
A Review of Coastal Anthropogenic Impacts on Mytilid Mussel Beds: Effects on Mussels and Their Associated Assemblages

 ,
,  , and
, and
Abstract
:1. Introduction
2. Literature Selection
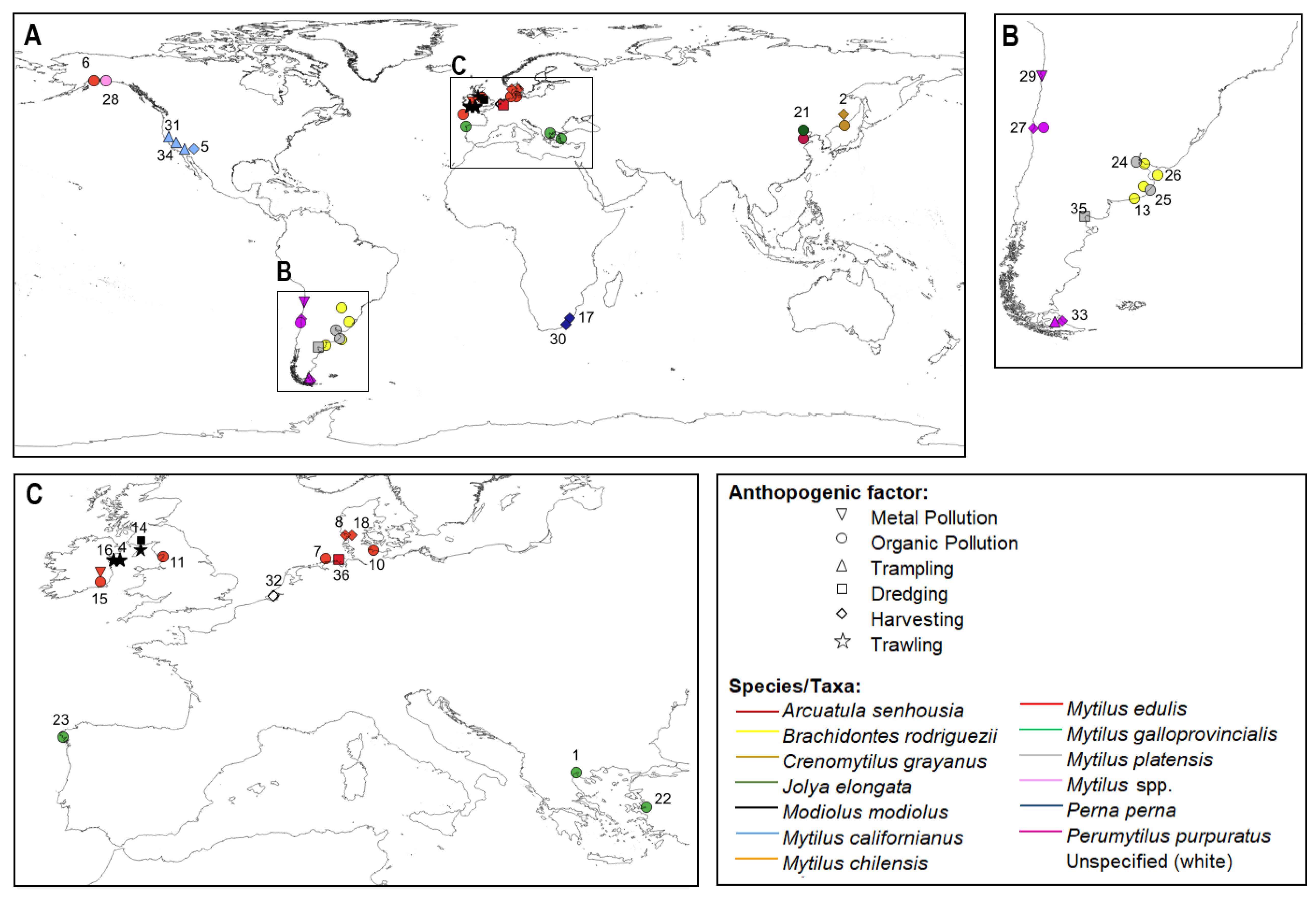
3. Geographic Area and Taxa
4. Anthropogenic Perturbations
4.1. Pollution Impacts
4.1.1. Organic Compounds
4.1.2. Heavy Metals
4.2. Small- to Medium-Scale Physical Impacts
4.2.1. Harvesting
4.2.2. Trampling
4.3. Large-Scale Physical Impacts
4.3.1. Dredging
4.3.2. Trawling
5. Synthesis and Final Remarks
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Antoniadou, C.; Sarantidis, S.; Chintiroglou, C. Small-scale spatial variability of zoobenthic communities in a commercial Mediterranean port. J. Mar. Biol. Assoc. U. K. 2011, 91, 77–89. [Google Scholar] [CrossRef]
- Galysheva, Y.A. Current state and long-term changes of the Crenomytilus grayanus community in Vostok Bay, Sea of Japan. Russ. J. Ecol. 2008, 39, 272–278. [Google Scholar] [CrossRef]
- Veiga, P.; Ramos-Oliveira, C.; Sampaio, L.; Rubal, M. The role of urbanisation in affecting Mytilus galloprovincialis. PLoS ONE 2020, 15, e0232797. [Google Scholar] [CrossRef]
- Fariñas-Franco, J.M.; Allcock, A.L.; Roberts, D. Protection alone may not promote natural recovery of biogenic habitats of high biodiversity damaged by mobile fishing gears. Mar. Environ. Res. 2018, 135, 18–28. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.R.; Fong, P.; Ambrose, R.F. The Impacts of Human Visitation on Mussel Bed Communities Along the California Coast: Are Regulatory Marine Reserves Effective in Protecting These Communities? Environ. Manag. 2008, 41, 599–612. [Google Scholar] [CrossRef] [PubMed]
- Lees, D.C.; Houghton, J.P.; Driskell, W.B. Effects of shoreline treatment methods on intertidal biota in Prince William Sound. In Proceedings of the International Oil Spill Conference, Tampa, FL, USA, 29 March–1 April 1993; American Petroleum Institute: Washington, DC, USA, 1993; Volume 1993, pp. 345–354. [Google Scholar]
- Reichert, K.; Buchholz, F. Changes in the macrozoobenthos of the intertidal zone at Helgoland (German Bight, North Sea): A survey of 1984 repeated in 2002. Helgol. Mar. Res. 2006, 60, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Riesen, W.; Reise, K. Macrobenthos of the subtidal Wadden Sea: Revisited after 55 years. Helgoländer Meeresunters. 1982, 35, 409–423. [Google Scholar] [CrossRef] [Green Version]
- Gestoso, I.; Arenas, F.; Rubal, M.; Veiga, P.; Peña, M.; Olabarria, C. Shifts from native to non-indigenous mussels: Enhanced habitat complexity and its effects on faunal assemblages. Mar. Environ. Res. 2013, 90, 85–95. [Google Scholar] [CrossRef]
- Anger, K. On the influence of sewage pollution on inshore benthic communities in the South of Kiel Bay. Helgoländer Wiss. Meeresunters. 1975, 27, 408–438. [Google Scholar] [CrossRef] [Green Version]
- Crowe, T.P.; Smith, E.L.; Donkin, P.; Barnaby, D.L.; Rowland, S.J. Measurements of sublethal effects on individual organisms indicate community-level impacts of pollution. J. Appl. Ecol. 2004, 41, 114–123. [Google Scholar] [CrossRef]
- Quintino, V.; Azevedo, A.; Magalhães, L.; Sampaio, L.; Freitas, R.; Rodrigues, A.M.; Elliott, M. Indices, multispecies and synthesis descriptors in benthic assessments: Intertidal organic enrichment from oyster farming. Estuar. Coast. Shelf Sci. 2012, 110, 190–201. [Google Scholar] [CrossRef]
- Elías, R.; Rivero, M.S.; Palacios, J.R.; Vallarino, E.A. Sewage-induced disturbance on polychaetes inhabiting intertidal mussel beds of Brachidontes rodriguezii off Mar del Plata (SW Atlantic, Argentina). Sci. Mar. 2006, 70, 187–196. [Google Scholar] [CrossRef] [Green Version]
- Cook, R.; Fariñas-Franco, J.M.; Gell, F.R.; Holt, R.H.F.; Holt, T.; Lindenbaum, C.; Porter, J.S.; Seed, R.; Skates, L.R.; Stringell, T.B.; et al. The Substantial First Impact of Bottom Fishing on Rare Biodiversity Hotspots: A Dilemma for Evidence-Based Conservation. PLoS ONE 2013, 8, e69904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Browne, M.A.; Brooks, P.R.; Clough, R.; Fisher, A.S.; Pinto, M.M.; Crowe, T.P. Simulating regimes of chemical disturbance and testing impacts in the ecosystem using a novel programmable dosing system. Methods Ecol. Evol. 2015, 7, 609–618. [Google Scholar] [CrossRef]
- Magorrian, B.H.; Service, M. Analysis of underwater visual data to identify the impact of physical disturbance on horse mussel (Modiolus modiolus) beds. Mar. Pollut. Bull. 1998, 36, 354–359. [Google Scholar] [CrossRef]
- Cole, V.J.; McQuaid, C.D.; Nakin, M.D.V. Marine protected areas export larvae of infauna, but not of bioengineering mussels to adjacent areas. Biol. Conserv. 2011, 144, 2088–2096. [Google Scholar] [CrossRef]
- Saier, B. Subtidal and intertidal mussel beds (Mytilus edulis L.) in the Wadden Sea: Diversity differences of associated epifauna. Helgol. Mar. Res. 2002, 56, 44–50. [Google Scholar] [CrossRef] [Green Version]
- World Register of Marine Species. At VLIZ. Available online: http://www.marinespecies.org (accessed on 12 January 2021).
- Cárdenas, L.; Leclerc, J.-C.; Bruning, P.; Garrido, I.; Détrée, C.; Figueroa, A.; Astorga, M.; Navarro, J.M.; Johnson, L.E.; Carlton, J.T.; et al. First mussel settlement observed in Antarctica reveals the potential for future invasions. Sci. Rep. 2020, 10, 5552. [Google Scholar] [CrossRef]
- Zhou, H.; Zhang, Z.N.; Liu, X.S.; Tu, L.H.; Yu, Z.S. Changes in the shelf macrobenthic community over large temporal and spatial scales in the Bohai Sea, China. J. Mar. Syst. 2007, 67, 312–321. [Google Scholar] [CrossRef]
- Çinar, M.E.; Katagan, T.; Koçak, F.; Öztürk, B.; Ergen, Z.; Kocatas, A.; Önen, M.; Kirkim, F.; Bakir, K.; Kurt, G.; et al. Faunal assemblages of the mussel Mytilus galloprovincialis in and around Alsancak Harbour (Izmir Bay, eastern Mediterranean) with special emphasis on alien species. J. Mar. Syst. 2008, 71, 1–17. [Google Scholar] [CrossRef]
- García-Regueira, X.; Tato, R.; Moreira, J.; Urgorri, V. Temporal evolution of polychaete assemblages on intertidal hard substrata at two localities of the Galician coast after the ‘Prestige’ oil spill. Thalassas 2010, 26, 33–45. [Google Scholar]
- Adami, M.L.; Tablado, A.; Gappa, J.L. Spatial and temporal variability in intertidal assemblages dominated by the mussel Brachidontes rodriguezii (d’Orbigny, 1846). Hydrobiologia 2004, 520, 49–59. [Google Scholar] [CrossRef]
- Vallarino, E.A.; Rivero, M.S.; Gravina, M.C.; Elías, R. The community-level response to sewage impact in intertidal mytilid beds of the Southwestern Atlantic, and the use of the Shannon index to assess pollution. Rev. Biol. Mar. Oceanog. 2002, 37, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, M.A.; Jaubet, M.L.; Garaffo, G.V.; Elías, R. Spatial and long-term analyses of reference and sewage-impacted sites in the SW Atlantic (38° S, 57° W) for the assessment of sensitive and tolerant polychaetes. Mar. Pollut. Bull. 2013, 74, 325–333. [Google Scholar] [CrossRef]
- Valdivia, N.; Thiel, M. Effects of point-source nutrient addition and mussel removal on epibiotic assemblages in Perumytilus purpuratus beds. J. Sea Res. 2006, 56, 271–283. [Google Scholar] [CrossRef] [Green Version]
- Houghton, J.P.; Fukuyama, A.K.; Lees, D.C.; Driskell, W.B.; Shigenaka, G.; Mearns, A.J. Impacts on Intertidal Epibiota: Exxon Valdez Spill and Subsequent Cleanup. In Proceedings of the International Oil Spill Conference, Tampa, FL, USA, 29 March–1 April 1993; American Petroleum Institute: Washington, DC, USA, 1993; Volume 1993, pp. 293–300. [Google Scholar]
- Acevedo, J.; Orellana, I.F.; Guiñez, R. Evaluación experimental de la toxicidad de cobre in situ sobre la fauna asociada a Perumytilus purpuratus (Bivalvia: Mytilidae), un ingeniero ecosistémico. Rev. Biol. Mar. Oceanog. 2010, 45, 497–505. [Google Scholar] [CrossRef]
- Lasiak, T.A.; Field, J.G. Community-level attributes of exploited and non-exploited rocky infratidal macrofaunal assemblages in Transkei. J. Exp. Mar. Biol. Ecol. 1995, 185, 33–53. [Google Scholar] [CrossRef]
- Beauchamp, K.A.; Gowing, M.M. A quantitative assessment of human trampling effects on a rocky intertidal community. Mar. Environ. Res. 1982, 7, 279–293. [Google Scholar] [CrossRef]
- Meire, P.M.; Dereu, J. Use of the Abundance/Biomass Comparison Method for Detecting Environmental Stress: Some Considerations Based on Intertidal Macrozoobenthos and Bird Communities. J. Appl. Ecol. 1990, 27, 210–223. [Google Scholar] [CrossRef]
- Calcagno, J.A.; Curelovich, J.N.; Fernandez, V.M.; Thatje, S.; Lovrich, G.A. Effects of physical disturbance on a sub-Antarctic middle intertidal bivalve assemblage. Mar. Biol. Res. 2012, 8, 937–953. [Google Scholar] [CrossRef]
- Van de Werfhorst, L.C.; Pearse, J.S. Trampling in the rocky intertidal of central California: A follow-up study. Bull. Mar. Sci. 2007, 81, 245–254. [Google Scholar]
- Morsan, E.M. Impact on biodiversity of scallop dredging in San Matías Gulf, northern Patagonia (Argentina). Hydrobiologia 2009, 619, 167–180. [Google Scholar] [CrossRef]
- Buhs, F.; Riese, K. Epibenthic fauna dredged from tidal channels in the Wadden Sea of Schleswig-Holstein: Spatial patterns and a long-term decline. Helgolander Meeresun. 1997, 51, 343–359. [Google Scholar] [CrossRef] [Green Version]
- Amoroso, R.O.; Pitcher, C.R.; Rijnsdorp, A.D.; McConnaughey, R.A.; Parma, A.M.; Suuronen, P.; Eigaard, O.R.; Bastardie, F.; Hintzen, N.T.; Althaus, F.; et al. Bottom trawl fishing footprints on the world’s continental shelves. Proc. Natl. Acad. Sci. USA 2018, 115, E10275–E10282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kristensen, L.D.; Stenberg, C.; Støttrup, J.G.; Poulsen, L.K.; Christensen, H.T.; Dolmer, P.; Røjbek, M.; Thorsen, S.W.; Holmer, M.; Deurs, M.V.; et al. Establishment of blue mussel beds to enhance fish habitats. Appl. Ecol. Environ. Res. 2015, 13, 783–798. [Google Scholar]


{kind=link}
| Article | Mussel Species | Habitat | Disturbance | Sampling Period | Metazoan Taxa | Mussel Response | Impact on Fauna | Indices and Statistics | Other Measures | Location |
|---|---|---|---|---|---|---|---|---|---|---|
| [1] | Mytilus galloprovincialis | Intertidal, Subtidal: Rock, Sand | Organic pollution (hydrocarbons and sewage) | 2004 | 88 spp. [only mussel beds]: Arthropoda (20.5%); Echinodermata (1.1%), Mollusca (18.2%), Annelida (36.4%), Other (23.8%) | (+) | Moderate impact | N, S, H, J, ANOVA, ANOSIM, nMDS | - | Thermaikos Gulf, Northern Aegean Sea, Greece |
| [2] | Crenomytilus grayanus | Subtidal: Rock, Sand | Organic pollution (sewage), Harvesting | 2000–2004 | 138 spp.: Arthropoda (11.6%), Echinodermata (11.6%), Mollusca (23.2%), Annelida (32.6%), Other (21.0%) | (−) | High impact | N, S, H, J | Biomass, trophic guilds | Vostok Bay, Sea of Japan, Russia |
| [4] | Modiolus modiolus | Subtidal: Mud, Rock, Shells | Trawling | 2010 | 273 spp.: Annelida (34.1%), Arthropoda (17.9%), Echinodermata (6.2%), Mollusca (17.2%), Other (24.6%) | (−) | High impact | N, S, H, J, Margalef, PERMANOVA, PERMDISP, SIMPER | - | Strangford Lough, Ireland |
| [5] | Mytilus californianus | Intertidal: Rock | Harvesting, Trampling | not specified | 22 spp. [highest species richness in a site]: not specified | (−) | Low impact | N, S, H, J, t-test | Biomass, cover, size, thickness | CA, USA |
| [6] | Mytilus edulis | Intertidal: Rock | Organic pollution (hydrocarbons) | 1989 | 18 spp. [highest species richness in a site]: not specified | (−) | High impact | N, S | Cover | Prince William Sound, AK, USA |
| [7] | Mytilus edulis | Intertidal: Rock | Unspecified pollutants Alien species | 2002 | 154 spp.: Annelida (18.2%), Arthropoda (27.9%), Echinodermata (1.9%), Mollusca (18.2%), Other (33.8%) | - | Moderate impact | S, Conspicuousness, Cluster Analysis, nMDS | - | Helgoland, German Bight, North Sea |
| [8] | Mytilus edulis | Subtidal: Gravel, Mud, Rock, Sand, Shell | Human interference: Harvesting (Ostrea edulis), | 1980 | 89 spp.: not specified | (+) | High impact | N, S, Cluster Analysis, Linear Regression | - | Island of Sylt, German Bight, North Sea |
| [10] | Mytilus edulis | Subtidal: Mud, Sand | Organic pollution (sewage) | 1971, 1972 | 38 spp. [only mussel beds]: Annelida (44.7%), Arthropoda (28.9%); Mollusca (21.1%), Echinodermata (2.6%), Other (2.6%) | (-) | Moderate impact | N, S, H, J, Simpson | Biomass | Kiel Bay, Baltic Sea |
| [11] | Mytilus edulis | Intertidal: Rock, Sand, Shells | Organic pollution (hydrocarbons) | 1999 | 57 spp.: Annelida (22.8%), Arthropoda (35.1%), Mollusca (31.6%), Other (10.5%) | (−) | Moderate impact | S, H, BIOENV, MANOVA, nMDS, SIMPER | - | West Coast of UK |
| [13] | Brachidontes rodriguezii | Intertidal: Rock, Sand | Organic pollution (sewage) | 1997–2000 | 12 spp.: only Polychaeta | (−) | Moderate impact | ANOSIM, SIMPER | - | Mar del Plata, Argentina |
| [14] | Modiolus modiolus | Subtidal: Rock | Dredging, Trawling | 2007–2009, 2012 | 29 spp.: not specified | (−) | High impact | N, S, H, J, Margalef, ANOVA, nMDS, PERMANOVA, PERMDISP, SIMPER | - | Isle of Man and Wales, UK |
| [15] | Mytilus edulis | Subtidal: Artificial structures | Metal and Organic pollution (experimental) | 2010 | not specified | (−) | High impact | ANOVA | - | Malahide Marina, Ireland |
| [16] | Modiolus modiolus | Subtidal: Mud, Rock, Shells | Trawling | not specified | not specified | (−) | High impact | Cluster Analysis, DECORANA | - | Strangford Lough, Ireland |
| [17] | Perna perna | Intertidal: Rock | Harvesting | 2008 | not specified | (−) | Moderate impact | N, S, PERMANOVA | Cover | Transkei, South Africa |
| [18] | Mytilus edulis | Intertidal, Subtidal: Rock | Harvesting (subtidal) | 1997, 1998 | 19 spp. [Intertidal], 22 spp. [Subtidal]: not specified | (−) | Moderate impact | N, S, H, J, Sørensen’s index, Renkon’s index, ANOVA, Kruskal-Wallis Test, Mann-Whitney U-Test | - | Island of Sylt, German Bight, North Sea |
| [21] | Arcuatula senhousia, Jolya elongata | Subtidal: Mud, Sand | Organic pollution (unspecified), | 1985–1987, 1997–1999 | 460 spp.: not specified | (−) | Moderate impact | N, S, H, J, ANOSIM, nMDS | - | Bohai Sea, China |
| [22] | Mytilus galloprovincialis | Intertidal: Artificial structures | Organic pollution (unspecified) | 2004 | 155 spp.: Annelida (43.2%); Arthropoda (18.1%) Echinodermata (0.6%), Mollusca (9.7%), Other (28.4%) | (+) | Moderate impact | N, S, H, J, Margalef, ANOVA, ANOSIM, BIOENV, nMDS, SIMPER | Biomass | Izmir Bay, Eastern Mediterranean, Turkey |
| [23] | Mytilus galloprovincialis | Intertidal: Rock | Organic pollution (hydrocarbons) | 2004, 2005 | 104 spp.: only Annelida | - | Moderate impact | N, S, H, nMDS | - | Caldebarcos and O Segaño, Galicia, Spain |
| [24] | Brachidontes rodriguezii, Mytilus platensis | Intertidal: Rock, Sand | Organic pollution (sewage) | 1999, 2000 | 24 spp.: Annelida (33.3%), Arthropoda (29.2%), Mollusca (20,8%), Other (16.7%) | (+) | Moderate impact | S, H, Margalef, ANOVA, ANOSIM, nMDS | Biomass | Quequén and Necoche, Argentina |
| [25] | Brachidontes rodriguezii, Mytilus platensis | Intertidal: Rock, Sand | Organic pollution (sewage) | 1997 | 43 spp.: not specified | (−) | Moderate impact | N, S, H, J, Jack-Knife test | - | Mar del Plata, Argentina |
| [27] | Perumytilus purpuratus | Intertidal: Rock | Harvesting, Organic enrichment (experimental) | 2004 | 45 spp.: Annelida (17.8%), Arthropoda (31.1%), Mollusca (35.6%), Echinodermata (4.4%) Other (11.1%) | - | Moderate impact | N, S, ANOVA, ANOSIM, SIMPER | Trophic guilds | Bahía Totoralillo, Northern-Central Chile |
| [28] | Mytilus sp. | Intertidal: Rock, Heterogenous sediment | Organic pollution (hydrocarbons), | 1990, 1991 | 42 spp. [highest species richness in a site]: not specified | (−) | High impact | N, S, H, ANOVA | Cover | Prince William Sound, AK, USA |
| [29] | Perumytilus purpuratus | Intertidal: Rock | Metal pollution (experimental) | 2007 | 46 spp.: Annelida (30.4%), Arthropoda (19.6%), Echinodermata (8.7%), Mollusca (23.9), Other (17.4%) | null | Moderate impact | S, H, J | - | Bahia San Jorge, Northern Chile |
| [30] | Perna perna | Intertidal: Rock | Harvesting | not specified | not specified | (−) | Low impact | N. S, H, nMDS | Biomass, trophic guilds | Transkei, South Africa |
| [31] | Mytilus californianus | Intertidal: Rock | Trampling | 1977, 1978 | 67 spp.: not specified | null | Low impact | N, S, H, Dominance curves, Kruskall-Wallis Test, Mann-Whitney U-Test | - | Santa Cruz, CA, USA |
| [32] | unspecified | Intertidal: Mud, Sand | Harvesting | 1981–1984, 1987 | 44 spp. [unpolluted site]: Annelida (50.0%), Arthropoda (20.4%), Echinodermata (3.7%), Mollusca (25.9%) | (−) | Moderate impact | ABC K-Dominance curves | - | Netherlands and Belgium |
| [33] | Perumytilus purpuratus, Mytilus chilensis | Intertidal: Mud, Rock, Sand | Harvesting, Trampling (experimental) | 2001 | not specified | (−) | Moderate impact | ANOVA, Mann-Whitney U-Test | Biomass, cover, size | Rio Grande, Tierra del Fuego, Argentina |
| [34] | Mytilus californianus | Intertidal: Rock | Trampling | 1995 | 20 spp. [highest species richness in a site]: not specified | (−) | Low impact | S, H, ANOVA, ANCOVA, t-test | Cover | Santa Cruz, CA, USA |
| [35] | Mytilus platensis | Subtidal: Gravel, Mud, Sand, Shells | Dredging | 1986–1997 | 46 spp. [highest species richness in a site]: not specified | (−) | Moderate impact | S, ANOSIM, MDS | - | Golfo de San Matías, Argentina |
| [36] | Mytilus edulis | Subtidal: Gravel, Sand, Shells | Dredging | 1988, 1992 | 42 spp.: Arthropoda (23.8%), Echinodermata (4.8%), Mollusca (21.4%), Annelida (4.8%), Other (45.2%) | (+) | High impact | Mann-Whitney U-Test | - | Schleswig-Holstein, Wadden Sea, North Sea |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sampaio, L.; Moreira, J.; Rubal, M.; Guerrero-Meseguer, L.; Veiga, P. A Review of Coastal Anthropogenic Impacts on Mytilid Mussel Beds: Effects on Mussels and Their Associated Assemblages. Diversity 2022, 14, 409. https://0-doi-org.brum.beds.ac.uk/10.3390/d14050409
Sampaio L, Moreira J, Rubal M, Guerrero-Meseguer L, Veiga P. A Review of Coastal Anthropogenic Impacts on Mytilid Mussel Beds: Effects on Mussels and Their Associated Assemblages. Diversity. 2022; 14(5):409. https://0-doi-org.brum.beds.ac.uk/10.3390/d14050409
Chicago/Turabian StyleSampaio, Leandro, Juan Moreira, Marcos Rubal, Laura Guerrero-Meseguer, and Puri Veiga. 2022. "A Review of Coastal Anthropogenic Impacts on Mytilid Mussel Beds: Effects on Mussels and Their Associated Assemblages" Diversity 14, no. 5: 409. https://0-doi-org.brum.beds.ac.uk/10.3390/d14050409