
Species Diversity and Community Composition of Macroinvertebrates in Headwater Streams of Two Subtropical Neighboring Lowland Basins


Abstract
:1. Introduction
2. Materials and Methods
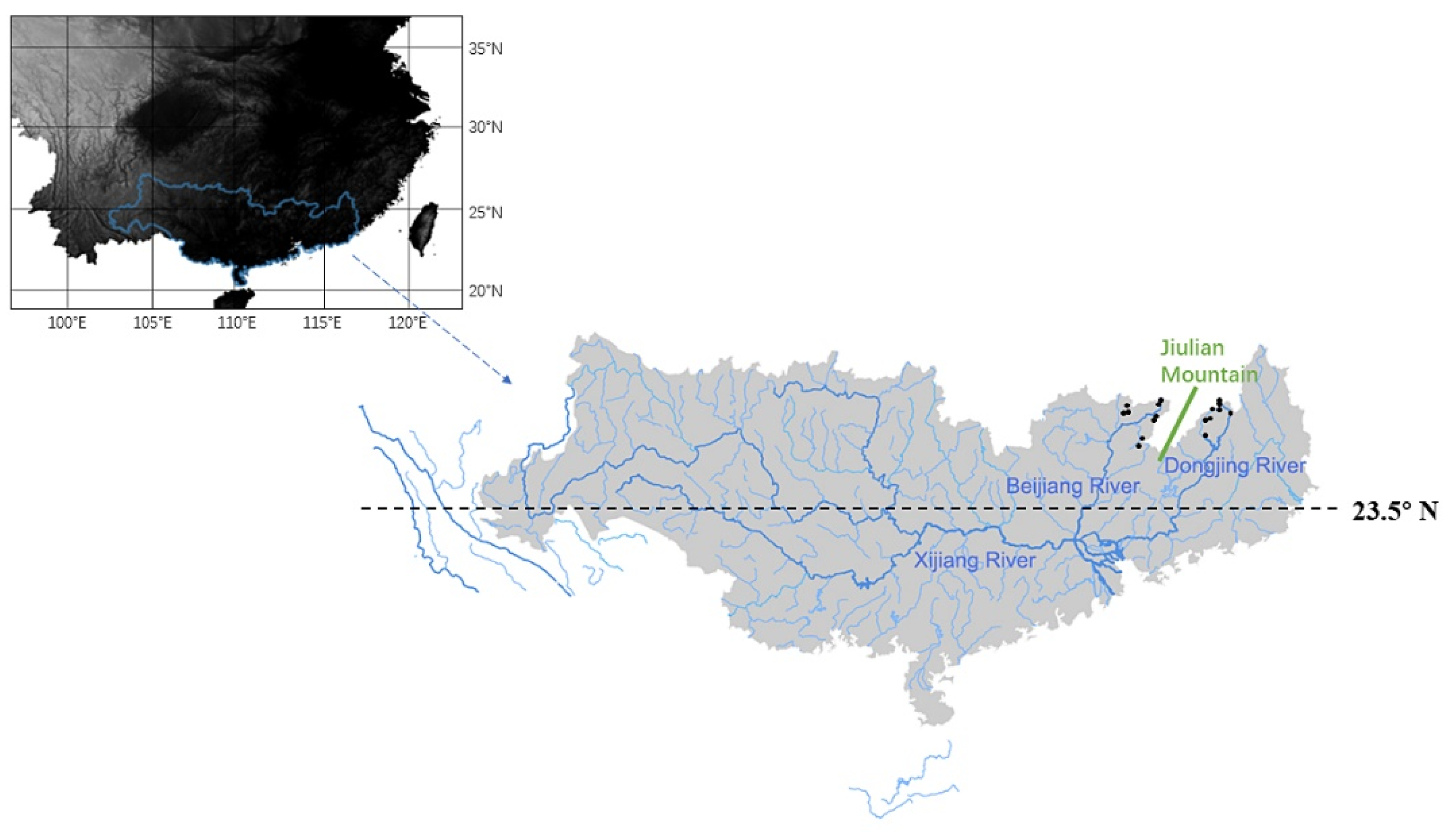
2.1. Study Area
2.2. Sampling and Identification of Macroinvertebrates
2.3. Measurements of Environmental Variables
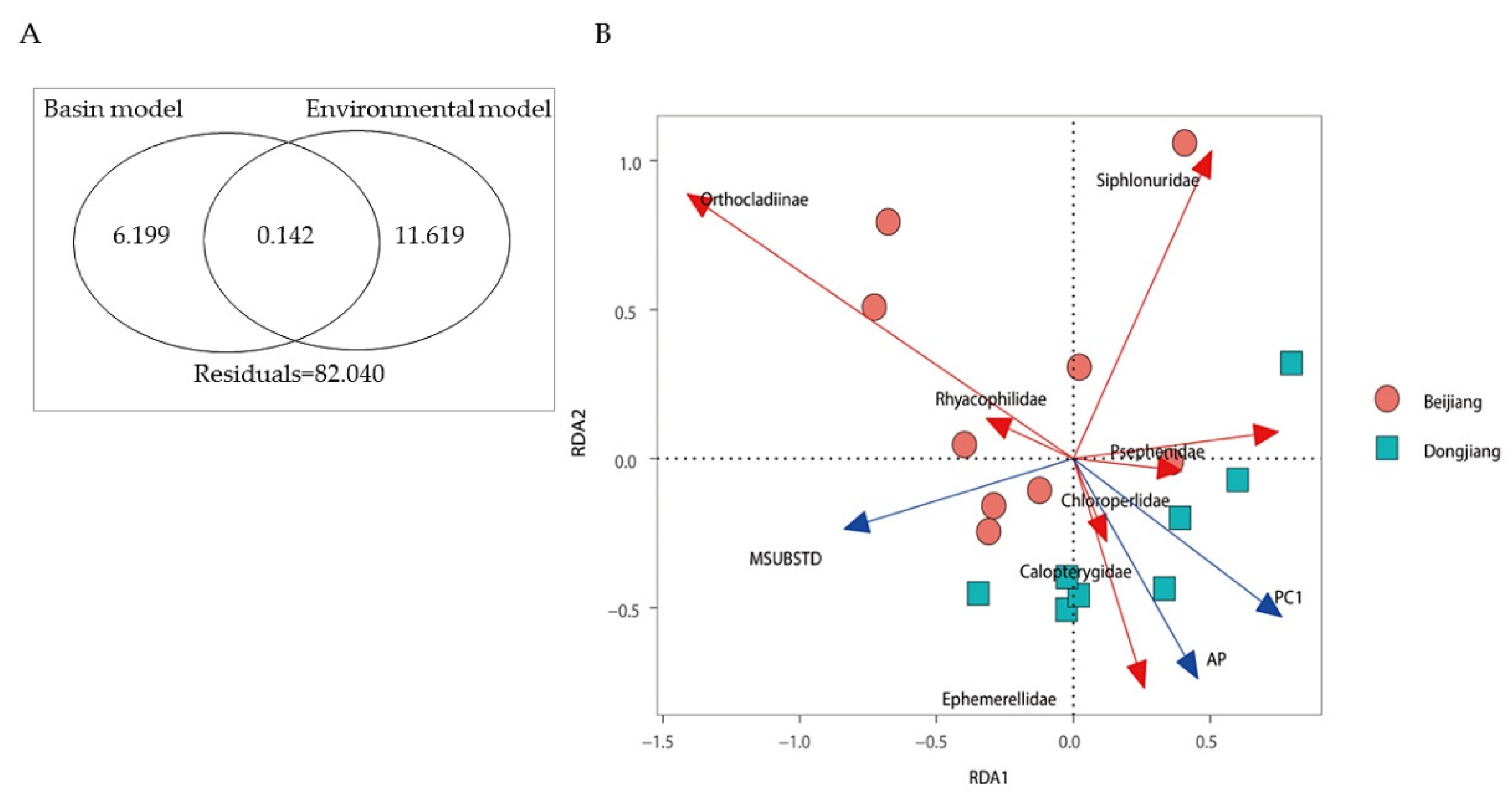
2.4. Data Analyses
3. Results
3.1. Environment Factors
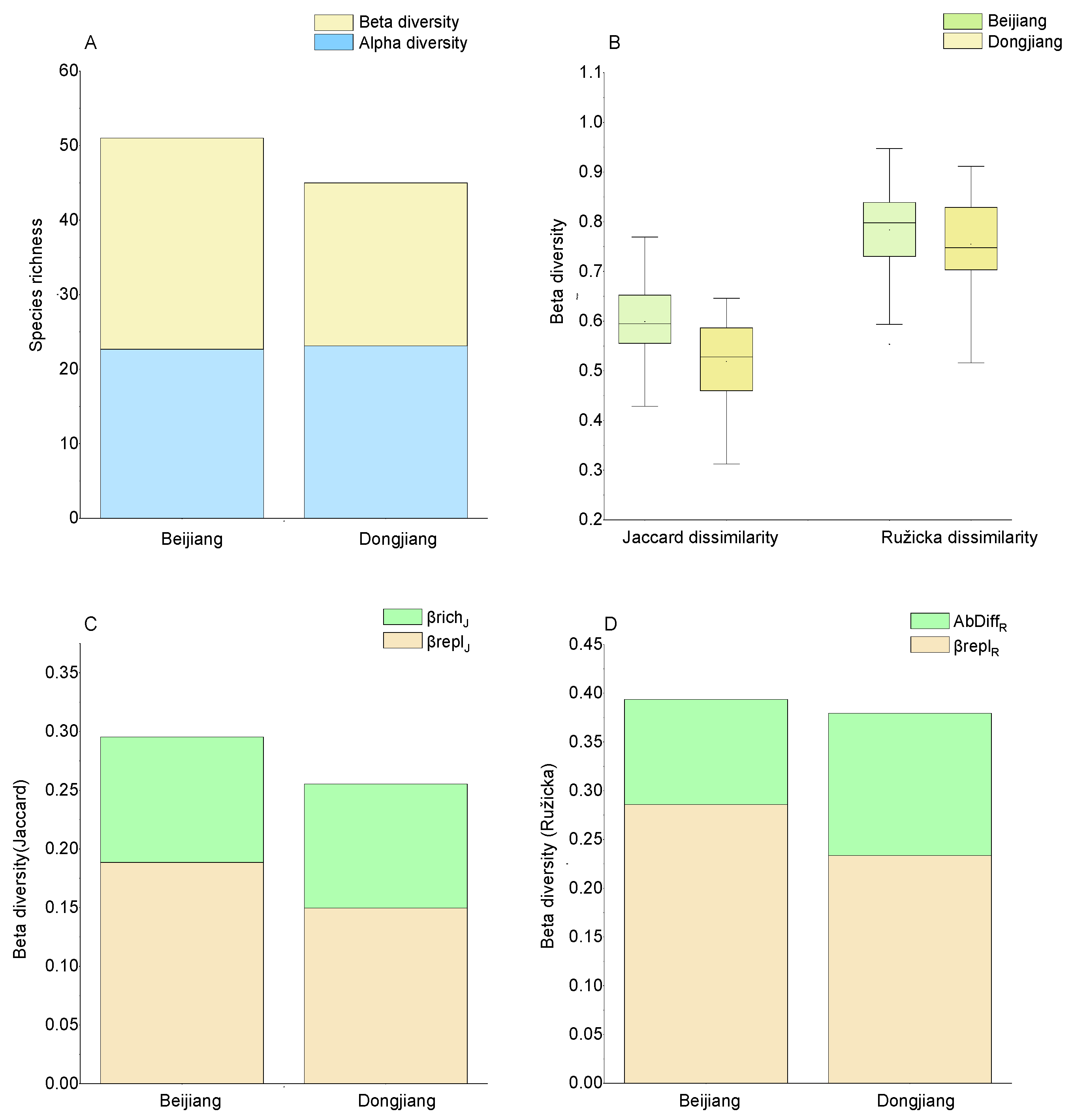
3.2. Taxa Composition, Alpha and Gamma Diversity
3.3. Beta Diversity and Its Components
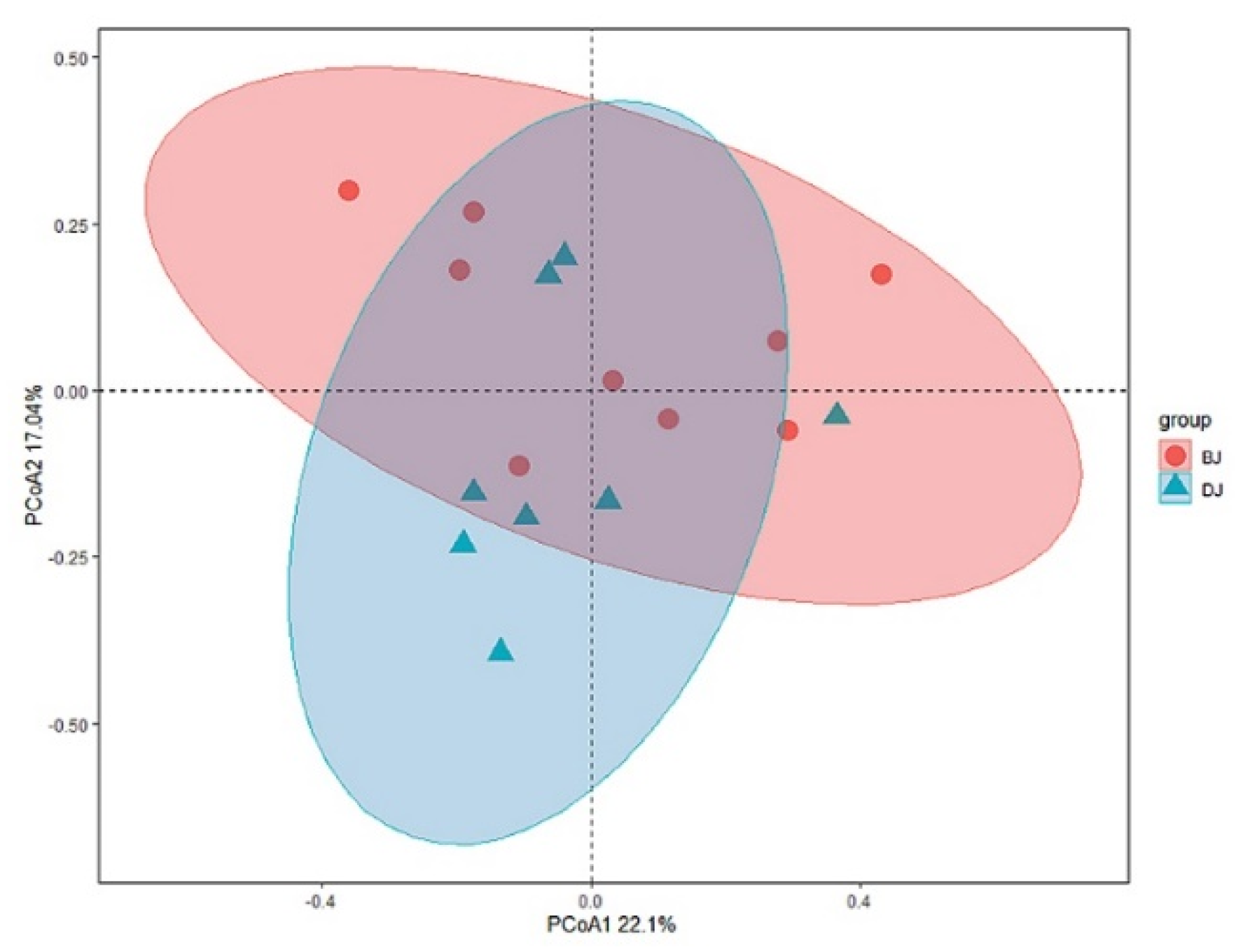
3.4. Environmental Selection in Structuring Macroinvertebrate Community
4. Discussion
4.1. Local and Regional Diversity of Macroinvertebrates
4.2. Environmental Factors Shaping Macroinvertebrate Communities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Whittaker, R.H. Vegetation of the Siskiyou mountains, Oregon and California. Ecol. Monogr. 1960, 30, 279–338. [Google Scholar] [CrossRef]
- Cottenie, K. Integrating environmental and spatial processes in ecological community dynamics. Ecol. Lett. 2005, 8, 1175–1182. [Google Scholar] [CrossRef] [PubMed]
- Sanders, N.J.; Rahbek, C. The patterns and causes of elevational diversity gradients. Ecography 2012, 35, 1–3. [Google Scholar] [CrossRef]
- Wilson, D.S. Complex interactions in metacommunities, with implications for biodiversity and higher levels of selection. Ecology 1992, 73, 1984–2000. [Google Scholar] [CrossRef]
- Leibold, M.A.; Chase, J.M.; Ernest, S.K.M. Community assembly and the functioning of ecosystems: How metacommunity processes alter ecosystems attributes. Ecology 2017, 98, 909–919. [Google Scholar] [CrossRef]
- Leibold, M.A.; Holyoak, M.; Mouquet, N.; Amarasekare, P.; Chase, J.M.; Hoopes, M.F.; Holt, R.D.; Shurin, J.B.; Law, R.; Tilman, D.; et al. The metacommunity concept: A framework for multi-scale community ecology. Ecol. Lett. 2004, 7, 601–613. [Google Scholar] [CrossRef]
- Legendre, P. Interpreting the replacement and richness difference components of beta diversity. Glob. Ecol. Biogeogr. 2014, 23, 1324–1334. [Google Scholar] [CrossRef]
- Podani, J.; Schmera, D. A new conceptual and methodological framework for exploring and explaining patterns in presence-absence data. Oikos 2011, 120, 1625–1638. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Kraft, N.J.B.; Comita, L.S.; Chase, J.M.; Sanders, N.J.; Swenson, N.G.; Crist, T.O.; Stegen, J.C.; Vellend, M.; Boyle, B.; Anserson, M.J.; et al. Disentangling the drivers of β diversity along latitudinal and elevational gradients. Science 2011, 333, 1755–1758. [Google Scholar] [CrossRef] [Green Version]
- Ricklefs, R.E. A comprehensive framework for global patterns in biodiversity. Ecol. Lett. 2004, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Harrington, R.A.; Poff, L.R.; Kondratieff, B.C. Aquatic insect β-diversity is not dependent on elevation in Southern Rocky Mountain streams. Freshw. Biol. 2016, 61, 195–205. [Google Scholar] [CrossRef]
- da Silva, P.G.; Lobo, J.M.; Hensen, M.C.; Vaz-de-Mello, F.Z.; Hernández, M.I.M. Turnover and nestedness in subtropical dung beetle assemblages along an elevational gradient. Divers. Distrib. 2018, 24, 1277–1290. [Google Scholar] [CrossRef] [Green Version]
- Finn, D.S.; Blouin, M.S.; Lytle, D.A. Population genetic structure reveals terrestrial affinities for a headwater stream insect. Freshw. Biol. 2007, 52, 1881–1897. [Google Scholar] [CrossRef]
- Tonkin, J.D.; Altermatt, F.; Finn, D.S.; Heino, J.; Olden, J.D.; Pauls, S.U.; Lytle, D.A. The role of dispersal in river network metacommunities: Patterns, processes, and pathways. Freshw. Biol. 2018, 63, 141–163. [Google Scholar] [CrossRef] [Green Version]
- Boulton, A.J. Parallels and contrasts in the effects of drought on stream macroinvertebrate assemblages. Freshw. Biol. 2003, 48, 1173–1185. [Google Scholar] [CrossRef] [Green Version]
- Clarke, A.; Mac Nally, R.; Bond, N.; Lake, P.S. Macroinvertebrate diversity in headwater streams: A review. Freshw. Biol. 2008, 53, 1707–1721. [Google Scholar] [CrossRef]
- Strayer, D.L. Challenges for freshwater invertebrate conservation. J. N. Am. Benthol. Soc. 2006, 25, 271–287. [Google Scholar] [CrossRef]
- Pearson, R.G.; Christidis, F.; Connolly, N.M.; Nolen, J.A.; St Clair, R.M.; Cairns, A.; Davis, L. Stream macroinvertebrate assemblage uniformity and drivers in a tropical bioregion. Freshw. Biol. 2017, 62, 544–558. [Google Scholar] [CrossRef]
- Paller, M.H.; Blas, S.A.; Kelley, R.W. Macroinvertebrate taxonomic richness in minimally disturbed streams on the Southeastern USA Coastal Plain. Diversity 2020, 12, 459. [Google Scholar] [CrossRef]
- Brysiewicz, A.; Czerniejewski, P.; Dąbrowski, J.; Formicki, K. Characterisation of benthic macroinvertebrate communities in small watercourses of the european central plains ecoregion and the effect of different environmental factors. Animals 2022, 12, 606. [Google Scholar] [CrossRef]
- Heino, J.; Tolonen, K.T. Ecological drivers of multiple facets of beta diversity in a lentic macroinvertebrate metacommunity. Limnol. Oceanogr. 2017, 62, 2431–2444. [Google Scholar] [CrossRef]
- Declerck, S.A.J.; Coronel, J.S.; Legendre, P.; Brendonck, L. Scale dependency of processes structuring metacommunities of cladoc-erans in temporary pools of High-Andes wetlands. Ecography 2011, 34, 296–305. [Google Scholar] [CrossRef] [Green Version]
- García-Girón, J.; Wilkes, M.; Fernández-Aláez, M.; Fernández-Aláez, C. Processes structuring macrophyte metacommunities in Mediterranean ponds: Combining novel methods to disentangle the role of dispersal limitation, species sorting and spatial scales. J. Biogeogr. 2019, 46, 646–656. [Google Scholar] [CrossRef]
- Heino, J.; Melo, A.S.; Siqueira, T.; Soininen, J.; Valanko, S.; Bini, L.M. Metacommunity organisation, spatial extent and dispersal in aquatic systems: Patterns, processes and prospects. Freshw. Biol. 2015, 60, 845–869. [Google Scholar] [CrossRef]
- Heino, J.; Melo, A.S.; Jyrkänkallio-Mikkola, J.; Petsch, K.D.; Saito, S.V.; Tolonen, K.T.; Bini, L.M.; Landeiro, V.L.; Silva, T.S.F.; Pajunen, V.; et al. Subtropical streams harbour higher genus richness and lower abun-dance of insects compared to boreal streams, but scale matters. J. Biogeogr. 2018, 45, 1983–1993. [Google Scholar] [CrossRef]
- Willig, M.R.; Kaufman, D.M.; Stevens, R.D. Latitudinal gradients of biodiversity: Pattern, process, scale, and synthesis. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 273–309. [Google Scholar] [CrossRef]
- Chen, W.; Ren, K.; Isabwe, A.; Chen, H.; Liu, M.; Yang, J. Stochastic processes shape microeukaryotic community assembly in a subtropical river across wet and dry seasons. Microbiome 2009, 7, 138. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Zhou, T.; Cui, Y.; Li, Z.; Wang, W.; Chen, Y.; Xie, Z. Environmental filtering and spatial processes equally contributed to macroinvertebrate metacommunity dynamics in the highly urbanized river networks in Shenzhen, South China. Ecol. Processes 2021, 10, 1–12. [Google Scholar] [CrossRef]
- Al-Shami, S.; Heino, J.; Che Salmah, M.R.; Abu Hassan, A.; Suhaila, A.H.; Madrus, M.R. Drivers of beta diversity of macroinvertebrate communities in tropical forest streams. Freshw. Biol. 2013, 58, 1126–1137. [Google Scholar] [CrossRef]
- Li, Z.; Wang, J.; Meng, X.; Heino, J.; Sun, M.; Jiang, X.; Xie, Z. Disentangling the effects of dispersal mode on the assembly of macroinvertebrate assem-blages in a heterogeneous highland region. Freshw. Sci. 2019, 38, 170–182. [Google Scholar] [CrossRef]
- Morse, J.C.; Yang, L.; Tian, L. Aquatic Insects of China Useful for Monitoring Water Quality; Hohai University Press: Nanjing, China, 1994. [Google Scholar]
- Thorp, J.P.; Covich, M.P. Ecology and Classification of North American Freshwater Invertebrates, 2nd ed.; Academic Press: Cambridge, MA, USA, 2001. [Google Scholar]
- Wiggins, G.B. Larvae of the North American Caddisfly Genera (Trichoptera), 2nd ed.; University of Toronto Press: Toronto, ON, Canada, 1996. [Google Scholar]
- Cummins, K.W. Structure and function of stream ecosystems. BioScience 1974, 24, 631–641. [Google Scholar] [CrossRef]
- Tomanova, S.; Goitia, E.; Helešic, J. Trophic levels and functional feeding groups of macroinvertebrates in neotropical streams. Hydrobiologia 2006, 556, 251–264. [Google Scholar] [CrossRef]
- EPBC. Standard Methods for the Monitoring and Analysis on Water and Wastewater, 4th ed.; Chinese Environmental Science Press: Beijing, China, 2002. [Google Scholar]
- Cummins, K.W. An evaluation of some techniques for the collection and analysis of benthic samples with special emphasis on lotic waters. Am. Midl. Nat. 1962, 67, 477–504. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. A J. R. Meteorol. Soc. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Fu, L.; Jiang, Y.; Ding, J.; Liu, Q.; Peng, Q.-Z.; Kang, M.-Y. Impacts of land use and environmental factors on macroinvertebrate functional feeding groups in the Dongjiang River basin, southeast China. J. Freshw. Ecol. 2016, 31, 21–35. [Google Scholar] [CrossRef] [Green Version]
- Lande, R. Statistics and partitioning of species diversity, and similarity among multiple communities. Oikos 1996, 76, 5–13. [Google Scholar] [CrossRef]
- Crist, T.O.; Veech, J.A.; Gering, J.C.; Summerville, K.S. Partitioning species diversity across landscapes and regions: A hierarchical analysis of α, β, and γ diversity. Am. Nat. 2003, 162, 734–743. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, J.C.; Cardoso, P.; Gomes, P. Determining the relative roles of species replacement and species richness differences in generating beta-diversity patterns. Glob. Ecol. Biogeogr. 2012, 21, 760–771. [Google Scholar] [CrossRef]
- Podani, J.; Ricotta, C.; Schmera, D. A general framework for analyzing beta diversity, nestedness and related community-level phenomena based on abundance data. Ecol. Complex. 2013, 15, 52–61. [Google Scholar] [CrossRef] [Green Version]
- Lai, J.; Zou, Y.; Zhang, J.; Peres-Neto, P.R. Generalizing hierarchical and variation partitioning in multiple regression and canonical analyses using the rdacca.hp R package. Methods Ecol. Evol. 2022, 13, 782–788. [Google Scholar] [CrossRef]
- Jacobsen, D.; Schultz, R.; Encalada, A. Structure and diversity of stream invertebrate assemblages: The influence of tem-perature with altitude and latitude. Freshw. Biol. 1997, 38, 247–261. [Google Scholar] [CrossRef] [Green Version]
- Ligeiro, R.; Melo, A.S.; Callisto, M. Spatial scale and the diversity of macroinvertebrates in a Neotropical catchment. Freshw. Biol. 2010, 55, 424–435. [Google Scholar] [CrossRef]
- Pearson, R.G.; Boyero, L. Gradients in regional diversity of freshwater taxa. J. N. Am. Benthol. Soc. 2009, 28, 504–514. [Google Scholar] [CrossRef]
- Balian, E.V.; Segers, H.; Lévèque, C.; Martens, K. The freshwater animal diversity assessment: An overview of the results. Hy-Drobiologia 2008, 595, 627–637. [Google Scholar] [CrossRef]
- Cortés-Guzmán, D.; Alcocer, J.; Cummins, K.W. Benthic macroinvertebrates of tropical streams: Functional and trophic diversity of the Lacantún River, Mexico. Limnology 2021, 22, 313–328. [Google Scholar] [CrossRef]
- Ilmonen, J.; Paasivirta, L.; Virtanen, R.; Muotka, T. Regional and local drivers of macroinvertebrate assemblages in boreal springs. J. Biogeogr. 2009, 36, 822–834. [Google Scholar] [CrossRef]
- Culp, J.M.; Lento, J.; Curry, R.A.; Luiker, E.; Halliwell, D. Arctic biodiversity of stream macroinvertebrates declines in response to latitudinal change in the abiotic template. Freshw. Sci. 2019, 38, 465–479. [Google Scholar] [CrossRef] [Green Version]
- Irons, J.G.; Oswood, M.W.; Stout, R.J.; Pringle, C.M. Latitudinal patterns in leaf litter breakdown: Is temperature really important? Freshw. Biol. 1994, 32, 401–411. [Google Scholar] [CrossRef]
- Yule, C.M. Trophic relationships and food webs of the benthic invertebrate fauna of two seasonal tropical streams on Bougainville Island, Papua New Guinea. J. Trop. Ecol. 1996, 12, 517–534. [Google Scholar] [CrossRef]
- Dobson, M.; Mathooko, J.M.; Magana, A.; Ndegwa, F.K. Macroinvertebrate assemblages and detritus processing in Kenyan high-land streams: More evidence for the paucity of shredders in the tropics? Freshw. Biol. 2002, 47, 909–919. [Google Scholar] [CrossRef]
- Gonçalves, J.F.; Graça, M.A.F.; Callisto, M. Leaf-litter breakdown in 3 streams in temperate, Mediterranean, and tropical Cerrado climates. J. N. Am. Benthol. Soc. 2006, 24, 344–355. [Google Scholar] [CrossRef]
- Yule, C.M.; Leong, M.Y.; Liew, K.C.; Ratnarajah, L.; Schmidt, K.; Wong, H.M.; Pearson, R.G.; Boyero, L. Shredders in Malaysia: Abundance and richness are higher in cool upland tropical streams. J. N. Am. Benthol. Soc. 2009, 28, 404–415. [Google Scholar] [CrossRef]
- Cheshire, K.; Boyero, L.; Pearson, R.G. Food webs in tropical Australian streams: Shredders are not scarce. Freshw. Biol. 2005, 50, 748–769. [Google Scholar] [CrossRef]
- Camacho, R.L.; Boyero, A.; Cornejo, A.; Ibáñez, A.; Pearson, R.G. Local variation in shredder numbers can explain their oversight in tropical streams. Biotropica 2009, 4, 625–632. [Google Scholar] [CrossRef]
- Mathuriau, C.; Chauvet, E. Breakdown of leaf litter in a neotropical stream. J. N. Am. Benthol. Soc. 2002, 21, 384–396. [Google Scholar] [CrossRef] [Green Version]
- Paine, R.T. Food web complexity and species diversity. Am. Nat. 1966, 100, 65–75. [Google Scholar] [CrossRef]
- Fox, L.R. Species richness in streams-an alternative mechanism. Am. Nat. 1977, 111, 1017–1021. [Google Scholar] [CrossRef]
- Danehy, R.J.; Chan, S.S.; Lester, G.T.; Langshaw, R. Periphyton and macroinvertebrate assemblage structure in headwaters bor-dered by mature, thinned, and clearcut Douglas-Fir stands. For. Sci. 2007, 53, 294–307. [Google Scholar]
- Arellano, G.; Cayola, L.; Loza, M.I.; Torrez, V. Commonness patterns and the size of the species pool along a tropical elevational gradient: Insights using a new quantitative tool. Ecography 2014, 37, 536–543. [Google Scholar] [CrossRef]
- Farooq, M.; Li, X.; Li, Z.; Yang, R.; Tian, Z.; Tan, L.; Fornacca, D.; Li, Y.; Cili, N.; Ciren, Z.; et al. The joint contributions of environmental filtering and spatial processes to macroinvertebrate metacommunity dynamics in the alpine stream environment of Baima Snow Mountain, Southwest China. Diversity 2022, 14, 28. [Google Scholar] [CrossRef]
- Li, Z.; Heino, J.; Chen, X.; Liu, Z.; Meng, X.; Jiang, X.; Ge, Y.; Chen, J.; Xie, Z. Understanding macroinvertebrate metacommunity organization using a nested study design across a mountainous river network. Ecol. Indic. 2021, 121, 107188. [Google Scholar] [CrossRef]
- Viana, D.S.; Figuerola, J.; Schwenk, K.; Manca, M. Assembly mechanisms determining high species turnover in aquatic com-munities over regional and continental scales. Ecography 2016, 39, 281–288. [Google Scholar] [CrossRef]
- Beisel, J.N.; Usseglio-Polatera, P.; Thomas, S.; Moreteau, J.-C. Stream community structure in relation to spatial variation: The influence of mesohabitat characteristics. Hydrobiologia 1998, 389, 73–88. [Google Scholar] [CrossRef]
- Jowett, I.G.; Richardson, J.; Biggs, B.; Hickey, C.W. Microhabitat preferences of benthic invertebrates and the development of generalised Deleatidium spp. habitat suitability curves, applied to four New Zealand rivers. N. Z. J. Mar. Freshw. Res. 1991, 25, 187–199. [Google Scholar] [CrossRef] [Green Version]
- Poff, N.L. Landscape filters and species traits: Towards mechanistic understanding and prediction in stream ecology. J. N. Am. Benthol. Soc. 1997, 16, 391–409. [Google Scholar] [CrossRef]
- Rocha, M.P.; Bini, L.M.; Domisch, S.; Tolonen, K.T.; Jyrkänkallio-Mikkola, J.; Soininen, J.; Hjort, J.; Heino, J. Local environment and space drive multiple facets of stream macroinvertebrate beta diversity. J. Biogeogr. 2018, 45, 2744–2754. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
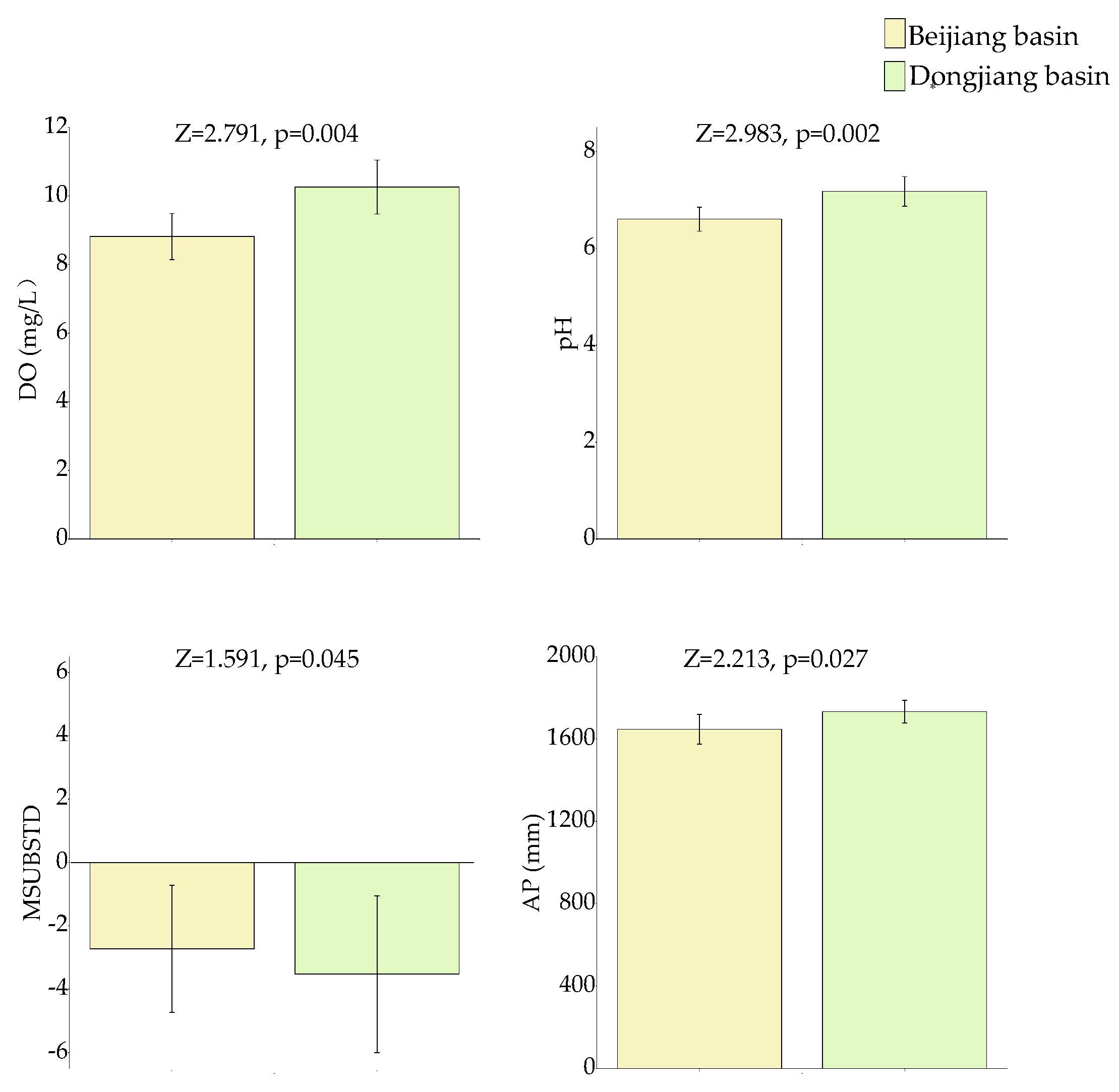{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Beijiang Basin | Dongjiang Basin | p Value | |
|---|---|---|---|
| Total macroinvertebrate | 823.951 ± 305.155 | 785.556 ± 283.806 | 0.793 |
| Gather-collector | 428.889 ± 191.804 | 397.222 ± 140.700 | 0.707 |
| Filter-collector | 149.63 ± 129.438 | 113.056 ± 92.950 | 0.519 |
| Scraper | 53.086 ± 31.140 | 83.889 ± 67.086 | 0.263 |
| Predator | 100.247 ± 59.695 | 83.333 ± 57.814 | 0.563 |
| Shredder | 92.099 ± 76.369 | 108.056 ± 69.241 | 0.660 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Lv, X.; Li, J.; Tan, L.; Rizo, E.Z.; Han, B.-P. Species Diversity and Community Composition of Macroinvertebrates in Headwater Streams of Two Subtropical Neighboring Lowland Basins. Diversity 2022, 14, 402. https://0-doi-org.brum.beds.ac.uk/10.3390/d14050402
Wang L, Lv X, Li J, Tan L, Rizo EZ, Han B-P. Species Diversity and Community Composition of Macroinvertebrates in Headwater Streams of Two Subtropical Neighboring Lowland Basins. Diversity. 2022; 14(5):402. https://0-doi-org.brum.beds.ac.uk/10.3390/d14050402
Chicago/Turabian StyleWang, Lu, Xiaochen Lv, Jiaxu Li, Lin Tan, Eric Zeus Rizo, and Bo-Ping Han. 2022. "Species Diversity and Community Composition of Macroinvertebrates in Headwater Streams of Two Subtropical Neighboring Lowland Basins" Diversity 14, no. 5: 402. https://0-doi-org.brum.beds.ac.uk/10.3390/d14050402