
Human–Wildlife Conflicts across Landscapes—General Applicability vs. Case Specificity


1
Research Center [Sustainability–Transformation–Transfer], Eberswalde University for Sustainable Development, 16225 Eberswalde, Germany
2
Albrecht Daniel Thaer-Institute of Agricultural and Horticultural Sciences, Faculty of Life Sciences, Humboldt-Universität zu Berlin, 10009 Berlin, Germany
*
Author to whom correspondence should be addressed.
Diversity 2022, 14(5), 380; https://0-doi-org.brum.beds.ac.uk/10.3390/d14050380
Submission received: 22 April 2022
/
Revised: 6 May 2022
/
Accepted: 8 May 2022
/
Published: 11 May 2022
(This article belongs to the Special Issue Human Wildlife Conflict across Landscapes)
Abstract
:Here, we address the question of if/to what extent human–wildlife conflict (HWC) can be reduced or mitigated by supra-regional or even global approaches, or whether case- and region-specific strategies are necessary. First, we try to shift the perspective from humans towards wildlife and regard modern era (near) extinctions of selected wildlife species as an ultimate expression of HWC. We then reflect on the legitimacy of ecosystem comparisons beyond the borders of biogeographical regions—an important prerequisite for global approaches. In the following, we present two case scenarios that exemplarily illustrate the topic from different perspectives: carnivore–livestock conflicts (classical view, human perspective) and wind turbine-induced mortality of bats (wildlife perspective, rarely regarded as a form of HWC). On this basis, we try to develop a framework that enables a global approach, while adequately considering case- and region-specificity. We think that a globally valid and generally approachable dimension can be defined, as long as there is a natural link: in the present case the established monophyly of the orders Carnivora and Chiroptera, i.e., representatives descending from common ancestors, thus sharing common ecological features. This natural relationship among all representatives of a systematic group (specification of the “wildlife” concept) is key for the development of an overarching strategy that can be adjusted to a specific case.
1. Introduction
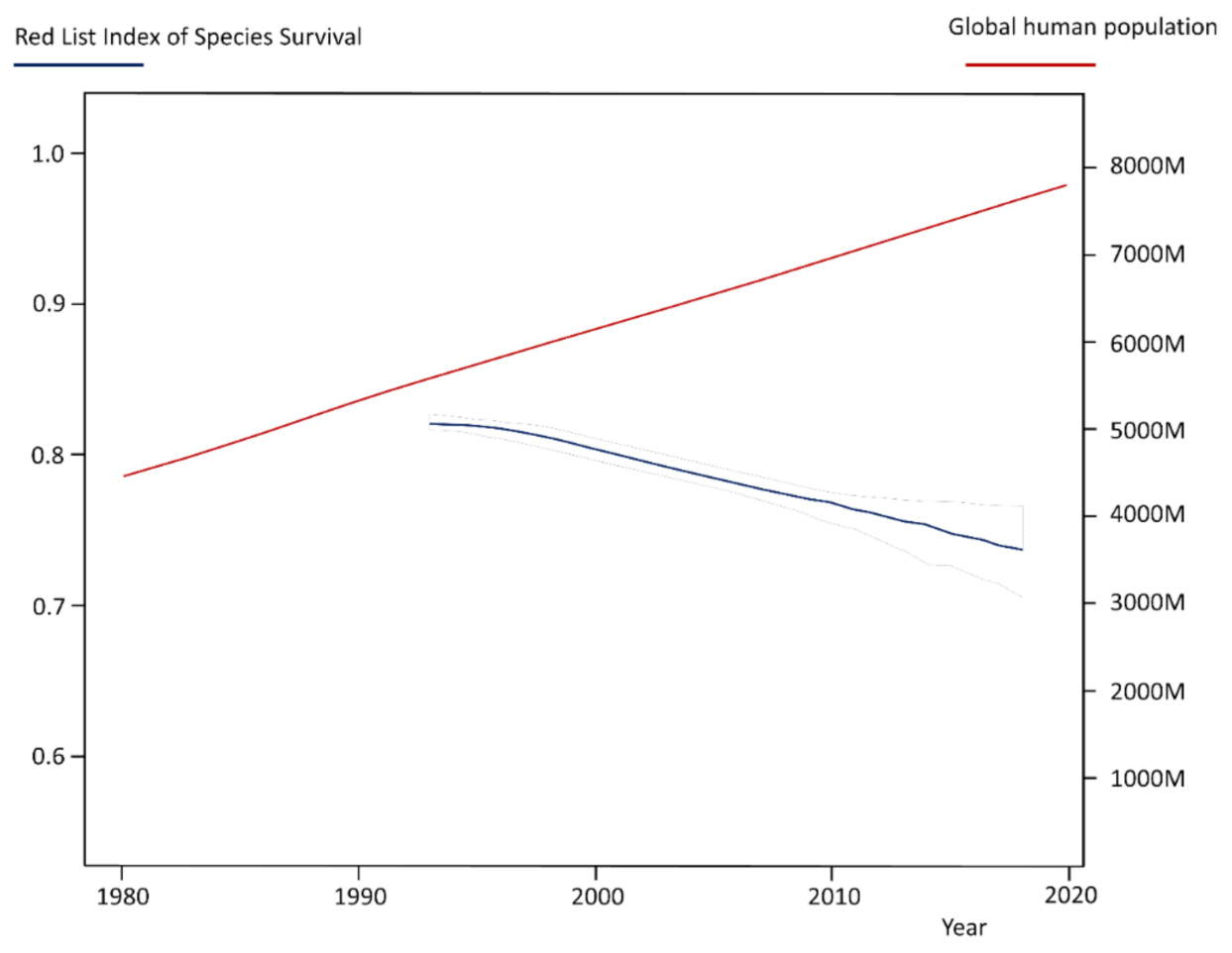
The relationships between humans and wildlife are varied, complex, and dynamic, and an urgent issue of our time. On the one hand the growing human population and the associated increasing demand for a limited and unevenly distributed pool of natural resources [1] is the main driver of biodiversity loss and environmental degradation (Figure 1) [2,3,4]. On the other hand, societies progressively reflect about the fragility of ecosystems (e.g., biodiversity crisis) and—as a result of these considerations—we are experiencing an increasing environmental awareness and appreciation [5], which is linked with efforts to better protect biodiversity and a more sustainable use of natural resources (e.g., transboundary protected area networks) [6,7]. However, crucial to this dilemma is that both developments have the potential to create new or intensify existing conflicts between humans and wildlife [6,8].
Consequently, the topic calls for multi-perspective and transdisciplinary approaches, including ecological, socio-ecological, socio-economic, and socio-cultural research concepts. Viewing existing conflicts through a local lens and in a case-specific context is important, but it does not always comply with the complexity of the underlying phenomenon. This is especially true when it comes to transregionally or globally occurring forms of conflict. Against this background, it seems advisable and necessary to exemplify the various types of existing conflicts in different contexts, provide inspiration for cross-regional and cross-landscape approaches, as well as encouraging the discovery and better understanding of fundamentally valid mechanisms and generally applicable strategies.
In a widely cited review article, Francine Madden clearly outlined the underlying dilemma: “Two interesting contradictory misconceptions seem to exist simultaneously. On one hand, many conservationists believe that “their” species or protected area is unique such that “lessons learned” from elsewhere or widely held principles of how to assess, address, process, and evaluate conflict do not apply.” “On the other hand, it is a mistake to think that a one-size-fits-all, standardized prescription for mitigation can be applied successfully across the wide spectrum of specific conflict situations.” [9] (p. 251).
The value of generally applicable mechanisms arises from the need to develop clear recommendations for action. However, this should not lead to oversimplification of complex issues through generalization. At the same time, and in view of the urgency of the issue, overestimation of case specificity at the local scale should be cautioned.
Here, we try to approach the broad topic of cross-landscape considerations of human–wildlife conflict (HWC) and outline that a cross-landscape view is possible, if the level of abstraction is adjusted accordingly and a balanced weighing of similarities and differences is conducted. In a first step, we try to clarify the terminology and in particular the necessity to address the concept of “conflict” from the perspective of selected wildlife species. In a second step, we encourage longitudinal investigations that may even cross the borders of zoogeographical regions and thus enable HWC to be compared on a supra-regional scale. We then present two case scenarios that exemplarily illustrate the topic from different perspectives (classical view, human perspective vs. wildlife perspective, rarely regarded as a form of HWC). In view of the challenges of our time, it seems necessary to work out and uncover these globally effective mechanisms as far as possible.

2. Trying to Adopt a Wildlife Species’ Perspective
Human–wildlife conflict (HWC) can be regarded as a phenomenon that involves and impacts on both, humans and wildlife. While many definitions explicitly address the interdependence of the two entities (humans vs. wildlife) [9,12,13,14], others stress negative impacts on humans caused by wildlife [15,16]. There are also those for whom any instances in which human and nonhuman interests overlap and clash can be seen as HWC, although this view completely disregards the concept of wildlife [17]. However, interestingly, much of the work published on the subject rather disproportionately sees wildlife causing a negative impact on the goals of humans, sometimes complemented by the statement that conflicts can be regarded as human–human conflicts or conflict between conservation and other human interests [18,19]. Social sciences undoubtedly contributed to a better understanding of the so-called “human dimension of wildlife” that can be approached from various directions, including a temporal, cultural and political dimension [17,18]. Characteristic of the anthropocentric nature of such considerations is the debate as to whether the term “coexistence” is more appropriate than the term “conflict”, because “coexistence” has a more positive connotation [20,21]. In their review paper “Rearticulating the myth of human wildlife conflict”, Peterson and others argue that the concept of HWC was used to frame high-profile conservation cases and that the concept initially was rooted in the science of wildlife damage management [16].
However, we think that we should use a more symmetrical view on HWC by also looking at cases where human activities impact non-human species. Thus, let us approach the issue from the perspective of selected wildlife species. A result of a particularly obvious form of conflict is the extinction of wildlife species following human activity, e.g., blue antelope (Hippotragus leucophaeus) [22], Tasmanian tiger (Thylacinus cynocephalus) [23], passenger pigeon (Ectopistes migratorius) [24], dodo (Raphus cucullatus) [25], moas (Dinornithiformes) [26] or giant tortoises (Cylindraspis spp.) [25]. However, these human-caused extinctions of wildlife species are termed “modern era extinctions” or “extinctions in the Anthropocene” [25] but are rarely subsumed under the umbrella of HWC.
A successful strategy of preventing these extinction events can be the re-introduction of remaining individuals into the natural habitat, if this still exists. Reintroduction is “the intentional movement and release of an organism inside its indigenous range from which it has disappeared” in order to re-establish “a viable population of the focal species within its indigenous range” [27] (p. 3). Table 1 lists examples of selected avian and mammalian species that have been pushed to the brink of extinction as a result of human activity—often overexploitation of natural resources. The remaining “populations” of some of these animal species were little more than a handful of individuals. Reintroduction is a globally applicable tool/procedure to counteract one of the most extreme forms of HWC. Reintroduction can be seen as a key-measure in preventing the extinction of these species at the last moment. The example of this reintroduction of endangered species is a lesson in the value of taking action when a conflict has progressed to the point of having an almost irretrievable impact: in this case, the disappearance of a charismatic wildlife species from the surface of the earth. At the same time, the example also shows that extremely time-consuming and labor-intensive tasks are required, and that success is by no means guaranteed. While this strategy is globally applicable, of course, there are region-specific, but especially species-specific differences that need to be considered.
Our examples from Table 1 are particularly suitable for showing case specificity and global relevance. Many of the animal species that are listed there and that came to the brink of extinction due to human intervention showed a spatially limited natural distribution range. It seems obvious and understandable that such species are particularly endangered by extinction events. The strategies and measures differ greatly among the species and according to their ecological conditions and biological requirements. In the case of the kakapo (Strigops habroptila), the animal species was completely removed from the original natural distribution area (New Zealand), the remaining animals were kept in captivity and then the offspring were released into offshore islands after alien predators (representatives of the order Carnivora) had been completely eradicated there—a very specific procedure that can hardly be transferred to other cases. In the case of the Przewalski’s horse (Equus ferus przewalskii), the remaining population existed exclusively ex situ for decades before wildlife conservationists could start to slowly reintroduce horses in their original habitat. For example, captive-born golden lion tamarins (Leontopithecus rosalia) or black-footed ferrets (Mustela nigripes) cannot simply be released into the wild but need to be trained pre-release in order to learn to find food (catch prey) and avoid dangers [28]. A zoo-born white rhino (Ceratotherium simum), on the other hand, was successfully reintroduced into a national park in Namibia without any behavioral training pre-release [29]. All of this shows the highly species-specific and case-specific dimension behind reintroduction. Yet, there is also a global dimension behind it. On one hand, the case studies show that the phenomenon exists all over the world. The procedure is similar: populations that have shrunk to little more than a handful of individuals are saved from certain extinction by protecting them with great effort and mostly reintroducing them into parts of the natural distribution range. An interesting and transregional peculiarity is the willingness of humans to undertake sometimes considerable efforts when the state of near extinction is reached. Sometimes, extreme effort is made to avert the extinction event. It could probably have been saved earlier in the process with much less effort.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Overview of selected bird and mammal species that were at the brink of extinction owing to human exploitation, populations were reduced to very few individuals that formed the basis for (more or less successful) reintroduction or translocation programs. These examples can be useful to adopt a more symmetrical view on HWC, while also illustrating global relevancy of the underlying phenomenon (resource-overexploitation by humans) and global applicability of mitigation strategies (reintroduction and translocation). Superscript digits refer to references.
Table 1.
Overview of selected bird and mammal species that were at the brink of extinction owing to human exploitation, populations were reduced to very few individuals that formed the basis for (more or less successful) reintroduction or translocation programs. These examples can be useful to adopt a more symmetrical view on HWC, while also illustrating global relevancy of the underlying phenomenon (resource-overexploitation by humans) and global applicability of mitigation strategies (reintroduction and translocation). Superscript digits refer to references.
| Scientific Name | Extinct in the Wild | Population Bottleneck | Nucleus of Initial Reintroduction Program (Founders) | Initial Release into Wild | Area of Initial Release | Current In Situ Population [30] |
|---|---|---|---|---|---|---|
| Strigops habroptila | 1997 [31] | 62 animals, 1999 [31] | 61 animals, 1980–1991 [31] | - | Offshore islands of New Zealand [31] | 116 mature individuals |
| Leucopsar rothschildi | - | 6 animals in the wild in 2001 + unknown number in captivity [32] | 426 animals, 1998–2019 [32] | around 1983 [32] | Bali Barat National Park [32] | 1–49 mature individuals |
| Gymnogyps californianus | 1987 [33] | 22 animals, 1981 [33] | 85 animals, 1982–2003 [33] | 1992 [34] | Sespe Condor Sanctuary, Los Padres [34] | 93 mature individuals |
| Canis rufus | 1980 [35] | 14 animals, 1976 [35] | 102 animals, 1987–2002 [35] | 1987 [35] | Alligator River National Wildlife Refuge, Carolina [35] | 20–30 mature individuals |
| Mustela nigripes | 1980s [36] | 18 animals, 1985–87 [36] | 37 animals, 1991 [37] | From 1991 [36] | Shirley Basin, Wyoming [36] | 206 mature individuals |
| Dama mesopotamica | - | 25 animals, 1956 [38] | Iran: 6 animals, 1964–1965 [39] Israel: 7 animals, 1976 [38] | From 1977 [39] 1996 [38] | Different sites in Iran [39] Nahal Kziv Nature Reserve [38,40] | >250 adults |
| Elaphurus davidianus | >1500 years ago [41] | 11 animals, 1900–1910 [41] | 77 animals, 1985–1987 [41] | 1987 [41] | Dafeng Milu Nature Reserve [41] | ~600 individuals |
| Oryx leucoryx | 1972 [42] | 14 animals, 1962–67 (“World Herd”) [43] +few animals from private collections [44] | Oman: 40 animals, 1980–1995 [45] | 1982 [45] | Arabian Oryx Sanctuary [45] | 850 mature individuals |
| Jordan: 14 animals, 1978–84 [46] | 1983 [46] | Shaumari Nature Reserve [46] | ||||
| Israel: 7 animals, 1978 [44] | 1997–1998 [44] | Shahak Spring [44] | ||||
| Saudi Arabia: 35 animals, 1989 [47] | 1990–1993 [48] 1995–2003 [48] | Mahazat as-Sayd [48] Uruq Bani Ma’arid [48] | ||||
| UAE: 90 animals [49] | 2007 [49] | Um El Zumool [49] | ||||
| Bison bonasus | 1927 [50] | 54 animals, 1920s [51] | 18 animals [50] | 1952–1966 [50] | Puszcza Bialowieska [50] | 2518 mature individuals |
| Equus ferus przewalskii | 1960s [52] | 53 animals, 1920s [53] | 16 animals, 1900–1947 [53,54] | 1992–2004 [53] 1992–2000 [53] | Great Gobi B via Takhin Tal [54] Hustai National Park [53] | 178 mature individuals |
| Ceratotherium s. simum | - | 20–50 animals [55] ~200 animals [56], 1900 | South Africa: 330 animals [57] | 1961–67 [57] | Different reserves in South Africa [57] | 10,080 mature individuals |
3. Comparing Ecosystems via Longitudinal Studies—Even beyond the Borders of Biogeographical Regions
Since Alfred Russel Wallace (1823–1913) divided the world into six zoogeographical regions in his work “The Geographical Distribution of Animals” [58], cross-landscape considerations of ecological relationships appear permitted and meaningful if they relate to habitats located within one zoogeographical region. For example, it seems legitimate and obvious to consider faunal elements and the conflicts they may be associated with throughout their distribution range, e.g., in terms of longitudinal studies [59]. However, when such a comparison relates to areas within different zoogeographical regions, things get much more complicated. A basic assumption behind biogeographical thinking is that a different evolutionary history of ecosystems within different biogeographic regions and the associated distinct evolutionary scenarios imply a separate consideration. Strictly speaking, this approach would lead to the realization that cross-landscape considerations of HWC are only possible if conflicts are addressed within one-and-the-same zoogeographical region. However, would this approach do justice to the complexity of the topic?
There are, on the other hand, generally applicable and globally valid ecological rules, and concepts, as becomes clear from the example of ecological equivalence in the sense of similar ecological interrelationships involving convergent species in different regions. Moreover, it can also happen that certain geographical regions show parallels and similarities that only become apparent at second glance, e.g., when the time scale is extended. Central Eurasia and southern Africa provide a good example in this regard. Although these regions appear to have little in common at first glance, there are several striking natural similarities [60,61,62]. Natural links and parallels between these regions include: (i) Tertiary tectonic events and shifting in the African–Eurasian plate boundary zone [63], (ii) Tertiary similarities in vegetation—a Mediterranean-type woodland thorn scrub [64], (iii) Quaternary Western European complex of Mediterranean/African affinities with North–South and South–North fluctuations of faunistic elements [65], and (iv) the Palearctic–African bird migration system, the largest bird migration system in the world [66].
During the last interglacial (Eemian interglacial, 126–115 ky BP), western Eurasia (today central Europe) was home to large mammal communities very similar to those of modern-day Africa. There were species that we can still find in Africa today (e.g., Crocuta crocuta, Panthera leo, Hippopotamus amphibius) or those that were relatively closely related to recent African species and may have fulfilled a similar ecological function in the Eurasian context (e.g., Palaeoloxodon antiquus) [62]. Having in mind that the large mammal communities of central Europe were significantly reduced by human activity, we come to the realization that sub-Saharan Africa represents an important model region for understanding the interactions between ecosystems and the associated large mammal communities. This example of comparing the situation in central Europe and southern Africa illustrates the legitimacy of comparing ecological contexts (such as HWC) beyond the borders of zoogeographical regions. Rather, the decisive factor here is the shift in the time scale and the awareness of a suitable reference point in time.
Such comparative considerations of the interrelations between human activity and biodiversity, including HWC, lead to an awareness of the transregional or even globally valid dimension of this phenomenon. From our point of view, two aspects are particularly important: Firstly, knowing about the natural connections between different and distinct appearing regions leads to an appreciation of the legitimacy of applying similar strategies in different contexts. The inclusion of time as an important dimension shows that relations within ecosystems can be more similar than it might first appear and reveals parallels and natural connections that justify transregional approaches.
Secondly, raising awareness of the similarities among ecosystems in different geographical regions can show the extent of the degree of ecosystem-transformation resulting from human activities. Reflecting about the existence of megafauna communities in central Europe during the Eemian interglacial gives us an idea of how highly transformed the current situation actually is. This in turn can lead to a reassessment of the concept of conflict. Observations that we initially evaluate as a conflict may be an expression of a high-level disruption of a formerly near-natural system. The reference value is decisive for the assessment of a supposed conflict situation. Considering a cleared agricultural landscape as a reference, into which large predatory mammals suddenly migrate, harbors an enormous potential for conflict per se. If, on the other hand, one realizes that this very landscape was characterized by the existence of megafauna communities including various very large representatives of carnivores, the recolonization of individual carnivore species appears in a completely different light. It is important to recognize that the development of strategies to avoid conflict depends heavily on what is perceived as conflict and what is defined as a desirable state. Such considerations can help us to shift from an anthropocentric perspective towards a more ecocentric view. This sovereignty of interpretation of this near-natural state is a very crucial point when considering HWC. In view of the fact that there are numerous initiatives that are active in nature conservation and that aim to mitigate or minimize existing conflicts, it is important that these initiatives have a valid and realistic idea of near-natural conditions. Here, is a very important lever to improve measures to avoid HWC by defining the states worth protecting as scientifically as possible. We think that cross-regional comparisons of ecological interrelations, including comparisons of HWC in different regional contexts and even on different continents can play an important role in this regard.
4. Carnivore–Livestock Conflicts—A Classic Example for Transregional Considerations
Carnivores (Carnivora) are monophyletic, globally distributed, highly diverse in form and function, and share similar ecological adaptations associated with their ecological function as predators [67]. Large carnivores require vast areas to establish home ranges and territories and are typical top predators of ungulates [67,68,69]. In the course of an anthropogenically controlled process (domestication), humans have transformed little more than a handful of ungulate species into domesticated forms (livestock) [70]. Some of these domesticated forms, whose natural “stem species” are either extinct (Bos primigenius) or occur in spatially clearly defined areas (Ovis orientalis, Capra aegagrus), are now spread globally as a result of this cultural achievement of humans and the number of individuals exceeds that of their stem species many times over [71]. However, despite their phenotypic diversity and adaptation to diverse regions, these domesticated livestock forms represent very few biological species. Domesticated ungulates often displace naturally occurring ungulate species and become prey for large carnivores. Owing to the cultural component of the domestication process, carnivore–livestock conflict undoubtedly represents a special form of HWC. Owing to human population growth, increasing livestock numbers, and intensifying agricultural practices on the one hand and ecotourism and an increasing awareness of the importance of nature conservation on the other hand, carnivore–livestock conflict is a global phenomenon. Comparative approaches seem effective to better understand and mitigate such conflicts on a transnational or even global scale. Thus, a comparative framework is needed that allows us to differentiate between case specificity and common applicability of the underlying ecological processes and the resulting management strategies in different regions.
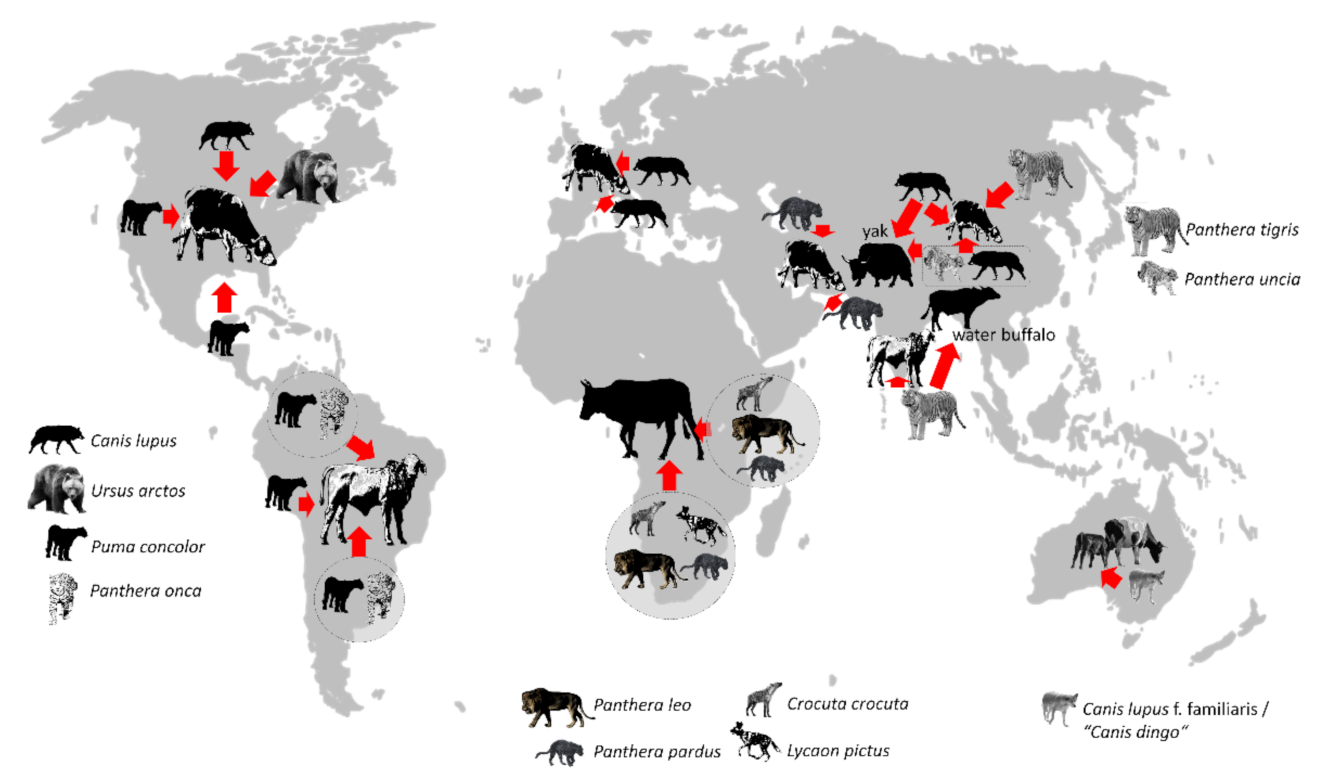
The global phenomenon of carnivore–cattle conflicts (Figure 2) is a good illustration of the principle according to which general aspects and case-specific characteristics can be weighed up if different levels are considered. An obvious global similarity is that large representatives of a systematic group (carnivores), as well as domesticated forms of a today-extinct wild species (cattle), exist all over the world. As a result of a growing human population and an increase in livestock-related biomass [60], humans are simulating an increasing abundance of a large ungulate species, which has also been modified by humans and is often more readily available as prey for large carnivores than naturally occurring ungulate species.
From a biological point of view, we have a globally comparable phenomenon on an abstract level, which, to some extent, enables the derivation of globally transferrable management strategies and conservation measures. However, a closer look reveals pronounced differences in the life history strategies of the carnivore species, as it makes a difference whether this species is a representative of the families Ursidae, Canidae, Felidae, or Hyaenidae to stick with our selected examples from Figure 2. Ursus arctos, for example, can induce conflicts resulting from depredation of cattle, but owing to its omnivorous feeding habits, the species can also be seen as a potential crop raider [72,73]. It also makes a difference for the development of management strategies whether the conflict is induced by canids that hunt in packs, such as wild dogs or wolves, or by felids that mostly live and hunt solitary and hidden, such as puma, jaguar, or leopard. The habitat also plays an important role. This becomes clear with the example of the leopard, which occurs in very different habitats. Additionally, it seems obvious that management strategies need to be adjusted depending on whether the animal is hunting in open areas or dense rainforest. This shows that besides considering phylogeny and habitat, it can be advisable to address a certain degree of case specificity even in view of one and the same species. This can also be observed by looking at the wolf (Canis lupus). The wolf is a particularly interesting case study because the species has a vast range that includes much of the entire Holarctic. Additionally, the domesticated and then feralized form of the wolf, the dingo, is the apex predator on the Australian continent. In central Europe, wolves are recolonizing areas, where the species was formerly extinct [74]. The emergence of this top predator in parts of Europe forces wildlife managers to develop appropriate strategies. To better understand the recovery of the wolf in Europe, it is important to investigate regions, where the same species causes comparable conflicts. Given the large distribution range of Canis lupus, comparable investigations on similar longitudinal levels, including the entire Holarctic region, seem feasible. However, it should also be taken into account that such a wide distribution area is also a consequence of the extreme ecological plasticity of this species, i.e., its ability to cope with different environmental conditions (euryoecious). This ecological plasticity also implies a high level of responsiveness to management strategies designed to avoid wolf–cattle conflicts. In other words, even though a conflict is of the same nature in different areas, it may be necessary to develop case-specific management strategies. Alternatively, to put it another way, there is no species-specific avoidance tactic to prevent a certain type of conflict in different regions or even under different circumstances (individual variability).
Figure 2.
Selected examples of carnivore–cattle conflicts on a global scale—if not further specified, ungulate silhouettes refer to different forms of cattle derived from the aurox Bos primigenius. All animal silhouettes derived from photos: T. Göttert. References: Canis lupus, North America: [75,76]; Ursus arctos: [77]; Puma concolor, western North America: [78]; Puma concolor, central America: [78,79]; circle, northern South America: [80]; Puma concolor, western South America: [78,81,82]; circle, southern South America: [81,82]; circle, eastern Africa: [83,84]; circle, southern Africa: [85]; Canis lupus, southern Europe: [86]; Canis lupus, eastern Europe: [87,88]; Canis lupus, central Asia: [89,90]; Panthera pardus, Near East: [91]; Panthera pardus, South Asia: [92]; box: Panthera uncia and Canis lupus: [93,94]; Panthera tigris, South Asia: [95,96]; Panthera tigris, East Asia: [97,98]; Canis lupus f. familiaris: [99].
Figure 2.
Selected examples of carnivore–cattle conflicts on a global scale—if not further specified, ungulate silhouettes refer to different forms of cattle derived from the aurox Bos primigenius. All animal silhouettes derived from photos: T. Göttert. References: Canis lupus, North America: [75,76]; Ursus arctos: [77]; Puma concolor, western North America: [78]; Puma concolor, central America: [78,79]; circle, northern South America: [80]; Puma concolor, western South America: [78,81,82]; circle, southern South America: [81,82]; circle, eastern Africa: [83,84]; circle, southern Africa: [85]; Canis lupus, southern Europe: [86]; Canis lupus, eastern Europe: [87,88]; Canis lupus, central Asia: [89,90]; Panthera pardus, Near East: [91]; Panthera pardus, South Asia: [92]; box: Panthera uncia and Canis lupus: [93,94]; Panthera tigris, South Asia: [95,96]; Panthera tigris, East Asia: [97,98]; Canis lupus f. familiaris: [99].

The experience from regions, where other carnivore species cause conflicts with livestock, is also part of the comparative framework. In Africa, for example, pastoralists coexist with large predators for millennia [60]. This knowledge might contribute to a common understanding of carnivore–livestock conflicts and mitigation strategies (Figure 3). Besides distinguishing between case-specific and species-specific patterns, the detection of generally applicable practices is needed to achieve the coexistence of ecologically highly relevant faunal elements, as well as human well-being. In other words, appropriately dealing with the topic requires a differentiation between similarities and case specificity. Similar aspects are: global drivers of conflicts, global distribution of carnivores (and livestock), similar ecological functions of large carnivores, similar spatial and dietary requirements of large carnivores. Case-specific aspects can be related to different social, cultural and economic conditions applying to the countries/regions, where the conflict emerges (e.g., developed countries vs. developing countries). However, they can also be the result of different behavioral patterns of representatives of different members of the systematic group Carnivora (e.g., a hunting pack of wolves vs. a solitary hunting large felid). Additionally, these aspects can be linked with different historic or political backgrounds of countries or regions that may exclude certain strategies to avoid or mitigate carnivore–livestock conflicts (e.g., re-settlement of people from national parks to adjacent areas seems problematic in former Apartheid countries). Once this appreciation of similarities and differences has been conducted, there is a globally effective “toolbox” [9] available for wildlife managers and livestock keepers including: using/optimizing artificial and natural barriers, guarding/optimizing livestock husbandry practices, effective waste management systems, compensation or insurance schemes and lethal control of carnivores in the last instance.
5. A Novel and Rapidly Emerging, Global Human–Wildlife Conflict Resulting from Wind Energy Development
The above-described example (carnivore livestock conflicts) is classic because it is a millennia-old phenomenon, and it causes direct harm to humans (direct threat to livelihoods and survival). It exemplifies all the forms of conflict that are evident when viewed from the human perspective. Numerous other examples of conflicts could be added here, e.g., damage to crops caused by wildlife or impairment (up to and including devastation) of infrastructure by wildlife. These are generally examples that are often cited when reflecting on the HWC theme. The starting point for the following topic (bats vs. wind energy) is different. Firstly, because a relatively new form of land use is involved. In addition, the conflict is clearly directed towards wildlife, which is probably one reason why the topic has so far hardly been subsumed under the umbrella of HWC. Since the conflict is originated by a human-induced threat (wind energy), the phenomenon can be viewed as one form of threat–target relationship belonging to the disciplinary arena of threat analysis [100]. In the following, we will look at it as an example for various forms of conflicts that come to light when viewed from the perspective of wildlife. Due to the complexity of the interrelationships between human action and ecosystem stability and integrity, one-sided considerations (human vs. wildlife perspective) do not do justice to the topic.
The worldwide expansion of the wind energy sector makes an important contribution to environmental protection with regard to avoiding CO2 emissions when compared to the use of fossil fuels. On the other hand, the construction and operation of wind turbines interferes with nature and the landscape and can threaten wildlife directly (e.g., collision mortality) and indirectly (e.g., displacement) [101,102,103,104]. Bat fatalities at wind energy facilities were first reported from Australia >40 years ago [105]. In Europe and the United States, observations of bat carcasses beneath wind turbines were reported in the late 1990s [106]. However, while impacts of wind energy on the avifauna have been intensively discussed for many years, public awareness on the impacts on bats was raised only at the end of the twentieth century, with increasing observations of dead bats at wind farms [107,108,109,110]. It has since been noted that the number of bat fatalities often exceed those of birds, e.g., in [101,103,111]. Bats are killed at wind turbines worldwide across different landscapes, and in fact, with regard to mortality, wind turbines have been identified as one of the greatest anthropogenic threats for bats worldwide [109,112,113]. Thus, hardly any other form of renewable energy is subject to as much global controversy as wind energy, resulting in a complicated trade-off-situation between environmental and nature conservation concerns [114,115]. Today, most post-construction fatality monitoring studies and related findings stem from North America and Europe. However, energy demand will rise dramatically in South America, Asia and Africa over the next 20 years, which, in the face of climate change mitigation, will certainly lead to the construction of thousands of wind turbines across these continents with definite implications for local bat faunas. For example, first studies of the few turbines installed in Africa to date have already documented bat fatalities [116].
For bats, similarly to birds, a distinction can be made between lethal and nonlethal effects. Bats can die or be injured by collisions with wind turbines [117,118,119,120], either when colliding with the rotors of wind turbines or when being exposed to air turbulences at the rotor blades [121]. Due to large pressure differences, the latter seems to cause a fatal barotrauma [109,122]. As the wind industry continues to expand, cumulative impacts of bat collisions on some species and/or local populations can be expected, causing population declines, particularly in species with a high collision risk [123,124,125]. Brinkmann et al. have shown that even a 0.5% increase in mortality rates can have negative effects on local populations of individual species [126]. Non-lethal impacts include (i) habitat loss due to structural alterations [127], e.g., when forests are cut down to establish roads and platforms for the erection and operation of wind turbines [112,128], (ii) barrier effects of the facility(ies) within important flight routes, (iii) deterrence effects due to construction work, noise during operation [129], or the constant movement of the rotors, or (iiii) disturbance of ultrasonic orientation due to facility-related emissions in the frequency range of bat vocalizations. The sum of these disturbances could severely affect species by decreasing their habitats’ availability which can cause behavioral changes (e.g., roost abandoning), significantly weaken local populations in the long term [130,131].
Several non-mutually exclusive hypotheses exist to explain the high collision risk of bats across continents [132,133,134]. The feeding-attraction hypothesis states that bats are attracted to the turbine nacelle because of relatively high insect abundances [135] and fatal collisions may occur while bats are foraging at high altitudes in the rotor area or inspecting the turbines for roosting possibilities. Furthermore, opening up forests as sites for wind turbines seems to considerably increase the potential for collisions as significantly more bat carcasses were found under wind turbines in forests than in the open country [126]. Worldwide, forests function as habitats for wildlife with a high ecological value, and several studies have shown that even monotonous coniferous forests can host rich and abundant bat communities [136,137,138,139]. For Northern Europe and North America there seem to exist similar patterns: clear correlations have been observed between weather variables and turbine-related bat activity: bats are particularly active during warm nights with low wind speed. Additionally, temporal patterns of bat fatalities seem to be consistent in the northern hemisphere. Most fatalities occur in late summer and autumn during autumn migration, and (in some parts of Europe) during spring migration. It is assumed that while migrating, bats use other mechanisms for orientation than echolocation, e.g., landscape structures, geomagnetism. There is a comparatively high number of affected individuals of migratory species, leading to potential impacts on supra-regional populations and thus, on bat populations over large geographical areas, many times over-exceeding the used for wind energy production itself [140]. This fact makes the conflict a cross-boundary/cross-landscape human–wildlife conflict. However, while in North America and Northern Europe tree-roosting migratory species seem mostly affected, Arnett et al. suggest that wind turbines are equally dangerous to resident cave bats in South or Central America (assumed to be non-migratory) as to migratory tree-roosting species [112]. Thus, the unifying finding across continents is that the species most vulnerable to collisions are adapted to flight at high altitudes and echolocation in the open air (aerial-hawking and relatively fast flying open air species), irrespective of continent, habitat, migratory pattern or roosting preferences.
Given current knowledge, the impacts of wind energy development seem predictable. This must be an incentive to plan wind turbines particularly carefully. The most important prerequisite for avoiding conflicts seems careful site selection [141]. Relating fatalities to features within the immediate area of a turbine (e.g., proximity to water or forest edge) will help with designing future facilities and locating turbines to avoid higher risk areas within a site (112,135). Proximity to bat-relevant areas such as roosting, hunting, and foraging habitats must also be considered in the selection process. Moreover, within nature conservation research avoidance measures are to be developed and tested that reduce conflicts between wind energy and the conservation of bats and possible impacts on the bat fauna may be addressed in conditions attached to construction permits. In this context, operational adjustments, i.e., technical systems on wind turbines that should prevent collisions with bats as far as possible, in combination with multi-year follow-up monitoring are a proven tool [142]. Proposed mitigation methods include shutdown algorithms, i.e., programming the wind turbine to intervene in operations by shutting it down during high-risk periods, and increasing cut-in speed (wind speed at which wind turbines begin to produce electrical power) have been established to reduce the probability of bat fatalities [143,144]. Additionally, increasing the start-up speed (wind speed at which the rotors begin to spin) as a relevant parameter has been discussed. Such adaptations during turbine operation are referred to as “adaptive management” and are seen as a possibility to accelerate wind energy development while at the same time taking nature conservation concerns into account [145]. These technical innovations could be further pursued, e.g., in combination with intelligent detectors (cameras, radar systems) that observe and measure bat species and activity around the airspace around a turbine and automatically shut down the wind turbine temporarily [146]. Yet, the detection of species and the reaction of the turbine to it must be reliable, and until now, current technical measures do not yet achieve the desired effects [147]. Another approach, the acoustic bat deterrent system, uses ultrasound to deter bats from entering the zone swept by the rotor of a wind turbine [148,149].
However, until now there is no international, legally binding standard in the licensing of wind turbines or the implementation of mitigation measures. Even on a national level, there is a lack of guidelines, standardized approval procedures and the legal framework to ensure effective implementation of bat protection measures across the countries [150]. Finally, it needs to be considered, that, irrespective of legislations, climate protection, or biodiversity conservation goals, competing interests, values and views of people will likely continue to influence the future development of this HWC [115,151].
6. Conclusions
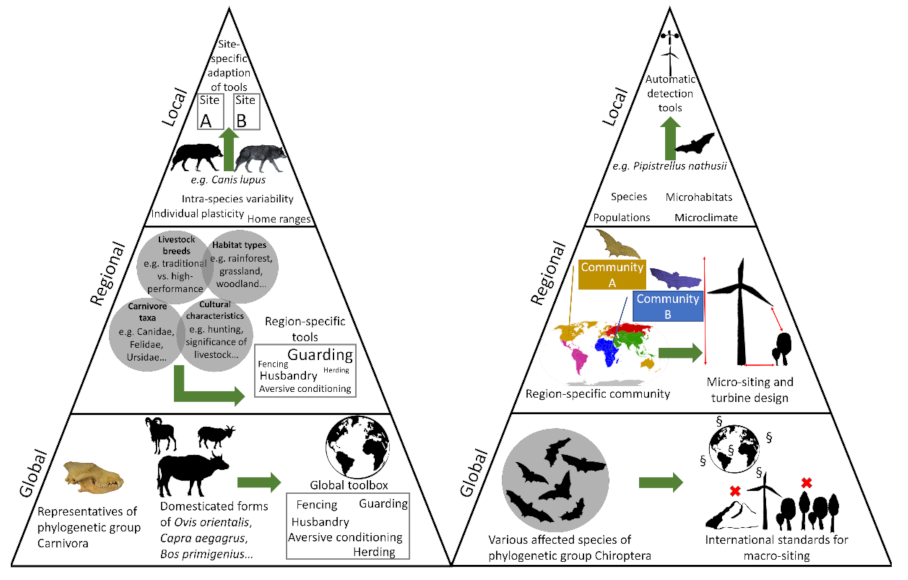
Our paper aims at stimulating reflection on the extent to which human–wildlife conflicts (HWC) can be addressed by global strategies, or to what extent the conflicts are region- or case-specific and therefore require an independent and special approach. In order to deal with this topic, we have selected two examples that are exemplary for different perspectives. On one hand, there is the classic perspective from a human’s point of view, whose livelihood is under threat because expressions of artificial biodiversity (domesticated animals) are threatened by natural faunal elements (carnivores). Another perspective is described by wind turbine-induced mortality of bats, an example that is rarely regarded as a form of HWC. We selected these two examples (carnivores vs. domesticated ungulate species and bats vs. wind farms) not only because they represent different viewpoints from which HWC can be looked at. Moreover, each example involves a clearly definably wildlife entity (monophyla Carnivora as well as Chiroptera), which, to our opinion, helps to develop an approach on different spatial scales (see below). It turns out that despite very different perspectives and types of conflict, a comparable development plan of strategies can very well be developed that focuses on different spatial (global, regional, local) and systematic scales (order, family, population, species, individuals) (Figure 4).
In the first step (recording and depicting the global dimension of the conflict) it is important to work out a phylogenetic entity that is represented globally (here: orders Carnivora and Chiroptera). Both groups represent monophyla, communities of individuals descending from a common ancestor, thus sharing a set of common ecological characteristics. This entitles us to look at the conflict from a supra-regional perspective. If we can define a phylogenetically related (and ecologically similar) community with global distribution, it legitimizes the approach at the global level. Of course, the ecological plasticity of the taxa representing a monophylum needs to be adequately considered, as phylogenetically closely-related species can differ in important ways. At the same time, functionally similar but phylogenetically dissimilar taxa (e.g., a mesocarnivore and a carnivorous marsupial, or a bat species and a bird species) may offer useful and comparable insights. Then, in the following steps (regional and local level), the principle is applied that we refine and adjust the approach according to site-, species- and sometimes even population-specific conditions. In the case of carnivore–livestock conflicts, the solution is the global toolbox of conflict mitigation measures, and in the case of bats, the solution is the adequate and meaningful construction of wind turbines with the aim of minimizing conflicts. These two basic approaches can be stepwise refined from the global level to the regional level to the local level and adapted to the specifics of the case. So, when considering generality or case specificity, our approach is characterized by the fact that a biological commonality of a wildlife category exists. This natural relationship among all representatives of a systematic group subsumed under the term “wildlife” is key for the development of an overarching strategy.
Author Contributions
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
No datasets were generated during the current study.
Acknowledgments
We acknowledge three anonymous reviewers for the positive feedback and the constructive and straightforward comments helping us to increase the quality of this paper. Moreover, we express our sincere gratitude to Dipl. Ing. agr. Christel Simantke for providing the excellent photo of the shepherd in Germany (Figure 3, top).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Steffen, W.; Richardson, K.; Rockström, J.; Cornell, S.E.; Fetzer, I.; Bennett, E.M.; Biggs, R.; Carpenter, S.R.; de Vries, W.; de Wit, C.A.; et al. Planetary boundaries: Guiding human development on a changing planet. Science 2015, 347, 1259855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhary, A.; Kastner, T. Land use biodiversity impacts embodied in international food trade. Glob. Environ. Chang. 2016, 38, 195–204. [Google Scholar] [CrossRef]
- Chaudhary, A.; Pfister, S.; Hellweg, S. Spatially explicit analysis of biodiversity loss due to global agriculture, pasture and forest land use from a producer and consumer perspective. Environ. Sci. Technol. 2016, 50, 3928–3936. [Google Scholar] [CrossRef] [PubMed]
- Marques, A.; Martins, I.S.; Kastner, T.; Plutzar, C.; Theurl, M.C.; Eisenmenger, N.; Huijbregts, M.A.J.; Wood, R.; Stadler, K.; Bruckner, M.; et al. Increasing impacts of land use on biodiversity and carbon sequestration driven by population and economic growth. Nat. Ecol. Evol. 2019, 3, 628–637. [Google Scholar] [CrossRef] [PubMed]
- Gadenne, D.L.; Kennedy, J.; McKeiver, C. An empirical study of environmental awareness and practices in SMEs. J. Bus. Ethics 2009, 84, 45–63. [Google Scholar] [CrossRef]
- Stoldt, M.; Göttert, T.; Mann, C.; Zeller, U. Transfrontier Conservation Areas and Human-Wildlife Conflict: The Case of the Namibian Component of the Kavango-Zambezi (KAZA) TFCA. Sci. Rep. 2020, 10, 7964. [Google Scholar] [CrossRef]
- Thornton, D.; Branch, L.; Murray, D. Distribution and connectivity of protected areas in the Americas facilitates transboundary conservation. Ecol. Appl. 2020, 30, e02027. [Google Scholar] [CrossRef]
- Distefano, E. Human-Wildlife Conflict Worldwide: Collection of Case Studies, Analysis of Management Strategies and Good Practices; Food and Agricultural Organization of the United Nations (FAO) Sustainable Agriculture; Rural Development Initiative (SARDI): Rome, Italy, 2005. [Google Scholar]
- Madden, F. Creating coexistence between humans and wildlife: Global perspectives on local efforts to address human-wildlife conflict. Hum. Dimens. Wildl. 2004, 9, 247–257. [Google Scholar] [CrossRef]
- Red List Index. Available online: https://www.iucnredlist.org/assessment/red-list-index (accessed on 20 April 2022).
- United Nations—World Population Prospects 2019. Available online: https://population.un.org/wpp/Download/Standard/Population/ (accessed on 20 April 2022).
- Conover, M.R. Resolving Human-Wildlife Conflicts: The Science of Wildlife Damage Management, 1st ed.; Lewis Publishers: Boca Raton, FL, USA, 2001; p. 440. [Google Scholar]
- Woodroffe, R.; Thirgood, S.; Rabinowitz, A. People and Wildlife, Conflict or Co-Existence? Cambridge University Press: New York, NY, USA, 2005. [Google Scholar]
- Nyhus, P.J. Human–wildlife conflict and coexistence. Annu. Rev. Environ. Resour. 2016, 41, 143–171. [Google Scholar] [CrossRef] [Green Version]
- Treves, A.; Karanth, K.U. Human-carnivore conflict and perspectives on carnivore management worldwide. Conserv. Biol. 2003, 17, 1491–1499. [Google Scholar] [CrossRef]
- Peterson, M.N.; Birckhead, J.L.; Leong, K.; Peterson, M.J.; Peterson, T.R. Rearticulating the myth of human–wildlife conflict. Conserv. Lett. 2010, 3, 74–82. [Google Scholar] [CrossRef]
- Fletcher, R.; Toncheva, S. The political economy of human-wildlife conflict and coexistence. Biol. Conserv. 2021, 260, 109216. [Google Scholar] [CrossRef]
- Dickman, A.J. Complexities of conflict: The importance of considering social factors for effectively resolving human–wildlife conflict. Anim. Conserv. 2010, 13, 458–466. [Google Scholar] [CrossRef]
- Redpath, S.M.; Bhatia, S.; Young, J. Tilting at wildlife: Reconsidering human–wildlife conflict. Oryx 2015, 49, 222–225. [Google Scholar] [CrossRef] [Green Version]
- Frank, B.; Glikman, J.A.; Marchini, S. Human–Wildlife Interactions: Turning Conflict into Coexistence; Cambridge University Press: Cambridge, UK, 2019. [Google Scholar] [CrossRef]
- Hodgson, I.D.; Redpath, S.M.; Sandstrom, C.; Biggs, D. The State of Knowledge and Practice on Human-Wildlife Conflicts; Research Report; The Luc Hoffman Institute: Gland, Switzerland, 2020. [Google Scholar]
- Hempel, E.; Bibi, F.; Faith, J.T.; Brink, J.S.; Kalthoff, D.C.; Kamminga, P.; Paijmans, J.L.A.; Westbury, M.V.; Hofreiter, M.; Zachos, F.E. Identifying the true number of specimens of the extinct blue antelope (Hippotragus leucophaeus). Sci. Rep. 2021, 11, 2100. [Google Scholar] [CrossRef]
- Waterhouse, J.; Mitchell, C. Has Anybody Seen a Tasmanian Tiger Lately? Ethical and Ontological Considerations of Thylacine De-Extinction. Green Lett. 2021, 25, 1–14. [Google Scholar] [CrossRef]
- Cuker, B.E. Passenger pigeon and waterfowl: Flights to extinction and not. In Diet for a Sustainable Ecosystem, 1st ed.; Cuker, B.E., Ed.; Springer: Cham, Switzerland, 2020; pp. 155–174. [Google Scholar] [CrossRef]
- Turvey, S.T.; Crees, J.J. Extinction in the Anthropocene. Curr. Biol. 2019, 29, R982–R986. [Google Scholar] [CrossRef]
- Boast, A.P.; Weyrich, L.S.; Wood, J.R.; Metcalf, J.L.; Knight, R.; Cooper, A. Coprolites reveal ecological interactions lost with the extinction of New Zealand birds. Proc. Natl. Acad. Sci. USA 2018, 115, 1546–1551. [Google Scholar] [CrossRef] [Green Version]
- IUCN/SSC. Guidelines for Reintroductions and Other Conservation Translocations; Version 1.0; IUCN Species Survival Commission: Gland, Switzerland, 2013; p. 57. [Google Scholar]
- Kleiman, D.G.; Beck, B.B.; Dietz, J.M.; Dietz, L.A.; Ballou, J.D.; Coimbra-Filho, A.F. Conservation program for the golden lion tamarin: Captive research and management, ecological studies, educational strategies, and reintroduction. In Primates—The Road to Self-Sustaining Populations; Benirschke, K., Ed.; Springer: New York, NY, USA, 1986; pp. 959–979. [Google Scholar]
- Böer, M.; Brain, C.; Cantzler, T.; Hamza, H.; Venzke, K. Reintroduction of a captive born white rhinoceros (Ceratotherium simum simum) to the Etosha National Park. Part II: Behaviour in the boma, procedures before and locomotion during the first ten weeks after release and observations from June until September 1997. Zool. Gart. 1999, 69, 73–81. [Google Scholar]
- The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/ (accessed on 20 April 2022).
- BirdLife International. Strigops habroptila. The IUCN Red List of Threatened Species; International Union for Conservation of Nature: Gland, Switzerland, 2018; p. e.T22685245A129751169. [Google Scholar] [CrossRef]
- BirdLife International. Leucopsar rothschildi. The IUCN Red List of Threatened Species; International Union for Conservation of Nature: Gland, Switzerland, 2021; p. e.T22710912A183006359. [Google Scholar] [CrossRef]
- BirdLife International. Gymnogyps californianus. The IUCN Red List of Threatened Species; International Union for Conservation of Nature: Gland, Switzerland, 2020; p. e.T22697636A181151405. [Google Scholar] [CrossRef]
- Cohn, J.P. Saving the California condor. Bioscience 1999, 49, 864–868. [Google Scholar] [CrossRef] [Green Version]
- Phillips, M. Canis rufus (Errata Version Published in 2020). The IUCN Red List of Threatened Species; International Union for Conservation of Nature: Gland, Switzerland, 2018; p. e.T3747A163509841. [Google Scholar] [CrossRef]
- Belant, J.; Biggins, D.; Garelle, D.; Griebel, R.G.; Hughes, J.P. Mustela nigripes. The IUCN Red List of Threatened Species; International Union for Conservation of Nature: Gland, Switzerland, 2015; p. e.T14020A45200314. [Google Scholar] [CrossRef]
- Biggins, D.E.; Miller, B.J.; Hanebury, L.R.; Powell, R.A. Mortality of Siberian polecats and black-footed ferrets released onto prairie dog colonies. J. Mammal. 2011, 92, 721–731. [Google Scholar] [CrossRef] [Green Version]
- Bar-David, S.; Saltz, D.; Dayan, T.; Shkedy, Y. Using spatially expanding populations as a tool for evaluating landscape planning: The reintroduced Persian fallow deer as a case study. J. Nat. Conserv. 2008, 16, 164–174. [Google Scholar] [CrossRef]
- Werner, N.Y.; Rabiei, A.; Saltz, D.; Daujat, J.; Baker, K. Dama mesopotamica. The IUCN Red List of Threatened Species; International Union for Conservation of Nature: Gland, Switzerland, 2015; p. e.T6232A97672550. [Google Scholar] [CrossRef]
- Saltz, D. A long-term systematic approach to planning reintroductions: The Persian fallow deer and the Arabian oryx in Israel. Anim. Conserv. Forum 1998, 1, 245–252. [Google Scholar] [CrossRef]
- Jiang, Z.; Harris, R.B. Elaphurus davidianus. The IUCN Red List of Threatened Species; International Union for Conservation of Nature: Gland, Switzerland, 2016; p. e.T7121A22159785. [Google Scholar] [CrossRef]
- Henderson, D.S. Were they the last Arabian oryx? Oryx 1974, 12, 347–350. [Google Scholar] [CrossRef]
- Marshall, T.C.; Spalton, J.A. Simultaneous inbreeding and outbreeding depression in reintroduced Arabian oryx. Anim. Conserv. Forum 2000, 3, 241–248. [Google Scholar] [CrossRef]
- Gilad, O.; Grant, W.E.; Saltz, D. Simulated dynamics of Arabian Oryx (Oryx leucoryx) in the Israeli Negev: Effects of migration corridors and post-reintroduction changes in natality on population viability. Ecol. Model. 2008, 210, 169–178. [Google Scholar] [CrossRef]
- Spalton, J.A.; Lawerence, M.W.; Brend, S.A. Arabian oryx reintroduction in Oman: Successes and setbacks. Oryx 1999, 33, 168–175. [Google Scholar] [CrossRef] [Green Version]
- Harding, L.E.; Abu-Eid, O.F.; Hamidan, N.; al Sha’lan, A. Reintroduction of the Arabian oryx Oryx leucoryx in Jordan: War and redemption. Oryx 2007, 41, 478–487. [Google Scholar] [CrossRef] [Green Version]
- Vassart, M.; Granjon, L.; Greth, A. Genetic variability in the Arabian oryx (Oryx leucoryx). Zoo Biol. 1991, 10, 399–408. [Google Scholar] [CrossRef]
- Mésochina, P.; Bedin, E.; Ostrowski, S. Reintroducing antelopes into arid areas: Lessons learnt from the oryx in Saudi Arabia. Comptes Rendus Biol. 2003, 326, 158–165. [Google Scholar] [CrossRef]
- El Alqamy, H.; Senn, H.; Roberts, M.F.; McEwing, R.; Ogden, R. Genetic assessment of the Arabian oryx founder population in the Emirate of Abu Dhabi, UAE: An example of evaluating unmanaged captive stocks for reintroduction. Conserv. Genet. 2012, 13, 79–88. [Google Scholar] [CrossRef]
- Pucek, Z.; Belousova, I.; Krasińska, M.; Krasiński, Z.; Olech, W. Status survey and conservation action plan. European bison. In IUCN/SSC Bison Specialist Group; Pucek, Z., Ed.; IUCN: Gland, Switzerland; Cambridge, UK, 2004; p. 54. [Google Scholar]
- Olech, W. Bison bonasus. The IUCN Red List of Threatened Species; International Union for Conservation of Nature: Gland, Switzerland, 2008; p. e.T2814A9484719. [Google Scholar] [CrossRef]
- Slotta-Bachmayr, L.; Boegel, R.; Kaczensky, P.; Stauffer, C.; Walzer, C. Use of population viability analysis to identify management priorities and success in reintroducing Przewalski’s horses to southwestern Mongolia. J. Wildl. Manag. 2004, 68, 790–798. [Google Scholar] [CrossRef]
- King, S.R.B.; Boyd, L.; Zimmermann, W.; Kendall, B.E. Equus ferus. The IUCN Red List of Threatened Species; International Union for Conservation of Nature: Gland, Switzerland, 2015; p. e.T41763A97204950. [Google Scholar] [CrossRef]
- Bowling, A.T.; Ryder, O.A. Genetic studies of blood markers in Przewalski’s horses. J. Hered. 1987, 78, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Emslie, R. Ceratotherium simum. The IUCN Red List of Threatened Species; International Union for Conservation of Nature: Gland, Switzerland, 2012; p. e.T4185A16980466. [Google Scholar]
- Rookmaaker, K. The alleged population reduction of the southern white rhinoceros (Ceratotherium simum simum) and the successful recovery. Säugetierkd. Mitt. 2000, 45, 55–70. [Google Scholar]
- Player, I. Translocation of white rhinoceros in South Africa. Oryx 1967, 9, 137–150. [Google Scholar] [CrossRef] [Green Version]
- Wallace, A.R. The Geographical Distribution of Animals with a Study of the Relations of Living and Extinct Faunas as Elucidating the Past Changes of the Earth’s Surface; Macmillan and Co.: London, UK, 1876; Volume II, p. 607. [Google Scholar]
- de Silva, S. Need for longitudinal studies of Asian wildlife in the face of crises. Glob. Ecol. Conserv. 2016, 6, 276–285. [Google Scholar] [CrossRef] [Green Version]
- Zeller, U.; Starik, N.; Göttert, T. Biodiversity, land use and ecosystem services—An organismic and comparative approach to different geographical regions. Glob. Ecol. Conserv. 2017, 10, 114–125. [Google Scholar] [CrossRef]
- Rottstock, T.; Göttert, T.; Zeller, U. Relatively undisturbed African savannas-an important reference for assessing wildlife responses to livestock grazing systems in European rangelands. Glob. Ecol. Conserv. 2020, 23, e01124. [Google Scholar] [CrossRef]
- Zeller, U.; Göttert, T. Humans, megafauna and landscape structure–Rock engravings from Namibia encourage a comparative approach to central Europe and southern Africa. Vertebr. Zool. 2021, 71, 631. [Google Scholar] [CrossRef]
- Meulenkamp, J.E.; Sissingh, W. Tertiary palaeogeography and tectonostratigraphic evolution of the Northern and Southern Peri-Tethys platforms and the intermediate domains of the African–Eurasian convergent plate boundary zone. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 196, 209–228. [Google Scholar] [CrossRef]
- Janis, C.M. Tertiary mammal evolution in the context of changing climates, vegetation, and tectonic events. Annu. Rev. Ecol. Evol. Syst. 1993, 24, 467–500. [Google Scholar] [CrossRef]
- Kahlke, H.D. Biostratigraphical correlations (mammals) of the Quaternary continental deposits of Europe and the Far East. Quartärpaläontologie 1986, 6, 83–86. [Google Scholar]
- Newton, I. The Migration Ecology of Birds, 1st ed.; Academic Press: London, UK, 2008. [Google Scholar]
- Eisenberg, J.F. An introduction to the Carnivora. In Carnivore Behavior, Ecology, and Evolution; Gittleman, J.L., Ed.; Springer: Boston, MA, USA, 1989; pp. 1–9. [Google Scholar] [CrossRef]
- Gervasi, V.; Nilsen, E.B.; Linnell, J.D. Body mass relationships affect the age structure of predation across carnivore–ungulate systems: A review and synthesis. Mammal Rev. 2015, 45, 253–266. [Google Scholar] [CrossRef]
- Prugh, L.R.; Sivy, K.J.; Mahoney, P.J.; Ganz, T.R.; Ditmer, M.A.; van de Kerk, M.; Gilbert, S.L.; Montgomery, R.A. Designing studies of predation risk for improved inference in carnivore-ungulate systems. Biol. Conserv. 2019, 232, 194–207. [Google Scholar] [CrossRef]
- Bohlken, H. Haustiere und zoologische Systematik. Z. Tierzuechtung Zuechtungsbiol. 1961, 76, 107–113. [Google Scholar] [CrossRef]
- Zeller, U.; Göttert, T. The relations between evolution and domestication reconsidered-implications for systematics, ecology, and nature conservation. Glob. Ecol. Conserv. 2019, 20, e00756. [Google Scholar] [CrossRef]
- Ambarlı, H.; Bilgin, C.C. Human–brown bear conflicts in Artvin, northeastern Turkey: Encounters, damage, and attitudes. Ursus 2008, 19, 146–153. [Google Scholar] [CrossRef]
- Zarzo-Arias, A.; Delgado, M.M.; Palazón, S.; Afonso Jordana, I.; Bombieri, G.; González-Bernardo, E.; Bettega, O.C.; García-González, R.; Penteriani, V. Seasonality, local resources and environmental factors influence patterns of brown bear damages: Implications for management. J. Zool. 2021, 313, 1–17. [Google Scholar] [CrossRef]
- Fechter, D.; Storch, I. How many wolves (Canis lupus) fit into Germany? The role of assumptions in predictive rule-based habitat models for habitat generalists. PLoS ONE 2014, 9, e101798. [Google Scholar] [CrossRef]
- Musiani, M.; Muhly, T.; Gates, C.C.; Callaghan, C.; Smith, M.E.; Tosoni, E. Seasonality and reoccurrence of depredation and wolf control in western North America. Wildl. Soc. Bull. 2005, 33, 876–887. [Google Scholar] [CrossRef]
- Wielgus, R.B.; Peebles, K.A. Effects of wolf mortality on livestock depredations. PLoS ONE 2014, 9, e113505. [Google Scholar] [CrossRef] [PubMed]
- Wells, S.L.; McNew, L.B.; Tyers, D.B.; Van Manen, F.T.; Thompson, D.J. Grizzly bear depredation on grazing allotments in the Yellowstone Ecosystem. J. Wildl. Manag. 2019, 83, 556–566. [Google Scholar] [CrossRef]
- Guerisoli, M.D.L.M.; Luengos Vidal, E.; Caruso, N.; Giordano, A.J.; Lucherini, M. Puma–livestock conflicts in the Americas: A review of the evidence. Mammal Rev. 2020, 51, 228–246. [Google Scholar] [CrossRef]
- Zarco-González, M.M.; Monroy-Vilchis, O.; Rodríguez-Soto, C.; Urios, V. Spatial factors and management associated with livestock predations by Puma concolor in Central Mexico. Hum. Ecol. 2012, 40, 631–638. [Google Scholar] [CrossRef]
- Polisar, J.; Maxit, I.; Scognamillo, D.; Farrell, L.; Sunquist, M.E.; Eisenberg, J.F. Jaguars, pumas, their prey base, and cattle ranching: Ecological interpretations of a management problem. Biol. Conserv. 2003, 109, 297–310. [Google Scholar] [CrossRef]
- Michalski, F.; Boulhosa, R.L.P.; Faria, A.; Peres, C.A. Human–wildlife conflicts in a fragmented Amazonian forest landscape: Determinants of large felid depredation on livestock. Anim. Conserv. 2006, 9, 179–188. [Google Scholar] [CrossRef]
- Palmeira, F.B.; Crawshaw Jr, P.G.; Haddad, C.M.; Ferraz, K.M.P.; Verdade, L.M. Cattle depredation by puma (Puma concolor) and jaguar (Panthera onca) in central-western Brazil. Biol. Conserv. 2008, 141, 118–125. [Google Scholar] [CrossRef]
- Kolowski, J.M.; Holekamp, K.E. Spatial, temporal, and physical characteristics of livestock depredations by large carnivores along a Kenyan reserve border. Biol. Conserv. 2006, 128, 529–541. [Google Scholar] [CrossRef]
- Ogada, M.O.; Woodroffe, R.; Oguge, N.O.; Frank, L.G. Limiting depredation by African carnivores: The role of livestock husbandry. Conserv. Biol. 2003, 17, 1521–1530. [Google Scholar] [CrossRef]
- LeFlore, E.G.; Fuller, T.K.; Tomeletso, M.; Stein, A.B. Livestock depredation by large carnivores in northern Botswana. Glob. Ecol. Conserv. 2019, 18, e00592. [Google Scholar] [CrossRef]
- Magrini, C. First data on canids depredation on livestock in an area of recent recolonization by wolf in Central Italy: Considerations on conflict survey and prevention methods. Ekológia 2014, 33, 81. [Google Scholar] [CrossRef]
- Gula, R. Wolf depredation on domestic animals in the Polish Carpathian Mountains. J. Wildl. Manag. 2008, 72, 283–289. [Google Scholar] [CrossRef]
- Sidorovich, V.E.; Tikhomirova, L.L.; Jędrzejewska, B. Wolf Canis lupus numbers, diet and damage to livestock in relation to hunting and ungulate abundance in northeastern Belarus during 1990–2000. Wildl. Biol. 2003, 9, 103–111. [Google Scholar] [CrossRef]
- Nakazawa, C.; Tungalagtuya, K.; Maruyama, N.; Suda, K. Food habits of gray wolves in the Bogdkhan mountain strictly protected area, Mongolia. Biosph. Conserv. Nat. Wildl. Hum. 2008, 9, 1–8. [Google Scholar] [CrossRef]
- Kirilyuk, A.; Ke, R. Wolf depredation on livestock in Daursky State Nature Biosphere Reserve, Russia. J. Nat. Conserv. 2020, 58, 125916. [Google Scholar] [CrossRef]
- Khorozyan, I.; Ghoddousi, S.; Soufi, M.; Soofi, M.; Waltert, M. Studded leather collars are very effective in protecting cattle from leopard (Panthera pardus) attacks. Ecol. Solut. Evid. 2020, 1, e12013. [Google Scholar] [CrossRef]
- Akrim, F.; Mahmood, T.; Belant, J.L.; Nadeem, M.S.; Qasim, S.; Asadi, M.A. Livestock depredations by leopards in Pir Lasura National Park, Pakistan: Characteristics, control and costs. Wildl. Biol. 2021, 2021, 00782. [Google Scholar] [CrossRef]
- Chetri, M.; Odden, M.; Devineau, O.; Wegge, P. Patterns of livestock depredation by snow leopards and other large carnivores in the Central Himalayas, Nepal. Glob. Ecol. Conserv. 2019, 17, e00536. [Google Scholar] [CrossRef]
- Suryawanshi, K.R.; Bhatnagar, Y.V.; Redpath, S.; Mishra, C. People, predators and perceptions: Patterns of livestock depredation by snow leopards and wolves. J. Appl. Ecol. 2013, 50, 550–560. [Google Scholar] [CrossRef]
- Kumar, S.; Khan, J.A.; Khan, A.; Musavi, A.; Malik, P.K.; Kushwaha, S.P.S.; Khati, D.S.; Sarin, G.D. An ecological study in the buffer zone of the Corbett Tiger Reserve: Tiger abundance and cattle depredation. Int. J. Ecol. Environ. Sci. 2008, 34, 133–140. [Google Scholar]
- Rahman, H.A.; Barlow, A.C.D.; Greenwood, C.J.; Islam, M.A.; Ahmad, I.U. Livestock Depredation by Tiger on the Edge of the Bangladesh Sundarbans: A Technical Report; Wildlife Trust of Bangladesh: Dhaka, Bangladesh, 2009. [Google Scholar]
- Soh, Y.H.; Carrasco, L.R.; Miquelle, D.G.; Jiang, J.; Yang, J.; Stokes, E.J.; Tang, J.; Kang, A.; Liu, P.; Rao, M. Spatial correlates of livestock depredation by Amur tigers in Hunchun, China: Relevance of prey density and implications for protected area management. Biol. Conserv. 2014, 169, 117–127. [Google Scholar] [CrossRef]
- Bing, L.I.; Endi, Z.H.A.N.G.; Zhongbao, L.I.U. Livestock depredation by Amur tigers in Hunchun Nature Reserve, Jilin, China. Acta Theriol. Sin. 2009, 29, 231. [Google Scholar]
- Campbell, G.; Coffey, A.; Miller, H.; Read, J.L.; Brook, A.; Fleming, P.J.S.; Bird, P.; Eldridge, S.; Allen, B.L. Dingo baiting did not reduce fetal/calf loss in beef cattle in northern South Australia. Anim. Prod. Sci. 2019, 59, 319–330. [Google Scholar] [CrossRef]
- Salafsky, N.; Salzer, D.; Stattersfield, A.J.; Hilton-Taylor, C.; Neugarten, R.; Butchart, S.H.; Collen, B.; Cox, N.; Master, L.L.; O’Connor, S.; et al. A standard lexicon for biodiversity conservation: Unified classifications of threats and actions. Conserv. Biol. 2008, 22, 897–911. [Google Scholar] [CrossRef]
- Saidur, R.; Rahim, N.A.; Islam, M.R.; Solangi, K.H. Environmental impact of wind energy. Renew. Sustain. Energy Rev. 2011, 15, 2423–2430. [Google Scholar] [CrossRef]
- Jaber, S. Environmental impacts of wind energy. J. Clean Energy Technol. 2013, 1, 251–254. [Google Scholar] [CrossRef] [Green Version]
- Schuster, E.; Bulling, L.; Köppel, J. Consolidating the state of knowledge: A synoptical review of wind energy’s wildlife effects. Environ. Manag. 2015, 56, 300–331. [Google Scholar] [CrossRef] [Green Version]
- Allison, T.D.; Diffendorfer, J.E.; Baerwald, E.F.; Beston, J.A.; Drake, D.; Hale, A.M.; Hein, C.D.; Huso, M.M.; Loss, S.R.; Lovich, J.E.; et al. Impacts to wildlife of wind energy siting and operation in the United States. Issues Ecol. 2019, 21, 2–18. [Google Scholar]
- Hall, L.S.; Richards, G.C. Notes on Tadarida australis (Chiroptera: Molossidae). Aust. Mammal. 1972, 1, 2. [Google Scholar]
- Johnson, G.D. A review of bat mortality at wind-energy developments in the United States. Bat Res. News 2005, 46, 45–49. [Google Scholar]
- Arnett, E.B.; Brown, W.K.; Erickson, W.P.; Fiedler, J.K.; Hamilton, B.L.; Henry, T.H.; Jain, A.; Johnson, G.D.; Kerns, J.; Koford, R.R.; et al. Patterns of bat fatalities at wind energy facilities in North America. J. Wildl. Manag. 2008, 72, 61–78. [Google Scholar] [CrossRef]
- Rydell, J.; Engström, H.; Hedenström, A.; Kyed Larsen, J.; Pettersson, J.; Green, M. The Effect of Wind Power on Birds and Bats: A Synthesis; Naturvårdsverket: Stockholm, Sweden, 2012. [Google Scholar]
- Voigt, C.C.; Lehnert, L.S.; Petersons, G.; Adorf, F.; Bach, L. Wildlife and renewable energy: German politics cross migratory bats. Eur. J. Wildl. Res. 2015, 61, 213–219. [Google Scholar] [CrossRef]
- Thaker, M.; Zambre, A.; Bhosale, H. Wind farms have cascading impacts on ecosystems across trophic levels. Nat. Ecol. Evol. 2018, 2, 1854–1858. [Google Scholar] [CrossRef] [PubMed]
- Bach, L.; Rahmel, U. Summary of wind turbine impacts on bats—Assessment of a conflict. Bremer Beiträge Nat. Nat. 2004, 7, 245–252. [Google Scholar]
- Arnett, E.B.; Baerwald, E.F.; Mathews, F.; Rodrigues, L.; Rodríguez-Durán, A.; Rydell, J.; Villegas-Patraca, R.; Voigt, C.C. Impacts of wind energy development on bats: A global perspective. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Voigt, C.C., Kingston, T., Eds.; Springer: Cham, Switzerland, 2016; pp. 295–323. [Google Scholar]
- O’Shea, T.J.; Cryan, P.M.; Hayman, D.T.; Plowright, R.K.; Streicker, D.G. Multiple mortality events in bats: A global review. Mammal Rev. 2016, 46, 175–190. [Google Scholar] [CrossRef]
- Voigt, C.C.; Straka, T.M.; Fritze, M. Producing wind energy at the cost of biodiversity: A stakeholder view on a green-green dilemma. J. Renew. Sustain. Energy 2019, 11, 063303. [Google Scholar] [CrossRef]
- Straka, T.M.; Fritze, M.; Voigt, C.C. The human dimensions of a green–green-dilemma: Lessons learned from the wind energy—Wildlife conflict in Germany. Energy Rep. 2020, 6, 1768–1777. [Google Scholar] [CrossRef]
- MacEwan, K.L.; Morgan, T.W.; Lötter, C.A.; Tredennick, A.T. Bat activity across South Africa: Implications for wind energy development. Afr. J. Wildl. Res. 2000, 50. [Google Scholar] [CrossRef]
- Dürr, T. Fledermäuse als Opfer von Windkraftanlagen in Deutschland. Nyctalus 2002, 8, 115–118. [Google Scholar]
- Dürr, T.; Bach, L. Bat deaths and wind turbines—A review of current knowledge, and of the information available in the database for Germany. Bremer Beiträge Nat. Nat. 2004, 7, 253–264. [Google Scholar]
- Rydell, J.; Bach, L.; Dubourg-Savage, M.J.; Green, M.; Rodrigues, L.; Hedenström, A. Mortality of bats at wind turbines links to nocturnal insect migration? Eur. J. Wildl. Res. 2010, 56, 823–827. [Google Scholar] [CrossRef] [Green Version]
- Rydell, J.; Bach, L.; Dubourg-Savage, M.J.; Green, M.; Rodrigues, L.; Hedenström, A. Bat mortality at wind turbines in northwestern Europe. Acta Chiropterologica 2010, 12, 261–274. [Google Scholar] [CrossRef]
- Grodsky, S.M.; Behr, M.J.; Gendler, A.; Drake, D.; Dieterle, B.D.; Rudd, R.J.; Walrath, N.L. Investigating the causes of death for wind turbine-associated bat fatalities. J. Mammal. 2011, 92, 917–925. [Google Scholar] [CrossRef] [Green Version]
- Baerwald, E.F.; D’Amours, G.H.; Klug, B.J.; Barclay, R.M. Barotrauma is a significant cause of bat fatalities at wind turbines. Curr. Biol. 2008, 18, R695–R696. [Google Scholar] [CrossRef]
- Ingersoll, T.E.; Sewall, B.J.; Amelon, S.K. Improved analysis of long-term monitoring data demonstrates marked regional declines of bat populations in the eastern United States. PLoS ONE 2013, 8, e65907. [Google Scholar] [CrossRef] [Green Version]
- Zahn, A.; Lustig, A.; Hammer, M. Potenzielle Auswirkungen von Windenergieanlagen auf Fledermauspopulationen. Anliegen Nat. 2014, 36, 21–35. [Google Scholar]
- Frick, W.F.; Baerwald, E.F.; Pollock, J.F.; Barclay, R.M.; Szymanski, J.A.; Weller, T.J.; Russell, A.L.; Loeb, S.C.; Medellin, R.A.; McGuire, L.P. Fatalities at wind turbines may threaten population viability of a migratory bat. Biol. Conserv. 2017, 209, 172–177. [Google Scholar] [CrossRef]
- Brinkmann, R.; Schauer-Weisshahn, H.; Bontadina, F. Survey of Possible Operational Impacts on Bats by Wind Facilities in Southern Germany; Report for Administrative District of Freiburg–Department; ECON Ecological Consultancy: Gundelfingen, Germany, 2006; p. 56. [Google Scholar]
- Frey-Ehrenbold, A.; Bontadina, F.; Arlettaz, R.; Obrist, M.K. Landscape connectivity, habitat structure and activity of bat guilds in farmland-dominated matrices. J. Appl. Ecol. 2013, 50, 252–261. [Google Scholar] [CrossRef] [Green Version]
- Claireau, F.; Bas, Y.; Pauwels, J.; Barré, K.; Machon, N.; Allegrini, B.; Puechmaille, S.J.; Kerbiriou, C. Major roads have important negative effects on insectivorous bat activity. Biol. Conserv. 2019, 235, 53–62. [Google Scholar] [CrossRef]
- Rodrigues, L.; Bach, L.; Dubourg-Savage, M.-J.; Karapandza, B.; Kovac, D.; Kervyn, T.; Dekker, J.; Kepel, A.; Bach, P.; Collins, J.; et al. Guidelines for Consideration of Bats in Wind Farm Projects—Revision 2014; EUROBATS Publication Serie; UNEP/EUROBATS: Bonn, Germany, 2015; p. 133. [Google Scholar]
- Barré, K.; Le Viol, I.; Bas, Y.; Julliard, R.; Kerbiriou, C. Estimating Habitat Loss Due to Wind Turbine Avoidance by Bats: Implications for European Siting Guidance. Biol. Conserv. 2018, 226, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Ferri, V.; Battisti, C.; Soccini, C. Bats in a mediterranean mountainous landscape: Does wind farm repowering induce changes at assemblage and species level? Environ. Manag. 2016, 57, 1240–1246. [Google Scholar] [CrossRef] [PubMed]
- Barclay, R.M.; Baerwald, E.F.; Gruver, J.C. Variation in bat and bird fatalities at wind energy facilities: Assessing the effects of rotor size and tower height. Can. J. Zool. 2007, 85, 381–387. [Google Scholar] [CrossRef]
- Cryan, P.M.; Barclay, R.M. Causes of bat fatalities at wind turbines: Hypotheses and predictions. J. Mammal. 2009, 90, 1330–1340. [Google Scholar] [CrossRef] [Green Version]
- Guest, E.E.; Stamps, B.F.; Durish, N.D.; Hale, A.M.; Hein, C.D.; Morton, B.P.; Weaver, S.P.; Fritts, S.R. An updated review of hypotheses regarding bat attraction to wind turbines. Animals 2022, 12, 343. [Google Scholar] [CrossRef] [PubMed]
- Kunz, T.H.; Arnett, E.B.; Erickson, W.P.; Hoar, A.R.; Johnson, G.D.; Larkin, R.P.; Strickland, M.D.; Thresher, R.W.; Tuttle, M.D. Ecological impacts of wind energy development on bats: Questions, research needs, and hypotheses. Front. Ecol. Environ. 2007, 5, 315–324. [Google Scholar] [CrossRef] [Green Version]
- Russ, J.M.; Montgomery, W.I. Habitat associations of bats in Northern Ireland: Implications for conservation. Biol. Conserv. 2002, 108, 49–58. [Google Scholar] [CrossRef]
- Kusch, J.; Schotte, F. Effects of fine-scale foraging habitat selection on bat community structure and diversity in a temperate low mountain range forest. Folia Zool. 2007, 56, 263. [Google Scholar]
- Buchholz, S.; Kelm, V.; Ghanem, S.J. Mono-specific forest plantations are valuable bat habitats: Implications for wind energy development. Eur. J. Wildl. Res. 2021, 67, 1–12. [Google Scholar] [CrossRef]
- Starik, N.; Göttert, T.; Heitlinger, E.; Zeller, U. Bat community responses to structural habitat complexity resulting from management practices within different land use types—A case study from north-eastern Germany. Acta Chiropterologica 2018, 20, 387–405. [Google Scholar] [CrossRef]
- Lehnert, L.S.; Kramer-Schadt, S.; Schönborn, S.; Lindecke, O.; Niermann, I.; Voigt, C.C. Wind farm facilities in Germany kill noctule bats from near and far. PLoS ONE 2014, 9, e103106. [Google Scholar] [CrossRef] [Green Version]
- Kuvlesky, W.P., Jr.; Brennan, L.A.; Morrison, M.L.; Boydston, K.K.; Ballard, B.M.; Bryant, F.C. Wind energy development and wildlife conservation: Challenges and opportunities. J. Wildl. Manag. 2007, 71, 2487–2498. [Google Scholar] [CrossRef]
- Rodrigues, L.; Bach, L.; Dubourg-Savage, M.-J.; Goodwin, J.; Harbusch, C. Guidelines for Consideration of Bats in Wind Farm Projects; EUROBATS Publication Series No. 3 (English Version); UNEP/EUROBATS Secretariat: Bonn, Germany, 2008; p. 51. [Google Scholar]
- Baerwald, E.F.; Edworthy, J.; Holder, M.; Barclay, R.M. A large-scale mitigation experiment to reduce bat fatalities at wind energy facilities. J. Wildl. Manag. 2009, 73, 1077–1081. [Google Scholar] [CrossRef]
- Arnett, E.B.; Huso, M.M.; Schirmacher, M.R.; Hayes, J.P. Altering turbine speed reduces bat mortality at wind-energy facilities. Front. Ecol. Environ. 2011, 9, 209–214. [Google Scholar] [CrossRef] [Green Version]
- Köppel, J.; Dahmen, M.; Helfrich, J.; Schuster, E.; Bulling, L. Cautious but committed: Moving toward adaptive planning and operation strategies for renewable energy‘s wildlife implications. Environ. Manag. 2014, 54, 744–755. [Google Scholar] [CrossRef]
- Richardson, S.M.; Lintott, P.R.; Hosken, D.J.; Economou, T.; Mathews, F. Peaks in bat activity at turbines and the implications for mitigating the impact of wind energy developments on bats. Sci. Rep. 2021, 11, 3636. [Google Scholar] [CrossRef]
- Voigt, C.C.; Russo, D.; Runkel, V.; Goerlitz, H.R. Limitations of acoustic monitoring at wind turbines to evaluate fatality risk of bats. Mammal Rev. 2021, 51, 559–570. [Google Scholar] [CrossRef]
- Romano, W.B.; Skalski, J.R.; Townsend, R.L.; Kinzie, K.W.; Coppinger, K.D.; Miller, M.F. Evaluation of an acoustic deterrent to reduce bat mortalities at an Illinois wind farm. Wildl. Soc. Bull. 2019, 43, 608–618. [Google Scholar] [CrossRef] [Green Version]
- Arnett, E.B.; Hein, C.D.; Schirmacher, M.R.; Huso, M.M.; Szewczak, J.M. Evaluating the effectiveness of an ultrasonic acoustic deterrent for reducing bat fatalities at wind turbines. PLoS ONE 2013, 8, e65794. [Google Scholar] [CrossRef]
- Fritze, M.; Lehnert, L.S.; Heim, O.; Lindecke, O.; Roeleke, M.; Voigt, C.C. Fledermäuse im Schatten der Windenergie: Deutschlands Expert*innen vermissen Transparenz und bundesweite Standards in den Genehmigungsverfahren. Nat. Landsch. 2019, 51, 20–27. [Google Scholar]
- Manfredo, M.J.; Bruskotter, J.T.; Teel, T.L.; Fulton, D.; Schwartz, S.H.; Arlinghaus, R.; Oishi, S.; Uskul, A.K.; Redford, K.; Kitayama, S.; et al. Why social values cannot be changed for the sake of conservation. Conserv. Biol. 2017, 31, 772–780. [Google Scholar] [CrossRef]
Figure 3.
Herding small livestock (sheep and goats) is labor and time intensive but also a very effective method of minimizing carnivore–livestock conflicts. Bottom picture: cattle herdsmen in East Africa, top picture: shepherd in Germany. While this type of livestock husbandry practice was widespread in Central Europe in the past, it has increasingly fallen into oblivion in the course of intensified agriculture and is practiced almost exclusively in the context of nature conservation activities (musealization). It is still applied on the African continent, although it is also subject to change there [61]. Supra-regional considerations can help to transfer the experiences and knowledge of the nomads of East Africa to the situation in Central Europe, recolonized by various large carnivore species. Photo top: Christel Simantke, photo bottom: T. Göttert.
Figure 3.
Herding small livestock (sheep and goats) is labor and time intensive but also a very effective method of minimizing carnivore–livestock conflicts. Bottom picture: cattle herdsmen in East Africa, top picture: shepherd in Germany. While this type of livestock husbandry practice was widespread in Central Europe in the past, it has increasingly fallen into oblivion in the course of intensified agriculture and is practiced almost exclusively in the context of nature conservation activities (musealization). It is still applied on the African continent, although it is also subject to change there [61]. Supra-regional considerations can help to transfer the experiences and knowledge of the nomads of East Africa to the situation in Central Europe, recolonized by various large carnivore species. Photo top: Christel Simantke, photo bottom: T. Göttert.

Figure 4.
Conceptual framework: stepwise adjustment of measures to mitigate HWC from the global to the local scale, including a gradual adjustment from a broad systematic category towards a certain species, sometimes a population, or group of individuals. Requirement: global distribution and a certain degree of ecological similarities among the representatives of a monophylum involved in HWC (specification of the wildlife concept); see text for further explanation.
Figure 4.
Conceptual framework: stepwise adjustment of measures to mitigate HWC from the global to the local scale, including a gradual adjustment from a broad systematic category towards a certain species, sometimes a population, or group of individuals. Requirement: global distribution and a certain degree of ecological similarities among the representatives of a monophylum involved in HWC (specification of the wildlife concept); see text for further explanation.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Göttert, T.; Starik, N. Human–Wildlife Conflicts across Landscapes—General Applicability vs. Case Specificity. Diversity 2022, 14, 380. https://0-doi-org.brum.beds.ac.uk/10.3390/d14050380
AMA Style
Göttert T, Starik N. Human–Wildlife Conflicts across Landscapes—General Applicability vs. Case Specificity. Diversity. 2022; 14(5):380. https://0-doi-org.brum.beds.ac.uk/10.3390/d14050380
Chicago/Turabian StyleGöttert, Thomas, and Nicole Starik. 2022. "Human–Wildlife Conflicts across Landscapes—General Applicability vs. Case Specificity" Diversity 14, no. 5: 380. https://0-doi-org.brum.beds.ac.uk/10.3390/d14050380
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.