
Molecular Identification and Novel Mitochondrial COI Gene Haplotypes of Nesting Kemp’s Ridley Turtles (Lepidochelys kempii) in Rancho Nuevo Sanctuary, Mexico

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
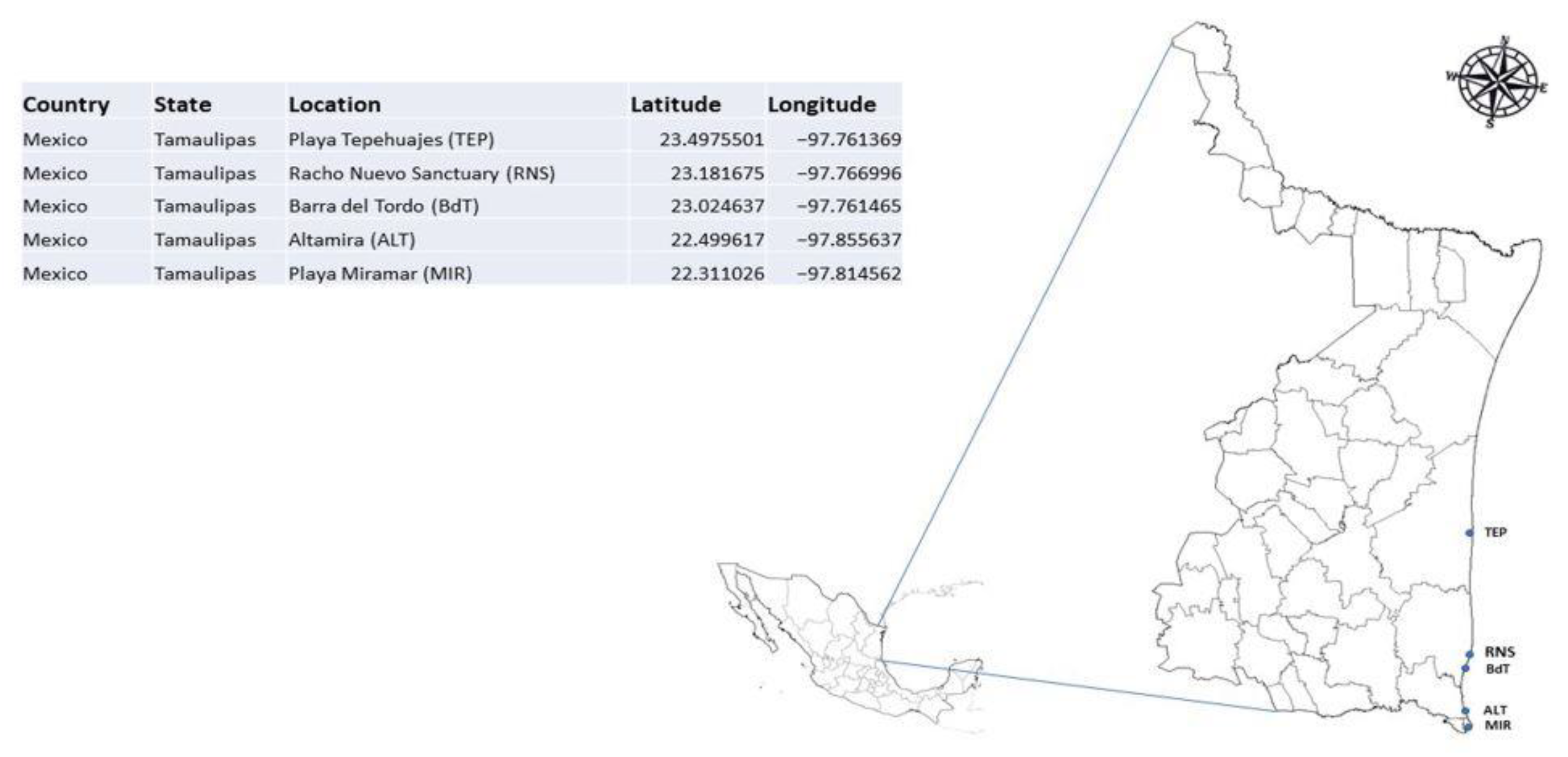
2.1. Field Specimens and DNA Biobank
2.2. Amplification and Sequence of Gene Segments
2.3. Data and Quality Sequence Analysis
2.4. Haplotype Analysis
2.5. Tree-Building for COI
2.6. Population Gene Parameters
3. Results
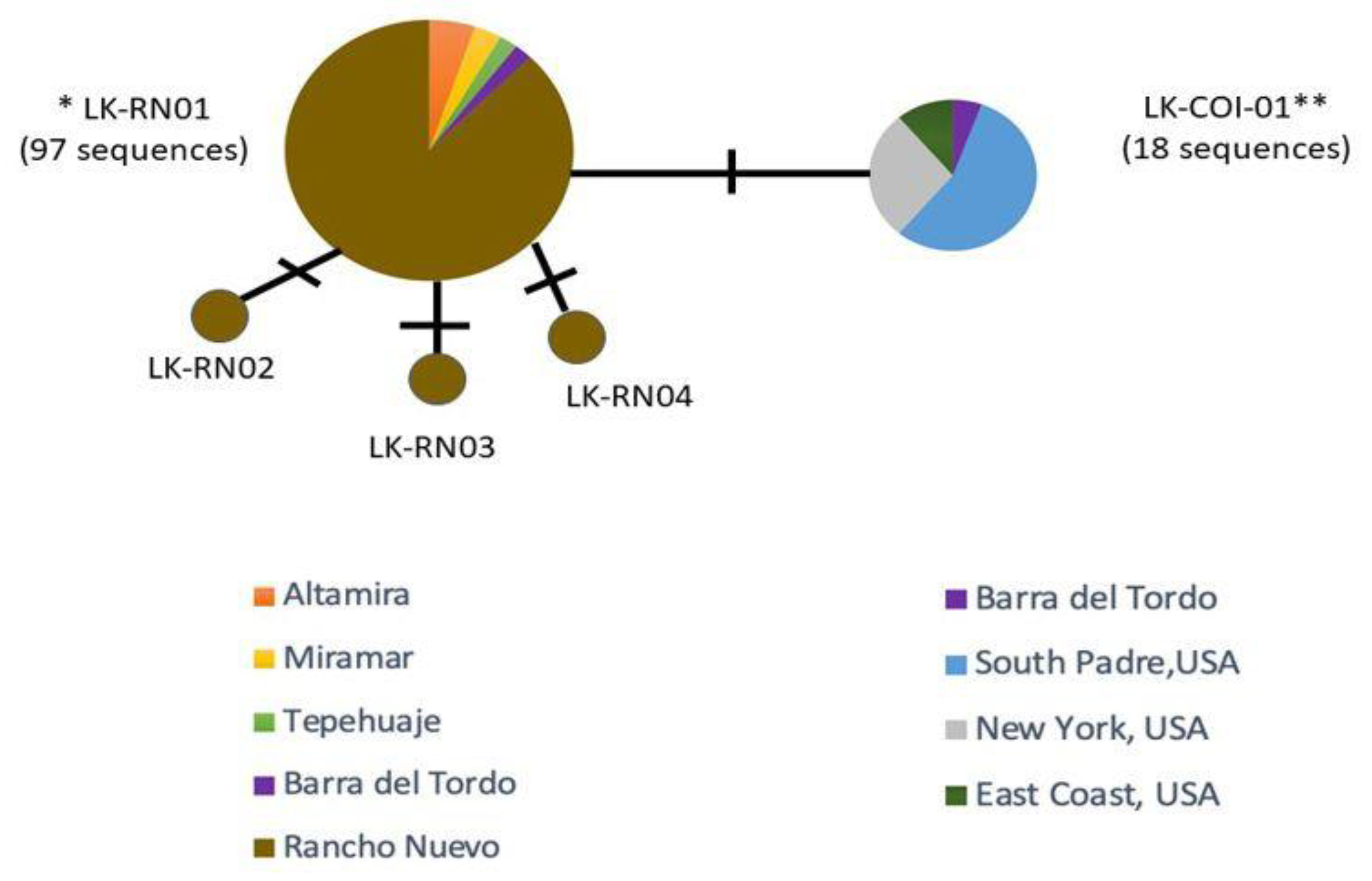
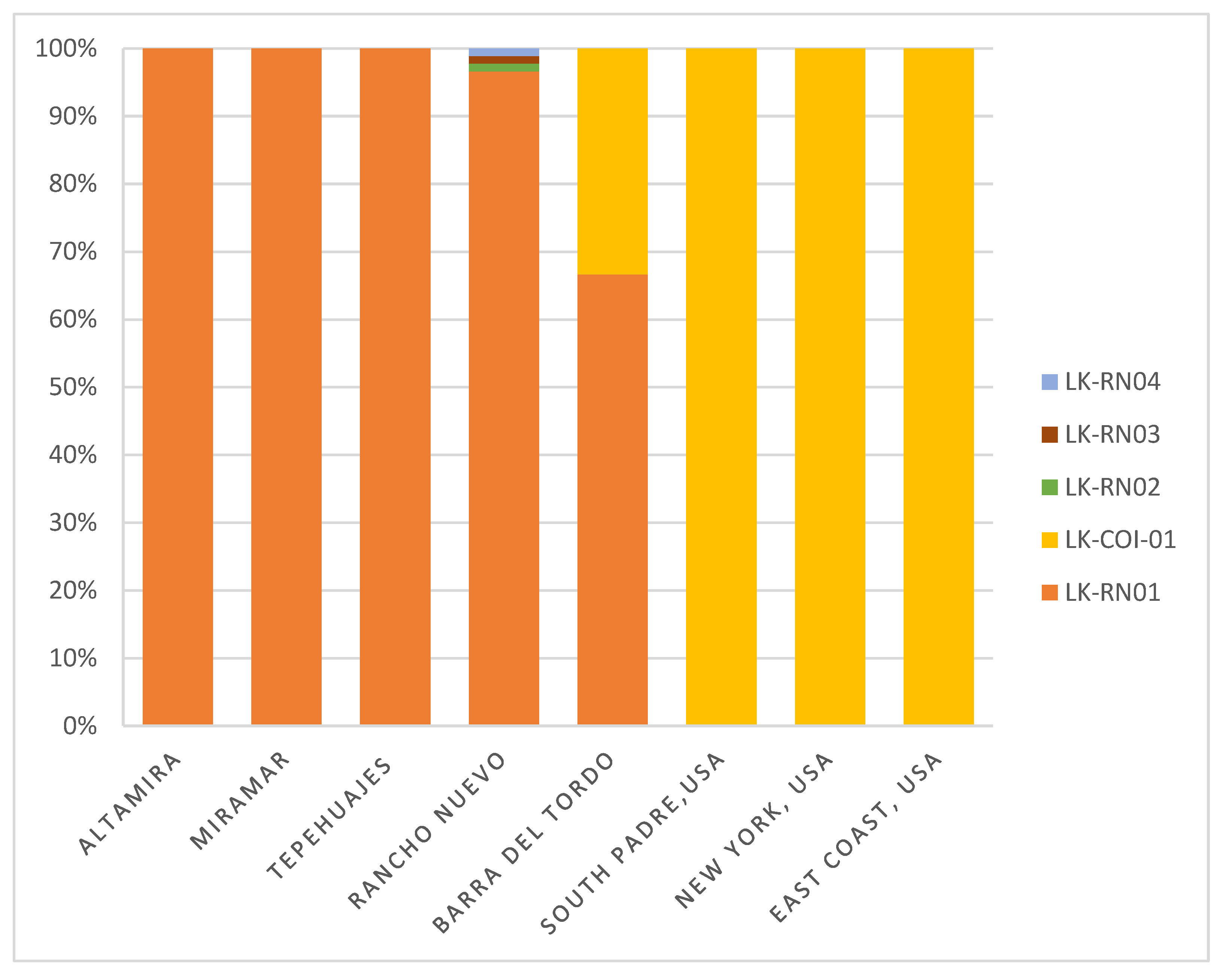
3.1. Haplotype Diversity
3.2. Genetic Divergence
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- CIT, Convención Interamericana para la Protección y Conservación de las Tortugas Marinas. 2006. Available online: http://www.iacseaturtle.org/docs/resolucionesCOP3CT/Informe-Final-COP3CIT-ESP.pdf (accessed on 2 April 2022).
- IUCN. The IUCN Red List of Threatened Species. Version 2021-3. 2021. Available online: https://www.iucnredlist.org (accessed on 2 April 2022).
- Morreale, S.J.; Meylan, A.B.; Sadove, S.S.; Standora, E.A. Annual occurrence of winter mortality of marine turtles in New York waters. J. Herpetol. 1992, 26, 301–308. [Google Scholar] [CrossRef]
- Gaona Pineda, O.; Barragán, A.R. Las tortugas marinas en México: Logros y perspectivas para su conservación. Cienc. Y Mar. 2017, XXI, 73–74. [Google Scholar]
- Liceaga Correa, M.d.l.Á. Hábitats críticos y hotspots de tortugas marinas. In Atlas de Línea Base Ambiental del Golfo de México, 2020th ed.; Herzka, S.Z., Alfonso, R., Álvarez, Z., Peters Recagno, E.M., Hernández, G., Coordinación, C., Vii, T., Uribe Martínez Y Edgar, A., Castro, R., Cartografía, P., et al., Eds.; CICESE: Mexico City, México, 2020; pp. 6–29. [Google Scholar]
- Bernardo, J.; Plotkin, P.T. An evolutionary perspective on the arribada phenomenon and reproductive behavioral polymorphism of olive ridley sea turtles (Lepidochelys olivacea). In The Biology and Conservation of Ridley Sea Turtles; Plotkin, P.T., Ed.; JHU Press: Baltimore, MD, USA, 2007; pp. 58–87. [Google Scholar]
- Márquez, M.R. Synopsis of Biological Data on the Kemp’s Ridley Turtle, Lepidochelys kempi (Garman, 1880); Southeast Fisheries Science Center: Miami, FL, USA, 1994. [Google Scholar]
- Caillouet, C.W., Jr. Interruption of the Kemp’s ridley population’s pre-2010 exponential growth in the Gulf of Mexico and its aftermath: One hypothesis. Mar. Turt. Newsl. 2014, 143, 1–7. [Google Scholar]
- Bevan, E.; Wibbels, T.; Najera, B.M.Z.; Sarti, L.; Martinez, F.I.; Cuevas, J.M.; Gallaway, B.J.; Pena, L.J.; Burchfield, P.M. Estimating the historic size and current status of the Kemp’s ridley sea turtle (Lepidochelys kempii) population. Ecosphere 2016, 7, e01244. [Google Scholar] [CrossRef] [Green Version]
- Reyes-López, M.A.; Camacho-Sánchez, F.Y.; Hart, C.E.; Leal-Sepúlveda, V.; Zavala-Felix, K.A.; Ley-Quiñónez, C.P.; Aguirre, A.A.; Zavala-Norzagaray, A.A. Rediscovering Kemp’s Ridley Sea Turtle Lepidochelys kempii: Molecular Analysis and Threats. In Natural History and Ecology of Mexico and Central America; IntechOpen: London, UK, 2021; Available online: https://www.intechopen.com/chapters/75625 (accessed on 2 April 2022). [CrossRef]
- Lara-Uc, M.; Mota-Rodríguez, C. Conociendo a la Tortuga Golfina, Lepidochelys olivacea (Eschscholtz, 1829). Bioma 2014, 5, 9. [Google Scholar]
- Avens, L.; Goshe, L.R.; Coggins, L.; Shaver, D.J.; Higgins, B.; Landry, A.M., Jr.; Bailey, R. Variability in age and size at maturation, reproductive longevity, and long-term growth dynamics for Kemp’s ridley sea turtles in the Gulf of Mexico. PLoS ONE 2017, 12, e0173999. [Google Scholar] [CrossRef]
- Ward, R.D. DNA barcode divergence among species and genera of birds and fishes. Mol. Ecol. Resour. 2009, 9, 1077–1085. [Google Scholar] [CrossRef]
- Beebe, N.W. DNA barcoding mosquitoes: Advice for potential prospectors. Parasitology 2018, 145, 622–633. [Google Scholar] [CrossRef]
- Nneji, L.M.; Adeola, A.C.; Ayoola, A.O.; Oladipo, S.O.; Wang, Y.Y.; Malann, Y.D.; Anyaele, O.; Nneji, I.C.; Rahman, M.M.; Olory, C.S. DNA barcoding and species delimitation of butterflies (Lepidoptera) from Nigeria. Mol. Biol. Rep. 2020, 47, 9441–9457. [Google Scholar] [CrossRef]
- Jamaluddin, J.A.F.; Mohammed Akib, N.A.; Ahmad, S.Z.; Abdul Halim, S.A.A.; Abdul Hamid, N.K.; Mohd Nor, S.A. DNA barcoding of shrimps from a mangrove biodiversity hotspot. Mitochondrial DNA Part A DNA Mapp. Seq. Anal. 2019, 30, 618–625. [Google Scholar] [CrossRef] [PubMed]
- Tikochinski, Y.; Carreras, C.; Tikochinski, G.; Vilaça, S.T. Population-specific signatures of intra-individual mitochondrial DNA heteroplasmy and their potential evolutionary advantages. Sci. Rep. 2020, 10, 1–14. [Google Scholar]
- Zhang, T.; Lin, L.; Gaillard, D.; Chen, F.; Chen, H.; Li, M.; Wu, S.; Wang, Z.; Shi, H. Rapid authenticity testing of artificially bred green turtles ( Chelonia mydas) using microsatellite and mitochondrial DNA markers. PeerJ 2021, 9, e12410. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; DeWaard, J.R. Biological identifications through DNA barcodes. Proc. Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Duchene, S.; Frey, A.; Alfaro-Núñez, A.; Dutton, P.H.; Thomas, M.; Morin, P.A. Marine turtle mitogenome phylogenetics and evolution. Mol. Phylogenet. Evol. 2012, 65, 241–250. [Google Scholar] [CrossRef]
- Vargas, S.M.; Jensen, M.P.; Ho, S.Y.; Mobaraki, A.; Broderick, D.; Mortimer, J.A.; Whiting, S.D.; Miller, J.; Prince, R.I.; Bell, I.P.; et al. Phylogeography, genetic diversity, and management units of hawksbill turtles in the indo-pacific. J. Hered. 2016, 107, 199–213. [Google Scholar] [CrossRef] [Green Version]
- Naro-Maciel, E.; Le, M.; FitzSimmons, N.N.; Amato, G. Evolutionary relationships of marine turtles: A molecular phylogeny based on nuclear and mitochondrial genes. Mol. Phylogenet. Evol. 2008, 49, 659–662. [Google Scholar] [CrossRef]
- Vargas, S.M.; Araújo, F.C.F.; Santos, F.R. DNA barcoding of Brazilian sea turtles (Testudines). Genet. Mol. Biol. 2009, 32, 608. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Gregory, T.R. The promise of DNA barcoding for taxonomy. Syst. Biol. 2005, 54, 852–859. [Google Scholar] [CrossRef]
- Komoroske, L.M.; Jensen, M.P.; Stewart, K.R.; Shamblin, B.M.; Dutton, P.H. Advances in the application of genetics in marine turtle biology and conservation. Front. Mar. Sci. 2017, 4, 156. [Google Scholar] [CrossRef] [Green Version]
- Naro-Maciel, E.; Reid, B.; Fitzsimmons, N.N.; Le, M.; Desalle, R.; Amato, G. DNA barcodes for globally threatened marine turtles: A registry approach to documenting biodiversity. Mol. Ecol. Resour. 2010, 10, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Morgada, M.N.; Abriata, L.A.; Cefaro, C.; Gajda, K.; Banci, L.; Vila, A.J. Loop recognition and copper-mediated disulfide reduction underpin metal site assembly of CuA in human cytochrome oxidase. Proc. Natl. Acad. Sci. USA 2015, 112, 11771–11776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porter, T.M.; Hajibabaei, M. Automated high throughput animal CO1 metabarcode classification. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Eckert, K.L.; Bjorndal, K.A.; Abreu-Grobois, F.A.; Donnelly, M. (Eds.) Técnicas de Investigación y Manejo para la Conservación de las Tortugas Marinas; Grupo Especialista en Tortugas Marinas (Spanish); SSC/IUCN Marine Turtle Specialist Group: Washington, DC, USA, 2000; Volume 4. [Google Scholar]
- Carreras, C.; Rees, A.F.; Broderick, A.C.; Godley, B.J.; Margaritoulis, D. Mitochondrial DNA markers of loggerhead marine turtles (Caretta caretta) (Testudines: Cheloniidae) nesting at Kyparissia Bay, Greece, confirm the western Greece unit and regional structuring. Sci. Mar. 2014, 78, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Craven, K.S.; Hodgson, J.Y.S.; Shaver, D.J.; Walker, J.S.; Villalba-Guerra, M.R.; Owens, D.W. Evaluation of gonadal tissue to validate size at reproductive maturity in Kemp’s ridley sea turtles found stranded in Texas, USA. Diversity 2019, 11, 76. [Google Scholar] [CrossRef] [Green Version]
- Owens, W.; Ruiz, G.J. New Methods of Obtaining Blood and Cerebrospinal Fluid From Marine Turtles. Herpetologica 1980, 36, 17–20. [Google Scholar]
- FitzSimmons, N.N.; Moritz, C.; Bowen, B. Population identification. In Research and management techniques for the conservation of sea turtles, 1st ed.; Eckert, K.L., Bjorndal, K.A., Abreu-Grobois, F.A., Donnelly, M., Eds.; SSC/IUCN Marine Turtle Specialist Group: Washington, DC, USA, 1999; pp. 82–88. [Google Scholar]
- Ivanova, N.V.; Zemlak, T.S.; Hanner, R.H.; Hebert, P.D.N. Universal primer cocktails for fish DNA barcoding. Mol. Ecol. Notes 2007, 7, 544–548. [Google Scholar] [CrossRef]
- Becker, S.; Hanner, R.; Steinke, D. Five years of FISH-BOL: Brief status report. Mitochondrial DNA 2011, 22, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Camacho-Sánchez, F.Y.; Aguirre, A.A.; Narváez-Zapata, J.A.; Zavala-Norzagaray, A.A.; Ley-Quiñónez, C.P.; Acosta-Sánchez, H.H.; Rodriguez-González, H.; Delgado-Trejo, C.; Reyes-López, M.A. DNA barcode analysis of the endangered green turtle (Chelonia mydas) in Mexico. Genome 2021, 64, 879–891. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leigh, J.W.; Bryant, D. Popart: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Nguyen, M.A.T.; Von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Hudson, R.R.; Slatkin, M.; Maddison, W.P. Estimation of levels of gene flow from DNA sequence data. Genetics 1992, 132, 583–589. [Google Scholar] [CrossRef]
- Phillips, J.D.; French, S.H.; Hanner, R.H.; Gillis, D.J. HACSim: An R package to estimate intraspecific sample sizes for genetic diversity assessment using haplotype accumulation curves. PeerJ Comput. Sci. 2020, 6, e243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archie Carr Center for Sea Turtle Research, “mtDNA Sequences”. Available online: https://accstr.ufl.edu/resources/mtdna-sequences/ (accessed on 15 March 2022).
- Pentinsaari, M.; Salmela, H.; Mutanen, M.; Roslin, T. Molecular evolution of a widely-adopted taxonomic marker (COI) across the animal tree of life. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef]
- Frandsen, H.R.; Figueroa, D.F.; George, J.A. Mitochondrial genomes and genetic structure of the Kemp’s ridley sea turtle (Lepidochelys kempii). Ecol. Evol. 2020, 10, 249–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gredzens, C.; Shaver, D.J. Satellite Tracking Can Inform Population-Level Dispersal to Foraging Grounds of Post-nesting Kemp’s Ridley Sea Turtles. Front. Mar. Sci. 2020, 7, 559. [Google Scholar] [CrossRef]
- Shaver, D.J.; Hart, K.M.; Fujisaki, I.; Rubio, C.; Sartain, A.R.; Peña, J.; Burchfield, P.M.; Gamez, D.G.; Ortiz, J. Foraging area fidelity for Kemp’s ridleys in the Gulf of Mexico. Ecol. Evol. 2013, 3, 2002–2012. [Google Scholar] [CrossRef]
- Uribe-Martínez, A.; Liceaga-Correa, M.A.; Cuevas, E. Critical in-water habitats for post-nesting sea turtles from the southern gulf of mexico. J. Mar. Sci. Eng. 2021, 9, 793. [Google Scholar] [CrossRef]
- Covelo, P.; Nicolau, L.; López, A. Four new records of stranded Kemp’s ridley turtle Lepidochelys kempii in the NW Iberian Peninsula. Mar. Biodivers. Rec. 2016, 9, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Bean, S.B.; Logan, J.M. Stable isotope analyses of cold-stunned Kemp’s ridley (Lepidochelys kempii) sea turtles at the northern extent of their coastal range. Mar. Biol. 2019, 166, 1–10. [Google Scholar] [CrossRef]
- Gallaway, B.J.; Gazey, W.J.; Caillouet, C.W., Jr.; Plotkin, P.T.; Grobois, F.A.A.; Amos, A.F.; Burchfield, P.M.; Carthy, R.R.; Martínez, M.A.C.; Cole, J.G.; et al. Development of a Kemp’s ridley sea turtle stock assessment model. Gulf Mex. Sci. 2016, 33, 138–157. [Google Scholar] [CrossRef] [Green Version]
- Caillouet, C.W., Jr.; Gallaway, B.J. Kemp’s Ridley Sea Turtle Emigration and Immigration Between The Gulf Of Mexico And North Atlantic Ocean Should Not Be Ignored In Age-Structured Population Modeling. Mar. Turt. Newsl. 2020, 161, 9–14. [Google Scholar]
- Caillouet, C.W.J. Substantial Reduction in Annual Production of Kemp’s Ridley Sea Turtle Hatchlings on Beaches of Tamaulipas, Mexico May Allow Abundance of Adults to Increase. Mar. Turt. Newsl. 2021, 163, 1–7. [Google Scholar]
- Miller, J.M.; Quinzin, M.C.; Edwards, D.L.; Eaton, D.A.R.; Jensen, E.L.; Russello, M.A.; Gibbs, J.P.; Tapia, W.; Rueda, D.; Caccone, A. Genome-Wide Assessment of Diversity and Divergence Among Extant Galapagos Giant Tortoise Species. J. Hered. 2018, 109, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Bhaskar, R.; Mohindra, V. Variability in DNA COI sequences reveals new haplotypes in freshwater turtles from northern region of India. Mitochondrial DNA Part B Resour. 2018, 3, 317–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamont, M.M.; Moreno, N.; Camacho-Sánchez, F.Y.; Acosta-Sánchez, H.H.; Glaberman, S.; Reyes-Lopez, M.A.; Chiari, Y. Genetic diversity of immature Kemp’s ridley (Lepidochelys kempii) sea turtles from the northern Gulf of Mexico. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 3003–3010. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID Sequence | Position | Number of Sequences (n) | Number of Haplotypes (H) | h | Pi | G + C | |||
|---|---|---|---|---|---|---|---|---|---|
| 71 | 550 | 590 | 604 | ||||||
| Population (LK-COI-01 */LK-RN01) | NA | NA | NA | NA | 118 | 2 | 0.303 | 0.00052 | 0.422 |
| LK-COI-01 * | C | T | T | A | 97 | 1 | 0 | 0 | ND |
| LK-RN01 | C | 18 | 1 | 0 | 0 | ND | |||
| LK-RN02 | C | C | 1 | 1 | ND | ND | ND | ||
| LK-RN03 | C | C | 1 | 1 | ND | ND | ND | ||
| LK-RN04 | T | C | 1 | 1 | ND | ND | ND | ||
| HAPLOTYPES | Altamira MEX | Miramar MEX | Tepehuajes MEX | Barra del Tordo MEX | Rancho Nuevo MEX | South Padre USA | New York USA | East Coast USA | TOTAL |
|---|---|---|---|---|---|---|---|---|---|
| LK-RN01 | 5 | 3 | 2 | 2 | 85 | 0 | 0 | 0 | 97 |
| LK-COI-01 | 0 | 0 | 0 | 1 | 0 | 10 | 5 | 2 | 18 |
| LK-RN02 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 |
| LK-RN03 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 |
| LK-RN04 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 |
| TOTAL | 5 | 3 | 2 | 3 | 88 | 10 | 5 | 2 | 118 |
| Species | Lk | Lo | Cc | CmP | CmA | Ei | Dc | Nd | Cs |
|---|---|---|---|---|---|---|---|---|---|
| Lepidochelys kempii (Lk) | 0.02 | 0.05 | 0.08 | 0.08 | 0.07 | 0.10 | 0.08 | 0.14 | |
| Lepidochelys olivacea (Lo) | 0.02 | 0.05 | 0.08 | 0.08 | 0.07 | 0.11 | 0.08 | 0.15 | |
| Caretta caretta (Cc) | 0.05 | 0.06 | 0.09 | 0.08 | 0.07 | 0.11 | 0.08 | 0.14 | |
| Chelonia mydas (CmP) | 0.08 | 0.08 | 0.10 | 0.01 | 0.08 | 0.12 | 0.06 | 0.14 | |
| Chelonia mydas (CmA) | 0.09 | 0.08 | 0.09 | 0.01 | 0.08 | 0.11 | 0.06 | 0.13 | |
| Eretmochelys imbricata (Ei) | 0.07 | 0.08 | 0.08 | 0.08 | 0.08 | 0.10 | 0.08 | 0.13 | |
| Dermochelys coriacea (Dc) | 0.12 | 0.12 | 0.13 | 0.13 | 0.13 | 0.11 | 0.11 | 0.15 | |
| Natator depressus (Nd) | 0.09 | 0.09 | 0.08 | 0.08 | 0.07 | 0.09 | 0.12 | 0.13 | |
| Centrochelys sulcata (Cs) | 0.16 | 0.17 | 0.16 | 0.16 | 0.15 | 0.15 | 0.17 | 0.14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camacho-Sánchez, F.Y.; Narváez-Zapata, J.A.; Acosta-Sánchez, H.H.; López-Hernández, M.; Luzariaga-Neira, A.; Rodríguez-González, H.; Aguirre, A.A.; Reyes-López, M.A. Molecular Identification and Novel Mitochondrial COI Gene Haplotypes of Nesting Kemp’s Ridley Turtles (Lepidochelys kempii) in Rancho Nuevo Sanctuary, Mexico. Diversity 2022, 14, 390. https://0-doi-org.brum.beds.ac.uk/10.3390/d14050390
Camacho-Sánchez FY, Narváez-Zapata JA, Acosta-Sánchez HH, López-Hernández M, Luzariaga-Neira A, Rodríguez-González H, Aguirre AA, Reyes-López MA. Molecular Identification and Novel Mitochondrial COI Gene Haplotypes of Nesting Kemp’s Ridley Turtles (Lepidochelys kempii) in Rancho Nuevo Sanctuary, Mexico. Diversity. 2022; 14(5):390. https://0-doi-org.brum.beds.ac.uk/10.3390/d14050390
Chicago/Turabian StyleCamacho-Sánchez, Fátima Yedith, José Alberto Narváez-Zapata, Héctor Hugo Acosta-Sánchez, Martha López-Hernández, Agusto Luzariaga-Neira, Hervey Rodríguez-González, A. Alonso Aguirre, and Miguel Angel Reyes-López. 2022. "Molecular Identification and Novel Mitochondrial COI Gene Haplotypes of Nesting Kemp’s Ridley Turtles (Lepidochelys kempii) in Rancho Nuevo Sanctuary, Mexico" Diversity 14, no. 5: 390. https://0-doi-org.brum.beds.ac.uk/10.3390/d14050390