observed and photographed by the Nikon Eclipse 80i microscope (magnifications × ≤ 1000) with a Nikon Digital Sight DS-Fi2 camera. The following abbreviations were used: L—mean length; W—mean width; Q—L/W ratio for the given specimen; n (a/b)— number of spores (a) measured from given number of specimens (b). The specimens were deposited in the herbarium of Beijing Forestry University (BJFC).
2.3. DNA Extraction, Polymerase Chain Reaction, and Sequencing
Total genomic DNA were extracted from the dried specimens using the CTAB rapid plant genome extraction kit (Aidlab Biotechnologies Co. Ltd, Beijing, China). The DNA sequence data were obtained from two regions. The internal transcribed spacer regions (ITS) were amplified with primer pairs ITS5/ITS4 [23], and the primer pairs LR0R/LR7 were used to amplify the large subunit of nuclear ribosomal DNA gene (nrLSU) [24].
Polymerase chain reaction (PCR) was performed using S1000™ Thermal Cycler (Bio-Rad Laboratories, Hercules, CA, USA). PCR was performed in Eppendorf tubes containing 30 μL of reaction mixture with the following composition: 1 μL extracted DNA, 12 μL ddH2O, 15 μL 2 × EasyTaq PCR Supermix (TransGen Biotech Co., Ltd., Beijing, China), and 1 μL of each primer. The PCR procedure for ITS was as follows: initial denaturation at 95 °C for 3 min, followed by 35 cycles of 94 °C for 40 s, 54 °C for 45 s, and 72 °C for 1 min, with a final extension at 72 °C for 10 min. The PCR procedure for nrLSU was as follows: initial denaturation at 94 °C for 1 min, followed by 35 cycles of 94 °C for 1 min, 50 °C for 1 min, and 72 °C for 1 min, and a final extension at 72 °C for 10 min. The PCR products were purified and sequenced in Beijing Genomics Institute, China, with the same primers.
2.4. Sequence Alignment and Phylogenetic Analysis
The sequences obtained from this study and downloaded from GenBank are listed in Table 1. Sequences were aligned with the MAFFT version 7 web tool (http://mafft.cbrc.jp/alignment/server/, accessed on 10 March 2022) using the Q-INS-i option for ITS and nrLSU. Coprinus comatus (O.F. Müll.) Pers. and Suillus pictus (Peck) Kuntze were used as the outgroups following work by Shirouzu et al. [15]. The combined ITS and nrLSU alignment contained 1588 characters (including gaps). Sequence alignment was deposited at TreeBase (http://purl.org/phylo/treebase, accessed on 14 March 2022; submission ID 29541).
Phylogenetic analyses were performed using maximum likelihood (ML) and Bayesian inference (BI) methods based on the combined ITS and nLSU dataset. All characters were equally weighted and gaps were treated as missing data. The optimal substitution model suitable for the combined dataset was determined using the AIC (Akaike information criterion) implemented in MrModeltest 2.3 [25]. The model GTR + I + G was selected for the combined ITS and nLSU dataset. RAxMLGUI 1.2 was used for ML analysis [26,27]. All parameters in the ML analysis used default settings. Statistical support values were obtained using non-parametric bootstrapping with 1000 replicates. The BI analysis was conducted using MrBayes 3.2.5 [28]. Two independent runs were performed, each starting from random trees with four chains for five million generations. Trees were sampled every 1000 generations. The first quarter of sampled trees was discarded as burn-ins, and the remaining trees were used to construct a 50% majority consensus tree and calculate BPP (Bayesian posterior probabilities). A majority rule tree consensus of all remaining trees was calculated. Branches were considered as significantly supported if they received bootstrap support (BS) for ML analysis and Bayesian posterior probabilities (BPP) greater than or equal to 75% (BS) and 0.90 (BPP).
3. Results
3.1. Phylogenetic Results
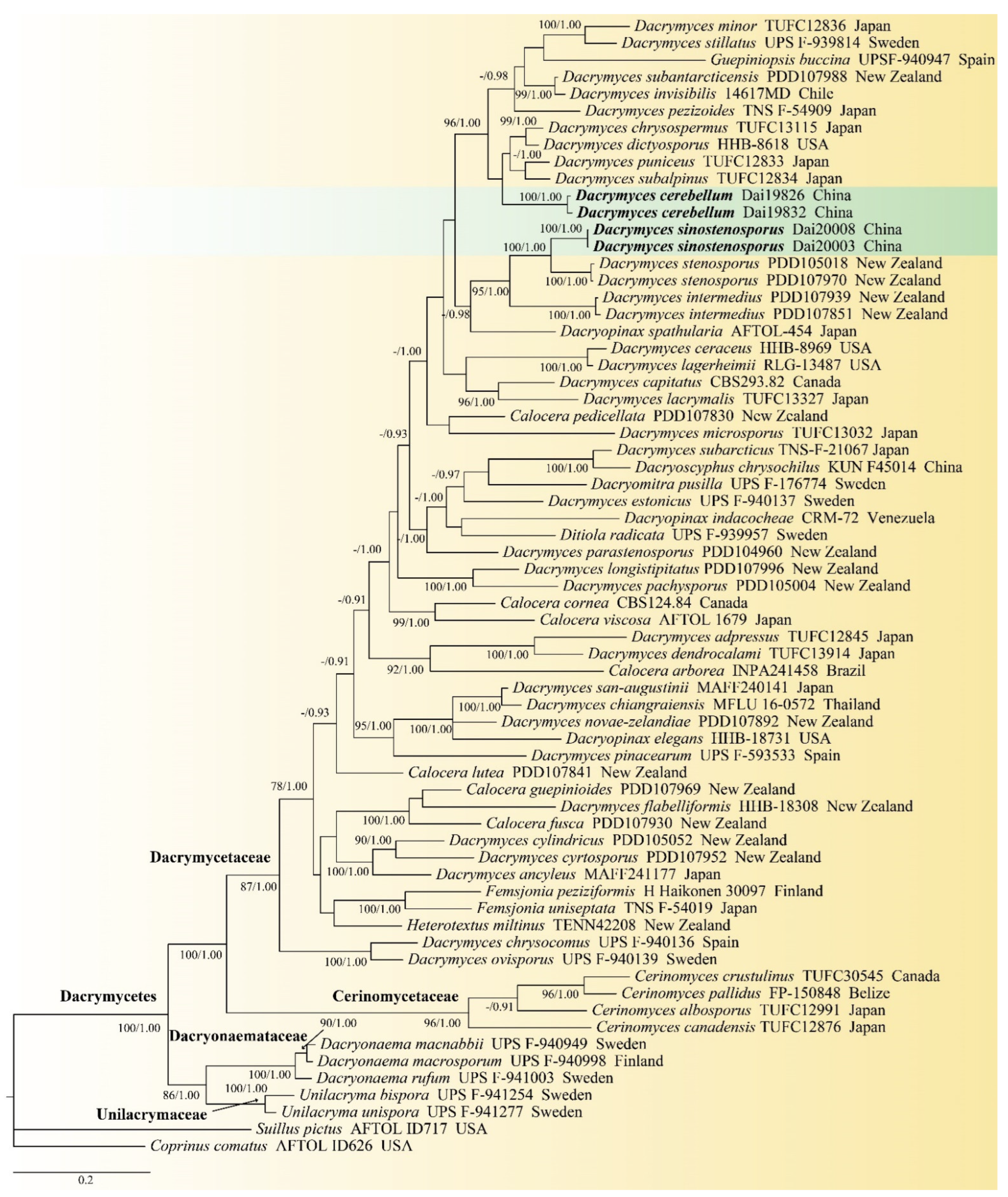
Sixty-seven fungal sequences that represented 63 species were included in the combined ITS and nrLSU dataset to construct the phylogenetic tree. The newly generated sequences of four Chinese specimens were included in the combined ITS and nrLSU dataset, because extracting DNA from specimen Dai 19805 failed. The BI analysis resulted in a similar topology as the ML analysis; thus, only the BI tree was presented (Figure 1). The phylogeny (Figure 1) demonstrated that samples of 35 Dacrymyces species were gathered into Dacrymycetaceae. The four specimens representing two new species, Dacrymyces cerebriformis and D. sinostenosporus, belonged to two distinct lineages with robust supports (100% BS, 1.00 BPP; 100% BS, 1.00 BPP) in Dacrymycetaceae. Dacrymyces sinostenosporus was found to be closely related to D. stenosporus with high support (100% BS, 1.00 BPP).
3.2. Taxonomy
Dacrymyces cerebriformis F. Wu and Y.P. Lian sp. nov. Figure 2.
MycoBank no.: 843769
Holotype—China, Yunnan province, Pingbian county, Daweishan National Forest Park, on fallen angiosperm branch, 26 June 2019, Dai 19832 (BJFC031507).
Etymology—Refers to the species having obviously cerebriform basidiomata when mature.
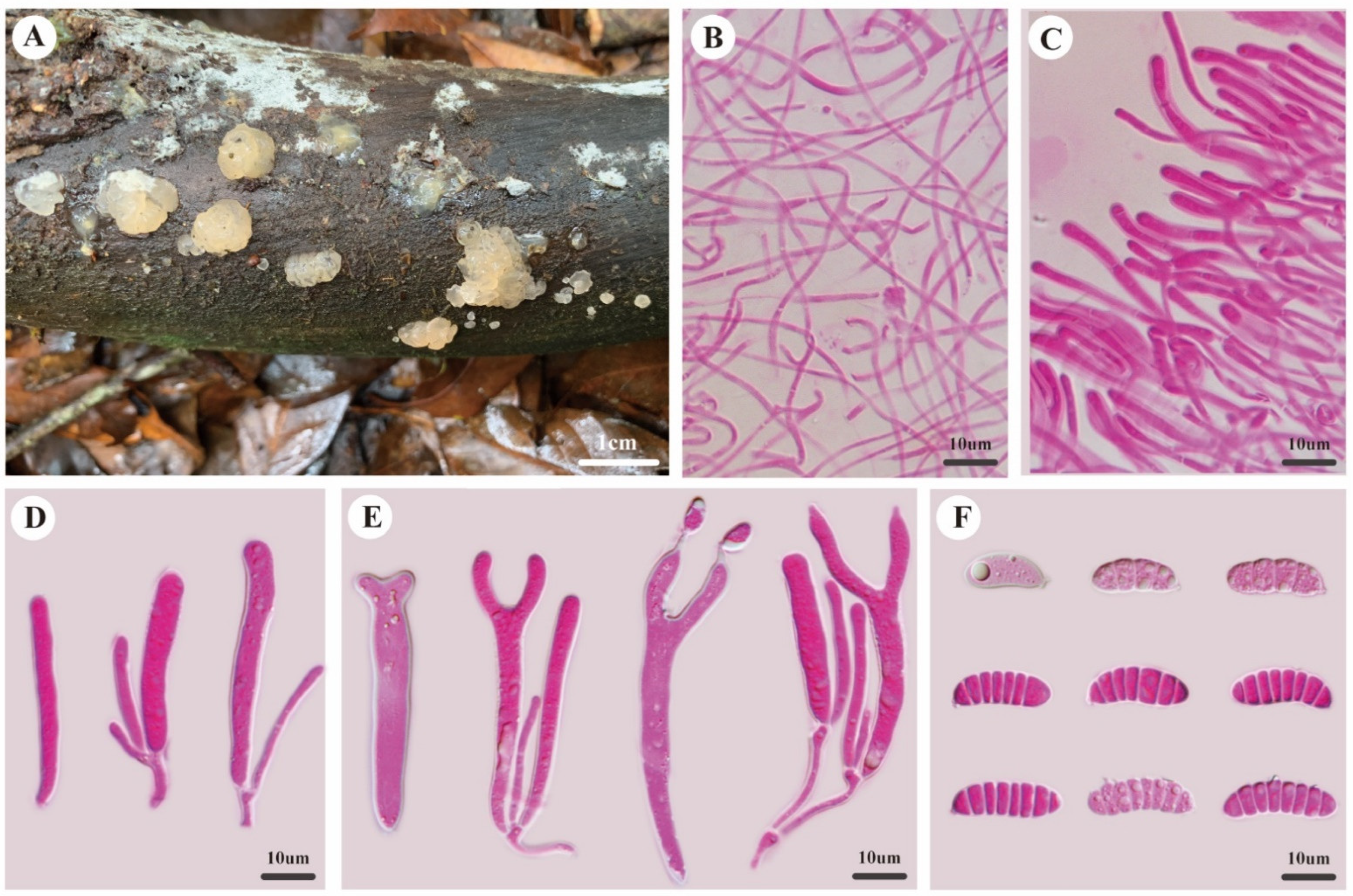
Description—Basidioma sessile, single or caespitose, gelatinous when fresh, cream to buff, pustulate to pulvinate when juvenile, obviously cerebriform when mature, up to 3.0 mm high, 5.0–10.0 mm in diam. Marginal hyphae of sterile surface hyaline, thin- to thick-walled, septate, straight or flexuous, with simple cylindrical thin- or thick-walled terminal cells measuring 11.0–18.0 × 1.7–3.5 μm, without clamp connections. Internal hyphae hyaline, thin-walled, septate, branched, without clamp connections, 1.0–3.0 μm in diam. Hymenium limited to the upper surface of the basidioma, composed of basidia and simple cylindrical hyphidia. Basidia hyaline, thin-walled, cylindrical to clavate, bifurcate when mature, 35.0–50.0 × 4.5–7.0 μm, probasidia similar to basidia in shape, but smaller. Basidiospores hyaline, thin-walled, allantoid, with an apiculum at the base, 0–7-septate, (18.1–)18.4–23.1(–23.8) × (5.4–)5.5–7.7(–8.0) μm, L = 20.7 μm, W = 6.5 μm, Q = 3.14–3.28 (n = 60/2).
Additional specimens examined—China, Yunnan province, Pingbian county, Daweishan National Forest Park, on rotten angiosperm wood, 26 June 2019, Dai 19805 (BJFC031480), Dai 19826 (BJFC 031501).
Notes—Dacrymyces cerebriformis is characterized by obviously cerebriform basidiomata when mature, hyphae without clamp connections, and hyaline, thin-walled, allantoid, 0–7-septate basidiospores. The species resembles D. adpressus Grognot and D. sichuanensis B. Liu and L. Fan by sharing cerebriform basidiomata when mature, but both differ from D. cerebriformis by their 0–3-septate basidiospores [13,14].
Dacrymyces sinostenosporus F. Wu and Y.P. Lian, sp. nov. Figure 3.
MycoBank no.: 838634
Holotype—China, Yunnan province, Wenshan county, Laojunshan Nature Reserve, on fallen branch of Lithocarpus, 30 June 2019, Dai 20003 (BJFC031677).
Etymology—Refers to the species similar to Dacrymyces stenosporus, but occurring in China.
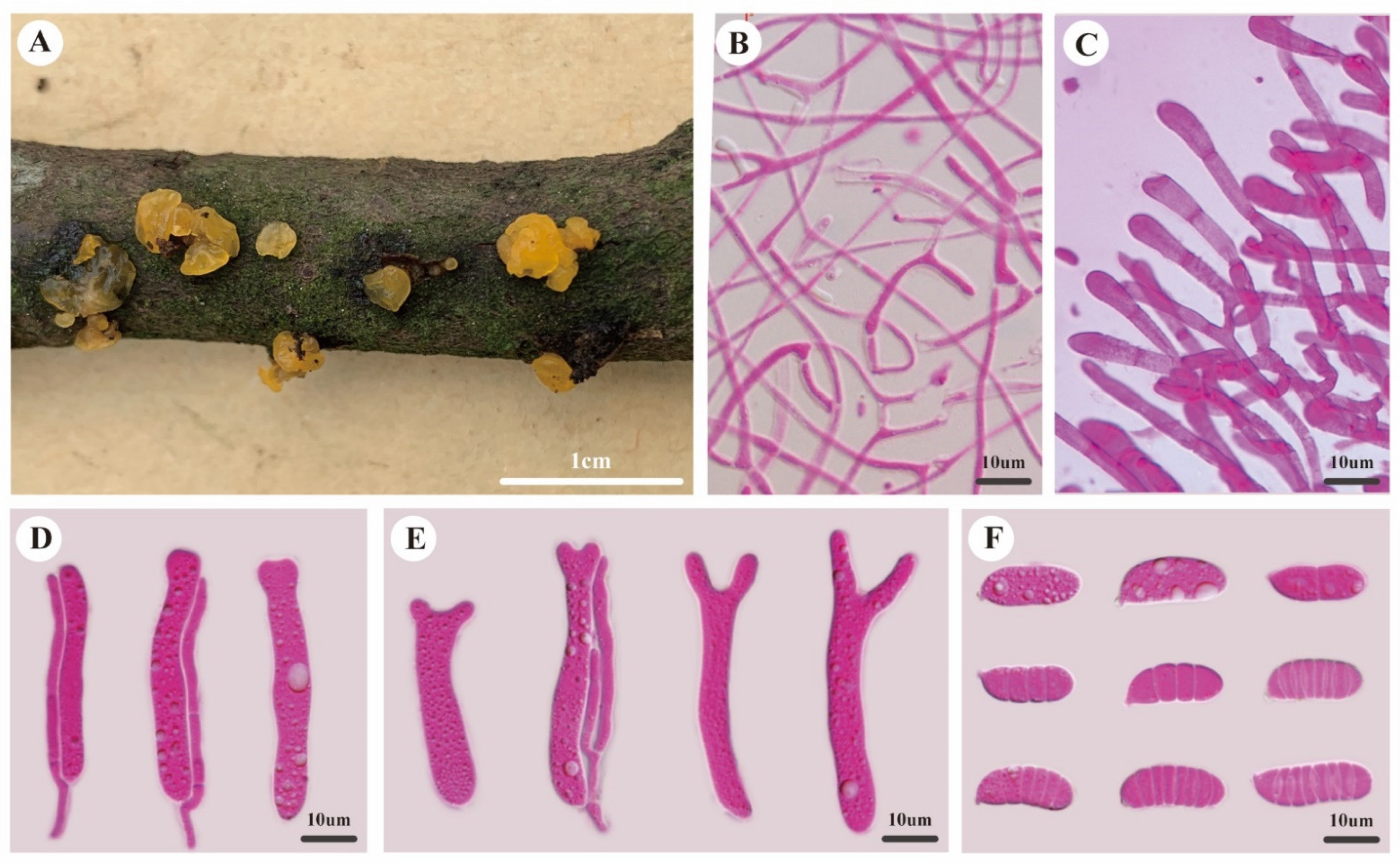
Description—Basidiomata sessile, single or caespitose, gelatinous when fresh, curry-yellow to orange yellow, pustulate to pulvinate when juvenile, discoid and applanate when mature, sometimes slightly cerebriform when coalescing, up to 1.2 mm high, 2–4 mm in diam. Marginal hyphae of sterile surface hyaline, thin- to thick-walled, septate, straight or flexuous, with cylindrical, thin-walled terminal cells measuring 7.0–17.0 × 3.5–6.0 μm, without clamp connections. Internal hyphae hyaline, thin-walled, septate, branched, without clamp connections, 0.8–3 μm in diam. Hymenium limited to the upper surface of the basidiomata, composed of basidia and simple or branched cylindrical hyphidia. Basidia hyaline, thin-walled, cylindrical to clavate, becoming bifurcate when mature, 40.0–60.0 × 4.0–9.0 μm, probasidia similar to basidia in shape, but smaller. Basidiospores hyaline, thin-walled, cylindrical to allantoid, with an apiculum at the base, 0–7-septate, (17.0–)18.0–23.5(–24.0) × (5.9–)6.3–8.0(–8.4) μm, L = 19.3 μm, W = 7.1 μm, Q = 2.63–2.77 (n = 60/2).
Additional specimen examined—China, Yunnan province, Wenshan county, Laojunshan Nature Reserve, on fallen angiosperm branch, 30 June 2019, Dai 20008 (BJFC031682).
Notes—Dacrymyces sinostenosporus is characterized by discoid and applanate basidiomata when mature, hyphae without clamp connections, and hyaline, thin-walled, cylindrical to allantoid, 0–7-septate basidiospores. The species is closely related to D. stenosporus in morphology and phylogeny, but D. stenosporus has shorter basidia (30–40 × 4 μm vs. 40.0–60.0 × 4.0–9.0 μm), is smaller, has 0–3-septate basidiospores (13–17 × 5–6 μm vs. 18.0–23.5 × 6.3–8 μm) [16], and it shares less than 90% similarity to D. stenosporus in ITS sequences.
4. Discussion
In order to improve knowledge on Dacrymyces, five specimens of the genus were collected from Yunnan province, China, which were identified as two new species, viz. D. cerebriformis and D. sinostenosporus, based on morphological characteristics and phylogenetic analyses using the combined ITS and nrLSU dataset. The two new species have typical characteristics of Dacrymyces, and they share 0–7-septate basidiospores, but they are phylogenetically distant from each other. Dacrymyces cerebriformis can be distinguished from D. sinostenosporus by its narrower, thin- or thick-walled terminal cells in the marginal hyphae of the sterile surface (1.7–3.5 μm vs. 3.5–6.0 μm), and thinner basidiospores (Q = 3.14–3.28 vs. Q = 2.63–2.77).
Five Dacrymyces species were described from China, but DNA data of these species were not available for a phylogenetic analysis. This study firstly obtained molecular data of Dacrymyces samples from China, which contributed to the knowledge of the species diversity in China. Among those previously described, Dacrymyces species from China, D. yunnanensis (B. Liu and L. Fan), and D. sichuanensis were also originally described from southwestern China, but D. yunnanensis differed from the two new species by having ellipsoid to subglobose basidiospores, with transverse and vertical septations (15.6–24.7 × 13–16 μm), and D. sichuanensis was readily distinguished from the two new species by the presence of clamp connections and 0–3-septate basidiospores [12,13]. Therefore, seven Dacrymyces species, including D. dendrocalami Oberw., D. duii (B. Liu and J.Z. Cao.), D. cerebriformis, D. pengii (B. Liu and L. Fan) A. Savchenko, D. sichuanensis, D. sinostenosporus and D. yunnanensis, have been found in China to date.
Other species, including D. stillatus, D. lacrymalis (Pers.) Nees, D. minor (Peck), D. subantarcticensis (Burds. and Laursen), D. novae-zelandiae (McNabb), and D. san-augustinii (Kobayasi), may be confused with D. cerebriformis and D. sinostenosporus due to sharing sessile, pulvinate, yellowish basidiomata when fresh [14,29], but the former four species differ from the two new species by having 0–3-septate basidiospores, D. novae-zelandiae differs by its longer basidia (55–72 μm vs. 35–40 μm in D. cerebriformis; 40–60 μm in D. sinostenosporus), and D. san-augustinii differs from D. cerebriformis and D. sinostenosporus in that it has wider and curved allantoid basidiospores (16–27.5 × 6–10 μm) [14].
The polyphyletic problem of Dacrymyces was still not well solved, and species in different genera of Dacrymycetaceae overlapped in phylogenies [17,18]. More samples with DNA data from different areas of the world are badly needed, and extensive examinations and phylogenetic analyses on these samples should be carried out in the future. The species presented here were intended to contribute to the understanding of the great diversity and taxonomy of the genus Dacrymyces, but further studies should analyze them as a whole.
Author Contributions
Conceptualization, F.W.; methodology, F.W. and Y.-P.L.; software, Y.-P.L.; investigation, F.W., Y.-P.L., A.T., and F.W.; validation, F.W. and Y.-P.L.; formal analysis, F.W. and Y.-P.L.; resources, F.W., Y.-P.L., and A.T.; writing—original draft preparation, F.W. and Y.-P.L.; writing—review and editing, F.W.; visualization, F.W. and Y.-P.L.; supervision, F.W.; project administration, F.W.; funding acquisition, F.W. All authors have read and agreed to the published version of the manuscript.
Funding
The research was financed by the Fundamental Research Funds for the Central Universities (no. 2021ZY91), and the National Natural Science Foundation of China (project nos. 31701978 and 32070006).
Institutional Review Board Statement
Not applicable for studies involving humans or animals.
Informed Consent Statement
Not applicable for studies involving humans.
Data Availability Statement
All data generated or analyzed during this study were included in this published article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nees von Esenbeck, C.G. Das System der Pilze und Schwämme; Stahelschen Buchhandlung: Würtzburg, Germany, 1816; Volume 2, pp. 1–329. [Google Scholar]
- Fries, E.M. Systema Mycologicum; Officina Berlingiana: Berlin, Germany, 1822; Volume 2, pp. 1–275. [Google Scholar]
- Fries, E.M. Summa Vegetabilium Scandinaviae; Bonnier: Stockholm, Sweden, 1849; Volume 2, pp. 259–572. [Google Scholar]
- Martin, G.W. New or noteworthy tropical fungi. IV. Lloydia 1948, 11, 111–122. [Google Scholar]
- Martin, G.W. The genus Ceracea Cragin. Mycologia 1949, 41, 77–86. [Google Scholar] [CrossRef]
- McNabb, R.F.R. Taxonomic studies in the Dacrymycetaceae I. Cerinomyces Martin. New Zealand J. Bot. 1964, 2, 415–424. [Google Scholar] [CrossRef]
- McNabb, R.F.R. Taxonomic studies in the Dacrymycetaceae III. Dacryopinax Martin. New Zealand J. Bot. 1965, 3, 59–72. [Google Scholar] [CrossRef] [Green Version]
- McNabb, R.F.R. Taxonomic studies in the Dacrymycetaceae VI. Femsjonia Fries. New Zealand J. Bot. 1965, 3, 223–228. [Google Scholar] [CrossRef]
- McNabb, R.F.R. Taxonomic studies in the Dacrymycetaceae VII. Ditiola Fries. New Zealand J. Bot. 1966, 4, 546–558. [Google Scholar] [CrossRef]
- Kobayasi, Y. On the Dacrymyces-group (Fungorum ordinis Tremellalium studia monographica III). Sci. Rep. Tokyo Bunrika Daigaku Sect. B 1939, 4, 105–128. [Google Scholar]
- Mcnabb, R.F.R. Taxonomic studies in the Dacrymycetaceae VIII. Dacrymyces Nees ex Fries. N. Z. J. Bot. 1973, 11, 461–564. [Google Scholar] [CrossRef]
- Liu, B.; Fan, L. Two new species of Dacrymycetaceae from China. Acta Microbiol. Sin. 1989, 8, 22–24. [Google Scholar]
- Liu, B.; Fan, L. Four new species and one new variety of Dacrymycetaceae from China. Acta Microbiol. Sin. 1990, 9, 12–19. [Google Scholar]
- Shirouzu, T.; Hirose, D.; Tokumasu, S. Taxonomic study of the Japanese Dacrymycetes. Persoonia 2009, 23, 16–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirouzu, T.; Hirose, D.; Oberwinkler, F.; Shimomura, N.; Maekawa, N.; Tokumasu, S. Combined molecular and morphological data for improving phylogenetic hypothesis in Dacrymycetes. Mycologia 2013, 105, 1110–1125. [Google Scholar] [CrossRef]
- Shirouzu, T.; Hosaka, K.; Nam, K.O.; Weir, B.S.; Johnston, P.R.; Hosoya, T. Phylogenetic relationships of eight new Dacrymycetes collected from New Zealand. Persoonia 2017, 38, 156–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamora, J.C.; Ekman, S. Phylogeny and character evolution in the Dacrymycetes, and systematics of Unilacrymaceae and Dacryonaemataceae fam. nov. Persoonia 2020, 44, 161–205. [Google Scholar] [CrossRef] [PubMed]
- Savchenko, A.; Zamora, J.C.; Shirouzu, T.; Spirin, V.; Malysheva, V.; Kõljalg, U.; Miettinen, O. Revision of Cerinomyces (Dacrymycetes, Basidiomycota) with notes on morphologically and historically related taxa. Stud. Mycol. 2021, 99, 100117. [Google Scholar] [CrossRef] [PubMed]
- Shirouzu, T.; Hirose, D.; Tokumasu, S. Sequence analyses of the 28S rRNA gene D1/D2 region suggest Dacrymyces (Heterobasidiomycetes, Dacrymycetales) is polyphyletic. Mycoscience 2007, 48, 388–394. [Google Scholar] [CrossRef]
- Dai, Y.C.; Yang, Z.L.; Cui, B.K.; Wu, G.; Yuan, H.S.; Zhou, L.W.; He, S.H.; Ge, Z.W.; Wu, F.; Wei, Y.L.; et al. Diversity and systematics of the important macrofungi in Chinese forests. Mycosystema 2021, 40, 770–805. [Google Scholar] [CrossRef]
- Fan, L.F.; Wu, Y.D.; Wu, F.; Dai, Y.C. Calocera tibetica sp. nov. (Dacrymycetaceae, Dacrymycetales) from southwestern China. Phytotaxa 2021, 500, 133–141. [Google Scholar] [CrossRef]
- Petersen, J.H. Farvekort. In The Danish Mycological Society’s Colour-Chart; Foreningen til Svampekundskabens Fremme: Greve, Denmark, 1996. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols, a Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analysis with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Silvestro, D.; Michalak, I. Raxmlgui: A graphical front-end for raxml. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burdsall, H.; Laursen, G.A.; Cripps, C.L. Fungi of New Zealand’s subantarctic islands. I: Two new species of Dacrymyces (Basidiomycotina). In Fungi in Forest Ecosystems: Systematics, Diversity and Ecology; Miller, O.K., Cripps, C.L., Eds.; New York Botanical Garden: New York, NY, USA, 2004; pp. 107–111. [Google Scholar]
Figure 1.
Phylogeny of Dacrymyces and related species generated by BI analysis based on the combined ITS and nrLSU dataset. Bootstrap support values for ML ≥ 75% and BI ≥ 0.90 were given near nodes, respectively. Two new species are in bold. Note: “-” means bootstrap without robust support (BS < 75%, BPP < 0.90); the arrows pointed the nodes.
Figure 1.
Phylogeny of Dacrymyces and related species generated by BI analysis based on the combined ITS and nrLSU dataset. Bootstrap support values for ML ≥ 75% and BI ≥ 0.90 were given near nodes, respectively. Two new species are in bold. Note: “-” means bootstrap without robust support (BS < 75%, BPP < 0.90); the arrows pointed the nodes.

Figure 2.
Morphology of Dacrymyces cerebriformis. (A) Basidiomata. (B) Internal hyphae. (C) Marginal hyphae. (D) Probasidia and hyphidia. (E) Developing basidia and hyphidia. (F) Basidiospores.
Figure 2.
Morphology of Dacrymyces cerebriformis. (A) Basidiomata. (B) Internal hyphae. (C) Marginal hyphae. (D) Probasidia and hyphidia. (E) Developing basidia and hyphidia. (F) Basidiospores.

Figure 3.
Morphology of Dacrymyces sinostenosporus. (A) Basidiomata. (B) Internal hyphae. (C) Marginal hyphae. (D) Probasidia and hyphidia. (E) Developing basidia and hyphidia. (F) Basidiospores.
Figure 3.
Morphology of Dacrymyces sinostenosporus. (A) Basidiomata. (B) Internal hyphae. (C) Marginal hyphae. (D) Probasidia and hyphidia. (E) Developing basidia and hyphidia. (F) Basidiospores.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Taxa information and GenBank accession numbers of the sequences used in this study.
| Species | Locality | Sample No. | GenBank Accession No. | |
|---|---|---|---|---|
| ITS | nrLSU | |||
| Calocera arborea | Brazil | INPA 241458 | AB744230 | AB723514 |
| Calocera cornea | Canada | CBS124.84 | AB712437 | AB472738 |
| Calocera fusca | New Zealand | PDD 107930 | LC131405 | LC131364 |
| Calocera guepinioides | New Zealand | PDD 107969 | LC131411 | LC131370 |
| Calocera lutea | New Zealand | PDD 107841 | LC131413 | LC131372 |
| Calocera pedicellata | New Zealand | PDD 107830 | LC131415 | LC131374 |
| Calocera viscosa (Type species) | Japan | AFTOL-ID 1679 | DQ520102 | DQ520102 |
| Cerinomyces albosporus | Japan | TUFC12991 | AB712440 | AB299050 |
| Cerinomyces canadensis | Japan | TUFC12876 | AB712441 | AB472696 |
| Cerinomyces crustulinus | Canada | TUFC 30545 | AB712443 | AB712423 |
| Cerinomyces pallidus (Type species) | Belize | FP-150848 | AB712446 | AB712426 |
| Dacrymyces adpressus | Japan | TUFC12845 | AB712447 | AB472707 |
| Dacrymyces ancyleus | Japan | MAFF241177 | AB712448 | AB472713 |
| Dacrymyces capitatus | Canada | CBS293.82 | AB712450 | AB472741 |
| Dacrymyces ceraceus | USA | HHB-8969 | AB712442 | AB712422 |
| Dacrymyces cerebriformis | China | Dai 19826 | OM955201 | OM955196 |
| Dacrymyces cerebriformis | China | Dai 19832 | OM955202 | OM955197 |
| Dacrymyces chiangraiensis | Thailand | MFLU:16-0572 | KY498587 | – |
| Dacrymyces chrysocomus | Spain | UPS F-940136 | MN595629 | MN595629 |
| Dacrymyces chrysospermus | Japan | TUFC 13115 | AB712452 | AB299073 |
| Dacrymyces cylindricus | New Zealand | PDD 105052 | LC131419 | LC131378 |
| Dacrymyces cyrtosporus | New Zealand | PDD 107952 | LC131421 | LC131380 |
| Dacrymyces dendrocalami | Japan | TUFC13914 | AB712453 | AB712428 |
| Dacrymyces dictyosporus | USA | HHB-8618 | AB712454 | AB712429 |
| Dacrymyces estonicus | Sweden | UPS F-940137 | MN595632 | MN595632 |
| Dacrymyces flabelliformis | New Zealand | HHB-18308 | AB712455 | AB712430 |
| Dacrymyces intermedius | New Zealand | PDD 107939 | – | LC131385 |
| Dacrymyces intermedius | New Zealand | PDD 107851 | – | LC131384 |
| Dacrymyces invisibilis | Chile | 14617MD | MH230101 | MH230103 |
| Dacrymyces lacrymalis | Japan | TUFC13327 | AB712456 | AB299069 |
| Dacrymyces lagerheimii | USA | RLG-13487 | AB712445 | AB712425 |
| Dacrymyces longistipitatus | New Zealand | PDD 107996 | LC131425 | LC131386 |
| Dacrymyces microsporus | Japan | TUFC 13032 | AB712457 | AB472712 |
| Dacrymyces minor | Japan | TUFC 12836 | AB712458 | AB299063 |
| Dacrymyces novae-zelandiae | New Zealand | PDD 107892 | LC131427 | LC131390 |
| Dacrymyces ovisporus | Sweden | UPS F-940139 | MN595635 | MN595635 |
| Dacrymyces pachysporus | New Zealand | PDD 105004 | LC131429 | LC131392 |
| Dacrymyces parastenosporus | New Zealand | PDD 104960 | LC131431 | LC131394 |
| Dacrymyces pezizoides | Japan | TNS-F-54909 | LC386890 | LC386894 |
| Dacrymyces pinacearum | Japan | UPS F-593533 | MN595637 | MN595637 |
| Dacrymyces puniceus | Japan | TUFC 12833 | AB712449 | AB299057 |
| Dacrymyces san-augustinii | Japan | MAFF240141 | AB712463 | AB299081 |
| Dacrymyces sinostenosporus | China | Dai 20003 | MW540888 | MW540890 |
| Dacrymyces sinostenosporus | China | Dai 20008 | MW540889 | MW540891 |
| Dacrymyces stenosporus | New Zealand | PDD 105018 | LC131433 | LC131396 |
| Dacrymyces stenosporus | New Zealand | PDD 107970 | LC131434 | LC131397 |
| Dacrymyces stillatus (Type species) | Sweden | UPS F-939814 | MN595677 | MN595677 |
| Dacrymyces subalpinus | Japan | TUFC 12834 | AB712465 | AB299060 |
| Dacrymyces subantarcticensis | New Zealand | PDD 107988 | LC131436 | LC131400 |
| Dacrymyces subarcticus | Japan | TNS-F-21067 | AB712467 | AB472727 |
| Dacryomitra pusilla (Type species) | Sweden | UPS F-176774 | MN595639 | MN595639 |
| Dacryonaema macnabbii | Sweden | UPS F-940949 | MN595650 | MN595650 |
| Dacryonaema macrosporum | Finland | UPS F-940998 | MN595660 | MN595660 |
| Dacryonaema rufum (Type species) | Sweden | UPS F-941003 | MN595645 | MN595645 |
| Dacryopinax elegans (Type species) | USA | HHB-18731 | AB712471 | AB712433 |
| Dacryopinax indacocheae | Venezuela | CRM-72 | AB712472 | AB712434 |
| Dacryopinax spathularia | Japan | AFTOL-ID 454 | AY854070 | AY701525 |
| Dacryoscyphus chrysochilus (Type species) | China | KUN F45014 | – | AY604567 |
| Ditiola radicata (Type species) | Sweden | UPS F-939957 | MN595641 | MN595641 |
| Femsjonia peziziformis | Finland | H Haikonen 30097 | MN595642 | MN595642 |
| Femsjonia uniseptata | Japan | TNS-F-54019 | LC222844 | LC222843 |
| Guepiniopsis buccina | Spain | UPS F-940947 | MN595643 | MN595643 |
| Heterotextus miltinus | New Zealand | TENN 42208 | MN595644 | MN595644 |
| Unilacryma bispora | Sweden | UPS F-941254 | MN595673 | MN595673 |
| Unilacryma unispora (Type species) | Sweden | UPS F-941277 | MN595665 | MN593500 |
| Coprinus comatus | USA | AFTOL-ID 626 | AY854066 | AY635772 |
| Suillus pictus | USA | AFTOL-ID 717 | AY854069 | AY684154 |
Note: newly generated sequences for this study are in bold.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lian, Y.-P.; Tohtirjap, A.; Wu, F. Two New Species of Dacrymyces (Dacrymycetales, Basidiomycota) from Southwestern China. Diversity 2022, 14, 379. https://0-doi-org.brum.beds.ac.uk/10.3390/d14050379
AMA Style
Lian Y-P, Tohtirjap A, Wu F. Two New Species of Dacrymyces (Dacrymycetales, Basidiomycota) from Southwestern China. Diversity. 2022; 14(5):379. https://0-doi-org.brum.beds.ac.uk/10.3390/d14050379
Chicago/Turabian StyleLian, Ya-Ping, Ablat Tohtirjap, and Fang Wu. 2022. "Two New Species of Dacrymyces (Dacrymycetales, Basidiomycota) from Southwestern China" Diversity 14, no. 5: 379. https://0-doi-org.brum.beds.ac.uk/10.3390/d14050379
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.